Биология: учебник: в 2 т. / под ред. В. Н. Ярыгина. - 2011. - Т. 1. - 736 с. : ил.
|
|
|
|
Глава 7. ПЕРИОДИЗАЦИЯ ОНТОГЕНЕЗА
7.1. этапы, периоды и стадии онтогенеза
Индивидуальное развитие представляет собой целостный непрерывный процесс, в котором отдельные события увязаны между собой в пространстве и времени. Существует несколько вариантов периодизации онтогенеза, каждый из которых наилучшим образом подходит для решения конкретных научных или практических задач.
С общебиологической точки зрения важнейшее событие онтогенеза - половое размножение. Если соотнести различные временные отрезки онтогенеза со способностью особи осуществлять функцию размножения, то его можно разделить на три периода: дорепродуктивный, активный репродуктивный и пострепродуктивный.
В дорепродуктивном периоде особь не способна к размножению. Основное содержание его заключается в развитии зрелого в половом отношении фенотипа. В этом периоде происходят наиболее выраженные структурные и функциональные преобразования, реализуется основная часть наследственной информации, организм обладает высокой чувствительностью к всевозможным воздействиям.
В активном репродуктивном периоде особь осуществляет функцию полового размножения, отличается наиболее стабильным функционированием органов и систем, а также относительной устойчивостью к воздействиям.
Пострепродуктивный период связан со старением организма и характеризуется ослаблением или полным прекращением участия в размножении. Снижаются приспособительные возможности и устойчивость к разнообразным воздействиям. Применительно к онтогенезу человека названные периоды дополнительно характеризуются специфическими социальными моментами (образование, трудоспособность, творчество). Для каждого из периодов характерны свои особенности заболеваемости.
Дорепродуктивный период подразделяют еще на четыре: эмбриональный, личиночный, метаморфоз и ювенильный.
Эмбриональный, или зародышевый, период онтогенеза начинается с момента оплодотворения и продолжается до выхода зародыша из яйцевых оболочек. Этот период отличается выраженностью процессов преобразования зиготы в организм, способный к более или менее самостоятельному существованию. У большинства позвоночных он включает стадии (фазы) зиготы, дробления, гаструляции, а также гисто- и органогенеза. Продолжительность его бывает различна. У плацентарных млекопитающих он особенно укорочен. Единственная яйцевая оболочка растворяется перед имплантацией бластоцисты в слизистую оболочку матки. Зародыш к этому моменту успевает пройти только стадии зиготы и дробления. Все дальнейшие процессы протекают под защитой и при участии материнского организма. Эволюционное значение этих особенностей рассмотрено в п. 13.2.
Дроблению предшествуют процессы гаметогенеза и оплодотворения, которые относятся непосредственно к индивидуальному развитию и могут даже не привести к нему, но которые во многом определяют дальнейшее развитие зародыша в том случае, если зачатие состоится. Эти процессы называют прогенезом, предшествующим собственно онтогенезу. Цитологически процессы гаметогенеза и оплодотворения представляют собой промежуточное звено, связывающее онтогенезы родителей с онтогенезом их потомства.
Личиночный период в типичном варианте наблюдается в развитии тех позвоночных, зародыши которых выходят из яйцевых оболочек и начинают вести самостоятельный образ жизни, не достигнув дефинитивных (зрелых) черт организации. Так, он встречается у некоторых представителей низших позвоночных - миног, большинства костистых рыб и земноводных. Наиболее характерные черты личинки: эмбриональный характер ее организации, наличие временных (провизорных) органов, раннее начало функционирования ряда органов, дающее возможность самостоятельного существования. Благодаря активному питанию личинка получает возможность завершить развитие, а благодаря активному перемещению имеет возможность выбирать условия среды, оптимальные для развития, и уйти таким образом от конкуренции со своими же взрослыми сородичами. У позвоночных продолжительность личиночного периода в сравнении с эмбриональным существенно больше.
Метаморфоз состоит в превращении личинки в ювенильную форму. В процессе метаморфоза происходят такие важные морфогенетические преобразования, как частичное разрушение, перестройка и новообразование органов. Степень преобразований тем больше, чем больше
различия между средой обитания личинки и взрослого организма, что хорошо иллюстрирует пример развития бесхвостых амфибий (сравни: головастик и лягушка).
Ювенильный период начинается с момента завершения метаморфоза (у плацентарных млекопитающих и человека - с рождения) и заканчивается половым созреванием и началом размножения. Особенности ювенильного периода проявляются в своеобразии питания молодого организма, его поведения и степени зависимости от родителей. С морфологической точки зрения для этого периода характерны интенсивный рост, установление окончательных пропорций между различными частями тела, завершение развития скелета, кожных покровов, смена зубов, завершение развития половых желез и гормональной регуляции. Продолжительность ювенильного периода у позвоночных варьирует от минимальной, равной 13-18 сут у мышей-полевок, до максимальной, равной 18-20 годам у белуги, крокодила, альбатроса, слона. У многих представителей позвоночных, особенно у человека, достижение половой зрелости и начало размножения могут быть разделены значительным промежутком времени.
Применение эколого-эмбриологического подхода позволяет разделить онтогенез на этапы, протекающие до рождения, во время и после рождения особи. Само рождение, т.е. выход развивающейся особи из оболочек яйца или из организма матери, у разных видов происходит на разных стадиях зрелости. В то же время у всех видов до рождения организм находится под защитой яйцевых оболочек или материнского организма и не способен питаться и осуществлять другие важные функции самостоятельно. Защищенность ранних, морфофункционально незрелых стадий обеспечивает выживаемость вида. После рождения особь устанавливает связи с новой средой, начинает самостоятельно питаться, передвигаться и осуществлять все другие функции.
Более подробное изложение периодизации онтогенеза человека, имеющее практическое значение в акушерской и педиатрической практике, см. п. 7.6.1 и табл. 7.3.
7.2. морфофизиологические и эволюционные особенности яиц хордовых
7.2.1. ПРОИСХОЖДЕНИЕ ЯЙЦЕКЛЕТОК
Яйца хордовых состоят из яйцеклетки, называемой иногда также яйцом, и яйцевых оболочек. Яйцеклетки образуются в женской половой
железе - яичнике. Они проходят долгий путь развития, который начинается в эмбриональном и продолжается в активном репродуктивном периоде онтогенеза особей женского пола.
Первичные половые клетки очень рано в эмбриогенезе обособляются от соматических клеток. Так, у бесхвостых амфибий гоноциты обособляются на стадии бластулы, у птиц они определяются при формировании первичного гипобласта, у млекопитающих в эпибласте на стадии гаструлы, у хвостатых амфибий этот процесс осуществляется позже, на стадии гаструлы или даже нейрулы (в мезодерме). Затем эти клетки совершают перемещения, достигая, в конце концов, зачатков половых желез. Гоноциты млекопитающих сначала оказываются во внезароды-шевой области в устье желточного мешка, а затем мигрируют в закладки гонад (рис. 7.1). Механизмы миграции изложены в пункте 6.5.3.
Попав в гонады, первичные половые клетки начинают пролифери-ровать. Они делятся митозом и называются ово(оо)гониями. У большинства низших позвоночных ово(оо)гонии сохраняют способность к делению на протяжении всего репродуктивного периода, так, например, рыбы за один нерест выделяют тысячи яиц, земноводные - сотни. У высших позвоночных число яиц, которые вызревают одномоментно, редко достигает 15, обычно их бывает меньше, иногда одно, чем можно объяснить и особенности ово(оо)генеза.
У человеческих эмбрионов женского пола ово(оо)гонии размножаются наиболее интенсивно между 2-м и 5-м месяцами внутриутробно-
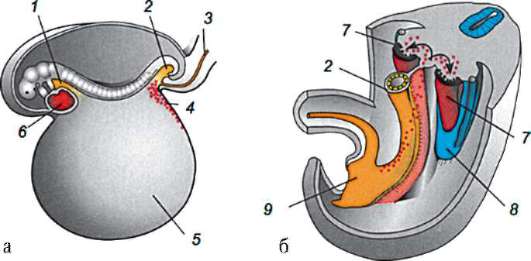
Рис. 7.1. Локализация первичных половых клеток у зародыша человека на стадии 16 сомитов (а) и их миграция в закладки гонад (б): 1 - передняя кишка; 2 - задняя кишка; 3 - аллантоис; 4 - первичные половые клетки; 5 - желточный мешок; 6 - сердце; 7 - развивающаяся гонада (половой бугорок); 8 - первичная почка; 9 - клоака
го периода развития, когда их число достигает примерно 7 млн. К 7-му месяцу многие ово(оо)гонии погибают, а сохранившиеся входят в профазу первого деления мейоза и останавливаются на стадии диакинеза. С наступлением полового созревания один овоцит ежемесячно овули-рует, достигнув стадии метафазы второго деления мейоза. Для части яйцеклеток это происходит в момент наступления полового созревания, а для других - непосредственно перед менопаузой. Овоцит завершает мейоз лишь в том случае, если происходит оплодотворение.
7.2.2. СПЕЦИФИКА И ЗНАЧЕНИЕ ХИМИЧЕСКОГО СОСТАВА ЦИТОПЛАЗМЫ ЯЙЦЕКЛЕТКИ
Зрелая яйцеклетка, как правило, бывает крупнее ово(оо)гонии и любых других соматических клеток. В ходе ово(оо)генеза в цитоплазме яйцеклетки резервируется большое количество веществ, необходимых для ее созревания и обеспечения раннего эмбриогенеза. Так, в течение периода роста объем ово(оо)цита мыши увеличивается примерно в 40 раз, а овоцита лягушки - более чем в 400. Скорость синтеза веществ в ово-цитах значительно возрастает, что обусловлено рядом особенностей. В созревающих яйцеклетках происходит амплификация (см. п. 2.4.3.4) отдельных генов (см. гл. 4), например генов рРНК. У амфибий (Xenopus laevis) в диплоидной клетке обнаруживается около 450 генов рРНК, а в ово(оо)ците - более миллиона. Кроме того, возрастает транскрипционная активность генома этих клеток. Так, скорость транскрипции в растущих ооцитах мыши в 10 раз выше, чем в соматических клетках.
Функциональная роль запасенных веществ различна.
Во-первых, это компоненты, необходимые для процессов репликации, транскрипции и трансляции, такие, как соответствующие ферменты, рибосомы, мРНК, тРНК и их предшественники.
Во-вторых, это набор специфических регуляторных веществ, которые обеспечивают координированное функционирование всех запасенных компонентов. К таким веществам относятся фактор дезинтеграции ядерной оболочки (с разрушения ядерной оболочки начинается прометафаза 1-го деления мейоза); фактор, вызывающий конденсацию хромосом; фактор, преобразующий ядро сперматозоида в пронуклеус и активирующий в нем синтез ДНК перед дроблением; цитостатический фактор CSF, ответственный за блок мейоза на стадии метафазы II (у многих позвоночных снятие этого блока происходит только в результате оплодотворения); система микрофиламентов цитоскелета, вовлеченная в поддержание блока
мейоза; циклические нуклеотиды (например, цАМФ), протеинкиназы и другие вещества, необходимые для передачи сигналов от окружающих ово(оо)цит клеток и внеклеточной среды; компоненты так называемой инозитолфосфатной системы, обеспечивающие активацию яйцеклетки после оплодотворения; фактор, ответственный за цитотомию во время дробления. Некоторые из них, будучи в овоците уже к моменту оплодотворения, начинают действовать только в фазе гаструляции.
В-третьих, это желток, в состав которого входят белки, фосфолипиды, нейтральные жиры, углеводы, минеральные соли. Желток представляет собой запас питательных веществ и энергетических ресурсов, необходимых для обеспечения эмбрионального периода. Многие вещества, вырабатываемые печенью, попадают в ово(оо)гонии в период их роста через фолликулярные клетки яичника. Это требует от самки больших энергетических затрат.
В-четвертых, это специфические вещества, называемые морфогенети-ческими детерминантами, довольно жестко определяющие судьбу клеток (бластомеров), в которые они попадают в ходе дробления. Подобные вещества обнаруживаются в цитоплазме яйцеклеток не у всех животных.
Благодаря описанным особенностям химического состава цитоплазмы яйцеклетки зародыш на протяжении периода дробления в ряде случаев не использует для развития генетическую информацию ядер зиготы и бластомеров. Специфический химический состав и закономерное распределение веществ в цитоплазме яйцеклетки имеют большое значение для начальных фаз эмбриогенеза. Запасенные питательные и энергетические вещества обеспечивают эмбриональное развитие без дополнительного поступления их извне.
7.2.3. РАЗМЕР ЯИЦ И ИХ РОЛЬ В ЭВОЛЮЦИИ. ТИПЫ ЯЙЦЕКЛЕТОК
В процессе развития выявляется закономерность, заключающаяся в том, что чем длиннее эмбриональный период, тем больше желтка должно быть накоплено в яйцеклетке. Продолжительность эмбрионального периода зависит от стадии, на которой зародыш переходит к самостоятельному существованию во внешней среде. Если постэмбриональное развитие прямое, т.е. без личинки и метаморфоза, то желтка в яйцеклетке должно быть больше. По количеству желтка яйцеклетки хордовых (табл. 7.1) делят на алецитальные, олиго-, мезо- и полилециталь-ные, т.е. с ничтожно малым, малым, средним и большим количеством желтка (от греч. лецитос - желток) (табл. 7.2).
Таблица 7.1. Систематика типа Хордовые
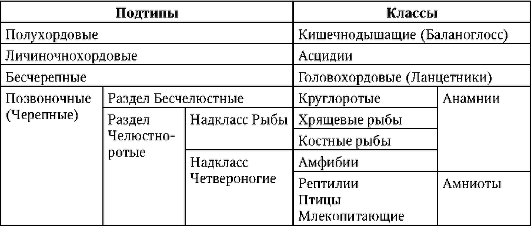
Таблица 7.2. Типы яйцеклеток, встречающиеся у хордовых

У ланцетника, представителя низших хордовых, яйцеклетка олиголецитальная. У большинства позвоночных в яйцеклетках содержится значительное количество желтка. Среди низших позвоночных (Anamnia) наиболее крупные яйца у миксин (кл. Круглоротые), у акул и химер (кл. Хрящевые рыбы) и у некоторых амфибий. У остальных амфибий, а также осетровых рыб яйцеклетки мезолецитальные, т.е. имеют среднее количество желтка. У высших позвоночных (Amniota), таких, как пресмыкающиеся, птицы и яйцекладущие млекопитающие, - полилецитальные, т.е. в яйцеклетке очень много желтка. Эмбриональное развитие у них протекает особенно долго.
Эта закономерность нарушена у сумчатых и плацентарных млекопитающих, которые имеют олиго- и алецитальные яйцеклетки соответственно. У сумчатых эмбрион выходит из яйцевых оболочек и матки при незавершенном органогенезе, переносится в сумку, где и продолжает развитие. У плацентарных, в том числе и человека, зародыш выходит
из яйцевых оболочек еще раньше, на стадии бластоцисты, но затем переходит к внутриутробному существованию, где и завершает все основные периоды развития, подготавливающие его к появлению на свет. Уменьшение количества желтка в яйцеклетках млекопитающих можно назвать вторичным, поскольку их предки, освоившие наземную среду, имели, как и все амниоты, полилецитальные яйца.
7.2.4. ПОЛЯРНОСТЬ ЯЙЦЕКЛЕТОК
Полярность яйцеклеток намечается еще на стадии накопления желтка в ово(оо)цитах во время их быстрого (большого) роста и закрепляется при выделении полярных (редукционных) телец. После выделения второго редукционного тельца полярность становится устойчивой и необратимой, что доказывается опытами Геррье по центрифугированию яйцеклеток на разных стадиях их созревания. Полюс, на котором выделяются редукционные тельца, называется анимальным, а противоположный ему - вегетативный. Полюса яйцеклетки отличаются по многим параметрам: концентрации различных веществ, количеству органоидов, активности протекания внутриклеточных процессов и ряду других. Так, эксперименты последних лет с применением вибрирующих электродов выявили электрические поля вокруг ово(оо)цитов и яйцеклеток ряда животных и растений и протекание через их цитоплазму электрических токов. Считают, что это обусловлено разной концентрацией ионных каналов и насосов на противоположных полюсах яйцеклеток. В яйце шпорцевой лягушки на анимальном полюсе выше концентрация ионных каналов, а на вегетативном - насосов.
Накопление яйцевой клеткой желтка - первое проявление ее поляризации. При малом количестве желтка в яйцеклетке он обычно распределен в цитоплазме равномерно, и ядро располагается примерно в центре. Такие яйцеклетки называют изолецитальными (от греч. изос - равный). У большинства позвоночных желтка много, и он распределен в цитоплазме яйцеклетки неравномерно. Это анизолецитальные клетки. Основная масса желтка скапливается у вегетативного полюса. Такие яйцеклетки называют телолецитальными (от греч. телос - конец). К противоположному анимальному полюсу оттесняется свободная от желтка активная цитоплазма. Если желток все же погружен в цитоплазму и не обособлен от нее в виде отдельной фракции, как у осетровых и земноводных, яйцеклетки называют умеренно телолеци-тальными. Если желток полностью отделен от цитоплазмы, как у амниот, то это резко телолецитальные яйцеклетки.
Особенности размеров и полярности яйцеклеток хордовых схематично сгруппированы в табл. 7.2.
В процессах поляризации яйцеклетки, по-видимому, принимает участие и кортикальный слой - это поверхностный слой цитоплазмы яйца, расположенный непосредственно под плазматической мембраной. В нем находятся микрофиламенты и кортикальные гранулы. Последние содержат целый спектр веществ, участвующих в формировании оболочки оплодотворения и препятствующих полиспермии. Наблюдаемая после оплодотворения сборка и перераспределение элементов цитоске-лета кортикального слоя обеспечивают приобретение им сократимости, что необходимо для осуществления делений дробления зиготы. У большинства животных первые две борозды дробления проходят по взаимно перпендикулярным анимально-вегетативным плоскостям (меридианам, соединяющим анимальный и вегетативный полюсы).
