Эндокринная регуляция. Биохимические и физиологические аспекты : учеб. пособие / А.Н. Смирнов ; под ред. В.А. Ткачука - 2009. - 368 с.
|
|
|
|
ВВЕДЕНИЕ
Процессы роста, дифференцировки, размножения, адаптации многоклеточного организма происходят при обязательном обмене информацией между клетками. Сигнальными свойствами обладают многие широко представленные в организме утилизируемые соединения, такие, как глюкоза, жирные кислоты, аминокислоты, кальций и т.д. Этот способ общения между клетками при всей своей значимости не покрывает, однако, всех потребностей целостного организма в координации функций разных клеток, органов и систем. Роль важнейших координаторов в ходе эволюции была возложена на специализированные на сигнальной функции секретируемые соединения - системно действующие гормоны и местно действующие гистогормоны, в том числе нейромедиаторы. Общение соседних клеток может осуществляться и другими способами, включая действие сигнальных соединений, заякоренных на плазматической мембране (в том числе молекул адгезии), и непосредственный обмен низкомолекулярными несекретируемыми сигнальными соединениями (вторыми посредниками) через так называемые щелевые контакты. Для перечисленных способов взаимодействия между клетками характерна одна общая черта: все они осуществляются с участием специализированных на проведении сигнала сенсоров/рецепторов сигнальных соединений (рис. 0-1).
Термины «рецептор» и «сенсор» близки, но не идентичны. Как правило, рецептором называют белок или белковый комплекс, предназначенный для восприятия действия специализированного на сигнальной функции соединения, такого, как гормон. Под сенсором же обычно понимают белок или белковый комплекс, воспринимающий сигнал обычного метаболита или его минорного производного. При этом по своей структуре сенсор может относиться к одному из классов известных рецепторов или представлять собой фермент или белок с иной активностью с аллостерической регуляцией. Как правило, рецепторы обладают более высокими сродством и избирательностью к своим лигандам, чем сенсоры.
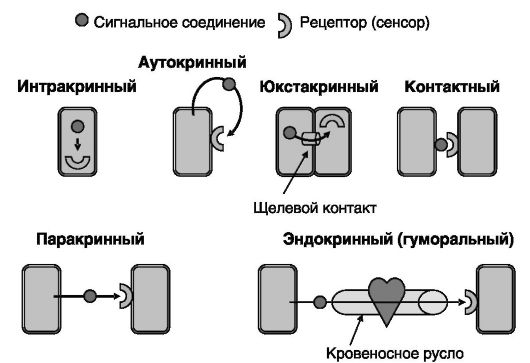 Рис. 0-1. Способы доставки/действия сигнальных соединений
Рис. 0-1. Способы доставки/действия сигнальных соединений
Одно и то же сигнальное соединение может действовать одновременно несколькими способами. Например, эстрогены - типичные дистантно действующие гормоны - в фолликулах яичников могут выполнять функцию паракринных регуляторов, стимулирующих развитие яйцеклетки, и интракринных регуляторов, повышающих продукцию клетками гранулезы ингибина (рис. 0-2).
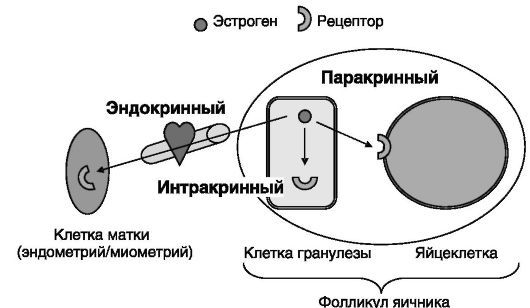 Рис. 0-2. Способы доставки/действия эстрогенов
Рис. 0-2. Способы доставки/действия эстрогенов
Концентрация сигнального соединения и его сродство к рецепто- ру/сенсору взаимно адаптированы, т.е. чем выше уровень сигнального соединения, тем ниже его сродство к рецептору. Примером природного эксперимента в этом плане могут служить приматы Нового Света. Обнаруженная у них повышенная (по сравнению с приматами Старого Света) концентрация стероидных гормонов, очевидно, является адаптацией к низкой аффинности рецепторов к соответствующим лигандам (из-за высокой ингибирующей активности одного из шаперонных белков, FKBP51). Иллюстрацией взаимной адаптации аффинности к рецептору и концентрации сигнального соединения может также служить сопоставление равновесных констант диссоциации кальция с его сенсором (CaR) и эстрадиола с его рецептором (Э-Рц), составляющих величины порядка 10-3 М и 10-10 М соответственно, с концентрациями этих сигнальных соединений в крови, имеющими такие же порядки величин.
Способ доставки/действия сигнального соединения зависит и от аффинности рецептора, и от соотношения скоростей продукции и разрушения сигнального соединения в организме. Многие дистантно действующие сигнальные соединения (гормоны) отличаются относительной устойчивостью к действию метаболизирующих их ферментов. Если деградация сигнального соединения все же происходит быстро, то для дистантной доставки необходима компенсация в форме высокой интенсивности его секреции. Способ доставки/действия сигнального соединения может меняться в зависимости от условий. Так, при сильных воспалительных реакциях продукция цитокинов, действующих обычно локально, может возрастать настолько, что они приобретают способность достигать отдаленных тканей и органов (рис. 0-3).
