Биология: учебник: в 2 т. / под ред. В. Н. Ярыгина. - 2011. - Т. 1. - 736 с. : ил.
|
|
|
|
Глава 6. РАЗМНОЖЕНИЕ В ЖИВОЙ ПРИРОДЕ
Среди фундаментальных свойств и проявлений земной жизни размножению принадлежит особое место (см. пп. 1.1 и 1.3). Действительно, смысл существования живого существа - выполнение им главной биологической функции: участие в процессе размножения. Основу функции размножения составляют определенные клетки (гаметы) и клеточные процессы.
Так как длительность жизни особи всегда короче продолжительности существования вида, к которому эта особь принадлежит, то история любого биологического вида заключается в истории сменяющих друг друга поколений организмов. Очередное или дочернее поколение образуется вследствие размножения представителей материнского поколения. Можно сказать, что благодаря размножению биологические виды а следовательно, и жизнь как явление материального мира сохраняются во времени.
Фенотипические различия, наблюдаемые среди особей разных поколений, представляют собой материал для естественного отбора, без чего эволюция живых форм и жизни в целом невозможна.
Размножение возникло в ходе исторического развития органического мира планеты рано, предположительно одновременно с (прото) клеткой. Размножение в мире жизни, наряду со сменой поколений и поддержанием на определенном уровне внутривидовой изменчивости, решает также задачи увеличения количества особей и сохранения путем «воспроизведения себе подобных» возникающих в эволюции типов и вариантов структурно-физиологической организации. Последнее обусловлено тем, что процесс биологического размножения предусматривает передачу в ряду поколений наследственного материала (ДНК) и, таким образом, специфической для конкретного вида биологической (генетической) информации.
6.1. способы и формы размножения
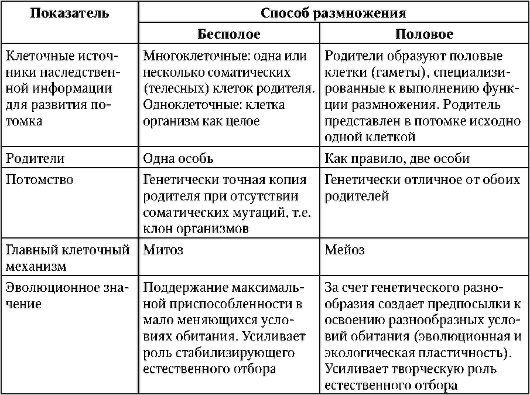
Способы и формы биологического размножения определяются характеристиками клеточного материала, который используется для размножения, а также рядом других обстоятельств (табл. 6.1). Выделяют бесполое и половое размножение.
Таблица 6.1. Сравнительная характеристика бесполого и полового размножения
6.2. бесполое размножение
Типичные формы бесполого размножения приведены на рис. 6.1.
Деление надвое приводит к образованию из одного материнского организма двух дочерних. Такая форма размножения преобладает у прокариот и простейших одноклеточных, но встречается и у многоклеточных, например, продольное - у медуз и поперечное - у кольчатых червей.
Множественное деление или шизогония наблюдается у простейших, в том числе паразитов человека (малярийный плазмодий).
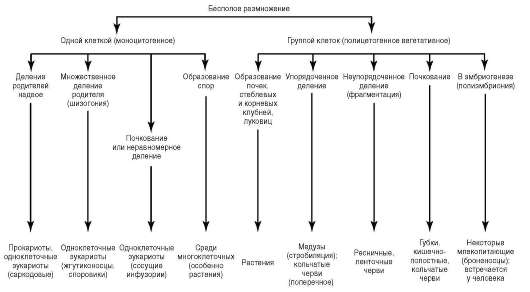
Рис. 6.1. Формы бесполого размножения
При размножении почкованием потомок первоначально формируется как вырост на теле родителя с последующей отшнуровкой - гидра.
Такая форма как фрагментация состоит в распаде тела многоклеточного организма на части, которые затем превращаются в самостоятельных особей - плоских червей, иглокожих.
У видов, размножающихся спорами, дочерняя особь развивается из специализированной клетки-споры.
Если при бесполом способе размножения клеточные источники наследственной информации для развития потомка представлены не одной, а несколькими или многими клетками родителя, говорят о вегетативном бесполом размножении. Оно распространено среди растений.
Бесполое размножение типично для организмов с относительно низким уровнем структурно-функциональной организации, среди которых встречается немало паразитов человека. У паразитов бесполое размножение служит не только решению задачи увеличения численности особей, но способствует расселению, помогает пережить периоды существования в неблагоприятных условиях.
Интересен способ, который по всем формальным признакам следует рассматривать как бесполое размножение, но сопряженное с процессом полового размножения - полиэмбриония (см. п. 6.3 и табл. 6.1).
6.3. половое размножение
Половое размножение, возникнув эволюционно позже бесполого, существует в природе тем не менее более 3 млрд лет. Оно обнаруживается в жизненных циклах представителей всех основных групп организмов. Эволюционный консерватизм и распространенность среди живых форм полового размножения имеет свои причины. Важнейшая из них - то, что оно обеспечивает значительное генетическое разнообразие особей в каждом поколении и, следовательно, высокий уровень фенотипической изменчивости потомства, благодаря чему решается задача эволюционной (сохранение жизни, пусть в измененных формах, во времени на фоне меняющихся жизненных условий) и экологической (расселение в разные среды, освоение разнообразных экологических ниш) пластичности живых существ.
Основу полового размножения составляет половой процесс. Суть его сводится к объединению в генетическом (биоинформационном) материале для развития потомка генетического материала из двух разных
источников, каковыми являются родители - самка и самец (мать и отец) или просто две особи, как это происходит у одноклеточных организмов. Представление о половом процессе дает явление конъюгации, в частности, у инфузорий, которая состоит во временном соединении путем образования «мостика» двух особей (родители) для обмена (рекомбинация) наследственным материалом. В итоге возникают две особи, генетически отличные друг от друга и от каждого из родителей. Эти особи затем размножаются бесполым путем (деление). Число инфузорий после завершения конъюгации не меняется, так что говорить о размножении в прямом смысле в этом случае нет оснований. Две задачи - использование феномена комбинативной генотипической изменчивости (половой процесс) для создания наследственного (биоинформационного) разнообразия среди потомков и увеличение числа особей (бесполое размножение) решаются путем сочетания двух разных способов. Напомним, что в обмене генетическим материалом у инфузорий участвуют микронуклеусы (см. п. 2.3).
У некоторых простейших половой процесс осуществляется в виде копуляции. В этом случае две особи (родители) соединяются в одну с объединением генетического (биоинформационного) материала и процессом рекомбинации. В дальнейшем такая особь вступает в фазу собственно размножения путем деления (бесполое размножение).