В целом, на анимальном полюсе яйцеклетки, как правило, больше свободной цитоплазмы, органоидов, запасенных РНК, обычно ядро также располагается на анимальном полюсе или ближе к нему. Вегетативный полюс характеризуется преобладанием гранул желтка.
Поляризация яйцеклетки сопровождается возникновением ово(оо)-плазматической сегрегации яйца, т.е. созданием внутренней разно-качественности участков цитоплазмы яйцеклетки.
Анимально-вегетативная поляризация яйца имеет решающее значение для всех последующих процессов эмбриогенеза, так как определяет будущую пространственную организацию зародыша. У взрослых животных переднезадняя ось тела совпадает с анимально-вегетативной осью яйцеклетки (например, у позвоночных) или перпендикулярна ей (например, у малощетинковых червей и некоторых членистоногих).
7.2.5. ЯЙЦЕВЫЕ ОБОЛОЧКИ
Яйцеклетки снаружи покрыты одной или несколькими оболочками, которые в дальнейшем выполняют в том числе и функцию защиты развивающегося зародыша.
Различают первичную оболочку, образуемую самой яйцевой клеткой, вторичную оболочку - продукт деятельности фолликулярных клеток яичников, и третичные оболочки, которыми яйцо окружается во время прохождения по яйцеводу.
Первичная оболочка, иногда называемая желточной, имеется у яйцеклеток всех животных. У позвоночных, в том числе млекопита-
ющих, первичная оболочка входит в состав плотной оболочки, образуя ее внутреннюю часть. Внешнюю часть плотной оболочки продуцируют фолликулярные клетки - это вторичная оболочка. Плотная оболочка изнутри пронизана микроворсинками яйцеклетки, а снаружи - микроворсинками фолликулярных клеток. За свои оптические свойства у млекопитающих она получила название блестящей оболочки (zona pellucida). Таким образом, эта оболочка совмещает в себе первичную и вторичную. Поверх блестящей оболочки яйцеклетки находится лучистый венец (corona radiata), образованный из фолликулярных клеток, которые прилипают к яйцеклетке, пока она находится в фолликуле яичника (рис. 7.2).
Блестящая оболочка представляет собой сложный комплекс внеклеточных гликопротеинов, которые у млекопитающих обозначаются ZP. У мышей он состоит из трех различных сульфатированных гликопротеинов - ZP1, ZP2, ZP3. С помощью ZP3 сперматозоиды связываются с блестящей оболочкой, а после проникновения одного спермия этот гликопротеин модифицируется, и проникновение дру-
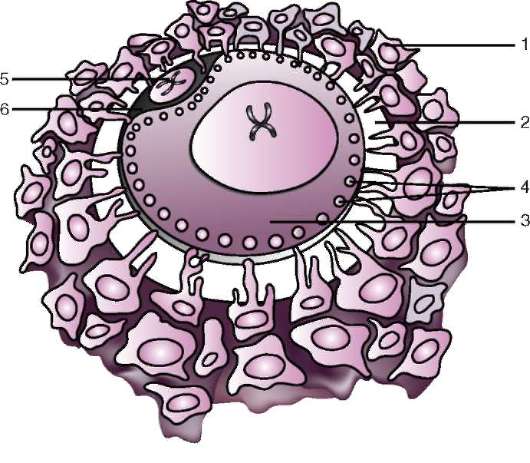
Рис. 7.2. Яйцеклетка (овоцит I порядка) млекопитающих: 1 - лучистый венец; 2 - блестящая оболочка; 3 - цитоплазма; 4 - кортикальные гранулы; 5 - полярное тельце; 6 - околожелточное пространство
гих спермиев становится невозможным. Кроме того, гликопротеины видоспецифичны, что предотвращает межвидовое оплодотворение. У других хордовых видоспецифичность оплодотворения определяется взаимодействием белков спермия с рецепторами желточной оболочки. Блестящая оболочка не примыкает непосредственно к мембране яйца, а отделяется перивителлиновым (околожелточным) пространством. Вслед за проникновением первого сперматозоида в яйцеклетку в это пространство попадает овопероксидаза кортикальных гранул. Считают, что действие этого фермента модифицирует ZP3 и ZP2, что приводит к затвердеванию блестящей оболочки. Она сохраняется вокруг зародыша на протяжении всего доимплантационного периода или в значительной части этого периода. Блестящая оболочка препятствует слипанию соседних зародышей и прилипанию зародышей к стенкам яйцевода и матки. Известно, что на начальных стадиях дробления вплоть до бластоцисты, бластомеры обладают высокой ад-гезивностью. Если трансплантировать зародыш, лишенный блестящей оболочки, в яйцевод, то неминуемо происходит адгезия бластомеров к стенке яйцевода, и зародыш погибает. Кроме того, благодаря блестящей оболочке бластомеры располагаются компактно и упорядоченно, что способствует образованию контактов и взаимодействию между ними и обеспечивает нормальное развитие зародыша на этой стадии. Если оболочку удалить, то дробление продолжится, но бластомеры будут располагаться в виде цепочки и их компактизация нарушится полностью или будет сильно запаздывать.
Третичные оболочки хорошо развиты у хрящевых рыб и амфибий, но особенную сложность они приобретают у наземных позвоночных - пресмыкающихся, птиц и низших млекопитающих. Образуясь из секретов желез яйцевода, эти оболочки не имеют клеточного строения. У всех позвоночных они выполняют функции защиты зародыша от механических повреждений и действия вредных биотических факторов, таких, как бактериальные, грибковые и протозойные. Кроме того, у наземных позвоночных появляются принципиально новые функции запасания воды и питательных веществ для обеспечения нужд зародыша. У пресмыкающихся скорлуповая оболочка действует как насос, забирая воду из почвы и воздуха. У птиц запас воды находится в белковой оболочке. Поглощение и испарение воды регулируется порами в скорлу-повой оболочке. Скорлупа содержит множество минеральных солей, необходимых для развития скелета зародыша.
7.3. ОПЛОДОТВОРЕНИЕ И ПАРТЕНОГЕНЕЗ
Оплодотворение - это процесс слияния половых клеток, завершающийся объединением их генетического материала. Образующаяся в результате оплодотворения диплоидная клетка - зигота - представляет собой начальный этап развития нового организма.
Процесс оплодотворения складывается из трех последовательных фаз:
• дистантного взаимодействия и сближения гамет;
• контактного взаимодействия гамет и активизации яйцеклетки;
• слияния гамет, или сингамии.
Сближение сперматозоида с яйцеклеткой обеспечивается совокупностью неспецифических факторов, повышающих вероятность их встречи и взаимодействия. К ним относят скоординированность наступления готовности к оплодотворению у самца и самки, поведение самцов и самок, обеспечивающее совокупление и осеменение, избыточную продукцию сперматозоидов, крупные размеры яйцеклетки, а также вырабатываемые яйцеклетками и сперматозоидами химические вещества, способствующие сближению и взаимодействию половых клеток. Эти вещества, называемые гамонами (гормоны гамет), с одной стороны, активируют движение сперматозоидов, а с другой - их склеивание. В движении сперматозоидов млекопитающих по верхним отделам яйцевода существенное значение имеет явление реотаксиса: их способности двигаться против встречного течения жидкости в маточных трубах.
У млекопитающих большое значение имеет пребывание сперматозоидов в половых путях самки, в результате чего происходит капаци-тация спермиев - приобретение ими оплодотворяющей способности. Сразу после попадания в половые пути самки спермии неспособны к проникновению в яйцеклетки. Под действием веществ секрета женских половых путей с плазмолеммы спермия в области акросомы удаляются гликопротеины и протеины семенной плазмы, которые блокируют активные центры рецепторных молекул плазмолеммы спермия, узнающие поверхность женской половой клетки. Кроме того, молекулы альбуминов, находящиеся в женском половом тракте, связываются с холестерином клеточной мембраны сперматозоидов. Это приводит к дестабилизации плазмолеммы спермия и его акросомальной мембраны, что облегчает последующее высвобождение ферментов акросом. Было обнаружено также, что в процессе капацитации изменяются свойства поверхности сперматозоидов, например ее заряд. Кроме того, происходят изменение подвижности (гиперактивация) сперматозоидов и активация акросомальных ферментов. У человека капацитация длится около 7 ч.
В фазе контактного взаимодействия спермий разрушает оболочки яйцеклетки (у млекопитающих - лучистый венец, прозрачную оболочку) и цитоплазматическую мембрану овоцита вследствие акросомной реакции. При контакте с оболочкой женской половой клетки под действием ее активирующих веществ (одно из которых - фертилизин), инициируется активное поступление катионов кальция в головку спермия. В результате происходят очаговые слияния клеточной и акросомальной мембран сперматозоида и их частичное разрушение. Через образовавшиеся микроотверстия выделяются ферменты спермия - гиалуронидаза, пенетраза и другие, которые разобщают контакты между клетками лучистого венца, а также между ними и овоцитом. Акросомальный фермент акрозин разрушает участок блестящей оболочки женской половой клетки, и спермий проникает в околожелточное пространство. В месте соприкосновения головки спермия с плазмолеммой овоцита происходят слияние и последующее разрушение мембран женской и мужской гамет. Через образующийся вследствие этого цитоплазматический мостик цитоплазмы обеих гамет объединяются. Затем в цитоплазму яйца переходят ядро и центриоль сперматозоида, а мембрана сперматозоида встраивается в мембрану яйцеклетки. Хвостовая часть сперматозоида либо остается снаружи, либо тоже входит в яйцо, но потом отделяется и рассасывается, не играя какой-либо роли в дальнейшем развитии.
Через участок мембраны сперматозоида в цитоплазму яйцеклетки начинают активно поступать ионы натрия, вследствие чего мембранный потенциал овоцита резко меняется, и женская половая клетка становится невосприимчивой к контактам с другими спермиями - быстрый блок полиспермии. Приток ионов натрия обусловливает высвобождение ионов кальция из внутриклеточных депо и увеличение его содержания в цитоплазме яйцеклетки, которое распространяется в виде волны от точки соприкосновения гамет. Вслед за этим начинается кортикальная реакция: мембраны кортикальных гранул сливаются с мембраной яйцеклетки и высвобождающиеся из них протеолитические ферменты попадают в околожелточное пространство. Под влиянием ферментов оболочка яйца уплотняется, утолщается, теряет рецепторные белки к сперматозоидам и превращается в оболочку оплодотворения. Кроме того, выделяемый из кортикальных гранул гликопротеид способствуют отслойке желточной оболочки от плазмолеммы яйцеклетки. В результате всего перечисленного проникновение других спермиев становится невозможным - медленный блок полиспермии. У млекопитающих
кортикальная реакция не вызывает образования оболочки оплодотворения, но суть ее та же.
В результате контакта сперматозоида с яйцеклеткой происходит ее активация. Она заключается в сложных структурных и физико-химических изменениях. Начальный этап активации - описанная выше кортикальная реакция. У таких животных, как иглокожие, костистые рыбы и земноводные, изменения цитоплазмы яйцеклетки сопровождаются видимыми морфологическими перестройками. Эти явления получили название расслоения или сегрегации плазмы. Значение ее для дальнейшего эмбрионального развития будет рассмотрено ниже. У многих видов бесхвостых амфибий проникновение сперматозоида в яйцеклетку приводит к перемещению пигментных гранул ани-мального полюса, и против места проникновения спермия появляется слабо окрашенная серповидная область, называемая «серым серпом» (рис. 7.3).
Активация яйцеклетки завершается началом синтеза белка на трансляционном уровне, поскольку мРНК, тРНК, рибосомы и энергия были запасены еще в ово(оо)генезе. Активация яйцеклетки может начаться и протекать до конца без ядра сперматозоида и без ядра яйцеклетки, что доказано опытами по энуклеации зиготы.
Яйцеклетка в момент встречи со сперматозоидом обычно находится на одной из стадий мейоза, заблокированной с помощью специфического фактора. У большинства позвоночных этот блок осуществляется
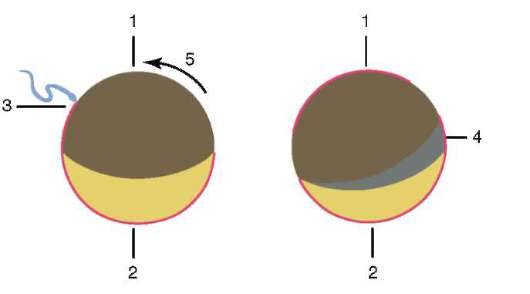
Рис. 7.3. Сегрегация цитоплазмы яйца амфибий после проникновения спермия: 1 - анимальный полюс; 2 - вегетативный полюс; 3 - место внедрения сперматозоида; 4 - серый серп; 5 - направление перемещения цитоплазмы с пигментными гранулами
на стадии метафазы II; у многих беспозвоночных, а также у трех видов млекопитающих (лошади, собаки и лисицы) блок происходит на стадии диакинеза (профаза I). В большинстве случаев блок мейоза снимается после активации яйцеклетки вследствие оплодотворения. В то время как в яйцеклетке завершается мейоз, ядро сперматозоида, проникшее в нее, видоизменяется. Оно принимает вид интерфазного, а затем профаз-ного ядра. За это время удваивается ДНК, и количество наследственного материала в мужском пронуклеусе становится п2с, т.е. он содержит гаплоидный набор редуплицированных хромосом.
Ядро яйцеклетки, закончившее мейоз, превращается в женский пронуклеус, также приобретая п2с. Оба пронуклеуса проделывают сложные перемещения, затем сближаются и сливаются (синкарион), образуя общую метафазную пластинку. Это, собственно, и есть момент окончательного слияния гамет - сингамия. Первое митотическое деление зиготы приводит к образованию двух клеток зародыша (бласто-меров) с набором хромосом 2п2с в каждом.
В некоторых случаях развитие происходит без оплодотворения - партеногенез (от греч. партеное - девственница). В случае естественного партеногенеза развитие идет на основе цитоплазмы и пронуклеуса яйцеклетки. Особи, формирующиеся из яйцеклетки, имеют либо гаплоидный, либо диплоидный набор хромосом, так как срабатывает один из механизмов удвоения числа хромосом. В одних случаях в ходе мейоза женской половой клетки выпадает стадия редукции числа хромосом, и яйцеклетка получается с диплоидным пронуклеусом. В других случаях диплоидизация происходит во время первого деления дробления, при котором не происходит цитотомии.
Естественный партеногенез, как правило, не бывает единственным способом размножения вида. Он либо чередуется с нормальным половым размножением, либо встречается у отдельных рас. Естественный партеногенез обнаружен у летних поколений некоторых ракообразных и коловраток, у пчел, ос, ряда чешуекрылых. Среди позвоночных пар-теногенетическое размножение описано у трех рас скальной ящерицы Армении, состоящих из одних самок. 40% яиц индеек, отложенных в отсутствие самца, могут начать развиваться, однако это развитие редко доходит до конца, чаще останавливается из-за возникающих аномалий. У других видов позвоночных естественное партеногенетическое размножение неизвестно.
Искусственный партеногенез возможен, по-видимому, у всех животных. Разработка методов партеногенетического развития - важ-
ная проблема в научном и прикладном отношениях. Большой вклад в эту проблему внесли отечественные исследователи А.А. Тихомиров, Б.Л. Астауров, В.А. Струнников. Обнаружено, что активация яйцеклетки сперматозоидом не является специфической. В качестве активирующих могут выступать многие физические и химические факторы. На тутовом шелкопряде было показано, что с помощью искусственного партеногенеза можно регулировать соотношение мужского и женского пола в популяции, получая большой экономический эффект.
Естественный партеногенез чаще всего случается при незавершенном оплодотворении, т.е. в тех случаях, когда имела место активация яйцеклетки, но ядро сперматозоида не участвовало в оплодотворении. В активированных яйцах используется информация только женского пронуклеуса. Такой вид партеногенеза называют гиногенезом. При искусственном партеногенезе можно удалить женский пронуклеус, и тогда развитие осуществится только за счет мужских пронуклеусов. Это ан-дрогенез. В специальных опытах на морских ежах было установлено, что потомки наследуют либо только признаки матери при гиногенезе, либо только признаки отца - при андрогенезе. Это указывает на то, что наследственные свойства особи определяются в основном ядром, а не цитоплазмой.
7.4. эмбриональное развитие
7.4.1. ДРОБЛЕНИЕ
7.4.1.1. Сущность стадии дробления
Дробление - это ряд последовательных митотических делений зиготы и образующихся бластомеров, заканчивающийся формированием многоклеточного однослойного зародыша - бластулы. Первое деление дробления начинается после объединения наследственного материала пронуклеусов и образования общей метафазной пластинки. Возникающие при дроблении клетки называют бластомерами (от греч. бласте - росток, зачаток). Особенность митотических циклов дробления - то, что с каждым делением клетки становятся все мельче и мельче, пока не достигнут обычного для соматических клеток соотношения объемов ядра и цитоплазмы. У морского ежа, например, для этого требуется 6 делений, и зародыш состоит из 64 клеток. Между очередными делениями не происходит роста клеток (период G1 отсутствует), но обязательно синтезируется ДНК. Все предшественники ДНК и необхо-
димые ферменты накоплены в процессе овогенеза. В результате митоти-ческие циклы укорочены, и деления следуют друг за другом значительно быстрее, чем в обычных соматических клетках. После нескольких циклов деления образовавшиеся бластомеры прилегают друг к другу, образуя скопление клеток, называемое морулой. Затем между клетками образуется полость - бластоцель, заполненная жидкостью. Клетки оттесняются к периферии, формируя стенку бластулы - бластодерму. Вследствие отсутствия роста бластомеров в период дробления общий размер зародыша на стадии бластулы не превышает размера зиготы.
7.4.1.2. Морфология дробления
Как правило, бластомеры располагаются в строгом порядке друг относительно друга и полярной оси яйца. Порядок, или способ дробления, зависит от количества, плотности и характера распределения желтка в яйце. По правилам Сакса-Гертвига, клеточное ядро стремится расположиться в центре свободной от желтка цитоплазмы, а веретено клеточного деления - в направлении наибольшей протяженности этой зоны.