Местно действующие сигнальные соединения нередко выступают в качестве средств распространения влияния системных регуляторов на клетки, не содержащие соответствующие рецепторы. Например, стимуляция прогестероном ветвления протоков и альвеологенеза в молочной железе опосредуется небольшой группой чувствительных к прогестерону стромальных и эпителиальных клеток, которые в ответ на поступление прогестерона секретируют паракринные регуляторы херегулин, Wnt-4 и RANKL, индуцирующие пролиферацию соседних, нечувствительных к прогестерону эпителиальных клеток.
Сигнальными свойствами обладают соединения разной химической природы: липиды, аминокислоты и их производные, нуклеотиды, пептиды и белки (рис. 0-4).
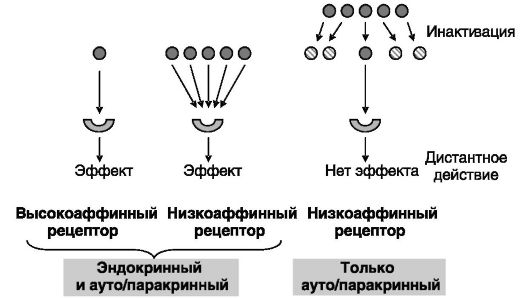 Рис. 0-3. Способ
доставки/действия сигнального соединения зависит от: 1) интенсивности
его продукции; 2) сродства к рецептору; 3) скорости деградации
Рис. 0-3. Способ