На определенном этапе эволюции (у многоклеточных) половой процесс как способ обмена генетической информацией между особями вида и, таким образом, увеличения наследственного и, следовательно, фено-типического разнообразия потомства оказался сопряженным с размножением как способом увеличения числа особей вида.
Необходимое условие полового размножения заключается в образовании родительскими особями (самкой и самцом, матерью и отцом) гамет (женских и мужских) - половых клеток, специализированных для выполнения генеративной функции (см. п. 6.4). Еще одна типичная характеристика полового размножения заключается в явлении оплодотворения - в слиянии материнской и отцовской гамет с образованием зиготы - клетки, представляющей собой дочернюю особь (потомка) на начальной одноклеточной, наиболее ранней, стадии индивидуального развития.
Есть виды организмов, у которых образование зиготы происходит путем слияния гамет, не отличимых по строению. У большинства видов, однако, по размерам, структурным, цитохимическим и цитофунк-циональным признакам гаметы делятся на женские или материнские
(яйцеклетки) и мужские или отцовские (сперматозоиды, спермии).
Как правило, яйцеклетки и сперматозоиды образуются разными особями - женскими (самки) и мужскими (самцы). В подразделении гамет на яйцеклетки и сперматозоиды (см. рис. 4.46), а особей на самок и самцов (см. рис. 4.48) состоит явление полового диморфизма. Наличие его в природе отражает различия в тех специфических задачах, которые в процессе полового размножения решают женские и мужские гаметы, самки и самцы.
Образование половых клеток обоих видов одним организмом, имеющим одновременно женские и мужские половые железы, - явление истинного гермафродитизма1.
Истинный гермафродитизм типичен для некоторых паразитов человека, например плоских червей. Хотя истинные гермафродиты производят оба типа гамет - женские и мужские, - самооплодотворения у них, как правило, не наблюдается. Обычная причина этого - несовпадение во времени созревания яйцеклеток и сперматозоидов.
Истинный гермафродитизм встречается у человека. Чаще он является результатом нарушения эмбриогенеза при одинаковой паре половых хромосом (либо XX, либо XY) во всех соматических клетках. У ряда людей-гермафродитов наблюдается мозаицизм по половым хромосомам: одна часть соматических клеток имеет пару XX, тогда как другая - XY.
Хотя оплодотворение представляет собой характерный признак полового размножения, дочерняя особь иногда развивается из неоплодот-воренной яйцеклетки - партеногенез или девственное развитие (от греч. parthenos - девственница, genos - рождение). Источником наследственного материала для развития потомка обычно бывает ДНК яйцеклетки - гиногенез. Реже наблюдается партеногенетическое развитие из клетки с цитоплазмой от яйцеклетки и ядром от сперматозоида - андрогенез. Существуют виды организмов, у которых все образующиеся яйцеклетки способны как к развитию с их оплодотворением сперматозоидом, так и к партеногенезу - факультативный партеногенез. В природе девственное развитие встречается среди растений, червей, насекомых, ракообразных - естественный партеногенез. Есть виды (кавказская скальная ящерица), размножающиеся исключительно пар-теногенетически - облигатный или обязательный партеногенез.
1 От истинного гермафродитизма следует отличать ложный гермафродитизм, для которого характерно сочетание у одного организма наружных половых органов и вторичных половых признаков обоих полов при наличии половой железы одного типа - женской или мужской (яичника или семенника).
Партеногенез как способ размножения путем образования многочисленного потомства, причем в отсутствие партнеров для спаривания, помогает решить задачу компенсации массивной гибели (часто неспецифической) организмов некоторых видов. Именно поэтому он распространен среди паразитов.
К девственному развитию яйцеклетку можно побудить в лабораторных условиях - искусственный партеногенез.
Исследования, выполненные на ранних эмбрионах человека, полученных путем экстракорпорального оплодотворения, показали, что развитие человека возможно только при наличии оплодотворения, т.е. в зиготе должны быть оба генома - материнский и отцовский. В отсутствие отцовского генома не образуются провизорные органы, и, следовательно, возникающий материал внутренней клеточной массы (эмбриобласт, согласно прежней эмбриологической терминологии) лишен возможности нормально развиваться. В таких случаях (дигиногенез - в диплоидных клетках оба генома материнские) обычно развитие приводит к образованию тератом, что равнозначно прекращению развития и гибели зародыша. В отсутствие материнского генома (диандрогенез - в диплоидных клетках оба генома отцовские) гипертрофированное развитие претерпевают ткани трофобласта, что приводит к патологическому состоянию в виде пузырного заноса. Внутренняя клеточная масса не образуется, зародыш гибнет. Можно заключить, что девственное развитие нового организма человека невозможно.
У представителей видов, для которых описан естественный партеногенез (пчелы), как и в случае типичного полового размножения, развиваются потомки с диплоидными соматическими клетками. Восстановление диплоидного набора хромосом обычно происходит путем слияния ово(оо)цита, т.е. яйцеклетки и редукционного тельца во втором делении мейоза.
Особого внимания, хотя бы потому, что речь идет об одном из возможных механизмов образования в процессе беременности женщины монозиготных близнецов, заслуживает полиэмбриония (см. табл. 6.1). Речь идет о бесполом размножении на стадии состоявшегося зародыша, начавшего развитие с оплодотворения и образования зиготы (типичная форма полового размножения), путем его разделения на две или большее число частей, каждая из которых дает в развитии полноценную особь. Среди животных полиэмбриония типична для броненосцев, у которых из первоначально одного зародыша образуется 4-8.
6.4. чередование поколений с бесполым и половым размножением
Многие виды организмов, обычно размножающиеся бесполым путем, в принципе способны к размножению половым путем. Обычно ряд поколений с бесполым размножением сменяется поколением с половым размножением или же осуществляющим половой процесс. Смена (чередование) бесполых и половых поколений у разных видов происходит с разной периодичностью, регулярно или через неодинаковые отрезки времени.
Первичное чередование поколений заключается в смене полового размножения на спорообразование. Оно описано для представителей классов споровиков - возбудителей разных форм малярии, малярийных плазмодий, и жгутиконосцев - лейшманий, трипаносом (тип Простейшие), у некоторых растений и объясняется сохранением в филогенезе соответствующих групп организмов как более древней (бесполой), так и более поздней и прогрессивной (половой) форм размножения.
Вторичное чередование поколений заключается в переходе на некоторых стадиях жизненного цикла к бесполому или партеногене-тическому размножению в группах животных, которые эволюционно уже освоили половое размножение - кишечнополостные, членистоногие.