В олиго- и мезолецитальных яйцах дробление полное, или голо-бластическое. Такой тип дробления встречается у ланцетников, миног, некоторых рыб, всех амфибий, а также у сумчатых и плацентарных млекопитающих. При полном дроблении плоскость первого деления соответствует плоскости двусторонней симметрии. Плоскость второго деления проходит перпендикулярно плоскости первого. Обе борозды первых двух делений меридианные, т.е. начинаются на анимальном полюсе и распространяются к вегетативному полюсу. Яйцевая клетка оказывается разделенной на четыре более или менее равных по размеру бластомера. Плоскость третьего деления проходит перпендикулярно первым двум в широтном направлении. У животных, имеющих изолеци-тальные яйца, все восемь образующихся бластомеров оказываются примерно равными - равномерное дробление. В мезолецитальных яйцах проявляется неравномерность дробления. На анимальном полюсе четыре более мелких бластомера - микромеры, на вегетативном - четыре более крупных - макромеры. Затем деление опять идет в меридианных плоскостях, а потом опять в широтных.
В полилецитальных яйцеклетках костистых рыб, пресмыкающихся, птиц, а также однопроходных млекопитающих дробление частичное, или меробластическое, т.е. охватывает только свободную от желтка цитоплазму. Она располагается в виде тонкого диска на анимальном полюсе, поэтому такой тип дробления называют дискоидальным.
При характеристике типа дробления учитывают также взаимное расположение и скорость деления бластомеров. Если бластомеры располагаются рядами друг над другом по радиусам, дробление называют радиальным. Оно типично для хордовых и иглокожих. В природе встречаются и другие варианты пространственного расположения бла-стомеров при дроблении, что определяет такие его типы, как спиральное у моллюсков, билатеральное у аскариды, анархичное у медузы.
Замечена зависимость между распределением желтка и степенью синхронности деления анимальных и вегетативных бластомеров. В оли-голецитальных яйцах иглокожих и головохордовых дробление почти синхронное, в мезолецитальных яйцах, например у амфибий, синхронность нарушена после третьего деления, так как вегетативные бластоме-ры из-за большого количества желтка делятся медленнее. У организмов с частичным дроблением (рептилии, птицы) деления с самого начала асинхронны и бластомеры, занимающие центральное положение, делятся быстрее.
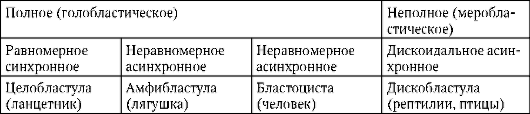
К концу дробления образуется бластула. Тип бластулы зависит от типа дробления, а значит, от типа яйцеклетки. Некоторые типы дробления и бластул представлены на рис. 7.4 и схеме (в таблице) 7.1. Более подробное описание дробления у млекопитающих, в том числе человека, см. пункт 7.5.1.
Таблица 7.3. Типы дробления и типы бластул хордовых
7.4.1.3. Особенности молекулярно-генетических и биохимических процессов при дроблении
Как было отмечено выше, митотические циклы в периоде дробления сильно укорочены, особенно в самом начале. Например, весь цикл деления в яйцах морского ежа длится 30-40 мин при продолжительности S-фазы всего 15 мин.
Gj-, а у многих животных и G2-периоды практически отсутствуют, так как в цитоплазме яйцеклетки создан необходимый запас всех веществ, и тем больший, чем она крупнее. Показано, что при удалении из зиготы
I
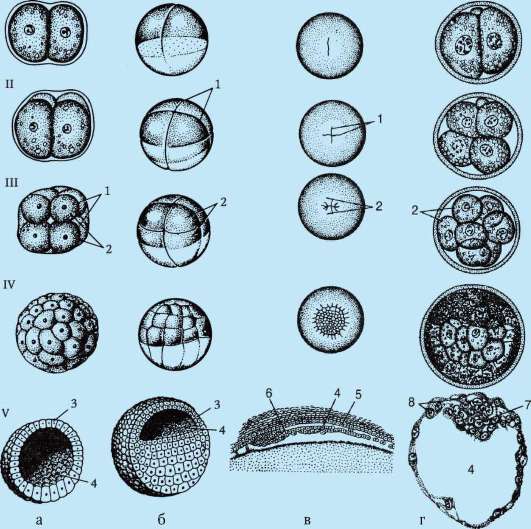
Рис. 7.4. Дробление у хордовых животных с разным типом яйцеклетки: а - ланцетник; б - лягушка; в - птица; г - млекопитающее: I - 2 бластомера; II - 4 бластомера; III - 8 бластомеров; IV - морула; V - бластула; 1 - борозды дробления; 2 - бластомеры; 3 - бластодерма; 4 - бластоцель; 5 - эпибласт; 6 - гипобласт; 7 - эмбриобласт; 8 - трофобласт; размеры зародышей на рисунке не отражают истинных размеров зародышей
ядра дробление происходит и зародыш доходит в своем развитии почти до стадии бластулы. Дальнейшее развитие прекращается.
Перед каждым делением происходит синтез ДНК и гистонов, однако длительность S-периода сокращена. При дроблении в ходе репликации ДНК скорость продвижения репликационной вилки обычная. Вместе с тем в ДНК бластомеров наблюдается больше точек инициации, чем в
соматических клетках, поэтому репликоны более короткие. Синтез ДНК идет во всех репликонах одновременно (синхронно). Поэтому время репликации ДНК в ядре совпадает со временем удвоения одного, притом укороченного, репликона.
В начале дробления другие виды ядерной активности, например транскрипция, практически отсутствуют. В разных типах яиц транскрипция генов и синтез РНК начинаются на разных стадиях. В тех случаях, когда в цитоплазме много различных веществ, как, например, у земноводных, транскрипция активируется не сразу. Синтез РНК у них начинается на стадии ранней бластулы. Напротив, у млекопитающих синтез РНК начинается уже на стадии двух бластомеров.
В периоде дробления образуются РНК и белки, аналогичные синтезируемым в процессе овогенеза. В основном это гистоны, белки клеточных мембран и ферменты, необходимые для деления клеток. Эти белки используются сразу же наравне с запасенными ранее в цитоплазме яйцеклеток. Наряду с этим в период дробления возможен синтез белков, которых не было ранее, в пользу чего свидетельствуют данные о наличии региональных различий в синтезе РНК и белков между бластомерами. Иногда эти вещества начинают действовать на более поздних стадиях.
Важную роль в дроблении играет деление цитоплазмы - цитотомия. Она имеет особое морфогенетическое значение, так как определяет тип дробления. В процессе цитотомии сначала образуется перетяжка с помощью сократимого кольца из микрофиламентов. Сборка этого кольца проходит под непосредственным влиянием полюсов митотического веретена. После цитотомии бластомеры олиголецитальных яиц остаются связанными между собой лишь тоненькими мостиками. Именно в это время их легче всего разделить. Это происходит потому, что цитотомия ведет к уменьшению зоны контакта между клетками из-за ограниченной площади поверхности мембран.
Сразу после цитотомии начинается синтез новых участков клеточной поверхности, зона контакта увеличивается, и бластомеры начинают плотно соприкасаться, что обеспечивает возможность межклеточного взаимодействия. Борозды дробления проходят по границам между отдельными участками овоплазмы, поэтому цитоплазма разных бла-стомеров различается по химическому составу. Это отражает явление овоплазматической сегрегации.
Основным результатом периода дробления становится превращение одноклеточного зародыша - зиготы - в многоклеточный и однослойный зародыш - бластулу. Помимо этого в ходе дробления происходят образо-
вание межклеточных контактов и увеличение их площади, распределение различных участков цитоплазмы зиготы между разными бластомерами зародыша, постепенная активация собственного генома зародыша, восстановление ядерно-цитоплазматического соотношения, характерного для соматических клеток. На стадии дробления все клетки зародыша однородны в отношении функционального состояния генетического аппарата. Различие цитоплазмы бластомеров и межклеточные взаимодействия детерминируют (предопределяют) направление дальнейшего развития клеток.
7.4.2. ГАСТРУЛЯЦИЯ
7.4.2.1. Сущность стадии гаструляции
Сущность стадии гаструляции заключается в том, что однослойный зародыш - бластула - превращается в многослойный - двух- или трехслойный, называемый гаструлой (от греч. гастер - желудок и уменьшительного суффикса «ул»).
У примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок - эктодерму - и внутренний зародышевый листок - энтодерму. Энтодерма формирует первичную кишку с полостью внутри - гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. Два зародышевых листка являются определяющим морфологическим признаком гаструляции. Их существование на определенной стадии развития у всех многоклеточных животных, начиная с кишечнополостных и кончая высшими позвоночными, позволяет думать о гомологии зародышевых листков и единстве происхождения всех этих животных.
У позвоночных помимо двух упомянутых зародышевых листков во время гаструляции образуется еще третий - мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка, представляющего собой хордомезодерму, - эволюционное усложнение фазы гаструляции у позвоночных, связанное с ускорением у них развития на ранних стадиях эмбриогенеза. У более примитивных хордовых животных, таких, как ланцетник, хордомезодерма обычно образуется в начале следующей после гаструляции фазы - органогенезе. Смещение времени развития одних органов относительно других у потомков по сравнению с предковыми группами является проявлением гетерохронии. Изменение времени закладки важнейших органов в процессе эволюции встречается достаточно часто.
Процесс гаструляции характеризуется важными, клеточными преобразованиями, такими, как направленные перемещения групп и отдельных клеток, избирательное размножение и сортировка клеток, начало цитодифференцировки и индукционных взаимодействий. Перечисленные клеточные механизмы онтогенеза подробно разбираются в п. 8.2.
Способы гаструляции различны. Выделяют четыре разновидности направленных в пространстве перемещений клеток, приводящих к преобразованию зародыша из однослойного в многослойный (двухили трехслойный).
Инвагинация - впячивание одного из участков бластодермы внутрь целым пластом. У ланцетника впячиваются клетки вегетативного полюса, у земноводных инвагинация происходит на границе между анимальным и вегетативным полюсами в области серого серпа. Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка.
Эпиболия - обрастание мелкими быстро делящимися клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных клеток вегетативного полюса. Такой процесс ярко выражен у земноводных.
Деламинация - расслоение клеток бластодермы на два слоя, лежащих друг над другом. Деламинацию можно наблюдать в дискобластуле зародышей с неполным дроблением таких животных, как пресмыкающиеся, птицы, яйцекладущие млекопитающие. Деламинация проявляется в эмбриобласте плацентарных млекопитающих, приводя к образованию гипобласта и эпибласта.
Иммиграция - перемещение групп или отдельных клеток, не объединенных в единый пласт. Иммиграция встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвоночных (рептилии, птицы и млекопитающие).
В каждом конкретном случае эмбриогенеза, как правило, сочетаются несколько способов гаструляции.
7.4.2.2. Морфология гаструляции
Более детальное рассмотрение гаструляции у ланцетника, лягушки, цыпленка и млекопитающих поможет глубже понять эволюционные связи и разобраться в закономерностях индивидуального развития.
7.4.2.2-а. Гаетруляция ланцетника
Гаструляция ланцетника показана на рис. 7.5. Разными маркерами на стадии бластулы (рис. 7.5, а) отмечены презумптивные (предполагаемые) зачатки. Это области бластулы, из клеточного материала ко-
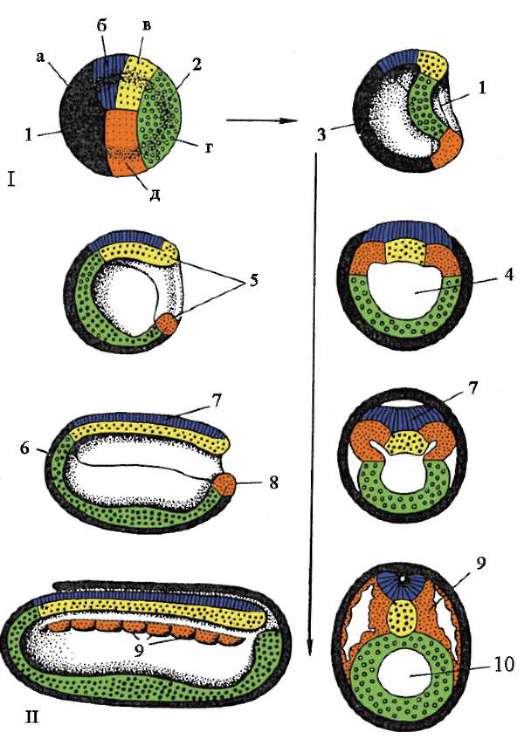
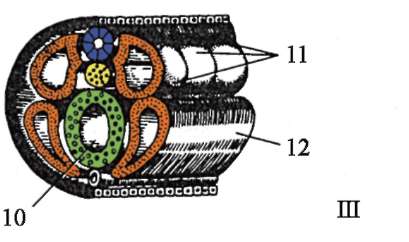
Рис. 7.5. Презумптивные зачатки, га-струляция и нейруляция у ланцетника. I - презумптивные зачатки на стадии бластулы (вид снаружи) и ранней га-струлы (вид на срезе); II - поздняя га-струла и нейруляция на сагиттальном (левый ряд) и поперечном (правый ряд) разрезах; III - пластическая модель зародыша в конце периода нейруляции:
1 - анимальный полюс; 2 - вегетативный полюс; 3 - бластоцель; 4 - гастро-цель; 5 - спинная и брюшная губы бластопора; 6 - головной конец зародыша; 7 - медуллярная пластинка; 8 - хвостовой конец зародыша; 9 - спинная часть мезодермы; 10 - полость вторичной кишки; 11 - сегментированные сомиты; 12 - брюшная часть мезодермы; а, б, в, г, д - обозначения презумптивных и развивающихся органов: а - эктодерма кожная; б - нервная трубка; в - хорда; г - энтодерма, эпителий кишки; д - мезодерма
торых в ходе гаструляции и раннего органогенеза (нейруляции) обычно образуются совершенно определенные зародышевые листки и органы (рис. 7.5, б и в).
Инвагинация начинается на вегетативном полюсе. Из-за более быстрого деления клетки анимального полюса разрастаются и толкают внутрь бластулы клетки вегетативного полюса. Этому способствует изменение состояния цитоплазмы в клетках, образующих губы бластопора и прилежащих к ним. Вследствие инвагинации бластоцель уменьшается, а гастроцель увеличивается. Одновременно с исчезновением бластоцеля эктодерма и энтодерма приходят в тесный контакт. У ланцетника, как и у всех вторичноротых животных (к ним относят тип Иглокожие, тип Хордовые и некоторые другие относительно малочисленные типы животных), область бластопора превращается в хвостовую часть организма, в отличие от первичноротых, у которых бластопор соответствует головной части. Ротовое отверстие у вторичноротых образуется на противоположном бластопору конце зародыша.
7.4.2.2-6. Гаетруляция у земноводных
Гаструляция у земноводных имеет много общего с гаструляцией ланцетника, но так как в яйцеклетках у них желтка намного больше и расположен он преимущественно на вегетативном полюсе, крупные бла-стомеры амфибластулы не способны впячиваться внутрь. Инвагинация проходит несколько иначе. На границе между анимальным и вегетативным полюсами в области серого серпа клетки сначала сильно вытягиваются внутрь, принимая вид «колбовидных» (рис. 7.6, а), а затем тянут за собой клетки поверхностного слоя бластулы. Возникают серповидная бороздка и спинная (дорзальная) губа бластопора (рис. 7.6, б).
Одновременно более мелкие клетки анимального полюса, делящиеся быстрее, начинают перемещаться в сторону вегетативного полюса. В области спинной губы они подворачиваются и впячиваются, а с боков и со стороны, противоположной серповидной бороздке, обрастают более крупные клетки. Затем процесс эпиболии приводит к тому, что образуются боковые (латеральные) и брюшная (вентральная) губы бластопора. Бластопор смыкается в кольцо, внутри которого некоторое время видны крупные светлые клетки вегетативного полюса в виде так называемой желточной пробки. Позднее они полностью погружаются внутрь, а бластопор сужается.
С помощью метода маркировки прижизненными (витальными) красителями у земноводных детально изучены перемещения клеток
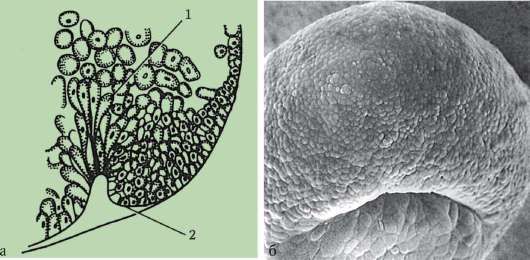
Рис. 7.6. Образование бластопора ранней гаструлы амфибий: а - колбовидные клетки в области спинной губы бластопора; б - общий вид серповидной бороздки и спинной губы; 1 - колбовидные клетки; 2 - спинная губа бластопора
бластулы во время гаструляции, Установлено, что конкретные области бластодермы, называемые презумптивными (от лат. praesumptio - предположение), при нормальном развитии оказываются сначала в составе определенных зачатков органов, а затем в составе самих органов (рис. 7.7). Известно, что у бесхвостых амфибий материал презумптив-ной хорды и мезодермы на стадии бластулы лежит не на ее поверхности, а во внутренних слоях стенки амфибластулы, однако примерно на тех уровнях, как это показано на рисунке.
Определенное сходство процессов гаструляции и областей презумп-тивных органов у земноводных и ланцетника, т.е. гомология основных органов, таких, как нервная трубка, хорда, вторичная кишка, указывает на их филогенетическое родство.