доставки/действия сигнального соединения зависит от: 1) интенсивности
его продукции; 2) сродства к рецептору; 3) скорости деградации
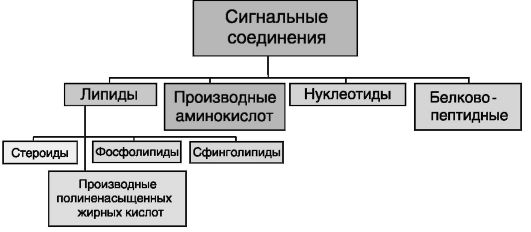 Рис. 0-4. Сигнальными свойствами обладают соединения разной химической природы
Рис. 0-4. Сигнальными свойствами обладают соединения разной химической природы
Ниже приведен перечень соединений, обладающих сигнальными функциями.
Независимо от химической природы в структуре сигнального соединения можно выделить несколько функциональных фрагментов: адресные, определяющие высокоаффинное связывание с рецептором; эффекторные, индуцирующие проведение сигнала рецептором; вспомогательные, влияющие на интенсивность деградации
Таблица 0-1. Соединения, обладающие сигнальными свойствами
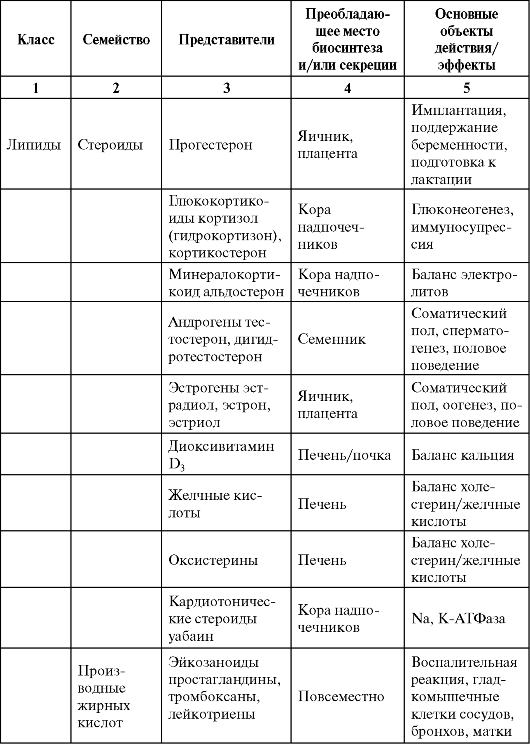 Продолжение табл. 0-1
Продолжение табл. 0-1
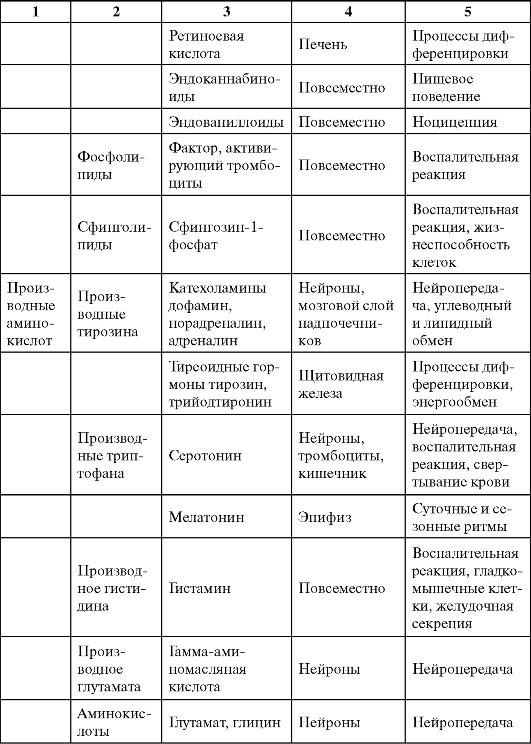 Продолжение табл. 0-1
Продолжение табл. 0-1
