Эндокринная регуляция. Биохимические и физиологические аспекты : учеб. пособие / А.Н. Смирнов ; под ред. В.А. Ткачука - 2009. - 368 с.
|
|
|
|
ГЛАВА 7. ИНТЕГРАЦИЯ ОБМЕННЫХ ПРОЦЕССОВ
Обмен веществ условно подразделяют на следующие типы:
• азотистый обмен (белков, аминокислот, нуклеиновых кислот, нуклеотидов);
• углеводный обмен;
• липидный обмен;
• обмен энергии;
• минеральный обмен.
В пределах каждого из типов обмена различают стадии обмена:
• поступление веществ с пищей, переваривание и всасывание;
• межуточный (тканевый) обмен - переработка веществ и энергии в клетках;
• образование конечных продуктов и их выведение.
Разные типы обмена тесно связаны между собой. Эта связь осуществляется на разных уровнях и в разных формах. Основой для взаимосвязи служат отношения между белками, углеводами и липидами как предшественниками и продуктами взаимопревращений с общим энергообеспечением и общими конечными продуктами обмена.
ВЗАИМОСВЯЗИ БЕЛКОВ, ЛИПИДОВ И УГЛЕВОДОВ
Общее энергетическое обеспечение. В анаболических процессах биосинтеза белков, липидов или углеводов используются одни и те же источники энергии фосфорилирования (АТФ) или восстановления (НАД(Ф)Н), которые пополняются за счет катаболических процессов распада поступающих с пищей или эндогенных белков, липидов или углеводов (рис. 7-1). Между процессами биосинтеза имеется конкуренция за источники энергии.
Общие предшественники и промежуточные продукты обмена. Среди многих общих продуктов и субстратов, образующихся и используемых в процессах обмена белков, липидов и углеводов, можно выделить как наиболее значимые образующиеся при гликолизе триозы, которые
могут быть использованы для образования глицерина (липосинтез) или ряда аминокислот (синтез белка), а также ацетил-CoA. Этот промежуточный продукт образуется при распаде белков, липидов и углеводов и может быть использован для липосинтеза или для извлечения энергии (см. рис. 7-1).
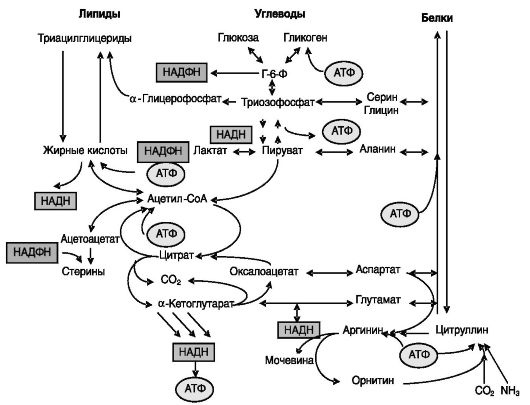 Рис. 7-1. Взаимосвязи липидного, углеводного и азотистого обмена
Рис. 7-1. Взаимосвязи липидного, углеводного и азотистого обмена
Общий конечный путь. Образующийся при распаде белков, липидов и углеводов ацетил-CoA поступает в цикл трикарбоновых кислот (цикл лимонной кислоты, или цикл Кребса), где подвергается окислению до CO2. Образующиеся при этом восстановительные эквиваленты в форме НАДН используются далее в дыхательной цепи для получения АТФ. CO2 может быть использован для синтеза жирных кислот (через малонил-CoA) и мочевины, пуринов и пиримидинов (через карбамоилфосфат), причем в мочевине конечные продукты обмена белков, липидов и углеводов (CO2 и NH3) связаны между собой непосредственно (см. рис. 7-1).
ВЗАИМОСВЯЗИ ТКАНЕЙ
В связи с разделением функций между клетками разных органов и тканей в многоклеточном организме между ними происходит постоянный обмен углеводами, липидами и аминокислотами. Центральную роль в координации обменных процессов и поддержании гомеостаза занимает печень. Она осуществляет внешнюю и внутреннюю секрецию и получает метаболическую информацию через системный кровоток и от кишечника (рис. 7-2). Организация структурно-функциональной единицы печени - печеночной дольки - показана на рис. 7-3.