7.4.2.2-в. Гаструляция у птиц
Гаструляция у зародышей с меробластическим типом дробления и развития имеет свои особенности. У птиц она начинается вслед за дроблением и образованием бластулы во время прохождения зародыша по яйцеводу. К моменту откладки яйца зародыш уже состоит из нескольких слоев: верхний слой называют эпибластом, нижний - первичным ги-побластом (см. рис. 7.4, в). Между ними находится узкая щель - бластоцель. Затем образуется вторичный гипобласт, способ образования которого не вполне ясен. Имеются данные о том, что в первичном ги-
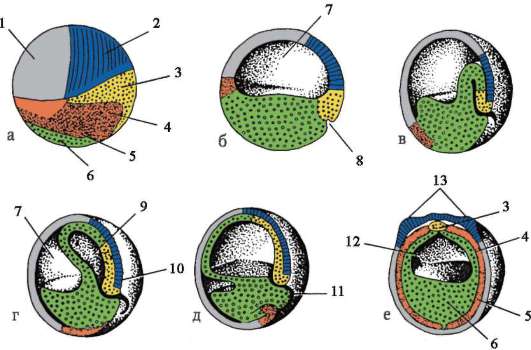
Рис. 7.7. Карта областей презумптивных зачатков органов на ранних стадиях эмбрионального развития амфибий: а - стадия бластулы (вид слева); б-д - последовательные этапы гаструляции (сагиттальные срезы); е - начало нейруля-ции (поперечный срез); 1 - кожная эктодерма; 2 - нервная трубка; 3 - хорда; 4 - мезодерма сомитов; 5 - мезодерма спланхнотомов; 6 - энтодерма; 7 - бла-стоцель; 8 - серповидная бороздка; 9 - гастроцель; 10 - спинная губа бласто-пора; 11 - желточная пробка; 12 - полость вторичной кишки; 13 - нервные валики
побласте птиц берут начало первичные половые клетки, а вторичный образует внезародышевую энтодерму. Образование первичного и вторичного гипобласта рассматривают как явление, предшествующее гаструляции.
Основные события гаструляции и окончательное образование трех зародышевых листков начинаются после откладки яиц с началом инкубации. Возникает скопление клеток в задней части эпибласта как результат неравномерного по скорости деления клеток и перемещения их с боковых участков эпибласта к центру, навстречу друг другу. Образуется так называемая первичная полоска, которая вытягивается в направлении головного конца. В центре первичной полоски образуется первичная бороздка, а по краям - первичные валики. На головном конце первичной полоски возникает утолщение - гензеновский узелок, а в нем - первичная ямка (рис. 7.8).
Когда клетки эпибласта входят в первичную бороздку, их форма изменяется. Они напоминают по форме «колбовидные» клетки гаструлы земноводных. Затем эти клетки приобретают звездчатую форму и погружаются под эпибласт, образуя мезодерму. Часть мигрирующих клеток, встраиваясь в гипобласт, дает в дальнейшем начало зародышевой энтодерме (рис. 7.9). Таким образом, энтодерма образуется на основе первичного и вторичного гипобласта с добавлением нового поколения энтодермаль-ных клеток, мигрирующих из верхних слоев бластодермы. Наличие нескольких генераций эн-тодермальных клеток указывает на растянутость периода гастру-ляции во времени.
Часть клеток, мигрирующая из эпибласта через гензеновский узелок, образует будущую хорду. Одновременно с закладкой и удлинением хорды гензеновский узелок и первичная полоска постепенно смещаются в направлении от головного к хвостовому концу и исчезают. Это соответствует сужению и закрытию бластопора. По мере сокращения первичная полоска оставляет за собой сформированные участки осевых органов зародыша в направлении от головных к хвостовым отделам. Представляется обоснованным рассматривать перемещения клеток в курином зародыше как гомологичные таковым у земноводных, а первичную полоску и ген-зеновский узелок - как гомологичные бластопору и спинной (дорсальной) губе гаструлы этих животных.
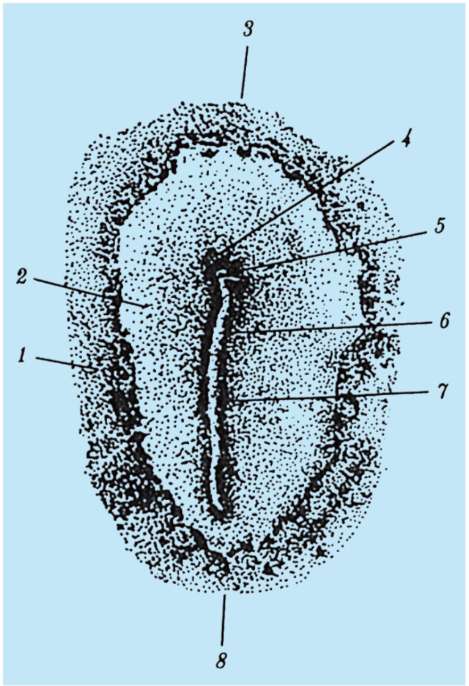
Рис. 7.8. Куриный зародыш на стадии первичной полоски (вид со спинной стороны): 1 - темная область; 2 - просвечивающая область зародышевого диска; 3 - головной конец; 4 - гензеновский узелок; 5 - первичная ямка; 6 - первичная бороздка; 7 - первичный валик; 8 - хвостовой конец
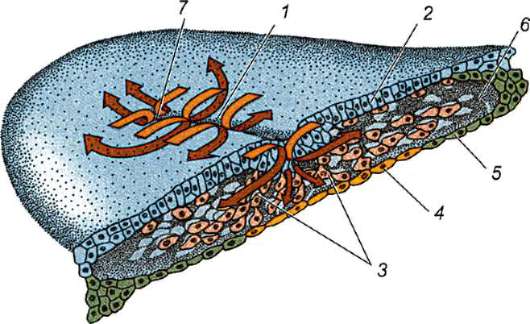
Рис. 7.9. Куриный зародыш на стадии первичной полоски (поперечный срез): 1 - первичная бороздка; 2 - эпибласт (эктодерма); 3 - мигрирующие клетки (мезодерма); 4 - зародышевая энтодерма; 5 - гипобласт (энтодерма); 6 - бла-стоцель; 7 - гензеновский узелок
7.4.2.2-г. Гаструляция у млекопитающих
Интересно отметить, что клетки зародышей млекопитающих (см. п. 7.5.1), несмотря на то, что у этих животных яйцеклетки имеют малое количество желтка, а дробление полное, в фазе гаструляции сохраняют перемещения, свойственные зародышам пресмыкающихся и птиц. Это подтверждает представление о происхождении млекопитающих от предковой группы, у которой яйца были богаты желтком.
7.4.2.3. Особенности стадии гаструляции
Гаструляция характеризуется разнообразными клеточными процессами. Продолжается митотическое размножение клеток, причем оно имеет разную интенсивность в разных частях зародыша. Интерфазы митотических циклов включают все периоды (G1, S, G2), поэтому начиная со стадии гаструляции наблюдается рост развивающегося организма. Вместе с тем наиболее характерная черта гаструляции состоит в перемещении клеточных масс. Это приводит к изменению строения зародыша и превращению его из бластулы в гаструлу. Происходит сортировка клеток по их принадлежности к разным зародышевым листкам, внутри которых они «узнают» друг друга.
На фазу гаструляции приходится начало дифференцировки клеток, что означает переход к активному использованию биологической информации собственного генома. Одним из регуляторов генетической активности является различный химический состав цитоплазмы клеток зародыша, установившийся вследствие ово(оо)плазматической сегрегации, Так, эктодермальные клетки земноводных имеют темный цвет из-за пигмента, попавшего в них из анимального полюса яйцеклетки, а клетки энтодермы - светлый, так как происходят из вегетативного полюса яйца.
Во время гаструляции очень велика роль эмбриональной индукции - взаимодействия между клеточными комплексами (частями) развивающегося зародыша. Показано, что появление первичной полоски у птиц - результат индукционного взаимодействия между гипобластом и эпибластом. Гипобласту присуща полярность. Изменение положения гипобласта по отношению к эпибласту вызывает изменение ориентации первичной полоски. Подробно обо всех перечисленных процессах рассказано в главе 8.
7.4.3. ОБРАЗОВАНИЕ ОРГАНОВ И ТКАНЕЙ 7.4.3.1. Сущность стадии органогенеза
Органогенезы, заключающиеся в образовании отдельных органов, составляют основное содержание эмбрионального периода. Они продолжаются в личиночном и завершаются в ювенильном периоде. Органогенезы отличаются наиболее сложными и разнообразными мор-фогенетическими (формообразующими) преобразованиями. Необходимая предпосылка перехода к органогенезам - достижение зародышем стадии гаструлы, а именно формирование зародышевых листков. Занимая определенное положение по отношению друг к другу, зародышевые листки, контактируя и взаимодействуя, обеспечивают такие взаимоотношения между различными клеточными группами, которые стимулируют их развитие в определенном направлении. Это так называемая эмбриональная индукция - важнейшее следствие взаимодействия между зародышевыми листками.
В ходе органогенезов изменяются форма, структура и химический состав клеток, обособляются клеточные группы, представляющие собой зачатки будущих органов. Постепенно развивается определенная форма органов, устанавливаются пространственные и функциональные связи между ними. Процессы морфогенеза сопровождаются дифферен-
цировкой клеток и образованием тканей, дифференциацией тканей и структур, а также их избирательным и неравномерным ростом. Обязательным условием органогенезов наряду с размножением, миграцией и сортировкой клеток является их избирательная гибель (см. гл. 8).
7.4.3.2. Нейруляция
Самое начало органогенеза называют нейруляцией. Нейруляция охватывает процессы от появления первых признаков формирования нервной пластинки до замыкания ее в нервную трубку (рис. 7.10). Параллельно формируются хорда и вторичная кишка, а лежащая по бокам от хорды мезодерма расщепляется в краниокаудальном направлении на сегментированные парные структуры - сомиты (формируется комплекс осевых органов - первичный органогенез).
Нервная система позвоночных, включая человека, отличается устойчивостью основного плана строения на протяжении всей эволюционной истории подтипа. В формировании нервной трубки у всех хордовых много общего. Вначале неспециализированная спинная эктодерма, отвечая на индукционное воздействие со стороны хордомезодермы, превращается в нервную пластинку, представленную нейроэпители-альными клетками цилиндрической формы.
Нервная пластинка недолго остается уплощенной. Вскоре ее боковые края приподнимаются, образуя нервные валики, которые лежат по обе стороны неглубокой продольной нервной бороздки. Края нервных валиков далее смыкаются, образуя замкнутую нервную трубку с каналом внутри - невроцелем. Раньше всего смыкание нервных валиков происходит на уровне начала (граница между спинным и головным мозгом) спинного мозга, а затем распространяется в головном и хвостовом
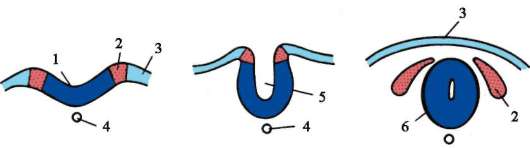
Рис. 7.10. Последовательные стадии формирования нервной трубки и нервного гребня (поперечный срез зародыша): 1 - нервная пластинка; 2 - нервный гребень; 3 - эктодерма; 4 - хорда; 5 - нервная бороздка; 6 - невроцель; 7 - нервные валики
направлениях. Показано, что в морфогенезе нервной трубки большую роль играют микротрубочки и микрофиламенты нейроэпителиальных клеток. Разрушение этих клеточных структур колхицином и цитоха-лазином В приводит к тому, что нервная пластинка остается открытой. Несмыкание нервных валиков ведет к врожденным порокам развития нервной трубки.
После смыкания нервных валиков клетки, первоначально располагавшиеся между нервной пластинкой и будущей кожной эктодермой, образуют нервный гребень. Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему телу (см. п. 8.2.2, рис. 8.9) и образуют два главных потока. Клетки одного из них - поверхностного - включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки. Другой поток мигрирует в брюшном направлении и образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые покровные кости черепа.
Энтодерма у всех зародышей в конечном счете образует эпителий вторичной кишки и многие ее производные. Сама вторичная кишка всегда располагается под хордой.
Таким образом, в процессе нейруляции возникает комплекс осевых органов: нервная трубка, хорда, кишка, представляющих собой характернейшую черту организации тела всех хордовых. Одинаковое происхождение, развитие и взаимное расположение осевых органов выявляют их полную гомологию и эволюционную преемственность.
При углубленном рассмотрении и сравнении процессов нейруляции у конкретных представителей типа хордовых выявляются некоторые различия, которые связаны в основном с особенностями, зависящими от строения яйцеклеток, способа дробления и гаструляции (рис. 7.11). Обращают внимание отличающаяся форма зародышей и смещение времени закладки осевых органов друг относительно друга, т.е. описанная выше гетерохрония.
7.4.3.3. Дифференцировка мезодермы
Мезодерма, занимающая место по бокам от хорды и распространяющаяся далее между кожной эктодермой и энтодермой вторичной кишки, подразделяется на дорсальную и вентральную области. Дорсальная
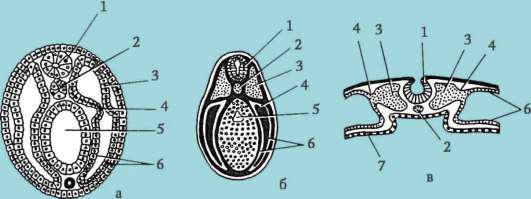
Рис. 7.11. Нейрулы различных хордовых животных: а - ланцетник; б - лягушка; в - цыпленок; 1 - нервная трубка; 2 - хорда; 3 - сомит; 4 - ножка сомита; 5 - вторичная кишка; 6 - боковая пластинка; 7 - энтодерма
часть сегментирована и представлена парными сомитами. 3акладка сомитов идет от головного к хвостовому концу. Вентральная несегментиро-ванная часть мезодермы, имеющая вид тонкого слоя клеток, называется боковой пластинкой. Сомиты соединены с боковой пластинкой промежуточной мезодермой в виде сегментированных ножек сомитов.
Все области мезодермы постепенно дифференцируются. В начале формирования сомиты имеют конфигурацию, характерную для эпителия с полостью внутри. Под индукционным воздействием, исходящим от хорды и нервной трубки, вентромедиальные части сомитов - скле-ротомы - превращаются во вторичную мезенхиму, выселяются из сомита и окружают хорду и вентральную часть нервной трубки. В конце концов из них образуются позвонки, ребра и лопатки.
Дорсолатеральная часть сомитов дифференцируется по двум направлениям. Области сомитов, обращенные к покровной эктодерме, образуют дерматомы, которые дают начало внутреннему слою кожи - дерме. Клетки, находящиеся с внутренней стороны дорсолатеральной части сомитов, формируют миотомы, из которых разовьются поперечнополосатые скелетные мышцы тела и конечностей. Из области ножек сомитов с зачатками нефротом и гонотом образуются органы выделения и половые железы.
Несегментированные боковые пластинки расщепляются на два листка, ограничивающих вторичную полость тела - целом. Внутренний листок, прилежащий к энтодерме, называют висцеральным. Он окружает кишку со всех сторон и образует брыжейку, покрывает легочную
паренхиму и мышцу сердца. Наружный листок боковой пластинки прилежит к эктодерме и называется париетальным. В дальнейшем он образует наружные листки брюшины, плевры и перикарда.
7.4.3.4. Производные зародышевых листков
Эктодерма, мезодерма и энтодерма в ходе дальнейшего развития, взаимодействуя друг с другом, участвуют в формировании конкретных органов (частные или локальные органогенезы). Возникновение зачатка органа связано с местными изменениями определенного участка соответствующего зародышевого листка. Так, из эктодермы развиваются эпидермис кожи и его производные (перо, волосы, ногти, кожные и молочные железы), компоненты органов зрения, слуха, обоняния, эпителий ротовой полости, эмаль зубов. Важнейшие эктодермальные производные - нервная трубка, нервный гребень и образующиеся из них все нервные структуры.
Производными энтодермы являются эпителий желудка и кишки, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Энтодерма переднего отдела эмбриональной кишки образует эпителий легких и воздухоносных путей, а также секретиру-ющие клетки передней и средней долей гипофиза, щитовидной и пара-щитовидной желез.
Мезодерма, помимо уже описанных выше скелетных структур, скелетной мускулатуры, дермы кожи, органов выделительной и половой систем, образует сердечно-сосудистую систему, лимфатическую систему, плевру, брюшину и перикард. Из мезенхимы, имеющей смешанное происхождение за счет клеток трех зародышевых листков, развиваются все виды соединительной ткани, гладкая мускулатура, кровь и лимфа (подробнее в п. 8.2.5).
Зачаток конкретного органа формируется первоначально из определенного зародышевого листка, но затем орган усложняется, и в итоге в его формировании принимают участие два или три зародышевых листка.
7.4.4. ПРОВИЗОРНЫЕ ОРГАНЫ ЗАРОДЫШЕЙ ПОЗВОНОЧНЫХ
Провизорные, или временные, органы образуются в эмбриогенезе ряда представителей позвоночных для обеспечения жизненно важных функций зародыша, таких, как дыхание, питание, выделение, движение и другие. Недоразвитые органы самого формирующегося животного еще не способны функционировать по назначению, хотя обязательно
играют какую-то роль в системе развивающегося целостного организма. Как только зародыш достигает необходимой степени зрелости, когда большинство органов способны выполнять жизненно важные функции, временные органы рассасываются или отбрасываются.
Время образования провизорных органов зависит от того, какие запасы питательных веществ были накоплены в яйцеклетке и в каких условиях среды происходит развитие зародыша. У бесхвостых земноводных, например, благодаря достаточному количеству желтка в яйцеклетке и тому, что развитие идет в воде, зародыш осуществляет газообмен и выделяет продукты диссимиляции непосредственно через оболочки яйца и достигает стадии головастика. На этой стадии образуются провизорные органы дыхания (жабры), пищеварения и движения, приспособленные к водному образу жизни. Перечисленные личиночные органы дают возможность головастику продолжить развитие. По достижении состояния морфофункциональной зрелости органов взрослого типа временные органы исчезают в процессе метаморфоза.
У пресмыкающихся и птиц запасов желтка в яйцеклетке больше, но развитие зародыша идет не в воде, а на суше. В связи с этим очень рано возникает потребность в обеспечении дыхания и выделения, а также и в защите от высыхания. У них уже в раннем эмбриогенезе, почти параллельно с нейруляцией, начинается формирование провизорных органов, таких, как амнион, хорион и желточный мешок. Чуть позднее формируется аллантоис. У плацентарных млекопитающих эти же провизорные органы образуются еще раньше, поскольку в яйцеклетке очень мало желтка. Развитие таких животных происходит внутриутробно, образование провизорных органов у них совпадает по времени с периодом гаструляции.