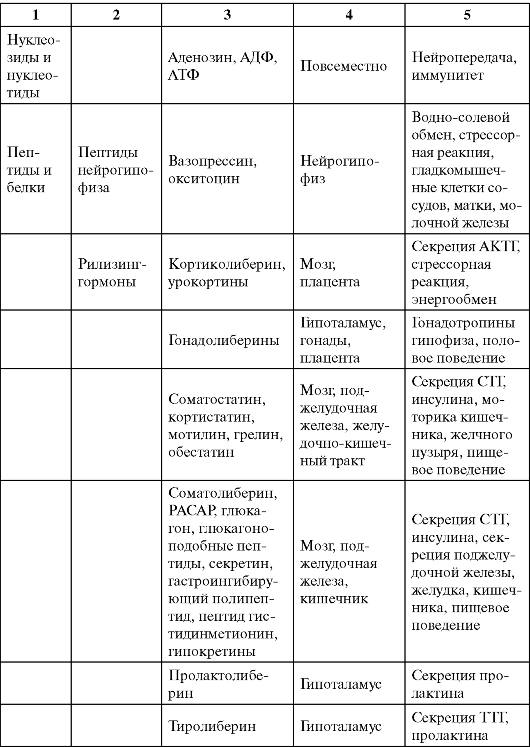 Продолжение табл. 0-1
Продолжение табл. 0-1
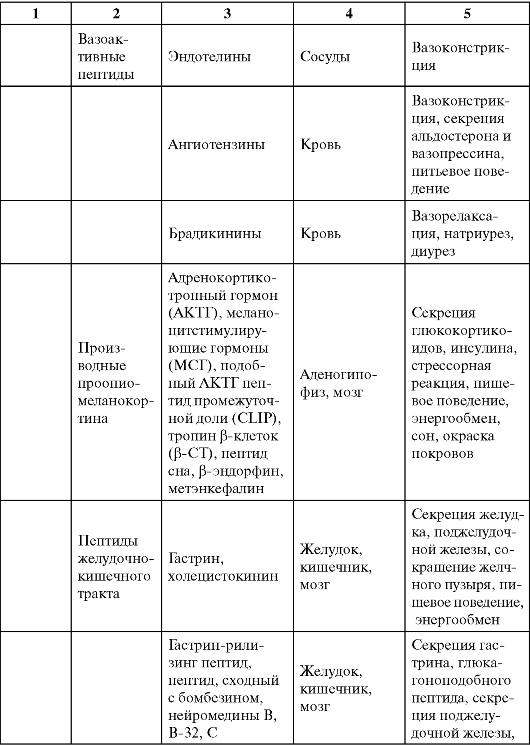 Продолжение табл. 0-1
Продолжение табл. 0-1
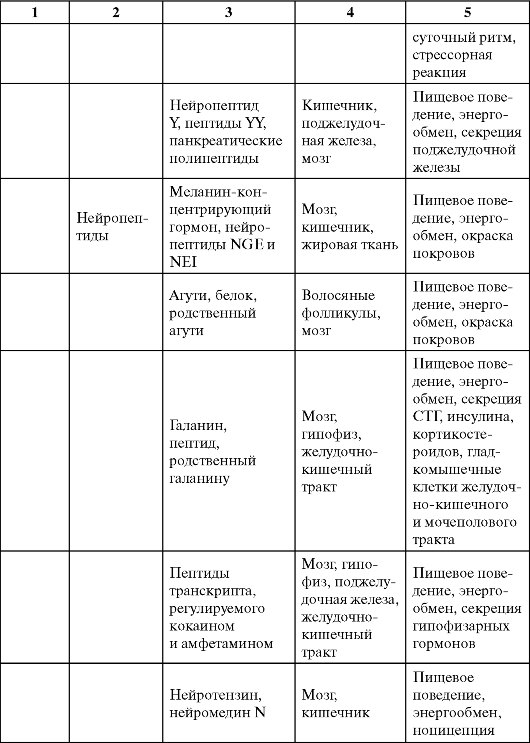 Продолжение табл. 0-1
Продолжение табл. 0-1
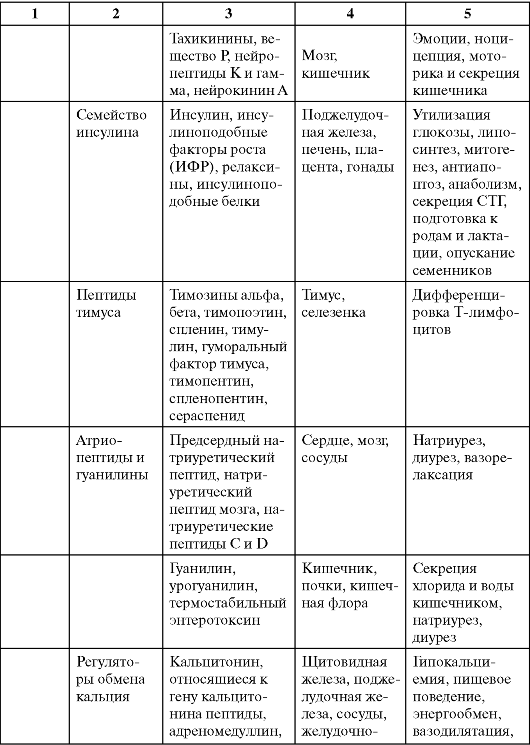 Продолжение табл. 0-1
Продолжение табл. 0-1
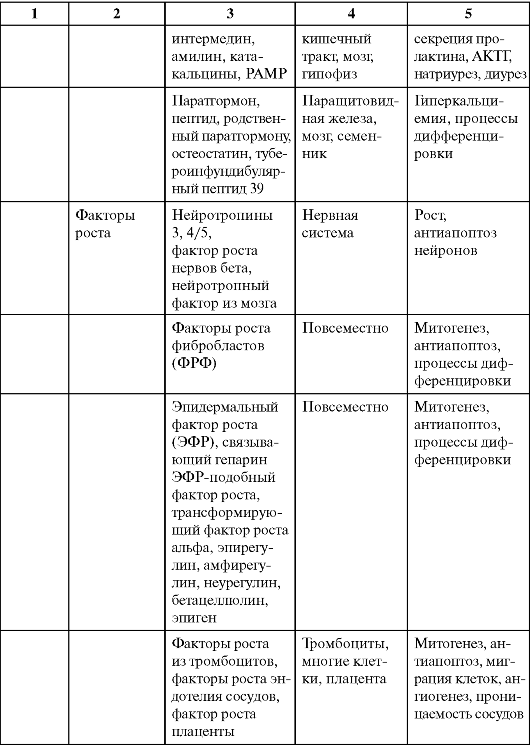 Продолжение табл. 0-1
Продолжение табл. 0-1
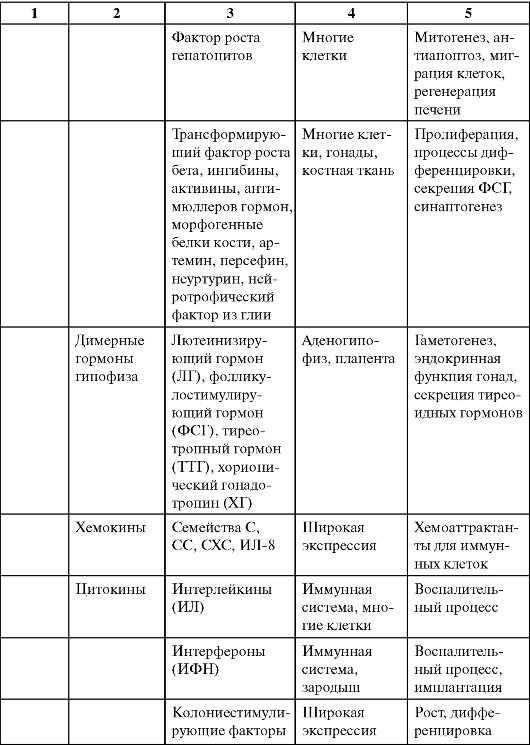 Продолжение табл. 0-1
Продолжение табл. 0-1
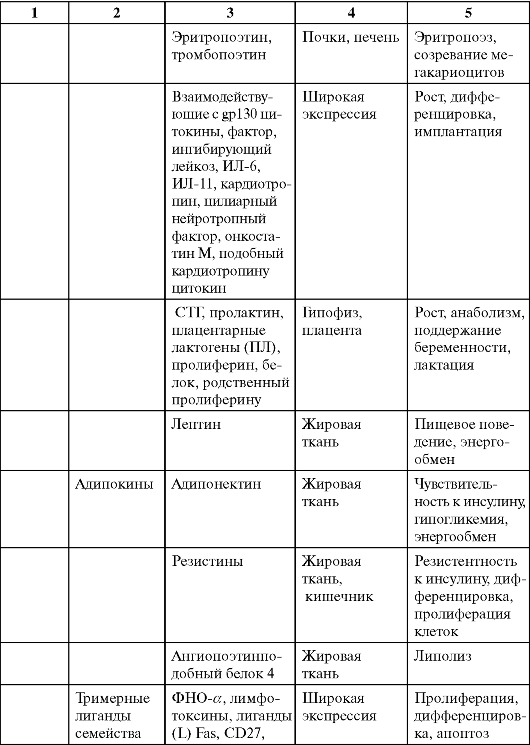 Окончание табл. 0-1
Окончание табл. 0-1
 и
некоторые другие характеристики сигнального соединения. Эти
функциональные фрагменты могут частично перекрываться. В ряде случаев
сигнальное соединение может действовать не через один, а через несколько
рецепторов. Соответственно в структуре такого сигнального соединения
может содержаться несколько адресных и эффекторных фрагментов.