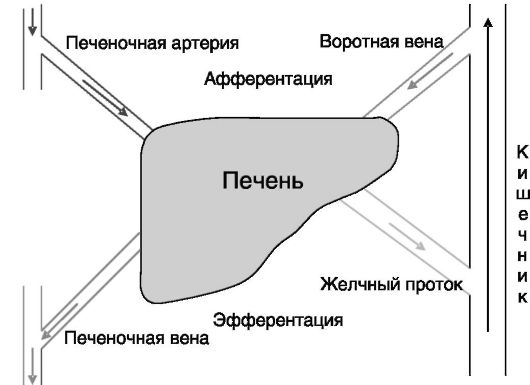 Рис. 7-2. Печень получает и перерабатывает метаболическую информацию
Рис. 7-2. Печень получает и перерабатывает метаболическую информацию
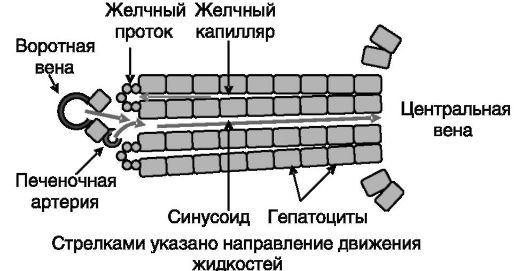 Рис. 7-3. Структура печеночной дольки
Рис. 7-3. Структура печеночной дольки
Благодаря экспрессии ряда ферментов, характерных преимущественно или исключительно для печени, в печени протекает ряд обменных процессов, которые не дублируются или дублируются лишь отчасти в других органах (рис. 7-4).
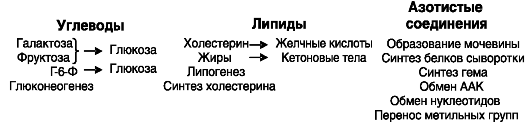 Рис. 7-4. Процессы, идущие исключительно или преимущественно в печени: ААК - ароматические аминокислоты
Рис. 7-4. Процессы, идущие исключительно или преимущественно в печени: ААК - ароматические аминокислоты
Печень выполняет функции глюко- и аминостата, играет важнейшую роль в адаптации обменных процессов к режиму питания, в том числе посредством перераспределения потоков питательных веществ между разными органами и тканями.
Взаимодействия между печенью и жировой тканью. Питательные вещества из кишечника направляются в печень, где подвергаются первичной переработке. Часть поступающих углеводов запасается в печени в форме гликогена, а избыток в виде глюкозы поступает в общий кровоток, откуда она извлекается жировой тканью для превращения в жиры и запасания. Жирные кислоты триглицеридов пищи в печени частично подвергаются дегидрогенизации и направляются (в форме липопротеинов очень низкой плотности - ЛОНП) преимущественно в жировые депо для запасания. В условиях голодания гидролиз гликогена печени обеспечивает поддержание эугликемии, а энерогозатраты большинства органов покрываются кетоновыми телами, которые образуются в печени из жирных кислот, поступающих из жировой ткани в результате липолиза (рис. 7-5).
Взаимодействия между печенью и скелетными мышцами. В условиях покоя энергетические затраты скелетной мышцы покрываются преимущественно за счет аэробного процесса окисления жирных кислот. При интенсивной работе в мышце начинает преобладать анаэробный процесс гликолиза. Образующийся в результате лактат поступает в печень, где вновь превращается посредством глюконеогенеза в глюкозу, которая направляется в мышцу. В условиях голодания аминокислоты белков мышцы становятся субстратом для глюконеогенеза в печени, что способствует поддержанию эугликемии, а основным
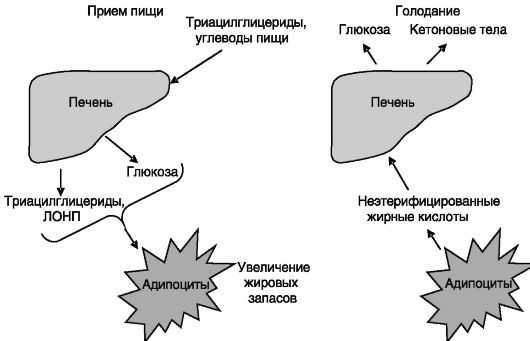 Рис. 7-5. Связи печени и жировой ткани
Рис. 7-5. Связи печени и жировой ткани
энергетическим субстратом для мышцы становятся кетоновые тела, образующиеся в печени из жирных кислот (рис. 7-6).