Наличие или отсутствие амниона и других провизорных органов лежит в основе деления позвоночных на две группы: Amniota и Anamnia (см. табл. 7.1). Эволюционно более древние позвоночные, развивающиеся исключительно в водной среде и представленные такими классами, как Круглоротые, Рыбы и Земноводные, не нуждаются в дополнительных водных и других оболочках зародыша и составляют группу анам-ний. К группе амниот относят первичноназемных позвоночных, т.е. тех, у кого эмбриональное развитие протекает в наземных условиях. Это три класса: Пресмыкающиеся, Птицы и Млекопитающие. Они относятся к высшим позвоночным, так как имеют скоординированные и высокоэффективные системы органов, обеспечивающие им существование в наиболее сложных условиях, каковыми являются условия суши. Эти
классы насчитывают большое количество видов, вторично перешедших в водную среду. Таким образом, высшие позвоночные оказались в состоянии освоить все среды обитания. Подобное совершенство было бы невозможным в том числе без внутреннего оплодотворения и образования специальных провизорных эмбриональных органов, называемых также зародышевыми оболочками. К зародышевым оболочкам амниот относят амнион, хорион (серозу), желточный мешок и аллантоис.
В строении и функциях провизорных органов различных амниот много общего. Характеризуя в самом общем виде провизорные органы зародышей высших позвоночных, следует отметить, что все они развиваются из клеточного материала уже сформировавшихся зародышевых листков. Некоторые особенности имеются в развитии зародышевых оболочек плацентарных млекопитающих, о чем будет сказано ниже.
Амнион представляет собой мешок, заключающий зародыша и заполненный амниотической жидкостью. Амниотическая оболочка образована внезародышевыми эктодермой и соматоплеврой. Экто-дермальная ее часть специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амнион играет первостепенную роль в защите зародыша от высыхания и от механических повреждений, создавая для него наиболее благоприятную и естественную водную среду. Мезодермальная часть амниона дает начало гладким мышечным волокнам. Сокращения этих мышц вызывают пульсацию амниона, а медленные колебательные движения, сообщаемые при этом зародышу, по-видимому, способствуют тому, что его растущие части не мешают друг другу.
Хорион (сероза) - самая наружная зародышевая оболочка, прилежащая к скорлупе (сероза) или материнским тканям (хорион), возникающая, как и амнион, из эктодермы и соматоплевры. Эта оболочка служит для обмена между зародышем и окружающей средой. У яйцекладущих видов основная функция серозы - участие в дыхании (газообмене); у млекопитающих хорион выполняет гораздо более обширные функции, участвуя, помимо дыхания, в питании, выделении, фильтрации и синтезе веществ, например гормонов.
Желточный мешок образован из внезародышевых энтодермы и висцеральной мезодермы и непосредственно связан с кишечной трубкой зародыша. У зародышей с большим количеством желтка он принимает участие в питании. У птиц, например, в спланхноплевре желточного мешка развивается сосудистая сеть. Желток не проходит через желточный проток, соединяющий мешок с кишкой. Сначала он переводится в
растворимую форму под действием пищеварительных ферментов, продуцируемых энтодермальными клетками стенки мешка. Затем попадает в сосуды и с кровью разносится по всему телу зародыша.
У млекопитающих нет запасов желтка, и сохранение желточного мешка может быть связано с важными вторичными функциями. Мезодерма желточного мешка продуцирует форменные элементы крови зародыша. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке.
Судьба желточного мешка у разных животных несколько различна. У птиц к концу периода инкубации остатки желточного мешка уже находятся внутри зародыша, после чего он быстро исчезает и к концу 6 суток после вылупления птенца полностью рассасывается. У млекопитающих желточный мешок бывает развит по-разному. У хищников он сравнительно большой, с сильно развитой сетью сосудов, а у приматов быстро сморщивается и исчезает без остатка до родов.
Аллантоис развивается несколько позднее других провизорных органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки. Следовательно, он образован энтодермой изнутри и спланхноплеврой снаружи. У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего, это вместилище для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении наружная часть аллантоиса отбрасывается, а внутренняя - сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту. Термин плацента означает тесное наложение или слияние зародышевых оболочек с тканями родительского организма. У приматов и некоторых других млекопитающих энтодермальная часть аллантоиса рудиментарна, а мезодермальные клетки образуют плотный тяж, протягивающийся от клоакального отдела к хориону. По мезодерме аллантоиса к хориону растут сосуды, посредством которых плацента выполняет выделительную, дыхательную и питательную функции.
Доступнее и проще изучить образование и строение зародышевых оболочек на примере зародыша курицы. На стадии нейрулы три зародышевых листка непосредственно переходят от зародыша к внезаро-
дышевой части, никак не отграничиваясь. По мере того как зародыш приобретает форму, вокруг него образуется несколько складок, которые как бы подсекают зародыш, отделяют его от желтка и устанавливают четкие границы между зародышем и внезародышевыми областями. Они называются туловищными складками (рис. 7.12).
Сначала образуется головная складка. Она подсекает снизу головную часть. Задние концы этой складки переходят в боковые туловищные складки, отграничивающие туловище зародыша с боков. Хвостовая складка отграничивает задний конец зародыша. Постепенно сужается ножка, соединяющая среднюю кишку и желточный мешок, образуются
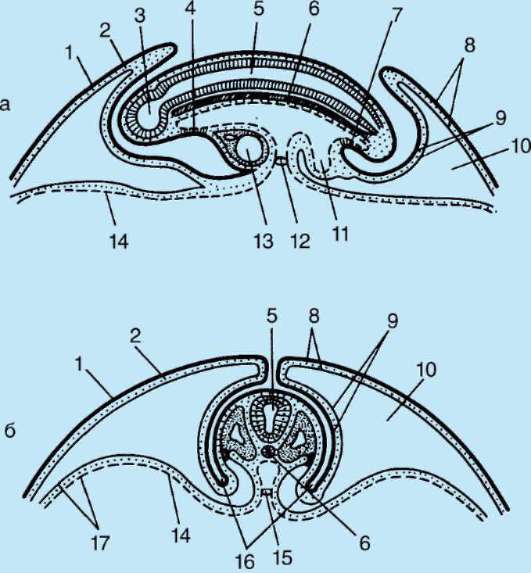
Рис. 7.12. Образование туловищных складок и зародышевых оболочек у зародыша цыпленка: а - продольный срез; б - поперечный срез; 1 - эктодерма; 2 - мезодерма; 3 - зачаток мозга; 4 - глоточная мембрана; 5 - нервная трубка; 6 - хорда; 7 - клоачная мембрана; 8 - хорион; 9 - амнион; 10 - экзоцелом; 11 - аллантоис; 12 - область пупка; 13 - зачаток сердца; 14 - энтодерма; 15 - закладка кишечника; 16 - туловищные складки; 17 - желточный мешок
передний и задний отделы кишки. Одновременно из эктодермы и прилежащей к ней соматоплевры образуется сначала головная складка (рис. 7.13), которая, как капюшон, нарастает на зародыш сверху. Концы головной складки образуют по бокам амниотические валики. Они растут поверх зародыша навстречу друг другу и срастаются, образуя сразу стенки амниона, прилежащего к зародышу, и хориона, лежащего снаружи.
Позднее образуется аллантоис (рис. 7.14). Общий вид куриного зародыша на 6-е сут инкубации изображен на рис. 7.15. У различных млекопитающих процессы образования провизорных органов больше или меньше похожи на описанные выше у птиц. Особенности развития их у приматов, в том числе человека, показаны в следующей главе.
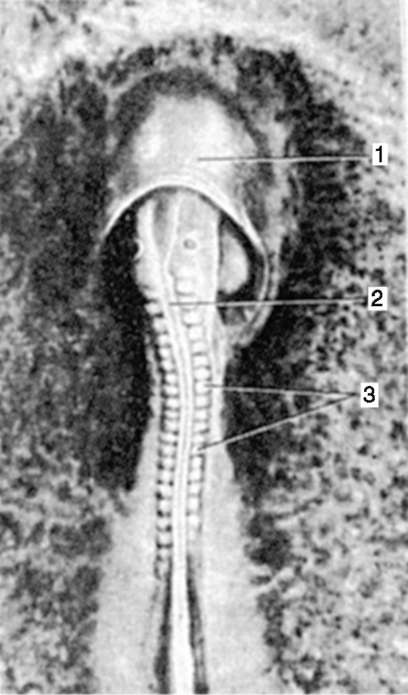
Рис. 7.13. Куриный зародыш около 40 ч инкубации: 1 - головная складка амниона; 2 - нервная трубка; 3 - сомиты
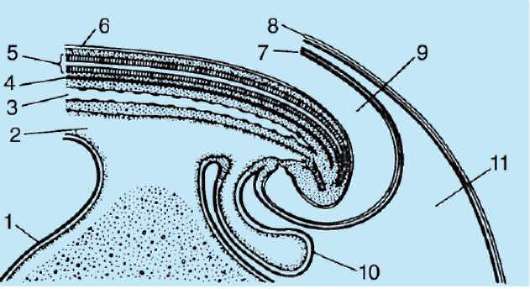
Рис. 7.14. Образование аллантоиса у зародыша цыпленка (продольный срез хвостовой части): 1 - желточный мешок; 2 - средняя кишка; 3 - аорта; 4 - хорда; 3 - нервная трубка; 6 - эктодерма; 7 - амнион; 8 - хорион; 9 - полость амниона; 10 - аллантоис; 11 - экзоцелом
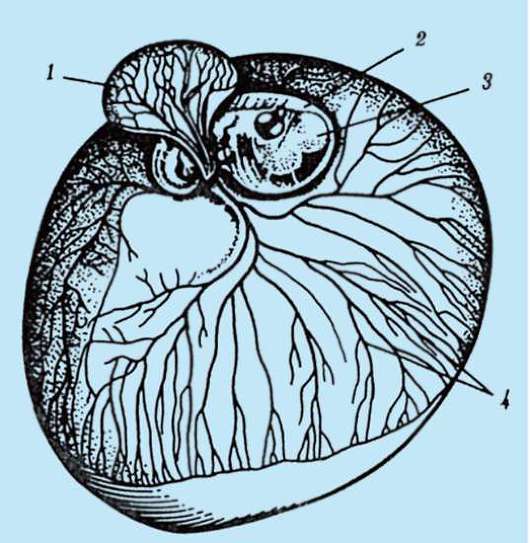
Рис. 7.15. Куриный зародыш на 6-е сут инкубации (белок и хорион удалены, аллантоис сдвинут вверх): 1 - аллантоис; 2 - амнион; 3 - зародыш; 4 - сосуды желточного мешка
7.5. эмбриональное развитие млекопитающих
7.5.1. ПЕРИОДИЗАЦИЯ И РАННЕЕ ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Изучение пренатального и, в частности, эмбрионального (внутриутробного) развития человека очень важно, так как помогает лучше понять взаимосвязи между органами, а также механизмы возникновения врожденных пороков развития. В эмбриональном развитии разных видов млекопитающих есть общие черты, но существуют и различия. У всех плацентарных, например, процессы раннего эмбриогенеза существенно отличаются от таковых, ранее описанных у других позвоночных. Вместе с тем и среди плацентарных есть видовые особенности.
Дробление зиготы человека характеризуется следующими чертами. Плоскость первого деления проходит через полюса яйцеклетки, т.е., как и у других позвоночных, является меридианной. При этом один из образующихся бластомеров оказывается крупнее другого, что указывает на неравномерность дробления. Два первых бластомера вступают в следующее деление асинхронно. Борозда проходит по меридиану и перпендикулярно первой борозде. Таким образом, возникает стадия трех бластомеров. Во время деления меньшего бластомера происходит поворот пары образующихся более мелких бластомеров на 90°, так, что плоскость борозды деления оказывается перпендикулярной к первым двум
Рис. 7.16. Ранние стадии дробления зиготы кролика: I - плоскость первой борозды дробления; IIа - плоскость второй борозды дробления одного из первых двух бластомеров; IIб - плоскость второй борозды дробления второго из первых двух бластомеров
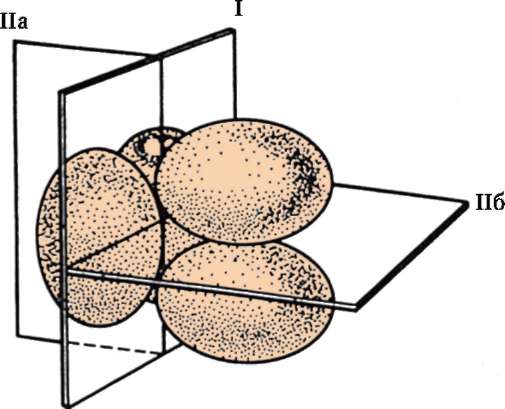
бороздам. Аналогичное расположение бластомеров на 4-клеточной стадии описано у мыши, кролика, норки и обезьяны (рис. 7.16). Благодаря асинхронному дроблению могут быть стадии с нечетным числом бла-стомеров - 3, 5, 7, 9, которые, вследствие отсутствия стадии роста, отличаются своими размерами. Таким образом, дробление человека полное асинхронное неравномерное.
В результате дробления образуется скопление клеток - морула. Поверхностно расположенные бластомеры образуют клеточный слой, а бластомеры, лежащие внутри морулы, группируются в центральный клеточный узелок. Вскоре внутри морулы появляется жидкость, образуется полость (бластоцель), и зародыш превращается в бластоцисту. Формирование морулы происходит на стадии 16 бластомеров, а кавитация (образование полости) начиная со стадии 32 бластомеров.
В бластоцисте различают наружный слой клеток (трофобласт) и внутреннюю клеточную массу (зародышевый узелок, или эмбрио-бласт). Внутренняя клеточная масса оттеснена жидкостью к одному из полюсов бластоцисты. Позднее из трофобласта разовьется наружная зародышевая оболочка - хорион, а из эмбриобласта - сам зародыш и некоторые внезародышевые органы. Показано, что собственно зародыш развивается из очень небольшого числа клеток зародышевого узелка.
Стадия дробления протекает под блестящей оболочкой. На рисунке 7.17 изображены ранние стадии эмбриогенеза человека с указанием, где в материнском организме располагается зародыш. Дробление человеческой зиготы и возникновение бластоцисты схематично представлены на рис. 7.18 и 7.19.
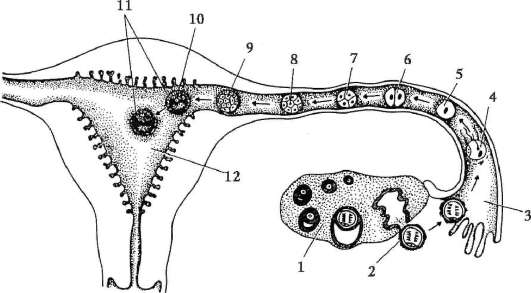
Рис. 7.17. Овуляция, оплодотворение и человеческий зародыш на 1-й неделе развития: 1 - яичник; 2 - овоцит II порядка (овуляция); 3 - яйцевод; 4 - оплодотворение; 5 - зигота; 6 - зародыш на стадии двух бластомеров; 7 - зародыш на стадии четырех бластомеров; 8 - зародыш на стадии восьми бластомеров; 9 - морула; 10, 11 - бластоциста; 12 - задняя стенка матки
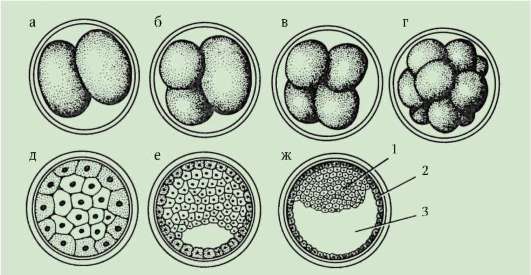
Рис. 7.18. Дробление зиготы человека: а - два бластомера; б - три бластомера; в - четыре бластомера; г - морула; д - разрез морулы; е, ж - разрез ранней и поздней бластоцисты: 1 - эмбриобласт; 2 - трофобласт; 3 - бластоцель
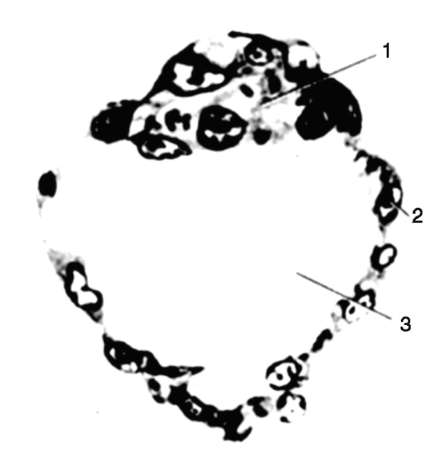
Рис. 7.19. Бластоциста зародыша человека (срез): 1 - эмбриобласт; 2 - трофобласт; 3 - бластоцель
Примерно на 6-7-е сутки после оплодотворения зародыш, который уже 2-3 сут свободно плавал в полости матки, готов к имплантации, т.е. к погружению в ее слизистую оболочку. Под действием ферментов, выделяемых трофобластом, блестящая оболочка частично разрушается и зародыш выходит из нее («вылупляется»). При участии белков-интегринов, синтезируемых и эпителием матки, и клетками трофобласта, бластула своим эмбриональным полюсом прикрепляется к стенке матки. Вступив в контакт с материнскими тканями, клетки трофобласта быстро размножаются и, выделяя протеолитические ферменты, разрушают их (эти ткани). Это вызывает реакцию стенки матки, сопровождающуюся активным образованием сосудов в месте имплантации. Нарушение синтеза интегринов и необходимых для имплантации ферментов делает этот процесс невозможным, что, в свою очередь, ведет к гипоксии и гибели зародыша. В ходе имплантации трофобласт дифференцируется на два слоя: внутренний, называемый цитотрофобластом, поскольку он сохраняет клеточное строение, и наружный, называемый синцитио-трофобластом, поскольку он представляет собой синцитий. На рисунке 7.20 показан зародыш человека в процессе имплантации.