и
некоторые другие характеристики сигнального соединения. Эти
функциональные фрагменты могут частично перекрываться. В ряде случаев
сигнальное соединение может действовать не через один, а через несколько
рецепторов. Соответственно в структуре такого сигнального соединения
может содержаться несколько адресных и эффекторных фрагментов.
Действие естественных регуляторов могут имитировать экзобиотики, т.е. соединения, поступающие извне: в составе пищи (например, изофлавоноиды сои с эстрогенной активностью), в составе продуктов кишечной флоры (например, термостабильный энтеротоксин с активностью гуанилинов) или в результате контакта с животными (например, натриуретический пептид D из змеиного яда). Нередко анализ механизма действия экзобиотиков позволяет выявить новые эндогенные сигнальные соединения. Примером может служить обнаружение группы производных жирных кислот с сигнальными свойствами (эндоканнабиноидов), действующих через те же рецепторы, что и активный ингредиент марихуаны (-)-Δ9-тетрагидроканнабинол. Экзобиотиками являются и лекарственные препараты. Рецепторы сигнальных соединений служат объектом действия более чем половины всех существующих терапевтических агентов.
Деградация сигнальных соединений, уравновешивающая их биосинтез, включает прежде всего модификацию адресных и эффекторных фрагментов. Если метаболические превращения начинаются с эффекторных фрагментов, то в результате могут возникать конкурентные антагонисты, т.е. соединения, оккупирующие рецептор, но не способные его активировать. Этот механизм служит, по-видимому, одним из способов терминации сигнализации. Терапевтический эффект многих фармацевтических агентов (например, бета-адреноблокаторов, антигистаминовых препаратов) основывается на том
же принципе конкурентного ингибирования рецепторов. Один из вариантов ингибирования основывается на необходимости для ряда рецепторов (например, рецептора гормона роста - ГР) формировать димерную структуру вокруг одной молекулы сигнального соединения. Разрушение в этой молекуле одного из двух сайтов взаимодействия с рецептором ведет к блокаде образования димера рецептора и проведения сигнала.
На пути от клетки-продуцента к клетке-мишени сигнальное соединение может вступить в комплекс с транспортным белком. В составе таких комплексов сигнальное соединение, как правило, оказывается временно недоступным ни для рецепторов, ни для разрушающих ферментов. Возникающий резерв сигнального соединения может быть мобилизован при локальном разрушении транспортного белка. Этот механизм, в частности, используют раковые клетки для поддержания своей жизнеспособности и пролиферативного потенциала: они секретируют протеазы, которые гидролизуют белки, связывающие инсулиноподобные факторы роста (ИФР). Высвобождающиеся ИФР оказывают антиапоптотическое и митогенное действие на опухолевые клетки (рис. 0-5, а). Направленная доставка сигнального соединения транспортным белком может также осуществляться за счет наличия на поверхности клетки-мишени рецептора транспортного белка (рис. 0-5, б). Допускается также, что сигнальное соединение может выступать в качестве аллостерического регулятора транспортного белка, выполняющего собственную сигнальную функцию (рис. 0-5, в). Изменение соотношения сигнальное соединение/транспорт- ный белок может служить фактором амплификации сигнализации. Например, значительное повышение секреции глюкокортикоидов в предродовой период на фоне неизменного или снижающегося уровня связывающего эти гормоны транскортина рассматривается в качестве одного из механизмов инициации родовой деятельности (рис. 0-5, г).