Взаимодействия между печенью и мозгом. Мозг является важнейшим потребителем глюкозы, и эугликемия, поддерживаемая печенью, служит одним из факторов его нормальной работы. Образующийся в процессе жизнедеятельности мозга аммиак (в частности, из нейромедиаторных аминокислот) является нейротоксичным и должен эффективно удаляться из мозга. Это удаление происходит путем связывания аммиака в глутамате и глутамине и их транспортировки в печень, где аммиак
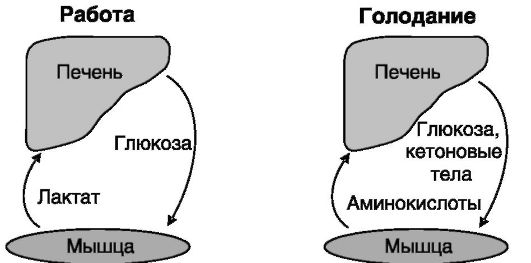 Рис. 7-6. Связи печени и мышечной ткани
Рис. 7-6. Связи печени и мышечной ткани
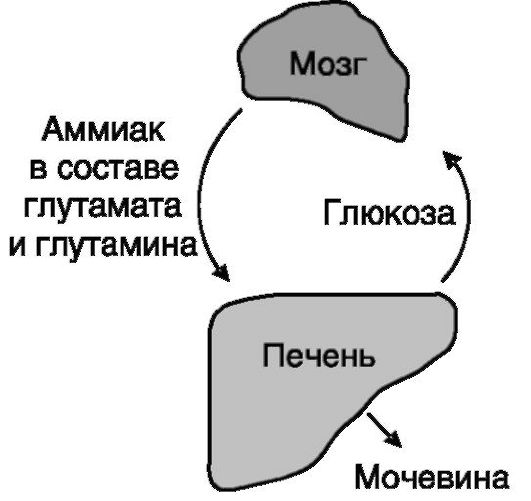 Рис. 7-7. Связи печени и мозга
Рис. 7-7. Связи печени и мозга
высвобождается и превращается в преобладающий конечный продукт азотистого обмена - мочевину. Остающийся после дезаминирования α-кетоглутарат превращается в печени в глюкозу, что способствует эугликемии (рис. 7-7).
Взаимодействия между печенью и почками. В условиях ацидоза, вызванного, например, интенсивной работой, голоданием или диабетом, главным способом выведения избытка протонов из организма становится их связывание с аммиаком
и экскреция почками в виде иона аммония. Источником аммиака служит глутамин, поставляемый печенью. Образующийся в почках после дезаминирования глутамина α-кетоглутарат используется для синтеза глюкозы (рис. 7-8). В условиях голодания это способствует поддержанию эугликемии, а при диабете усугубляет гипергликемию.
СЕНСОРЫ МЕТАБОЛИТОВ
Направленность и интенсивность обменных процессов зависят не только от доступности субстратов и энергии, но и от активности ферментов, катализирующих превращения веществ (а также транспортеров и других белков, участвующих в обменных процессах). Способы регуляции активности ферментов включают регуляцию субстратом и продуктом (в том числе влияние на стабильность фермента); аллостерическую регуляцию за счет наличия в ферменте сайтов связывания регуляторных соединений; регуляцию посредством ковалентной модификации фермента, чаще всего фосфорилирования-
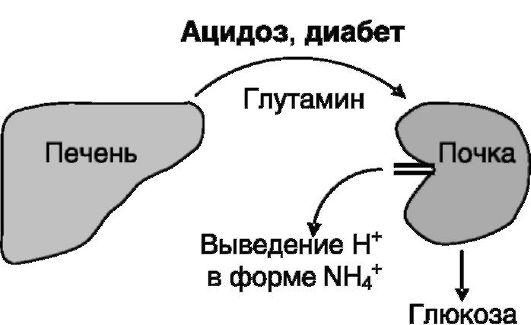 Рис. 7-8. Связь печени и почек
Рис. 7-8. Связь печени и почек
дефосфорилирования; регуляцию на претрансляционном уровне - на уровнях транскрипции генов, сплайсинга продуктов транскрипции, стабильности мРНК. Таким образом, решения об интенсивности и направленности обменных процессов принимаются как на уровне ферментов-исполнителей, так и надстроечными системами регуляции. Структуры, воспринимающие уровень метаболитов, называют сенсорами. Между сенсорами и рецепторами нет жесткой границы, хотя обычно термином «рецептор» обозначают структуры, воспринимающие соединения, специализирующиеся на сигнальной функции (например, гормоны). Так, к сенсорам метаболитов относится ряд ядерных рецепторов, например, рецептор X печени (LXR), выполняющий функцию сенсора окисленных производных холестерина. Важнейшие метаболиты воспринимаются несколькими сенсорами, функционально связанными с разными этапами обмена данного метаболита или даже другими типами обмена. Например, восприятие глюкозы может осуществляться через упомянутый выше ядерный рецептор LXR (связь углеводного и липидного обмена; опосредованно через минорный метаболит глюкозы, ксилулозо-5-фосфат, протеинфосфатазу PP2A (гликолиз, липосинтез); через другой метаболит глюкозы, фруктозо-2,6-бифосфат, фосфофруктокиназу и фруктозобифосфатазу (гликолиз и глюконеогенез).