Гаструляция у млекопитающих тесно связана с другими эмбриональными преобразованиями. Одновременно с разделением трофо-бласта на два слоя происходит уплощение зародышевого узелка, и он превращается в двухслойный зародышевый щиток. Нижний слой щитка - гипобласт, или первичная энтодерма, по мнению большинства авторов, образуется путем деламинации внутренней клеточной массы, примерно так, как это происходит в зародышевом диске птиц. Первич-
Рис. 7.20. Последовательные стадии имплантации и развития зародыша человека в конце 1-й и на 2-й неделе: а - бластоциста; б - бластоциста в самом начале имплантации (7-е сутки развития); в - частично имплантировавшая бла-стоциста (8-е сутки развития)
Рис. 7.20. Окончание: г - зародыш на 9-10-е сутки развития; д - зародыш на 13-е сутки развития: 1 - эмбриобласт, 2 - бластоцель, 3 - трофобласт, 4 - полость амниона, 5 - гипобласт, 6 - синцитиотрофобласт, 7 - цитотрофобласт, 8 - эпибласт, 9 - амнион, 10 - лакуна трофобласта, 11 - эпителий матки, 12 - ножка тела, 13 - почка аллантоиса, 14 - желточный меток, 15 - внезаро-дышевый целом, 16 - ворсинка хориона, 17 - первичный желточный мешок, 18 - вторичный желточный мешок
ная энтодерма полностью расходуется на образование внезародышевой энтодермы. Выстилая полость трофобласта, она образует первичный желточный мешок млекопитающих.
Верхний клеточный слой - эпибласт - источник будущей эктодермы, мезодермы и вторичной энтодермы. На 3-й неделе в эпибласте образуется первичная полоска, развитие которой сопровождается почти такими же перемещениями клеточных масс, как и при образовании первичной полоски птиц (рис. 7.21). В головном конце первичной полоски образуются гензеновский узелок и первичная ямка, гомологичные спинной губе бластопора других позвоночных. Клетки, которые перемещаются в области первичной ямки, направляются под эпибластом в сторону прехордальной пластинки.
Прехордальная пластинка находится на головном конце зародыша и обозначает место будущей ротоглоточной мембраны. Клетки, перемещающиеся по центральной оси, образуют зачаток хорды и мезодермы и составляют хордомезодермальный отросток. Гензеновский узелок постепенно смещается к хвостовому концу зародыша, первичная полоска укорачивается, а зачаток хорды удлиняется. По бокам от хордомезо-дермального отростка образуются мезодермальные пластинки, которые расширяются в обе стороны. На рисунке 7.22 представлена обобщающая схема некоторых процессов раннего эмбрионального развития.
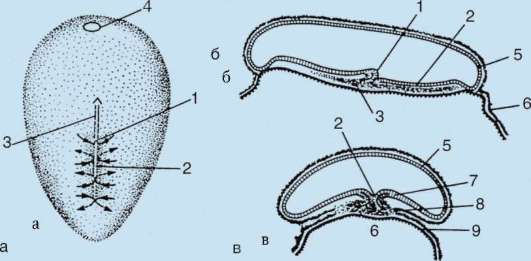
Рис. 7.21. Развитие зародыша человека на стадии первичной полоски (15-17-е сутки): а - вид на зародыш сверху (амнион снят); б - продольный срез; в - поперечный срез через первичную полоску: 1 - гензеновский узелок; 2 - первичная полоска; 3 - хорда; 4 - прехордальная пластинка; 5 - амнион; 6 - желточный мешок; 7 - эктодерма; 8 - мезодерма; 9 - энтодерма
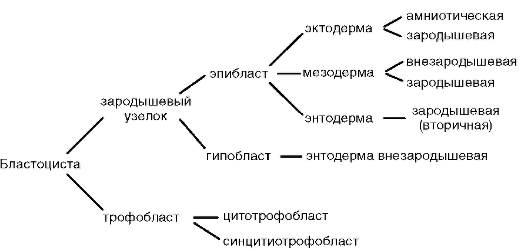
Рис. 7.22. Дифференциация зародышевых листков млекопитающих
К концу 3-й недели в эктодерме зародыша над зачатком хорды образуется нервная пластинка. Она состоит из высоких цилиндрических клеток. В центре нервной пластинки образуется прогиб в виде нервного желоба, а по бокам его возвышаются нервные валики. Это начало нейруляции. В средней части зародыша происходит смыкание нервных валиков - образуется нервная трубка. Затем смыкание распространяется в головном и хвостовом направлениях. Нервная трубка и прилежащие к ней участки эктодермы, из которых в дальнейшем развивается нервный гребень, полностью погружаются и отделяются от эктодермы, срастающейся над ними (см. рис. 7.10). Полоска клеток, лежащая под нервной трубкой, превращается в хорду. По бокам от хорды и нервной трубки в средней части зародыша появляются сегменты спинной мезодермы - сомиты. К концу 4-й недели они распространяются к головному и хвостовому концам, их число достигает примерно 40 пар.
К этому же времени относится начало формирования вторичной кишки, закладок сердца и сосудистой сети желточного мешка. На рисунке 7.23 видны соотношения размеров зародыша и внезародышевых органов на 21-е сутки развития. Более детально обособление тела зародыша от зародышевых оболочек и закладку органов можно видеть на рис. 7.24, где изображены не только общий вид зародыша, но и планы разрезов. Обращает внимание быстрое (за 7 сут 4-й недели) формирование зародыша в виде вытянутого в длину и изогнутого тела, приподнятого и отсеченного туловищными складками от желточного мешка. За
3 4
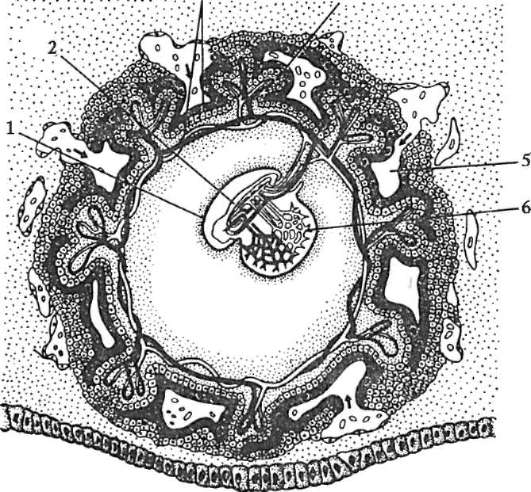
Рис. 7.23. Зародыш и внезародышевые органы человека на 21-е сутки развития: 1 - амнион; 2 - зародыш; 3 - хорион; 4 - третичная ворсина; 5 - материнская кровь; 6 - желточный мешок
это время закладываются все сомиты, четыре пары жаберных дуг, сердечная трубка, почки конечностей, средняя кишка, а также «карманы» передней и задней кишки.
В следующие четыре недели эмбрионального развития закладываются все основные органы. Нарушение процесса развития в этот период ведет к наиболее грубым и множественным врожденным порокам развития.
Как было отмечено выше, развитие внезародышевых провизорных органов у млекопитающих и человека имеет особенности. Эти органы образуются очень рано, одновременно с гаструляцией, и несколько иначе, чем у других амниот. Начало развития хориона и амниона приходится на 7-8-е сутки, т.е. совпадает с началом имплантации. Хорион возникает из трофобласта, который уже разделился на цитотрофобласт и синцитиотрофобласт. Последний под влиянием контакта со слизистой оболочкой матки разрастается и разрушает ее. К концу 2-й недели обра-
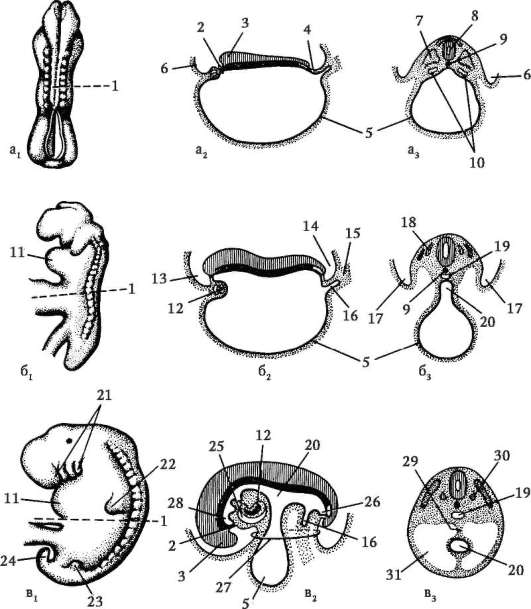
Рис. 7.24. Развитие зародыша человека на 4-й неделе: а1б1в1 - общий вид; а2б2в2 - продольный срез; а3б3в3 - поперечный срез; а1а2а3 - 22 сут; б1б2б3 - 24 сут; в1в2в3 - 28 сут: 1 - уровень поперечного среза; 2 - ротоглоточ-ная мембрана; 3 - мозг; 4 - клоачная мембрана; 5 - желточный мешок; 6 - амнион; 7 - сомиты; 8 - нервная трубка; 9 - хорда; 10 - парные закладки брюшной аорты; 11 - сердечный выступ; 12 - сердце; 13 - головная туловищная складка; 14 - хвостовая туловищная складка; 15 - ножка тела; 16 - аллантоис; 17 - боковые туловищные складки; 18 - нервный гребень; 19 - спинная аорта; 20 - средняя кишка; 21 - жаберные дуги; 22 - почка передней конечности; 23 - почка задней конечности; 24 - хвост; 25 - перикард; 26 - карман задней кишки; 27 - пупочный канатик; 28 - карман передней кишки; 29 - спинная брыжейка; 30 - нервный узел заднего корешка; 31 - внутризародышевый целом
зуются первичные ворсинки хориона в виде скопления эпителиальных клеток цитотрофобласта. В начале 3-й недели в них врастает мезодер-мальная мезенхима и возникают вторичные ворсинки, а когда к концу 3-й недели внутри соединительнотканной сердцевины появляются кровеносные сосуды, их называют третичными ворсинками. Область, где тесно прилежат ткани хориона и слизистая оболочка матки, называют плацентой.
У человека, как и у других приматов, сосуды материнской части плаценты утрачивают свою непрерывность и ворсинки хориона фактически омываются кровью и лимфой материнского организма. Такая плацента называется гемохориальной. По мере развития беременности ворсинки увеличиваются в размерах, разветвляются, но кровь плода с самого начала и до конца изолирована от материнской крови плацентарным барьером.
Плацентарный барьер состоит из трофобласта, соединительной ткани и эндотелия сосудов плода. Этот барьер проницаем для воды, электролитов, питательных веществ и продуктов диссимиляции, а также для антигенов эритроцитов плода и антител материнского организма, токсических веществ и гормонов. Клетками плаценты вырабатывается четыре гормона, в том числе хорионический гонадотропин, который обнаруживается в моче беременной женщины со 2-3-й недели беременности.
Амнион возникает путем расхождения клеток эпибласта внутренней клеточной массы. Амнион человека называют шизамнионом (см. рис. 7.20) в отличие от плеврамниона птиц и некоторых млекопитающих. Амниотическая полость некоторое время ограничена клетками эпибласта и частично участком трофобласта. Затем боковые стенки эпи-бласта образуют складки, направленные вверх, которые впоследствии срастаются. Полость оказывается полностью выстланной эпибластиче-скими (эктодермальными) клетками. Снаружи амниотическую эктодерму окружают внезародышевые мезодермальныс клетки. Амниотическая жидкость содержит разнообразные вещества, такие, как белки, углеводы, соли, микроэлементы, гормоны и др. Ее состав меняется в ходе развития, кроме того, она постоянно обновляется. В процессе развития зародыша она играет важную роль при формировании носовой и ротовой полостей, органов пищеварения и дыхания.
Желточный мешок появляется, когда от внутренней клеточной массы отделяется тонкий слой гипобласта и его внезародышевые энто-дермальныс клетки, перемещаясь, выстилают изнутри поверхность тро-
фобласта. Образовавшийся первичный желточный мешок на 12-13-е сутки спадается и преобразуется во вторичный желточный мешок, связанный с зародышем желточным протоком. Энтодермальные клетки покрываются снаружи внезародышевой мезодермой. На 3-й неделе развития в стенке желточного мешка начинают обнаруживаться первичные половые клетки, клетки крови и кровеносные сосуды. До 7-8-й недели развития этот желточный мешок является основным кроветворным органом зародыша.
Аллантоис возникает у зародыша человека, как и у других амниот, в виде кармана вентральной стенки задней кишки, но его энтодермаль-ная полость остается рудиментарной структурой. Тем не менее в его стенках развивается обильная сеть сосудов, соединяющаяся с главными кровеносными сосудами зародыша. Мезодерма аллантоиса соединяется с мезодермой хориона, отдавая в него кровеносные сосуды, которые примерно до второго месяца эмбриогенеза обеспечивают питание развивающегося зародыша. Таким образом происходит васкуляризация хо-риоаллантоисной плаценты.
Некоторые этапы и сроки развития органов у зародышей человека представлены в табл. 7.4.
Таблица 7.4. Основные периоды и события в раннем онтогенезе человека
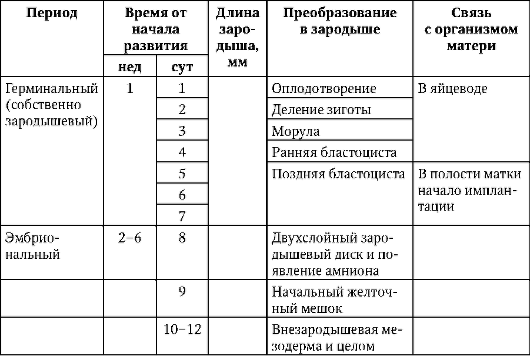
Продолжение табл. 7.4
Окончание табл. 7.4
7.5.2. ПРИМЕРЫ ОРГАНОГЕНЕЗОВ ЧЕЛОВЕКА, ОТРАЖАЮЩИХ ЭВОЛЮЦИЮ ВИДА
В этом разделе не ставится цель описать развитие всех органов человека. Будут рассмотрены лишь некоторые морфогенетические процессы, иллюстрирующие следующие общебиологические моменты:
• значение межклеточных, тканевых и межорганных взаимодействий в морфогенезе;
• отражение в морфогенезе человека эволюционно более древних черт организации позвоночных;
• особенности органогенезов, позволяющие оценить стадию и механизмы нарушений развития при тех или иных врожденных пороках.
Сегментарное строение позвоночника отражает его происхождение из сомитов зародыша. Кроме того, формирование позвонков тесно связано с хордой и спинным мозгом. Тела позвонков образуются из мигрирующих по направлению к хорде и окружающих ее скоплений ме-зенхимных клеток, которые происходят из склеротомов. В образовании тел позвонков участвуют мезенхимные клетки двух лежащих рядом пар сомитов, в связи с чем тела позвонков находятся не на уровне сомитов, а между ними. В то же время мышцы, развивающиеся из миотомов соми-
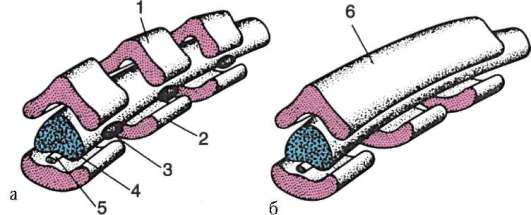
Рис. 7.25. Развитие спинных дужек позвонков у куриного зародыша: а - нормальное развитие; б - после удаления спинномозговых ганглиев: 1 - спинная дужка; 2 - тело позвонка; 3 - спинномозговой узел; 4 - спинной мозг; 5 - хорда; 6 - несегментированные спинные дужки позвонков
тов, как бы перекидываются через межпозвонковые сочленения и обеспечивают движение позвонков.
Дужки позвонков образуются из скопления мезенхимных скле-ротомных клеток, распространяющихся в спинном (дорсальном) направлении, а поперечные отростки и ребра образуются из скоплений склеротомных клеток, мигрирующих вбок. Спинной мозг и спинномозговые ганглии на соответствующих этапах внутриутробного развития участвуют в морфогенезе спинных дужек позвонков. Так, если удалить спинномозговые ганглии, то хрящ спинных дужек формируется, но имеет вид несегментированного стержня (рис. 7.25).
Постепенно изменяется гистологическое строение позвонков. На протяжении 4-5-й недель образуются мезенхимные позвонки, на 6-й неделе появляются очаги хондрогенеза, а затем на 8-й неделе начинается окостенение, продолжающееся почти до 25 лет. Участки хорды, расположенные внутри тел позвонков, постепенно исчезают. Внутри межпозвонковых дисков хорда сохраняется в виде слизеподобной структуры, известной под названием nucleus pulposus.
Формирование среднего уха связано с онтогенетическими преобразованиями первой и второй висцеральных (жаберных) дуг. На рисунке 7.26 схематично показаны области головы и шеи на 4-й и 24-й (конец первого-второй триместр периода внутриутробного развития) неделях развития человека. На 4-й неделе у эмбриона в головной и шейной областях справа и слева закладываются висцеральные (жаберные) дуги. Изнутри со стороны глотки между ними образуются глоточные карманы (рис. 7.27). Это выпячивания (или впячивания) энтодермы,
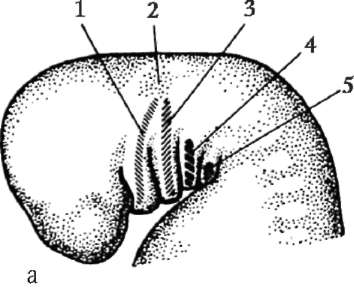
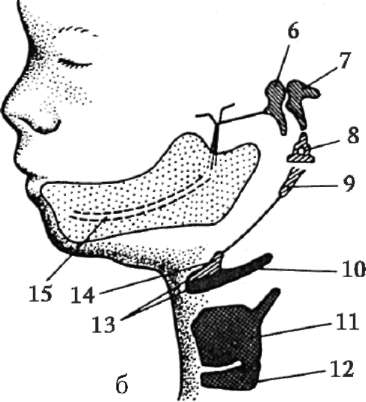
Рис. 7.26. Голова и шея зародыша человека, вид сбоку: а - на 4-й неделе; б - на 24-й неделе: 1 - первая дуга; 2 - местоположение развивающегося внутреннего уха; 3 - вторая дуга; 4 - третья дуга; 5 - четвертая луга; 6 - молоточек. 7 - наковальня; 8 - стремечко; 9 - шиловидный отросток; 10 - большой рог подъязычной кости; 11 - щитовидный хрящ; 12 - перстневидный хрящ; 13 - тело подъязычной кости; 14 - малый рог; 15 - нижняя челюсть; закладки висцеральных дуг (а) и развивающиеся из них органы (б) обозначены одинаковой штриховкой
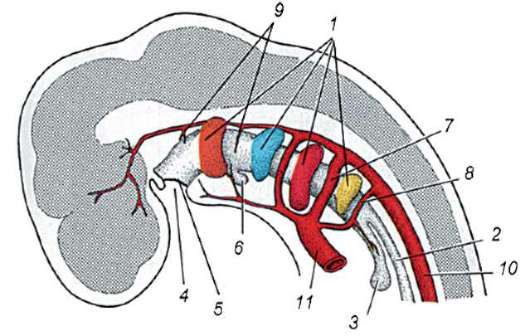
Рис. 7.27. Жаберная область 5-недельного зародыша человека (покровная эктодерма и мезенхима удалены). 1 - глоточные карманы; 2 - пищевод; 3 - трахея и легочная почка; 4 - стомодеум; 5 - стомодеальная (ротовая) пластинка; 6 - зачаток щитовидной железы; 7 - 4-я аортальная дуга; 8 - 6-я аортальная дуга; 9 - редуцирующиеся 1-я и 2-я аортальные дуги; 10 - спинная нисходящая аорта; 11 - брюшная восходящая аорта (артериальный мешок)
растущие вбок. Снаружи, соответственно глоточным выростам, впячиваются внутрь эктодермальные жаберные карманы.