Рецепторы сигнальных соединений по механизму проведения сигнала весьма разнообразны (рис. 0-6). Стероидные гормоны, производные холестерина и некоторые производные жирных кислот служат лигандами ядерных рецепторов - надсемейства белков, функционирующих в качестве транскрипционных факторов. Ряд ксенобиотиков и некоторые эндобиотики действуют через рецептор арильных углеводородов - транскрипционный фактор надсемейства, отличного от надсемейства ядерных рецепторов. Электрофильные ксенобиотики и некоторые эндобиотики (а через них и активные формы кислорода)
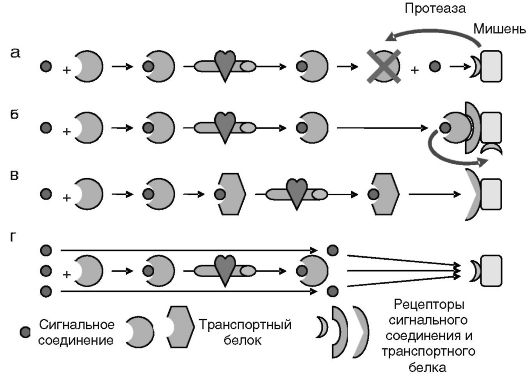 Рис. 0-5. Формы участия транспортных белков в доставке и действии сигнальных соединений
Рис. 0-5. Формы участия транспортных белков в доставке и действии сигнальных соединений
оказывают влияние на клетку посредством ковалентной модификации (присоединения к сульфгидрильным группам) репрессора одного из транскрипционных факторов. Группа лигандов Notch стимулирует протеолитическое расщепление своих рецепторов с высвобождением корегуляторов транскрипционных факторов. Ряд рецепторов обладает встроенной ферментативной функцией. Рецепторные гуанилатциклазы опосредуют эффекты натриуретических пептидов и гуанилинов. Рецепторные тирозинкиназы служат посредниками в действии инсулина и многих ростовых факторов (ИФР, факторов роста фибробластов - ФРФ, лигандов семейства эпидермального фактора роста - ЭФР, лигандов семейства фактора роста из тромбоцитов - PDGF, нейротропинов, фактора роста гепатоцитов - ФРГ и др.). Большинство представителей семейства трансформирующего фактора роста бета (ТФР-β), включая антимюллеров гормон (АМГ), ингибины и активины, действует через рецепторные серин/трео- нинкиназы. Сигналы ряда заякоренных на клеточной мембране лигандов могут восприниматься рецепторными протеинфосфата-
зами. Рецепторы многих сигнальных соединений, не обладая собственной ферментативной активностью, прямо или опосредованно (через адапторные белки) связаны с ферментами. Самую многочисленную группу составляют рецепторы, сопряженные с G-белками. Индуцированная замена в G-белках связанного гуанозиндифосфата (ГДФ) гуанозинтрифосфатом (ГТФ) обеспечивает регуляцию активности многих ферментов и ионных каналов. Через рецепторы этого типа действуют гипоталамические рилизинг-гормоны, нейрогипофизарные гормоны вазопрессин и окситоцин, гипофизарные гормоны семейства фолликулостимулирующего гормона (ФСГ) и производные проопиомеланокортина (адренокортикотропный гормон - АКТГ,
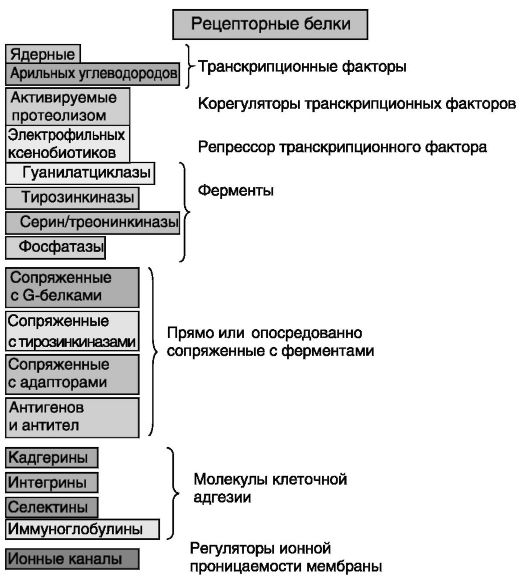 Рис. 0-6. Известные типы рецепторов сигнальных соединений
Рис. 0-6. Известные типы рецепторов сигнальных соединений
меланоцитстимулирующие гормоны - МСГ, бета-эндорфин - β-END и мет-энкефалин - met-ENK), вазоактивные пептиды, многие нейропептиды и гормоны желудочно-кишечного тракта, хемокины, гормональные регуляторы обмена кальция паратгормон, кальцитонин и родственные им полипептиды, релаксин, нейромедиаторы катехоламины и отчасти ацетилхолин, ряд липидов (эйкозаноиды групп простагландинов, тромбоксанов и лейкотриенов, эндоканнабиноиды, фактор активации тромбоцитов, сфингозин-1-фосфат), отчасти нуклеотиды и т.д. Цитокины (интерлейкины, интерфероны, колониестимулирующие факторы, эритропоэтин) и близкие к ним по пространственной организации гормоны пролактин, соматотропный гормон, плацентарный лактоген, лептин действуют через рецепторы, ассоциированные с растворимыми тирозинкиназами. Тримерные лиганды надсемейства фактора некроза опухолей альфа (ФНО-α), также относимые к цитокинам, действуют через рецепторы, либо сами включающие, либо ассоциированные с адапторными белками, содержащими так называемый домен смерти. Рецепторы антигенов и антител включают субъединицы, фосфорилируемые по остаткам тирозина, или ассоциированы с адапторными белками, содержащими подобные аминокислотные мотивы, что напоминает ситуацию с рецепторами цитокинов. Молекулы, выполняющие адгезивные функции, такие, как кадгерины, интегрины, селектины, иммуноглобулиноподобные молекулы клеточной адгезии, способны сообщать клетке о состоявшемся контакте с внеклеточным матриксом или соседней клеткой с использованием внутриклеточных путей сигнализации, включающих элементы, характерные для проведения сигналов другими типами рецепторов, такие, как малые G-белки, тирозин- и серин/ треонинкиназы и др. Ионотропные рецепторы представляют собой самую мобильную систему проведения сигнала и функционируют преимущественно при синаптической передаче действия нейромедиаторов, таких, как ацетилхолин (никотиновые рецепторы), глутамат и др. Ионные каналы могут служить объектами аллостерической регуляции со стороны сигнальных соединений, не являющихся нейромедиаторами, таких, как эндованиллоиды и нейростероиды. Не для всех сигнальных соединений рецепторы идентифицированы (например, «сиротами» остаются резистины, продукты транскрипта, регулируемого кокаином и амфетамином). Клонированы также рецепторы с неизвестным механизмом передачи сигнала (например, рецепторы адипонектина, хотя и напоминают рецепторы, сопряженные с G-бел-
ками наличием 7 трансмембранных доменов, но отличаются от них обратным расположением N- и С-концевых фрагментов относительно плазматической мембраны). С другой стороны, «сиротскими» остаются и многие члены известных надсемейств рецепторов, лиганды которых пока не идентифицированы. Не исключено, что в некоторых случаях таких лигандов просто не существует. Например, заполнение лигандсвязывающего кармана в гомологе 1 рецептора печени (надсемейство ядерных рецепторов) объемными аминокислотными заместителями никак не сказывалось на транскрипционной активности этого белка.