Включение в жизненные циклы организмов, обычно размножающихся бесполым путем, полового размножения или полового процесса активизирует комбинативную генотипическую изменчивость, чем способствует преодолению генетического однообразия потомков. Этим расширяются эволюционные и экологические перспективы группы.
6.5. половые клетки (гаметы)
В сравнении с функциями других дифференцированных клеток, функция половых клеток или гамет уникальна. Они обеспечивают передачу генетической (наследственной, биологической) информации между особями разных поколений (передача биоинформации по вертикали), чем сохраняют жизнь как явление во времени. Гаметы представляют собой одно из многих направлений дифференцировки клеток многоклеточных живых существ. У человека, например, таких направлений порядка 220250. Половые клетки образуют особую клеточную линию, специализированную для выполнения репродуктивной функции.
Предположительно клетки этой линии образуются из бластомеров, имеющих на вегетативном полюсе цитоплазму особого рода - зародышевую (половую) плазму1, богатую РНК.
У некоторых видов организмов (двукрылые насекомые) зародышевая плазма в виде специфических гранул обособляется очень рано - до начала дробления, фактически в яйцеклетке. Если зародышевую плазму разрушить, например, подействовав на нее УФ-лучами, то развиваются стерильные особи, у которых гаметы не образуются.
Разделение клеток начавшего индивидуальное развитие организма на линию половых клеток и соматические происходит обязательно. У одних видов это случается достаточно рано. Так, у веслоногого рака циклопа на 5-м делении дробления, у плодовой мухи - на 13-м делении, у бесхвостых амфибий (лягушки) - на стадии бластулы. Сравнительно поздно клетки, имеющие в цитоплазме зародышевую плазму, обособляются в клетки-непосредственные предшественницы половых клеток, у млекопитающих это происходит на стадии гаструляции.
По сравнению с соматическими клетками зрелые половые клетки имеют типичные отличия. Во-первых, это гаплоидный (у человека n = 23) набор хромосом в ядрах. Благодаря этому вследствие оплодотворения в зиготе восстанавливается типичный для вида диплоидный (у человека 2n = 46) набор хромосом. Во-вторых, это необычное для других клеточных типов значение ядерно-цитоплазматического отношения, которое у яйцеклеток снижено в силу значительного количества цитоплазмы (в частности, благодаря наличию желтка) - у соматических клеток оно обычно выражается дробью 1/6, тогда как у яйцеклеток - 1/15. У сперматозоидов ядерно-цитоплазматическое отношение повышено в силу малого количества цитоплазмы. В-третьих, это низкий уровень обменных процессов, близкий к состоянию анабиоза. В-четвертых, сперматозоиды неспособны вступать в митотический цикл, а у яйцеклеток эта способность восстанавливается вследствие оплодотворения или действия фактора, провоцирующего партеногенез. В-пятых, только зигота - клетка, образующаяся вследствие оплодотворения, т.е. в результате слияния мужской и женской гамет, характеризуется истинной тотипотент-ностью и в связи с этим может рассматриваться как универсальная стволовая клетка-родоначальница. Именно ее потомки дают
1 Первым термин «зародышевая плазма» использовал А. Вейсман, но совершенно в ином смысле - для обозначения наследственного вещества клеточного ядра (фактически хромосом).
в дальнейшем все цитотипы многоклеточного организма соответствующего вида (у человека таких цитотипов 220-250). При парте-ногенетическом развитии, не требующем оплодотворения, свойство универсальной тотипотентной стволовой клетки характеризует яйцеклетку.
Существуют различия между женскими и мужскими половыми клетками, что обусловлено различными функциями яйцеклетки и сперматозоида в процессе размножения. Так, яйцеклетки имеют оболочки, которые выполняют защитную функцию, обеспечивают требуемый уровень обмена веществ, препятствуют проникновению в яйцеклетку ядра более чем одного сперматозоида (блокируют полиспермию), способствуют у плацентарных животных имплантации (внедрению) зародыша в стенку матки, поддерживают форму зародыша. По крайней мере, у некоторых видов животных клетками (фолликулярными, питающими) оболочек яйцеклетки образуются некоторые виды и(м)РНК, которые затем используются в белковых синтезах зародыша. Так, в этих клетках транскрибируются так называемые гены с материнским эффектом - Bicoid (соответствующий белок экспрессируется на переднем полюсе) и Nanos (соответствующий белок экспрессируется на заднем полюсе) зародыша плодовой мухи. Эти белковые продукты создают градиенты, благодаря которым определяется положение переднего (головного) и заднего концов зародыша, краниальное (ростральное) и каудальное направления. В создании переднезадних координат участвует также ген Hunchback, транскрипция и трансляция которого осуществляются в клетках самого зародыша. Для яйцеклетки характерна ооплазма-тическая (плазматическая) сегрегация. После оплодотворения (у асцидий через 5 мин), но до начала делений дробления происходит закономерное перераспределение в объеме фактически яйцеклетки участков цитоплазмы разного химического состава. В последующем развитии цитоплазма этих участков закономерно обнаруживается в клетках разных, но конкретных эмбриональных зачатков (фактически тканей и органов). Можно думать, что способность бластомеров дифференцироваться в определенные клеточные типы каким-то образом связана со свойствами той цитоплазмы, которую они наследуют в процессе делений дробления. Наличие и закономерная локализация в цитоплазме зародышевой (половой) плазмы рассматриваются как частный случай ово(оо)плазматической сегрегации.
Сперматозоид имеет аппарат движения в виде жгутика. В семенной жидкости мужская половая клетка человека демонстрирует ско-
рость порядка 5 см/ч. Если исходить из соотношения преодолеваемого расстояния и длины движущегося объекта, то при названной скорости сперматозоид человека перемещается в 1,5 раза быстрее пловца олимпийского ранга. Женская половая клетка лишена аппарата активного движения. Расстояние до полости матки, равное примерно 10 см, она преодолевает с током жидкости в маточных (фаллопиевых) трубах за 4-7 сут.
Сперматозоиды некоторых видов животных имеют так называемый акросомный аппарат, выбрасывающий при контакте с яйцеклеткой особую нить. Путем растворения ферментами, выделяемыми акросомным аппаратом, оболочек яйцеклетки достигается образование своеобразного «канала» и проникновение ядра спермия в цитоплазму женской гаметы. Наряду с акросомным аппаратом, у представителей иных видов описаны другие приспособления и механизмы, способствующие оплодотворению.
6.5.1. ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ (ХРОМОСОМЫ, ХРОМАТИН, ДНК) ГАМЕТ И СОМАТИЧЕСКИХ КЛЕТОК. КЛОНИРОВАНИЕ МНОГОКЛЕТОЧНЫХ ЖИВОТНЫХ
В истории биологии был период, когда половые и соматические клетки противопоставляли друг другу, наделяя только гаметы всей полнотой свойств жизни, проносимых ими через поколения.