Энтодерма и эктодерма первого глоточного и жаберного карманов приходят в контакт на 4-й неделе развития (рис. 7.28, а). Контакт продолжается недолго, слепой конец глоточного кармана отходит от поверхности и окружается мезенхимой (рис. 7.28, б). Из дистальной части глоточного кармана начинает развиваться полость среднего уха (барабанная полость), а из проксимальной части образуется слуховая (евстахиева) труба. Из мезенхимы в дальнейшем дифференцируются закладки слуховых косточек, служащих для проведения звука (рис. 7.28, в). Молоточек и наковальня происходят из клеточного материала первой висцеральной дуги, стремечко - из клеточного материала дорсальной части второй висцеральной дуги. Мышцы и нервы, связанные со звукопроводящим косточковым аппаратом косточками среднего уха, формируются из клеточного материала области первой и второй висцеральных дуг.
В конце внутриутробной жизни и в течение нескольких месяцев после рождения эмбриональная соединительная ткань, находящаяся в барабанной полости, рассасывается. Только после этого слуховые косточки приобретают подвижность, и колебания барабанной перепонки могут свободно передаваться на мембрану овального окна внутреннего уха.
В формировании структур лицевой области и ротовой полости принимают участие клетки экто- и мезодермы. Большая часть струк-
Рис. 7.28. Развитие среднего уха, фронтальный разрез: а - на 4-й неделе; б - на 5-й неделе; в - более поздние стадии; 1 - первая жаберная пластинка; 2 - первый жаберный карман; 3 - первая висцеральная (жаберная) дуга; 4 - слуховой пузырек; 5 - первый глоточный карман; 6 - производные первой висцеральной дуги; 7 - производное второй висцеральной дуги; 8 - трубоба-рабанная полость; 9 - пробка наружного слухового прохода. 10 - первичный наружный слуховой проход; 11 - молоточек; 12 - наковальня; 13 - стремечко; 14 - барабанная полость; 15 - слуховая (евстахиева) труба
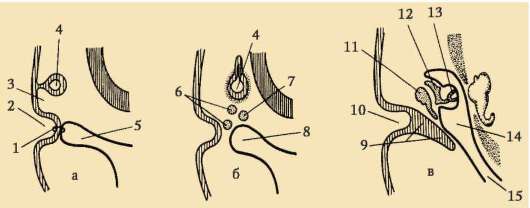
тур лицевой области развивается из масс мезенхимных клеток экто-дермального происхождения, мигрирующих из нервного гребня (4-й зародышевый листок). Из этой мезенхимы развиваются соединительная, хрящевая и костная ткани и ткань пульпы зубов. Поперечнополосатая мускулатура мышц лица и глотки формируется из мезенхимы мезодермального происхождения. Здесь важно понять, как шел процесс «расширения и смены функций» в антропогенезе.
Ротовая полость образована эктодермальным впячиванием стомо-деумом, примыкающим к слепо заканчивающейся передней кишке. Их разделяет тонкая стомодеальная пластинка, состоящая из экто-дермального (со стороны стомодеума) и энтодермального (со стороны кишки) листков (см. рис. 7-26). Эта пластинка прорывается в конце 4-й недели развития, и ротовая полость соединяется с первоначально слепо замкнутой кишечной трубкой.
Околоушные слюнные железы развиваются из эктодермы стомодеу-ма, а подъязычные и подчелюстные слюнные железы - из энтодермы вентральной части глотки.
У 5-недельного зародыша ротовое отверстие ограничено следующими структурами (рис. 7.29, а): по средней линии выше ротовой полости находится округлая нависающая область - лобный (фронтальный) выступ; по бокам от него располагаются подковообразные возвышения, окружающие обонятельные ямки. Средние отростки этих возвышений
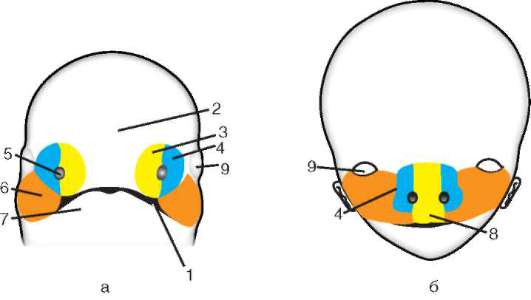
Рис. 7.29. Последовательные этапы формирования лица (вид спереди): а - на 5-й неделе; б - на 10-й неделе. 1 - стомодеум; 2 - лобный (фронтальный) выступ; 3 - медиальный носовой отросток; 4 - латеральный носовой отросток; 5 - обонятельная ямка; 6 - верхнечелюстной отросток; 7 - закладка нижнечелюстной дуги; 8 - фильтрум; 9 - глаз
называются медиальными носовыми, а боковые - латеральными
носовыми отростками. Из верхних боковых углов ротовой полости по направлению к средней линии растут верхнечелюстные отростки.
Ограничивающая ротовую полость снизу нижняя челюсть формируется из парных закладок нижнечелюстной дуги. Вначале они появляются с каждой стороны от средней линии, затем увеличиваются навстречу друг другу и срастаются.
В течение 6-й недели происходит быстрое развитие верхней челюсти. Верхнечелюстные отростки растут по направлению к средней линии. Одновременно с этим носовые отростки увеличиваются, оттесняя наверх расположенную между ними нижнюю часть лобного выступа. Особенно интенсивно растут медиальные носовые отростки, которые смещаются по направлению к средней линии и сливаются друг с другом, а затем срастаются с верхнечелюстными отростками, образуя тем самым на 10-й неделе развития полную верхнечелюстную дугу. Кожная эктодерма над слившимися медиальными отростками формирует фильтрум - продольный желобок над верхней губой под носом (рис. 7.29, б).
К концу второго месяца развития, когда образуется верхняя челюсть, начинает развиваться твердое нёбо. В его формировании, так же как и в образовании верхней челюстной дуги, участвуют средние носовые и верхнечелюстные отростки (рис. 7.30, а). Из срединной области верхней челюсти, возникшей в результате срастания средних носовых отростков, формируется небольшая треугольной формы часть - первичное нёбо. Основная часть - вторичное нёбо - развивается из латеральных нёбных выростов (отростков), которые появляются на верхней челюсти (рис. 7.30, б). Сначала они направлены вниз, а затем края нёбных отростков поднимаются вверх и к средней линии. Они контактируют и сливаются с образованием шва нёба. Сформировавшаяся основная часть нёба разделяет наиболее верхнюю часть исходной стомодеальной полости. Вторичное нёбо срастается с первичным, и к их краниальной поверхности прирастает носовая перегородка (рис. 7.30, в). Таким образом, одновременно с отделением носовой полости от ротовой происходит ее разделение на правую и левую половины.
В начальный период своего формирования (на 5-й неделе эмбриогенеза) язык представляет собой мешок, образованный слизистой оболочкой, заполняющийся растущей мышечной массой. Считают, что слизистая оболочка языка возникает из эктодермы стомодеума, а мышцы происходят из затылочных миотомов.
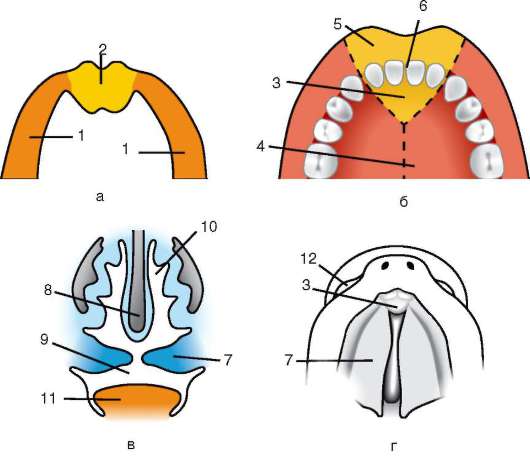
Рис. 7.30. Формирование твердого неба: а - схематичное изображение верхней челюсти (вид со стороны ротовой полости); б - 10-я неделя развития (вид со стороны ротовой полости); в - 8-я неделя развития (фронтальный срез); г - 8-я неделя развития (вид со стороны ротовой полости): 1 - верхнечелюстной отросток; 2 - слившиеся медиальные носовые отростки; 3 - первичное небо; 4 - вторичное небо (латеральные небные выросты); 5 - фильтрум губы; 6 - верхняя челюсть с резцами; 7 - латеральный небный вырост; 8 - носовая перегородка; 9 - ротовая полость; 10 - носовая полость; 11 - язык; 12 - глаз
В формировании слизистой оболочки языка участвует несколько структур дна ротовой полости. Тело языка образуется при срастании парных латеральных утолщений - латеральных язычных бугорков и расположенного между ними небольшого срединного возвышения, называемого tuberculum impar (рис. 7.31). Считают, что тело языка является производным клеточного материала области первой висцеральной дуги. В формировании корня языка принимает участие сорulа (скоба) - срединное возвышение, расположенное позади tuberculum impar и вклю-
Рис. 7.31. Развитие языка: а - 5-недельный зародыш; б - 5-месячный плод; 1 - латеральный язычный бугорок; 2 - tuberculum impar 3 - copula (скоба); 4 - foramen coecum; 5 - терминальная борозда; 6 - тело языка; 7 - корень языка; 8 - закладка надгортанного хряща; 9 - голосовая щель; 10 - черпаловидный бугорок; 11 - надгортанник, нёбная миндалина. I-IV - номера висцеральных дуг
чающее клеточный материал областей второй, третьей и четвертой висцеральных дуг. После срастания всех закладок и окончательного формирования языка его тело отделено от корня терминальной бороздой (sulcus terminalis), в центре которой находится слепое отверстие (foramen coecum) - след впячивания дна глотки, из которого возникает закладка щитовидной железы (см. рис. 7.27). Именно оно считается границей между телом и корнем языка.
Позднее язык почти полностью отделяется от дна ротовой полости, и связь между ними сохраняется в виде складки слизистой оболочки - уздечки языка.
Интересный пример органогенеза - формирование зубов, в котором участвуют покровная эктодерма - эпителий стомодеума (эмаль) и ме-зенхимные клетки, происходящие из нервного гребня (дентин, цемент и пульпа) (рис. 7.32). В ходе развития между мезенхимными и эпителиальными структурами отмечаются индукционные взаимодействия.
В течение жизни развивается два поколения зубов. Смена зубов происходит в детстве, первые зубы называют молочными. Их насчитывается 20: по 10 в верхней и в нижней челюсти. Молочные зубы полностью прорезываются приблизительно в возрасте 2 лет. Этот набор зубов служит ребенку последующие 4 года, после чего молочные зубы постепен-
Рис. 7.32. Постоянные зубы человека: 1 - резцы; 2 - клыки; 3 - малые коренные зубы; 4 - большие коренные зубы
но выпадают и замещаются постоянными зубами (второе поколение зубов), которые функционируют у человека в оставшуюся часть жизни. Период смены зубов продолжается приблизительно от 6 до 12 лет. Постоянных зубов 32: 16 верхних и 16 нижних (добавляются большие коренные зубы) (см. рис. 7.32). По форме они сходны с молочными, но имеют более крупные размеры.
Закладка и образование зубных зачатков молочных зубов начинаются с 6-7 нед внутриутробного развития. Эпителий ротовой полости погружается в подлежащую мезенхиму в виде плотного тяжа, называемого зубной пластинкой (рис. 7.33). На зубной пластинке появляются мелкие эпителиальные выпячивания - зубные зачатки, из которых будут развиваться молочные зубы. По мере роста зубной пластинки каждый зубной зачаток увеличивается в размере, глубже внедряется в мезенхиму и принимает форму перевернутой чаши. Эта структура образует эмалевый орган, а нижележащая мезенхима, заполняющая полость чаши, формирует зубной сосочек. В дальнейшем развитии последний образует пульпу зуба. На 3-м месяце внутриутробного развития эмалевый орган увеличивается в размерах, изменяет форму и постепенно отделяется от зубной пластинки. Форму будущего зуба определяет мезенхимный компонент. При сочетании in vitro мезенхимы закладки коренного зуба (моляра) с эпителиальным компонентом закладки резца, развивается моляр. И наоборот, в результате комбинации эктодермы моляра и мезенхимы резца развивается резец.
В ходе гистогенеза тканей зуба, который начинается с конца 4-го месяца, образуются дентин, эмаль и пульпа (рис. 7.33, г). Эмаль
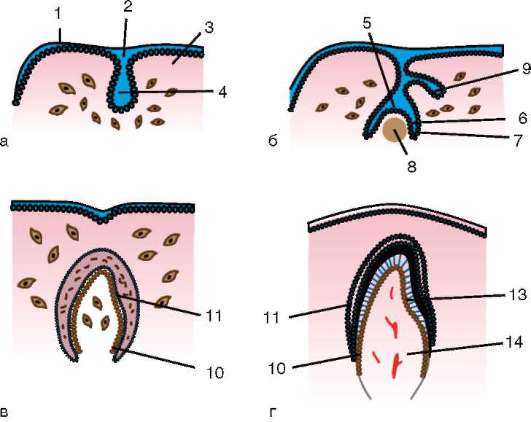
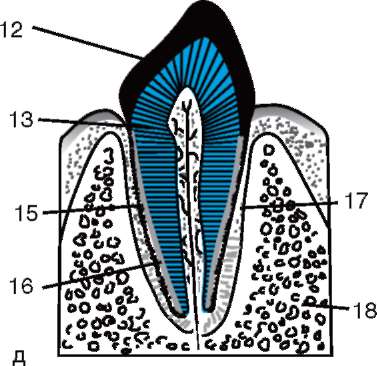
Рис. 7.33. Развитие зуба: а - 8-недельный зародыш; б - 10-недельный плод; в - 3-месячный плод; г - 6-месячный плод; д - после прорезывания; 1 - эпителий ротовой полости; 2 - зубная пластинка; 3 - мезенхима; 4 - зачаток молочного зуба; 5 - эмалевый орган; 6 - внутренний слой эмалевого органа; 7 - наружный слой эмалевого органа; 8 - зубной сосочек; 9 - зачаток постоянного зуба; 10 - одонтобласты; 11 - амелобласты; 12 - эмаль; 13 - дентин; 14 - пульпа зуба; 15 - цементобласты; 16 - цемент; 17 - периодонт; 18 - кость альвеолы
продуцируют клетки внутреннего слоя эмалевого органа - энамело-бласты. Их отростки выделяют органическую основу эмали - эмалевые призмы, которые затем обызвествляются. Источник развития дентина - одонтобласты (дентинобласты), поверхностные клетки пульпы. Верхушка дентинобластов также имеет отростки, выделяющие органические вещества фибриллярной структуры, называемые предентин и образующие матрицу дентина. С конца 5-го месяца в предентине откладываются соли кальция и фосфора, формируется окончательный дентин. Образование дентина и эмали отличается от развития кости тем, что клетки не замуровываются в межклеточное вещество, а отодвигаются: амелобласты - наружу, одонтобласты - внутрь. Кроме того различается содержание органических веществ в этих тканях: кость содержит примерно 45% органики, дентин - 28%, а эмаль - меньше 5%.
На наружной поверхности дентина из окружающей зуб мезенхимы (зубного мешочка) незадолго до прорезывания зуба появляются це-ментобласты. Эти клетки выделяют коллагеновые волокна и межклеточное вещество, образуя цемент. Он формируется только через 4-5 мес после рождения при развитии корней зубов. Из наружного слоя зубного мешочка формируется зубная связка (периодонт) (рис. 7.33, д). Таким образом, во внутриутробном развитии происходит развитие только коронок молочных зубов.
Прорезывание зуба обеспечивается тремя факторами. Первый и важнейший - рост и развитие корня, приводящие к выталкиванию коронки через слизистую оболочку. Второй фактор - рост пульпы, приводящий к повышению давления внутри зубного зачатка. Кроме того, на дне зубной альвеолы происходит дополнительное отложение костной ткани слоями.
Закладка постоянных зубов начинается очень рано, в начале 5-го месяца внутриутробного развития. Зачаток постоянного зуба находится позади зачатка молочного, и развивающийся корень последнего в первые два года после рождения оказывает индукционное влияние на развитие зачатка постоянного зуба. Гистологические процессы, в результате которых формируются молочные и постоянные зубы, одинаковы. Когда прорезываются молочные зубы, в зачатках постоянных происходит образование эмали и дентина. Некоторое время они пребывают в латентном состоянии и активируются только после достижения челюстью достаточно больших размеров. В процессе замены рост постоянного зуба и давление его эмали на корень молочного приводят к рассасы-
ванию остеокластами (клетками-разрушителями костной ткани) более мягкой ткани - дентина молочного зуба. Последний выталкивается и заменяется постоянным.