Рецепторы, локализованные на плазматической мембране, способны к быстрому проведению сигналов (миллисекунды для ионных каналов, секунды-минуты для других типов рецепторов) посредством регуляции ионных потоков, образования или высвобождения низкомолекулярных внутриклеточных (вторых) посредников (таких, как циклический аденозинмонофосфат - цАМФ, циклический гуанозинмонофосфат - цГМФ, циклическая аденозиндифосфатрибоза, фосфатидилинозитиды и инозитолфосфаты, церамид и родственные сфинголипиды, кальций), активирующего/инактивирующего фос- форилирования/дефосфорилирования белков. Вызванные изменения могут вести к модуляции структуры хроматина и активности транскрипционных факторов, что находит отражение в долгосрочной (часы-дни) перестройке работы клетки, включая процессы пролиферации и дифференцировки. Допускается также более прямое влияние сигнальных соединений и их мембранных рецепторов на процессы транскрипции за счет их транспорта в клеточное ядро и действия в качестве корегуляторов транскрипционных факторов. Для эффектов сигнальных соединений, действующих через ядерные рецепторы, характерно медленное развитие (часы-дни). Вместе с тем для таких соединений документированы и быстрые эффекты (минуты). В отдельных случаях (например, при индукции эстрогенами вазодилатации через аккумуляцию NO) посредником быстрого эффекта может служить изоформа ядерного рецептора, заякоренная на плазматической мембране. В других случаях (например, при стимуляции прогестероном развития яйцеклетки или акросомной реакции сперматозоида) проведение быстрого сигнала может реализоваться специальными мембранными рецепторами. Таким образом, граница между сигнальными соединениями, действующими через мембранные и ядерные рецепторы, весьма условна.
Клетки-продуценты сигнальных соединений могут специализироваться на синтезе и секреции одного или нескольких близких по структуре или биосинтетическому пути веществ. Например, в клетках промежуточной доли гипофиза из одного полипептидного предшественника проопиомеланокортина образуется несколько пептидов с сигнальными свойствами (меланоцитстимулирующие гормоны - МСГ, пептид промежуточнойдоли, подобныйкортикотропину - CLIP). Часто, однако, одна и та же клетка секретирует одновременно несколько неродственных сигнальных соединений, спектры действия которых могут частично перекрываться. Так, β-клетки поджелудочной железы наряду с гипогликемическим, стимулирующим липосинтез гормоном инсулином в ответ на поступление глюкозы секретируют гормон амилин, который за счет подавления пищевого поведения и повышения энергозатрат снижает жировые запасы, т.е. действует на липидный обмен противоположно инсулину. Практически все клетки в организме способны синтезировать те или иные сигнальные соединения, и эта способность чаще всего совмещается с выполнением клетками других функций (сократительной для гладкомышечных клеток сердца и сосудов, барьерной для эндотелия сосудов и т.д.). Лишь немногие клетки специализируются исключительно на продукции сигнальных соединений. Такие клетки часто формируют скопления (например, островки Лангерганса в поджелудочной железе) или целые железы (например, мозговой и корковый слои надпочечника), но могут быть рассеяны между клетками с иными функциями (например, эндокринные клетки в желудочно-кишечном тракте). Эндокринные железы обеспечивают действие продуцируемых ими сигнальных соединений на системном уровне. Ряд тканей и органов способен дублировать синтез гормональных соединений, но образующиеся здесь под контролем локальных регуляторов сигнальные соединения действуют местно, аутоили паракринно. Например, образующиеся в тимусе глюкокортикоиды обеспечивают тонкую регуляцию селекции развивающихся T-клеток.