В конце XIX века А. Вейсман сформулировал идею, которую можно рассматривать как первую попытку объяснить природу генетического механизма клеточной дифференцировки. Согласно этой идее, клеточные деления бывают двух типов - равнонаследственные и неравнонаследственные. Неравнонаследственно делятся соматические клетки. В результате таких делений дочерние клетки в сравнении с материнской теряют некоторую часть хромосомного материала (де-минуция хромосом), причем клетки разных направлений дифферен-цировки теряют разные фрагменты хромосомного материала. В своих воззрениях А. Вейсман исходил из данных наблюдений за поведением хромосомного материала делящихся клеток круглого червя лошадиной аскариды. Наследственный материал в неизменном объеме сохраняют гаметы. Поэтому, по мнению А. Вейсмана, именно половые клетки, и только они, способны обеспечить развитие нового полноценного организма.
Более поздние исследования показали, что неравнонаследственные клеточные деления, связанные с потерей части хромосомного материала, имеют ограниченное распространение и не представляют собой генетический механизм клеточной дифференцировки. Так, у другого круглого червя, популярного в настоящее время объекта генетических исследований Caenorhabditis elegans, клеточные деления не сопровождаются деминуцией хромосом. Даже у лошадиной аскариды хромосомный материал в связи с делениями теряется не во всех клеточных линиях, т.е.
не при всех направлениях клеточной дифференцировки. В настоящее время гипотеза А. Вейсмана имеет только историческое значение.
Определяющее значение имеют результаты современных экспериментальных исследований, доказавших возможность развития полноценного организма на основе наследственного материала (генетической, биологической информации) ядра соматической дифференцированной клетки, например кишечного эпителия (рис. 6.2). Особое место в этом плане принадлежит работам, выполненным в Великобритании группой Дж. Гердона на бесхвостых амфибиях (лягушка). Принципиально опыт заключался в следующем. Уничтожалось ядро яйцеклетки, после чего в нее вводилось ядро дифференцированной соматической клетки и обеспечивались условия для развития. Такие яйцеклетки в 1-2% экспериментов в процессе развития давали взрослых лягушек. Приведенный результат доказывал, что наследственный мате-
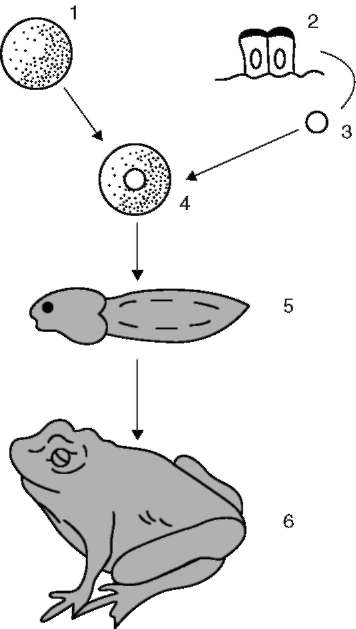
Рис. 6.2. Успешное клонирование лягушки, доказывающее полноценность наследственного материала соматической дифференцированной клетки: 1 - яйцеклетка с разрушенным УФ-лучами ядром - источник цитоплазмы; 2 - эпителиальные клетки кишечника головастика - источник ядерного наследственного материала; 3 - ядро; 4 - имплантация ядра соматической клетки в лишенную собственного ядра яйцеклетку; 5 - клонированный головастик; 6 - клонированная лягушка
риал (ДНК) дифференцированных соматических клеток позвоночных в количественном и качественном (информационно-содержательном) отношении является биологически полноценным. Вместе с тем уже тогда возникли подозрения, что клонированные животные чаще в сравнении с лягушками, появляющимися на свет обычным путем, имели дефекты развития.
В последующие годы были развернуты работы по клонированию высших животных, в частности млекопитающих. Знаковым событием, хотя единственный позитивный результат пришелся на 236 попыток, стало клонирование шотландскими учеными овцы Долли. Для получения Долли использовали энуклеированную (лишенную ядра) яйцеклетку овцы породы шотландская черномордая, в которую ввели диплоидное ядро клетки молочной железы беременной овцы породы финский дорсет. Описанную клеточную конструкцию активировали к дроблению посредством электрического разряда. По достижении развивавшимся ex vivo зародышем определенной стадии его имплантировали в матку приемной (суррогатной) матери-овцы. Есть сообщения об успешном клонировании млекопитающих других видов животных - коровы, мыши, лошади, собаки и др. (2-2,8% успеха).
Результативное клонирование млекопитающих разных видов породило амбициозную идею клонирования человека. Здесь, однако, сразу же последовали указания на наличие, наряду с техническими и биологическими проблемами, также проблем этического и правового порядка. В Нью-Йорке в ООН работает Комитет по клонированию человека, соответствующие комитеты национального уровня существуют во многих странах мира, в том числе в РФ. В настоящее время принято различать два вида клонирования человека: репродуктивное (задача - получить нового человека, генетически и, как предполагают некоторые люди, фенотипически близкого или даже идентичного человеку-донору соматического ядра) и терапевтическое (задача - вырастить зародыш в условиях ex vivo до стадии бластоцисты с целью получения из внутренней клеточной массы эмбриональных стволовых клеток, которые затем будут использованы в интересах регенеративной медицины - см. п. 3.2). Протесты против любых форм клонирования людей носят массовый, международный и многоконфессионный характер. Государственной думой РФ принято и подтверждено решение (Федеральный закон о временном запрете ...) о моратории на работы в области репродуктивного клонирования.
При обсуждении проблемы клонирования высших животных нередко обходят стороной известные и сейчас уже не единичные факты. Эти факты указывают на то, что клонированные особи в своей массе характеризуются сниженным здоровьем и жизнеспособностью. Так, овца Долли умерла, прожив лишь половину среднего для овец срока жизни. Мыши, полученные путем клонирования, причем в разных лабораториях, отличаясь пониженной жизнеспособностью, проживают в целом не более половины срока, соответствующего средней продолжительности жизни их линии. Способность к обучению у клонированных мышей снижена.
Настораживают данные о низком проценте успешного клонирования в сравнении с числом предпринимаемых попыток. Так, согласно мировой статистике, на начало 2002 г. из общего числа попыток получить новый организм путем клонирования успехом (рождением животного) завершилась лишь небольшая часть: овцы - создано 3156 эмбрионов, получено 50 ягнят; коровы - 8600 эмбрионов, 111 телят; мыши - 7613 эмбрионов, 54 мышонка; обезьяны - 78 эмбрионов, 2 родившиеся обезьянки. Смертность плодов и новорожденных среди клонируемых животных достигает 85%. Более 1/3 из числа родившихся и выживших клонированных животных имеют серьезные нарушения здоровья, в том числе угрожающие жизни.
