Биология: учебник: в 2 т. / под ред. В. Н. Ярыгина. - 2011. - Т. 1. - 736 с. : ил.
|
|
|
|
Раздел III. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ. Глава 4. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИЗНИ - РЕАЛИЗАЦИЯ СВОЙСТВ НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ КЛЕТОЧНОГО АППАРАТА НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ (ГЕНЕТИЧЕСКИЙ АППАРАТ)
Переход от «преджизни» к жизни связывают с появлением наследственности (см. п. 1.4.4.1). Есть неопровержимые доказательства непротиворечивости наследственности и изменчивости как фундаментальных свойств живых форм. В перечне всеобщих, обусловленных структурой эволюционного процесса, уровней организации земной жизни (см. п. 1.6) молекулярно-генетический уровень занимает первую строку. Элементарной структурой на этом уровне является ген, определяемый как фрагмент молекулы ДНК, несущий определенный объем биологически целесообразной (полезной для жизнедеятельности, репродукции и индивидуального развития) генетической информации. Элементарное явление на этом уровне состоит в конвариантной редупликации ДНК, т. е. в самоудвоении молекул нуклеиновой кислоты путем самокопирования с некоторым количеством ошибок. Такого рода ошибки (генные, истинные мутации) сохраняются в ряду поколений клеток и особей. Кроме генных мутаций, имеются также хромосомные и геномные мутации, к которым, однако, неприложимо определение «истинные». Различия состоят в том, что генные мутации способны производить принципиально новую биологическую информацию, тогда как хромосомные и геномные мутации заключаются либо в изменении
количества (дозы) уже имеющейся информации, либо в появлении ее новых комбинаций. Вне явлений наследственности и изменчивости, обусловливающих в своем взаимодействии консерватизм и динамичность структур и функций в мире жизни, реальность появления и сохранения во времени новых адаптаций (приспособлений) и, следовательно, процесс биологической эволюции, невозможны.
4.1. наследственность и изменчивость - фундаментальные свойства живого
Жизнь существует на планете более 3,5 млрд лет, что связано с особенностями ее временной организации в виде сменяющих друг друга поколений. Происходит смена органелл (например, митохондрий) в клетках, поколений клеток в многоклеточном организме, смена поколений организмов в популяциях, смена видов в биоценозе, смена биоценозов, образующих биосферу. Сохранение живых форм в меняющихся условиях окружающего мира возможно благодаря их эволюции, в ходе которой у них появляются и закрепляются в потомстве свойства, дающие возможность приспособиться к новой среде обитания. В основе эволюции, в свою очередь, лежит способность живых систем разного уровня к самовоспроизведению с изменениями. Такое самовоспроизведение обеспечено наличием в мире жизни двух универсальных свойств - наследственности и изменчивости.
Свойства наследственности и изменчивости традиционно рассматривают в связи с клеткой или организмом. На клеточном и организ-менном (онтогенетическом) уровнях организации живого (см. п. 1.6) под наследственностью понимают свойство соответственно клеток или организмов в процессе самовоспроизведения передавать новому поколению способность к определенному типу обмена веществ и развития. В ходе развития у потомства формируются общие признаки данного типа клеток или вида организмов, проявляются некоторые индивидуальные особенности родительских форм. На популяционно-видовом уровне организации жизни свойство наследственности состоит в поддержании во времени постоянного состава и соотношения генетических форм или гено(аллело)ти-пов в ряду поколений организмов данной популяции (вида). На био-ценотическом уровне длительное существование биоценоза обеспечено сохранением определенных соотношений популяций (видов) организмов. Это предполагает воспроизводство во времени совокупности
популяций (видов), присущих конкретному биоценозу, без изменения их гено(аллело)фондов.
В процессе исторического развития жизни (эволюции) наследственность, используя механизм репродукции, обеспечивает тиражирование и закрепляет в ряду поколений биологически полезные приобретения (консерватизм), что делает ее обязательным фактором эволюции.
Сохранение живых форм во времени на фоне меняющихся условий (в частности, абиотических - смена на планете восстановительной атмосферы на окислительную порядка 1,9 млрд лет назад, перемещение участков земной коры, континентов и субконтинентов со сменой на обширных территориях климата, покровные оледенения и межледниковые периоды) было бы невозможным при отсутствии наработки новой биологической информации. В результате появления новой биоинформации возникают отсутствовавшие ранее структурно-функциональные фенотипические признаки и их комплексы, которые оказываются биологически полезными в новых условиях среды.