ЭНДОКРИННАЯ РЕГУЛЯЦИЯ
Координация различных типов обмена, согласование обменных процессов в разных органах и адаптация этих процессов к режиму питания и внешним условиям происходят с участием гормональных механизмов. Практически во всех клетках организма имеются сенсоры углеводов и липидов, а также уровня энергообеспеченности клетки. Примерами клеток, реагирующими на такие сигналы изменением продукции гормонов служат α- и β-клетки поджелудочной железы (глюкагон и инсулин соответственно), а также адипоциты (лептин, адипонектин, резистин). Сигналы, поступающие в ЦНС извне и в форме изменений в концентрации глюкозы, аминокислот, гормонов желудочно-кишечного тракта и т.д., преобразуются в изменения активности симпатико-адреналовой и гипоталамо-гипофизарной систем, что сопровождается модуляцией уровня периферических гормонов, регулирующих активность разнообразных ферментов метаболизма.
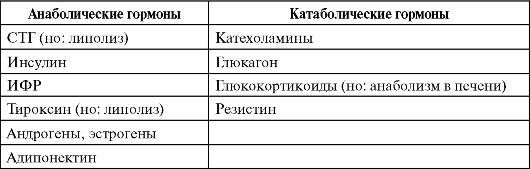
По общей направленности действия гормоны условно делят на две большие группы - анаболические и катаболические. Анаболические
гормоны способствуют процессам роста и запасания, интегральным показателем которых служит положительный азотистый баланс (преобладание задержки азота в организме над его выведением). Катаболические гормоны действуют противоположным образом.Необходимо отметить, что мишени действия разных гормонов одной группы могут совпадать лишь частично, и один и тот же гормон может действовать противоположным образом на клетки разных тканей (табл. 7-1).
Таблица 7-1. Анаболические и катаболические гормоны
 Помимо
указанных в регуляции обменных процессов принимают участие многие
другие сигнальные соединения. Например, ФНО-α служит важнейшим
индуктором кахексии (истощения) при воспалительных заболеваниях,
индуцируя, в частности, липолиз. При беременности к регуляции обменных
процессов подключаются гормоны плаценты, обеспечивающие мобилизацию
материнских запасов и направление их к растущему плоду. Индуцированное
стрессорными факторами поступление вазопрессина в передний гипофиз
меняет направленность действия этого гормона на катаболическую (через
стимуляцию секреции АКТГ и глюкокортикоидов).
Помимо
указанных в регуляции обменных процессов принимают участие многие
другие сигнальные соединения. Например, ФНО-α служит важнейшим
индуктором кахексии (истощения) при воспалительных заболеваниях,
индуцируя, в частности, липолиз. При беременности к регуляции обменных
процессов подключаются гормоны плаценты, обеспечивающие мобилизацию
материнских запасов и направление их к растущему плоду. Индуцированное
стрессорными факторами поступление вазопрессина в передний гипофиз
меняет направленность действия этого гормона на катаболическую (через
стимуляцию секреции АКТГ и глюкокортикоидов).
Рекомендуемая литература
Мак-Мюррей У Обмен веществ у человека.- М.: Мир, 1980.- 368 с.
Desvergne В., Michalik L., Wahli W Transcriptional regulation of metabolism. Physiol Rev. 2006;86:465-514.
Long Y.C., Zierath JR. AMP-activated protein kinase signaling in metabolic regulation. J Clin Invest. 2006;116:1776-1783.
Zollner G., Marschall H. U., Wagner M., Trauner M. Role of nuclear receptors in the adaptive response to bile acids and cholestasis: pathogenetic and therapeutic considerations. Mol Pharm. 2006;3:231-251.