Одно время немало можно было слышать о репродуктивном клонировании как инструменте получения гениев и/или возвращении в настоящую жизнь выдающихся личностей прошлого. На сегодняшний день есть основания говорить о несомненном успехе современной биологии, решившей технические вопросы клонирования даже высших животных. Открытым, однако, остается вопрос о том, насколько точно клонированные животные могут копировать свой прототип. Особенно остро названный вопрос стоит в отношении клонирования людей. Во всяком случае нельзя забывать о том, что стартовая генетическая программа индивидуального развития особи (индивидуума) проявляет себя во вполне определенных, достаточно вариабельных условиях среды - I-го порядка (генотипической), IIа, IIб и III-го порядка (см. п. 4.3.1.1). Следует также иметь в виду, что принципиальное место в формировании человека как личности принадлежит культурной (социальной) программе индивидуального развития.
Гаметогенез - процесс образования гамет или половых клеток: яйцеклеток (овогенез или оогенез) и сперматозоидов (сперматогенез).
В нем выделяют ряд стадий (рис. 6.3).
Между процессами ово(оо)генеза и сперматогенеза имеются различия. Так, стадия формирования выделяется фактически только в сперматогенезе. Стадия размножения сперматогенеза осуществляется в половой железе - в семеннике, начиная с момента достижения мужскими особями состояния половой зрелости. Размножение ово(оо)го-ний происходит в яичнике, главным образом в эмбриогенезе. Наиболее интенсивно у людей этот процесс протекает между 3-м и 7-м месяцами внутриутробного развития, а завершается на 3-м году жизни. Стадия роста ово(оо)генеза более сложна, отчасти в связи с накоплением в цитоплазме яйцеклетки питательного материала желтка, а также в связи с явлением ово(оо)плазматической сегрегации, а стадия созревания женских половых клеток растянута во времени и завершается в том случае, если происходит оплодотворение.
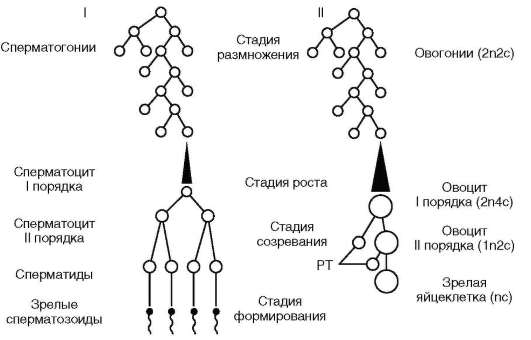
Рис. 6.3. Гаметогенез (схема): I - сперматогенез; II - ово(оо)генез; n - число хромосом в гаплоидном наборе; с - количество ДНК в гаплоидном наборе хромосом n; РТ - редукционные тельца
Образование функционально зрелых сперматозоидов в семенниках происходит на протяжении всей взрослой жизни мужчины. Интенсивность процесса может снижаться по достижении мужчиной 50-летнего возраста. Продукция зрелых половых клеток прекращается с достижением женским организмом климактерического периода онтогенеза.
На стадии размножения клетки-предшественницы гамет называются ово(оо)гониями и сперматогониями. Эти клетки осуществляют серию последовательных митотических делений, что приводит к существенному росту их количества. Так как клетки-предшественницы женских и мужских гамет размножаются обычным митозом, то ово(оо) гонии и сперматогонии вне митотического цикла так же, как все соматические клетки, характеризуются диплоидностью, и в отношении числа хромосом, и в отношении количества ДНК - 2n2c, где n - число хромосом в гаплоидном наборе, с - количество ДНК в гаплоидном наборе хромосом. В процессе митотического цикла (после завершения митоза и до синтетического периода интерфазы) хромосомы названных клеток представлены парами гомологичных аутосом и парой половых гетерохромосом, каждая из которых содержит по одной биспирали ДНК - 2n2с. По завершении синтетического периода (на протяжении постсинтетического периода интерфазы) число хромосом остается прежним, однако каждая из них содержит две биспирали ДНК - 2n4c. В метафазе митоза хромосомы представлены каждая двумя дочерними хроматидами, соединенными только в области центромеры, фактически хромосомами будущих дочерних клеток - 4n4с. По завершении митоза в части числа хромосомных наборов и количества ДНК диплоидные дочерние клетки приобретают обычный вид - 2n2с.
Среди сперматогоний выделяют клетки двух типов: светлые (А) и темные (Б). Темные клетки - неделящиеся или покоящиеся - рассматривают как стволовые. Светлые сперматогонии активно размножаются, поставляя клеточный материал для образования зрелых сперматозоидов.
На стадии роста наблюдается увеличение размеров клеток-предшественниц половых клеток, которые уже называются ово(оо)цитами и сперматоцитами I порядка (первичные овоциты и сперматоциты). При этом ово(оо)циты I порядка крупнее сперматоцитов I порядка. Увеличение клеточных размеров на названной стадии объясняют накоплением веществ, необходимых для предстоящего деления. Больший вклад в рост размеров ово(оо)цитов I порядка вносит накопление в их цитоплазме питательного материала - желтка. Так, растущие ово(оо)циты плодовых мух за три дня увеличивают объем в 90 000 раз, лягушек -
в 64 000 раз, мыши - более чем в 40 раз. Наиболее распространенный способ, обеспечивающий рост ово(оо)цитов I порядка и наблюдаемый, в частности, у млекопитающих, связан с наличием и трофической (питающей) активностью особых фолликулярных (питающих) клеток. Стадию роста делят на два периода - превителлогенеза (до образования и накопления желтка) и вителлогенеза (образование и накопление желтка). Превителлогенез (он же период малого или цитоплазматическо-го роста яйцеклетки) характеризуется относительно незначительным и пропорциональным увеличением объемов ядра и цитоплазмы без изменения значений ядерно-цитоплазматического отношения. Многие эмбриологи считают, что на стадию роста приходится предмейотическая репликация ДНК, тогда как другие эмбриологи относят это событие к профазе редукционного деления стадии созревания. Вителлогенез (он же период большого или трофоплазматического роста) характеризуется объемным увеличением цитоплазмы в связи с появлением в ней питательного материала - желтка, который представляет собой сложное вещество белково-липидно-углеводной природы. Следствием периода большого роста является выраженное снижение значений ядерно-цитоплазматического отношения.
Стадия роста ово(оо)цитов I порядка у некоторых видов животных укладывается в достаточно короткое время, тогда как у других занимает продолжительный отрезок времени. Так, у человека длительность стадии роста яйцеклеток может составлять около 30 лет.