Практически любая клетка организма испытывает действие одновременно нескольких сигнальных соединений. В некоторых случаях действие одного из них может быть превалирующим (например, антиапоптотическое, митогенное, анаболическое действие андрогенов на предстательную железу). В таких случаях говорят о гормонзависимости ткани. В других случаях (например, при адаптации к режиму питания) сигнальное соединение лишь корректирует функции клеток, не
затрагивая само их существование и базальную активность. Такие клетки и ткани называют гормончувствительными.
Действие двух сигнальных соединений на одну и ту же функцию клетки характеризуют как антагонистичное, если одно соединение подавляет эффект другого; аддитивное, если эффекты субоптимальных доз суммируются; синергичное, если результирующий эффект превышает аддитивный. За общим феноменологическим описанием в конкретных случаях могут лежать разные молекулярные механизмы. Примеры антагонистичного действия включают активацию сигнальными соединениями стимулирующего и ингибирующего аденилатциклазу G-белков (GS и Gi). Аддитивность наблюдается в действии гипергликемических гормонов на киназу гликогенфосфорилазы: активность фермента возрастает и под влиянием внутриклеточного кальция (через кальмодулин), и под влиянием фосфорилирования (протеинкиназой A - ПК-А). Синергизм может возникать в тех случаях, когда эффект одного сигнального соединения восполняет лимитирующее звено в действии другого сигнального соединения. Таким звеном, в частности, может служить уровень рецепторного белка. В этом случае говорят о сенсибилизирующем эффекте.
Для действия ряда сигнальных соединений могут быть характерны так называемые критические периоды, когда наличие сигнального соединения абсолютно необходимо для нормального развития органов и систем. Например, недостаточность поступления йода с пищей при беременности, сопровождающаяся недостаточностью образования тиреоидных гормонов у матери и плода, ведет к необратимому недоразвитию структур мозга у новорожденных, проявляющемуся в форме кретинизма. В отдельных случаях действие сигнального соединения в критический период проявляется не сразу, а спустя относительно долгое время. Таким образом у грызунов происходит программирование андрогенами семенников (конец внутриутробного развития - первые дни постнатальной жизни), мужского полового поведения, агрессивной реакции, ациклического типа функционирования репродуктивной системы, паттерна секреции гормона роста, проявляющихся после полового созревания. Подобный тип детерминации называют гормональным импринтингом.
Продукция конкретного сигнального соединения клеткой контролируется многими факторами, включая другие сигнальные соединения. Наиболее общим принципом такого контроля является отрицательная обратная связь, т.е. ингибирование контролируемым
сигнальным соединением продукции контролирующего соединения. Эта связь может включать несколько звеньев, расположенных последовательно или параллельно, причем эти звенья могут включать не только сигнальные соединения, но и другие регулируемые параметры, такие, как электролитный состав или давление крови. Например, секреция соматотропного гормона (СТГ) гипофизом ингибируется стимулируемой СТГ секрецией ИФР-I печенью. Это ингибирование включает прямое подавление активности соматотрофов, а также снижение секреции гипоталамусом соматолиберина (гормона роста рилизинг-гормон - ГГРГ) и стимуляцию секреции гипоталамусом соматостатина. Наличие параллельных систем ауторегуляции секреции сигнальных соединений, очевидно, придает этой регуляции повышенную устойчивость. В эмбриональный/плодный период развития формирование систем отрицательной обратной связи может отставать от появления способности к образованию сигнальных соединений, что способствует активному участию этих соединений в органогенезе. Системы положительной обратной связи между сигнальными соединениями распространены в гораздо меньшей степени и действуют на протяжении сравнительно коротких промежутков времени. Наиболее известным примером такого рода связи служит лавинообразное нарастание секреции лютеинизирующего гормона (ЛГ) в предовуляторный период под действием эстрогенов яичников. На протяжении остальной части овариального цикла эстрогены подавляют секрецию ЛГ, который является активатором их продукции.
Многие формы патологии связаны с нарушениями образования или действия сигнальных соединений. Например, к аганглиозу кишечника может приводить недостаточность эндотелина 3, его рецептора, неуртурина или нейротропного фактора из глии. Причиной рака предстательной железы нередко служат мутации рецептора андрогенов (А-Рц), расширяющие спектр взаимодействующих с ним лигандов (например, андрогенную активность начинают проявлять глюкокортикоиды). В ряде опухолей выявлены мутации рецепторных тирозинкиназ, обеспечивающие конститутивную активность этих митогенных ферментов. Геномы вирусов и микроорганизмов могут кодировать пептиды и белки, близкие к сигнальным соединениям или их рецепторам хозяев. Так, термостабильный энтеротоксин кишечной палочки имитирует действие гормонов кишечника гуанилинов, вызывая диарею. Вирусные гомологи рецепторных тирозинкиназ часто конститутивно активны и не контролируются сигнальными соединениями, что может служить причиной злокачественного перерождения клеток-хозяев.