Развитие отделов пищеварительной системы сложно рассмотреть в деталях, так как кишечная трубка сильно удлиняется, дифференцируется и связана в своем развитии со многими другими системами: эндокринной, дыхательной, выделительной, кровеносной, нервной и вторичной полостью - целомом. Рассмотрим только наиболее ранние и общие процессы, происходящие в кишечной трубке, а также изменения ее связи с желточным мешком.
Кишечная трубка возникает из энтодермы крыши желточного мешка и прилегающего к ней висцерального листка мезодермы в процессе обособления тела эмбриона от внезародышевых частей с помощью головной, хвостовой и боковых туловищных складок (см. рис. 7.24, б2, б3). На 4-й неделе она представлена сравнительно простой трубкой, состоящей из передней кишки, слепо замкнутой спереди стомодеальной (ротовой) пластинкой, средней кишки, связанной с желточным мешком
посредством желточного стебелька (протока), и задней кишки, слепо замкнутой на хвостовом конце клоакальной (анальной) мембраной
(рис. 7.34).
За период от 4-й до конца 8-й недели эмбрионального (внутриутробного) развития кишка удлиняется и начинает дифференцироваться. Из переднего ее отдела формируются глотка, пищевод, желудок и двенадцатиперстная кишка с ее производными (печень и поджелудочная железа) до впадения в нее общего желчного протока.
Средняя кишка образует U-об-разную петлю, которая связана с желточным мешком посредством желточного протока (рис. 7.35, а). Участок кишки между желточным стебельком и желудком превратится в тонкую кишку, а участок, ле-
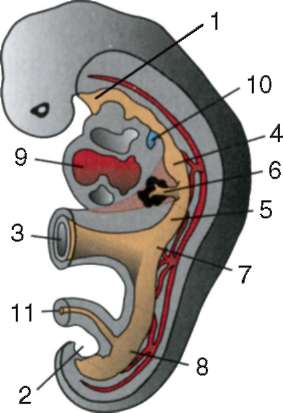
Рис. 7.34. Пищеварительная система 4-недельного зародыша: 1 - стомо-деальная пластинка; 2 - клоакальная мембрана; 3 - желточный проток; 4 - желудок; 5 - двенадцатиперстная кишка; 6 - зачаток печени; 7 - средняя кишка; 8 - задняя кишка; 9 - сердце; 10 - закладка органов дыхания; 11 - урахус
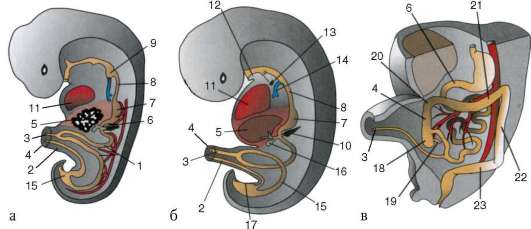
Рис. 7.35. Развитие пищеварительной системы человека: а - на 32-е сутки; б - на 36-е сутки; в - более поздние стадии; 1 - U-образная петля кишки; 2 - урахус; 3 - желточный проток; 4 - брюшной стебелек (пупочный канатик); 5 - печень; 6 - двенадцатиперстная кишка; 7 - желудок; 8 - пищевод; 9 - гортань; 10 - поджелудочная железа; 11 - сердце; 12 - язык; 13 - закладка щитовидной железы; 14 - зачаток трахеи и легких; 15 - задняя кишка; 16 - желчный пузырь; 17 - клоакальная мембрана; 18 - слепая кишка; 19 - аппендикс; 20-23 - отделы толстой кишки
жащий каудальнее желточного протока, - в толстую кишку. Средняя кишка удлиняется, часть ее выпячивается в полость брюшного стебелька, образуя грыжеподобное выпячивание брюшной стенки зародыша.
За это же время некоторые участки кишки, вращаясь, изменяют свое положение (рис. 7.35, б). К 10-й неделе выступающая часть кишечной петли втягивается назад через пупочный ободок, занимая свое окончательное положение в брюшной полости. Несколько ранее в месте перехода тонкой кишки в толстую развивается слепая кишка. Сначала она увеличивается в размере, но к 3-му месяцу ее дистальная часть начинает отставать от роста других частей (неравномерность роста), в результате чего диаметр этой части оказывается значительно меньшим и образуется червеобразный отросток. Из задней кишки формируется толстая кишка ниже селезеночного угла, в том числе прямая (рис. 7.35, в). Развитие клоакального конца задней кишки тесно связано с развитием мочеполового отверстия (рис. 7.36). Уроректальная перегородка, разделяющая клоаку на мочеполовой синус и прямую кишку, образуется на 5-6-й неделе. Прорыв анальной мембраны происходит на 8-й неделе.
На 2-м месяце внутриутробного развития начинается быстрая пролиферация эпителия пищевода и двенадцатиперстной кишки, что
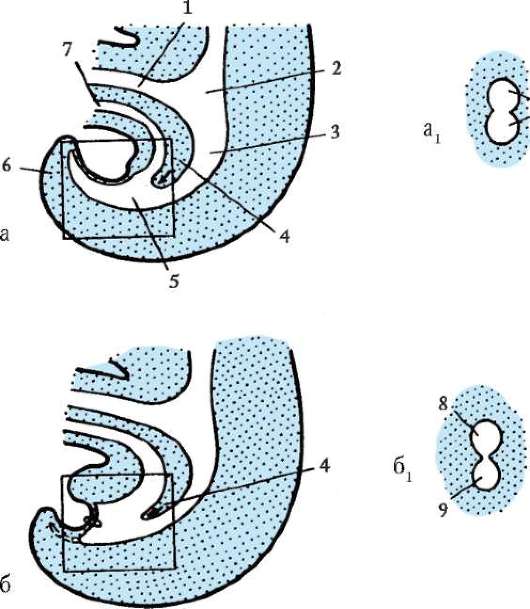
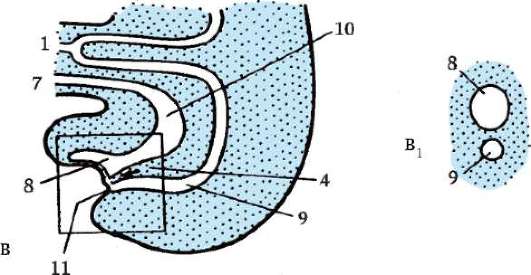
Рис. 7.36. Развитие клоакальной области задней кишки у зародыша человека: а - на 4-й неделе; б - на 6-й неделе; в - на 7-й неделе; а1 б1 и в1 - поперечные срезы через клоаку (уровни среза обозначены квадратами на рис. а; б; в). 1 - желточный стебелек; 2 - средняя кишка; 3 - задняя кишка; 4 - мочеполовая перегородка; 5 - клоака; 6 - хвост; 7 - аллантоис; 8 - мочеполовой синус; 9 - прямая кишка; 10 - мочевой пузырь; 11 - анальная мембрана; стрелками показано направление роста
приводит к временному закрытию их просвета. К концу 2-го месяца происходит реканализация названных органов, т.е. восстановление проходимости.
Сердце человека начинает развиваться с конца 3-й - начала 4-й недели из спланхномезодермы в виде парных зачатков, расположенных под глоткой (рис. 7.37). По мере отграничения тела самого зародыша туловищными складками и замыкания передней кишки с брюшной стороны парные эндокардиальные трубки смыкаются в одну, лежащую по средней линии (см. рис. 7.37). Для земноводных доказано существование индукционного действия энтодермы на прекардиальную мезодерму. Об этом же свидетельствует опыт на курином зародыше, когда после удаления дна передней кишки был получен зародыш с двойным сердцем. Парные закладки целомических полостей сливаются на брюшной стороне, образуя перикардиальную полость.
Региональная дифференцировка сердца начинается с быстрого удлинения первичной сердечной трубки, что приводит к ее изгибу и приобретению 5-образной формы (рис. 7.38). Отмечено, что образование изгиба с локальными изменениями формы клеток вдоль сердечной трубки стимулируется из самой сердечной трубки. Будучи эксплантиро-вана (ситуация «вне организма», или ex vivo), сердечная трубка сохраняет способность к образованию 5-образной структуры.
Рис. 7.37. Развитие сердца человека на 21-22-е сутки: а - общий вид зародыша; б - продольный срез головного конуса зародыша; в - поперечный срез зародыша: 1 - уровень среза; 2 - нервный валик; 3 - сомиты; 4 - первичная полоска; 5 - мозг; 6 - передняя кишка; 7 - сердечная трубка; 8 - перикард; 9 - амнион; 10 - ротоглоточная мембрана; 11 - нервный желобок; 12 - хорда; 13 - парные закладки спинной аорты; 14 - сливающиеся сердечные трубки; 15 - желточный мешок
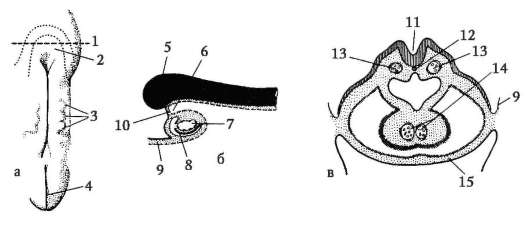
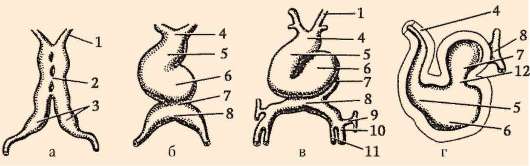
Рис. 7.38. Развивающееся сердце человека: а - на 21-22-е сутки; б - на 23-е сутки; в - на 24-е сутки (вид с брюшной стороны); г - на 28-е сутки (продольный срез): 1 - первая пара дуг аорты; 2 - сливающиеся сердечные трубки; 3 - несливающиеся сердечные трубки; 4 - артериальный ствол; 5 - луковица сердца; 6 - желудочек; 7 - предсердие; 8 - венозный синус; 9 - кювьеров проток; 10 - пупочная вена; 11 - желточная вена; 12 - предсердно-желудочковый канал
На первоначально хвостовом конце находится венозный синус, в который впадают крупные вены: кювьеровы протоки, пупочные и желточные. Венозный синус позже войдет в состав правого предсердия. Краниальнее венозного синуса из расширенной части сердечной трубки образуется предсердие, а из изогнутой средней части - желудочек. Переходную область, где желудочек сужается, называют конусом или луковицей. Позже он будет включен в стенку правого желудочка. Конус переходит в артериальный ствол, от которого отходят корни брюшной аорты.
После образования предсердия и желудочка проявляются внешние признаки предстоящего разделения сердца на правую и левую половины, появляется срединная борозда. В этот момент в тканях формирующегося сердца отмечаются очаговая гибель клеток и взаимодействие внеклеточных и клеточных элементов факторов. Некоторое время спустя на внутренней поверхности желудочка, соответственно уровню наружной борозды, образуется перегородка из мышечных тяжей, растущих от верхушки сердца по направлению к предсердию. На дорсальной и вентральной стенках суженного предсердно-желудочкового канала из рыхлой мезенхимы образуются эндокардиальные подушки. Позднее они трансформируются в плотную соединительную ткань, срастаются и разделяют канал на правый и левый протоки. В то же время появляется первичная срединная перегородка, которая позднее заменяется вторичной. В ней имеется отверстие, называемое овальным, через которое
кровь из правого предсердия попадает в левое. Это необходимо для кровообращения плода.
Одновременно с изменениями в основной части сердца как самостоятельного органа происходит разделение артериального ствола на два канала. Этот процесс начинается в корне брюшной аорты между четвертой и шестой висцеральными дугами. Разделение происходит за счет формирования продольных складок. Складки располагаются по спирали, растут внутрь артериального ствола и, встречаясь, делят его на аорту и легочный ствол. Кроме того, складки распространяются в сторону конуса, где из специализированных участков образуются полулунные клапаны аорты и легочного ствола, и далее в желудочки, где встречаются с перемещающейся эндокардиальной тканью предсердно-желудочкового канала и межпредсердной перегородки. Это приводит к полному зарастанию межжелудочкового отверстия.
До конца внутриутробной жизни остаются открытыми только клапан в овальном отверстии межпредсердной перегородки и артериальный (боталлов) проток, соединяющий легочный ствол с аортой. О нем подробнее будет сказано ниже. Артериальный проток служит для отведения части крови из правого желудочка в аорту, минуя легкие, пока они не достигли необходимого развития и не получили функционального стимула в связи с началом дыхательной функции (рис. 7.39).
Крупные артерии развиваются в комплексе с сердцем, начиная с 4-й недели. Первичная система кровообращения эмбриона функционирует с конца 5-й недели, к концу 8-й недели реализуется основной план строения артериальной системы. Закладывающиеся на 4-й неделе парные висцеральные дуги получают соответствующее артериальное обеспечение в виде аортальных дуг. Эти дуги аорты поднимаются от артериального ствола, а точнее, от его расширенной части - артериального мешка (называемого также брюшной или восходящей аортой) - и заканчиваются в корнях спинной, или нисходящей, аорты (см. рис. 7.27).
Несмотря на то что закладывается шесть пар аортальных (висцеральных) дуг, они не сосуществуют одновременно. В тот период, когда формируется шестая пара дуг, две первые пары уже дегенерируют. Пятая дуга появляется лишь на короткое время в виде рудиментарного сосуда.
В формировании магистральных сосудов существенную роль играют корни брюшной и спинной аорты, а также третья, четвертая и шестая пары аортальных дуг (рис. 7.40).
Проксимальные части третьей пары образуют общие сонные артерии (рис. 7.41). Левая четвертая дуга составляет часть дуги аорты.
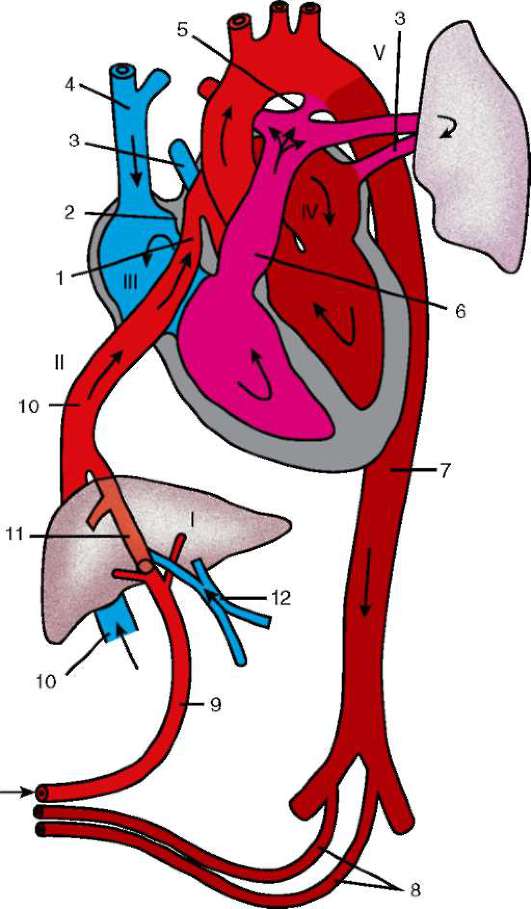
Рис. 7.39. Схема кровообращения плода: 1 - овальное окно; 2 - межпредсерд-ная перегородка; 3 - легочная вена; 4 - верхняя полая вена; 5 - артериальный (боталлов) проток; 6 - легочная артерия; 7 - нисходящая аорта; 8 - пупочные артерии; 9 - пупочная вена; 10 - нижняя полая вена; 11 - ductus venosus; 12 - портальная вена печени. I - печень; II - нижняя полая вена; III - правое предсердие; IV - левое предсердие; V - нисходящая аорта
Рис. 7.40. Развитие аортальных дуг: а - вид с брюшной стороны: 1-6 - аортальные дуги; 7 - легочная артерия; 8 - артериальный мешок, брюшная аорта; 9 - седьмая межсегментарная артерия, подключичная; 10 - спинная нисходящая аорта; 11 - артериальный ствол; 12 - наружная сонная артерия, корень брюшной восходящей аорты; 13 - внутренняя сонная артерия, корень спинной аорты
Правая четвертая дуга становится проксимальной частью правой подключичной артерии. Дистальная часть этой артерии образуется из правой спинной аорты. Проксимальная часть левой шестой дуги превращается в проксимальную часть левой легочной артерии, а дистальная часть - в артериальный (боталлов) проток. Проксимальная часть правой шестой дуги образует проксимальную часть правой легочной артерии, а дистальная часть редуцируется. Корни спинной аорты очень рано сливаются в непарную спинную аорту, но на 7-й неделе развития дистальная часть правого корня спинной аорты обычно включается в правую подключичную артерию, о чем уже было сказано выше. Это становится возможным благодаря обратному развитию участка, соединяющего правый корень с левым. Изложение материала, касающегося развития сердца и крупных артерий, дает основания для сопоставле-
Рис. 7.41. Формирование крупнейших артерий, производных артериального ствола, аортального мешка, аортальных дуг и спинной аорты: а - 6-недельный зародыш; б - 7-недельный зародыш; в - 6-месячный плод: 1-6 - аортальные дуги; 7 - артериальный ствол; 8 - легочная артерия; 9 - подключичная артерия; 10 - спинная нисходящая аорта; 11 - аортальный мешок; 12 - внутренняя сонная артерия; 13 - артериальный (боталлов) проток; 14 - восходящая аорта; 15 - легочный ствол; 1б - наружная сонная артерия; 17 - общая сонная артерия
ний онтогенеза человека с филогенетическим развитием позвоночных (см. гл. 14), а также позволяет приблизиться к пониманию механизмов возникновения врожденных пороков развития сердца и сосудов.
Вопросы для самоконтроля
1. Какие типы онтогенеза существуют? Перечислите и охарактеризуйте основные этапы онтогенеза.
2. Каковы особенности строения и функций половых клеток?
3. Как осуществляется оплодотворение, и какие этапы включает этот процесс?
4. Охарактеризуйте сущность и морфологические формы стадий эмбрионального развития у представителей разных классов
хордовых (дробления, гаструляции, органогенеза и формирования профизорных органов).
5. Каковы особенности эмбрионального развития млекопитающих?
6. Приведите примеры эмбрионального развития органов человека, отражающего эволюцию предковых групп хордовых животных.