6.5.2.1. Мейоз
Основное событие стадии созревания - мейоз, способ образования половых клеток, который состоит из двух последовательных быстро происходящих друг за другом делений - редукционного и эквационного.
Мейоз (рис. 6.4) решает две важные задачи. Во-первых, образуются клетки (гаметы) с гаплоидным набором хромосом. Этот результат достигается благодаря тому, что два деления мейоза происходят при однократной репликации ДНК. До настоящего времени нет полной ясности, к какой из стадий гаметогенеза следует отнести эту репликацию: происходит ли она в завершающей фазе стадии роста или в самом начале стадии созревания, непосредственно перед профазой 1 деления мейоза или даже во время профазы. С одной стороны, есть мнение, что ово(оо) цит I-го порядка, завершив цитоплазматические преобразования стадии роста, сразу же вступает в профазу первого деления стадии созревания. С другой стороны, ряд эмбриологов относят предмейотическую репли-
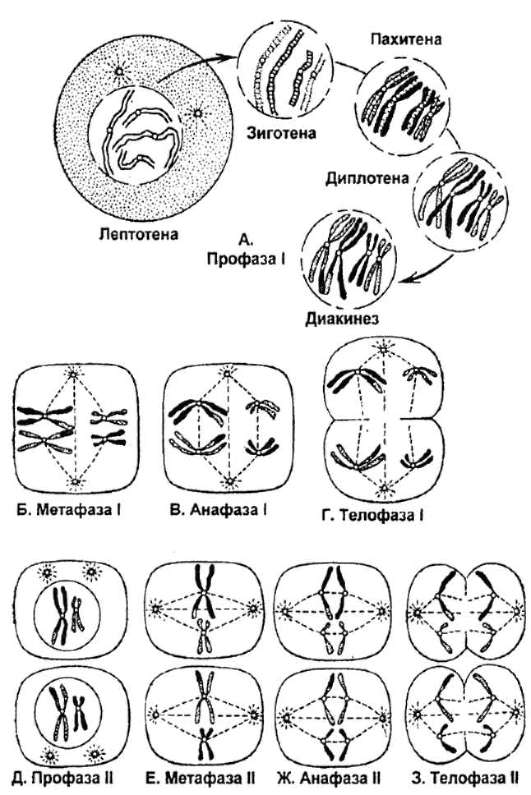
Рис. 6.4. Мейоз (схема)
кацию ДНК к началу профазы первого деления мейоза. Нельзя исключить, что репликация ДНК, начавшись на стадии роста, завершается в начале стадии созревания. Во-вторых, в профазе и анафазе первого деления мейоза заложены механизмы генотипической комбинативной изменчивости, что делает гаметы генотипически отличными от клеток-предшественниц половых клеток, а также в целом от соматических клеток обоих родителей.
Вступая в первое деление (редукционное) стадии созревания ,
клетки имеют диплоидный набор хромосом, но увеличенное вдвое количество ДНК - 2n4c.
Так же как в обычном митозе, в профазе названного деления происходит компактизация (спирализация) материала хромосом. Вместе с тем в отличие от обычного митоза в нем наблюдается попарное сближение (конъюгация) гомологичных хромосом, которые тесно контактируют друг с другом взаимосоответствующими (гомологичными) участками. Результат конъюгации - образование пар хромосом или бивалентов, число которых n. Поскольку каждая хромосома, вступающая в мейоз, состоит из двух хроматид, то бивалент представлен четырьмя хромати-дами - n4c. В профазе I мейоза отмечается формирование веретена деления. К концу профазы степень спирализации хромосом в бивалентах возрастает, и они укорачиваются. Профаза первого деления мейоза занимает в сравнении с профазой обычного митоза больше времени. В ней выделяют несколько стадий.
Лептотена - хромосомы начинают процесс спирализации и становятся видимыми в микроскоп как тонкие и достаточно длинные нитчатые структуры.
Зиготена - соответствует началу конъюгации гомологичных хромосом, объединяемых в биваленты особыми структурами - синап-тонемальными комплексами (рис. 6.5). Если не все гомологичные хромосомы конъюгируют и остаются неспаренные хромосомы вне бивалентов, клетка гибнет апоптозом.
Пахитена - на фоне продолжающейся спира-лизации хромосом и их укорочения гомологичные хромосомы осуществляют кроссинговер или перекрест, заключающийся в обмене взаимосоответствующими (гомологичными) участками. Кроссинговер обеспечивает перекомбинацию отцовских и материнских аллелей в группах сцепления (гомологичных
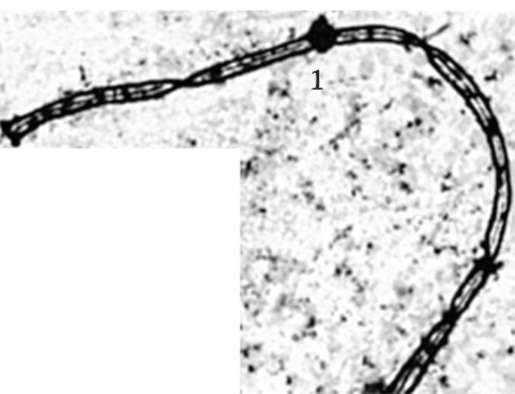
Рис. 6.5. Образование бивалентов конъюги-рующими гомологичными хромосомами в зи-готене профазы I мейоза: 1 - центромера
хромосомах). Перекрест хромосом может происходить в различных местах хромосом, в связи с чем кроссинговер в каждом конкретном случае приводит к обмену разными участками генетического материала. Возможны образование нескольких перекрестов между двумя хроматидами (рис. 6.6) или обмен взаимосоответствующими фрагментами происходит между более чем двумя хроматидами бивалента (рис. 6.7). Все это повышает эффективность кроссинговера как механизма генотипиче-ской комбинативной изменчивости.
Диплотена - гомологичные хромосомы начинают отдаляться друг от друга, в первую очередь в области центромер, но сохраняют связь в местах произошедшего кроссинговера - хиазмы. Можно говорить о продольном расщеплении конъюгировавших гомологичных хромосом по всей их длине. В итоге каждая пара хромосом воспринимается как комплекс из четырех структур-хроматид (дочерних хромосом) - тетрада (рис. 6.8).
Диакинез - завершает профазу первого деления мейоза; гомологичные хромосомы остаются в составе бивалентов, однако их связь ограничивается только отдельными точками хиазм (рис. 6.9). Сами биваленты приобретают форму колец, восьмерок, крестов.
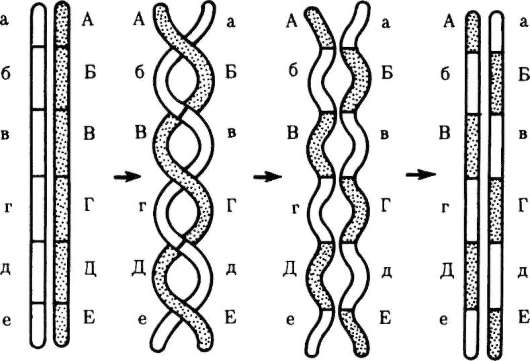
Рис. 6.6. Многократный кроссинговер между гомологичными хромосомами (схема): А-Е, а-е: локусы хромосом
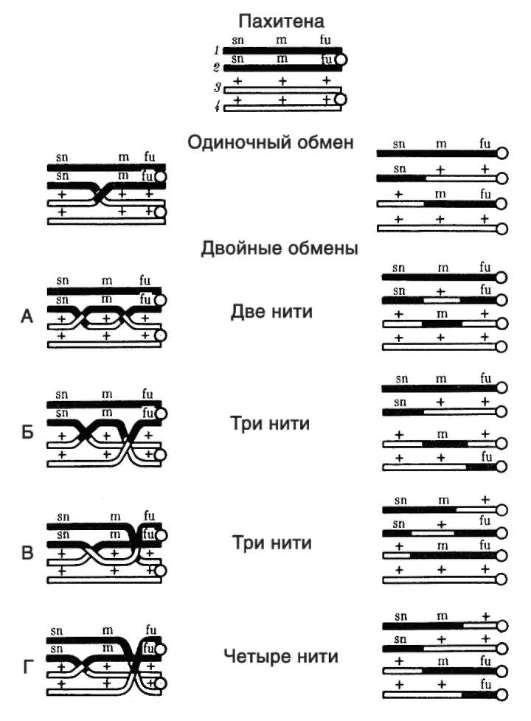
Рис. 6.7. Множественный обмен участками между четырьмя хроматидами в па-хитене профазы I мейоза (схема): в кроссинговере могут участвовать все четыре хроматиды бивалента; латинскими буквами обозначены мутантные аллели, знаком «+» - аллели дикого типа (нормальные)
В период диакинеза прохождение клетками-предшественницами гамет редукционного деления приостанавливается (согласно более ранним представлениям, это происходит уже в диплотене), в связи с чем этот период называют стационарным. Деление возобновляется и
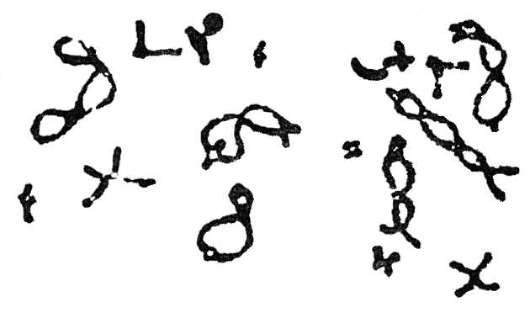
Рис. 6.8. Диплотена в профазе I мейоза кузнечика

Рис. 6.9. Диакинез в профазе I мейоза человека: стрелками показаны хиазмы
завершается в случае овуляции яйцеклетки (см. здесь же, ниже) и ее оплодотворения. Несмотря на характеристику периода диакинеза как стационарного, в нем активно происходят синтетические процессы. Эти процессы относятся к прогенезу (предзародышевому периоду онтогенеза), поскольку результаты этих процессов в виде синтезируемых
молекул и образуемых структур необходимы в основном для ранних стадий развития зародыша. Во-первых, речь идет об амплификации ДНК (см. также п. 2.4.3.4-а), которая заключается в образовании многочисленных копий генов рибосомных РНК - малой (18S) и большой (28S) субъединиц. Копии, став самостоятельными, преобразуются морфологически в ядрышки числом до нескольких тысяч. В таких ядрышках образуются субъединицы рибосом, которые используются для организации биосинтеза белков клетками зародыша. По завершении своей функции эти ядрышки перемещаются в цитоплазму и там разрушаются. В диакинезе амплифицируются гены 5S рибосомных РНК и тРНК. Эти РНК нарабатываются в необходимых (т.е. больших) количествах «впрок» для белковых синтезов тоже в эмбриогенезе. Благодаря амплификации генов время «наработки» требуемого для ранних стадий эмбриогенеза количества, например, рибосом у африканской шпорцевой лягушки (Xenopus laevis) сокращается с 500 лет до 3 мес. Во-вторых, в период диакинеза профазы I мейоза хромосомы приобретают вид «ламповых щеток» (см. п. 2.4.3.4-а), чем обеспечивается образование «впрок» для нужд зародыша определенного набора и(м)РНК. Описанные процессы наиболее полно изучены на бесхвостых амфибиях (лягушка), для которых характерна относительно поздняя (стадия га-струлы) активизация собственного генома. У млекопитающих, например, полное биоинформационное обеспечение процессов эмбриогенеза за счет функционально-генетической активности (транскрипции) собственных генов отмечается начиная со стадии 8 бластомеров.
В метафазе первого деления мейоза завершается формирование веретена деления. Нити этого веретена, связанные, в частности, с центромерами гомологичных хромосом, направляются к разным полюсам. Такое положение нитей обеспечивает закономерную ориентацию бивалентов в плоскости экватора веретена деления.
В анафазе первого деления мейоза благодаря ослаблению связей между гомологичными хромосомами в бивалентах и закономерной ориентации бивалентов в метафазной пластинке гомологи каждого бивалента расходятся к разным полюсам клетки. При этом гомологичные хромосомы отцовского и материнского происхождения каждой пары расходятся независимо друг от друга. В результате на полюсах клеток по завершении анафазы I стадии созревания мейоза собираются «случайные» ассоциации гомологичных хромосом отцовского и материнского происхождения. Независимое расхождение к полюсам в анафазе редукционного деления хромосом отцовского и материнского происхождения
разных бивалентов представляет собой, наряду с кроссинговером, еще один эффективный механизм генотипической комбинативной изменчивости. В этом случае происходит перекомбинация целых групп сцепления, причем с уже измененным в сравнении с хромосомами родителей вследствие прошедшего кроссинговера набором аллелей.
Благодаря особенностям анафазы, в результате телофазы первого деления мейоза образуются гаплоидные клетки. Однако хромосомы в таких клетках представлены двумя хроматидами, т.е. содержат две би-спирали ДНК - п2с.
Второе (эквационное) деление стадии созревания мейоза проходит без репликации ДНК и дает клетки с гаплоидным набором хромосом (к полюсам расходятся отдельные хроматиды), каждая из которых содержит одну биспираль ДНК - nc.
Особенность стадии созревания ово(оо)генеза в сравнении с одноименной стадией сперматогенеза заключается в асимметричном характере обоих мейотических делений. В результате в ово(оо)генезе из одного ово(оо)цита I порядка образуется одна функционально полноценная яйцеклетка и три так называемых редукционных или полярных тельца (одно - вследствие асимметричного деления яйцеклетки и два - вследствие симметричного деления редукционного тельца, возникшего при первом делении стадии созревания). Это мелкие клетки, которые гибнут (но: см. п. 6.2). По завершении первого деления мейоза и отделения первого полярного тельца клетка, которая даст зрелую яйцеклетку, приобретает название ово(оо)цит II порядка (вторичный овоцит).
Асимметричность делений способствует сохранению в одной женской гамете всего запаса питательных и иных, необходимых для развития нового организма, веществ.
По завершении стадии созревания сперматогенеза образуются четыре клетки, каждая из которых даст полноценный сперматозоид - пс.
Стадия созревания сперматогенеза завершается образованием клеток, называемых сперматидами. Сперматиды, чтобы стать функционально зрелыми сперматозоидами, проходят стадию формирования. На этой стадии хроматин уплотняется, изменяются форма и размеры ядра, формируется аппарат активного движения клетки - жгутик, образуется акросома (у представителей некоторых видов), перестраивается мито-хондриальный аппарат клетки, она теряет некоторую часть цитоплазмы.
Гаметогенез - высокопродуктивный процесс. За период половой жизни мужчина производит порядка 500 млрд сперматозоидов. На 5-м месяце внутриутробного развития в половой железе женского организ-
ма насчитывается 6-7 млн клеток-предшественниц яйцеклеток. К началу репродуктивного периода (постнатальный онтогенез) в яичниках присутствует примерно 100 000 ово(оо)цитов I порядка. От момента полового созревания женского организма до прекращения гаметогенеза (менопауза) в яичниках созревает 400-500 клеток-предшественниц яйцеклеток, готовых к оплодотворению. На протяжении репродуктивного периода постнатального онтогенеза в яичниках женщины под влиянием лютеинизирующего гормона гипофиза ежемесячно, как правило, одна женская гамета покидает яичник (овуляция - разрыв зрелого граафо-вого пузырька; яйцеклетка сначала попадает в свободную брюшную полость, а затем в маточную трубу, где может произойти оплодотворение) и, будучи оплодотворенной, возобновляет мейоз.
Виды, размножающиеся половым путем, характеризуются типичной структурой жизненного цикла, в котором происходит чередование гаплоидной и диплоидной фаз (см. п. 4.3.7.1 и рис. 4.47).
6.5.3. ПЕРВИЧНЫЕ ПОЛОВЫЕ КЛЕТКИ
Ово(оо)генез и сперматогенез (см. п. 6.4.3) происходят в дифференцированных по полу женских или мужских половых железах - яичниках или семенниках. Прежде чем попасть в эмбриональную закладку половой железы, клетки-предшественницы гамет, называемые первичными половыми клетками или гоноцитами , проходят достаточно сложный путь.
Половой зачаток, содержащий первичные половые клетки - гоно-бласт, - образуется бластомерами, имеющими в цитоплазме зародышевую или половую плазму (см. п. 6.4). У млекопитающих первичные половые клетки обособляются в эмбриогенезе относительно поздно (гаструла).
Первичные половые клетки имеют внегонадное происхождение. У людей, например, они обособляются во внутренней клеточной массе (эпибласт, эмбриобласт в более ранней эмбриологической терминологии) и представляют собой потомки тотипотентных эмбриональных стволовых клеток. Их можно видеть в составе зародышевых листков, хотя они не являются производными клеток ни одного из них. На гистологических препаратах первичные половые клетки идентифицируют по относительно крупным размерам (12-20 мкм в диаметре), форме, приближающейся к округлой, центральному положению светлого ядра с крупным ядрышком. Главный гистохимический маркер - высокая
активность фермента щелочной фосфатазы в цитоплазме. Из зародышевых структур они перемещаются во внезародышевые - стенку желточного мешка (внезародышевая мезодерма и энтодерма). В определенном участке энтодермы желточного мешка, вблизи места отхож-дения аллантоиса первичные половые клетки концентрируются перед тем, как начать движение в закладки половых желез. Миграция начинается после 25 сут развития. Она происходит в основном по интерсти-циальному типу, т.е. через мезенхиму различных внезародышевых и зародышевых образований путем так называемого контактного ориентирования. Тонковолокнистый слой толщиной порядка 30 нм на поверхности гоноцитов специфически взаимодействует с макромолекулами межклеточного матрикса, например, с фибронектином, лами-нином, коллагеном IV типа. Названные белки внеклеточного матрикса, взаимодействуя с белками клеточных оболочек, преимущественно из семейства интегринов, определяют маршруты движения клеток в эмбриогенезе (фибронектин). Часть первичных половых клеток достигает закладок половых желез пассивным образом, с кровотоком. Предположительно определенная роль в миграциях гоноцитов принадлежит хемотаксису, причем молекулы-аттрактанты, выполняя функцию ориентиров (своеобразных «маяков»), определяющих направление движения гоноцитов, образуются клетками половых валиков (см. п. 4.3.7.1). Есть мнение, что они ответственны также за задержку первичных половых клеток в капиллярах вблизи закладок половых желез. В ткани закладок гоноциты из кровотока попадают путем диапедеза, т.е. через сосудистую стенку без нарушения ее целостности. Первичные половые клетки у человека достигают закладок гонад и «обосновываются» там (осуществляют хоуминг) между клетками целомического эпителия, которые создают необходимые условия для их дальнейшего развития, на 28-30-е сутки внутриутробного периода. Первичные половые клетки, не достигшие закладок половых желез, гибнут путем апоптоза (см.
п. 3.1.2).
Если по какой-то причине в зачатке гонады или вне его гоноциты осуществляют несколько «лишних» митотических циклов, то они теряют перспективу стать гаметами и становятся полипотентными стволовыми клетками (стволовые гаметоциты). В таком случае они могут привести к образованию тератомы.
У позвоночных животных первичные половые клетки или гоно-циты являются единственными предшественницами зрелых половых клеток, или гамет. У низкоорганизованных многоклеточных животных
клетками-предшественницами гамет могут стать тотипотентные резервные стволовые клетки (археоциты у губок) или даже высоко дифференцированные соматические клетки (хоаноциты - воротничковые клетки, имеющие такие специализированные структуры, как жгутики, у губок).
Вопросы для самоконтроля
1. В чем различия полового и бесполого размножения, и каково биологическое значение этих способов размножения?
2. Как образуются половые клетки? Перечислите и охарактеризуйте этапы гаметогенеза?
3. Какова роль мейоза в рекомбинации генетического материала?