Свойство живых форм приобретать наследуемые изменения, и, комбинируя их в потомстве в различных сочетаниях, существовать в разных фенотипических вариантах, что обусловливает их эволюционную и экологическую пластичность, называется изменчивостью. Биологическая изменчивость как предпосылка создания, накопления и использования новой биоинформации лежит в основе способности живых форм выживать во времени, несмотря на смену параметров среды (эволюционная пластичность) или в один и тот же исторический период заселять территории, различающиеся по комплексу жизненно важных условий (экологическая пластичность).
У клеток определенного цитофенотипа и организмов определенного вида изменчивость, затрагивая процесс развития (гистогенез для клеток, эмбриогенез для организмов c половым размножением), проявляется в
наличии различий между разными клетками одного цитофено-типа (цитотипа) и особями одного вида. На популяционно-видовом уровне организации жизни это свойство проявляется в генетическом разнообразии популяций вида. На биоценотическом уровне появление новых видов и популяций вносит изменения в межвидовые отношения в биоценозах. Изменчивость обусловливает появление и использование новой биологической информации (динамизм), что создает почву для функционирования механизма естественного отбора. Это делает изменчивость, наряду с наследственностью, обязательным фактором эволюции.
4.1.1. ФОРМЫ БИОЛОГИЧЕСКОЙ ИЗМЕНЧИВОСТИ
С одной стороны, обязательная характеристика живых форм состоит в наличии у них генотипа и фенотипа (см. п. 1.3), с другой - биоинформация, связанная с генами (сайтами, нуклеотидными последовательностями ДНК) напрямую, не принимая участия в процессах жизнедеятельности и развития непосредственно, является в функционально-генетическом плане фактически «потенциальной», тогда как актуализированная (действующая) биоинформация связана с белками и, следовательно, фенотипическими признаками и свойствами клеток и организмов (см. п. 2.4.5.4). Это порождает проблемы, во-первых, реализации генотипической биоинформации в фенотипиче-скую биоинформацию (см. п. 2.4.5.4; 2.4.5.5 и 2.4.5.6) и, во-вторых, выделения разных форм биологической изменчивости.
Биологическую изменчивость подразделяют на генотипическую и фенотипическую. Генотипическая изменчивость распространяется на генетический аппарат - структурные (смысловые, кодирующие, транскрибируемые, транслируемые) гены или сайты (нуклеотидные последовательности) ДНК с иными функциями, хромосомы, геном, генотип, кариотип. Генотипическая изменчивость подразделяется на мутационную и комбинативную. Мутационная генотипическая изменчивость реализуется по уровням структурно-функциональной организации генетического аппарата (см. п. 4.3). Соответствующие изменения носят название мутаций, которые бывают генными, хромосомными и геномными. Примеры мутаций разного уровня приведены ниже (генные - см. п. 4.3.1.3; хромосомные - см. п. 4.3.2.2; геномные - см. п. 4.3.3.1). Отметим еще раз, что только с генными мутациями связано появление новой, ранее не существовавшей в природе биоинформации. Хромосомные и геномные мутации в функционально-генетическом отношении сводятся либо к изменению количества биоинформации (делеции, дупликации участков хромосом, гаплоидные, полиплоидные или анэуплоидные клетки и организмы), либо к перекомбинации блоков биоинформации разного объема (транслокации, транспозиции, инверсии, инсерции).
Современная (молекулярная) генетика расширяет область знаний, касающихся форм биологической изменчивости. В частности, в дополнение к генным, хромосомным и геномным мутациям классической генетики, возникающим скачкообразно (сальтаторно) и поэтому удовлетворяющим принципу «все или ничего» (мутация либо происходит,
либо нет), описаны «динамические» мутации - экспансия тринуклео-тидных повторов (см. п. 4.3.1.3).
Биоинформационное обеспечение функций митохондрий клеток имеет свои особенности. Речь идет, в частности, о взаимодействии генов митохондриальной и ядерной локализации. Такие взаимодействия необходимо учитывать, поскольку их наличие вносит дополнения в представления о полном объеме генотипических (мутационных) изменений, случающихся в эукариотических клетках. В сферу интересов современной медицинской генетики прочно вошли митохондриальные болезни, причиной которых могут быть в том числе изменения генов, приводящие к нарушению механизмов взаимодействия ядерного и митохондри-ального геномов (межгеномные сигнальные эффекты, см. п. 4.3.1.3).
Относительно недавно в профессиональном словаре генетиков и эмбриологов появился термин - геномный импринтинг (от англ. imprint - отпечаток), или геномная память. Суть явления заключается в том, что оба родителя передают потомству в принципе одинаковые гены, например, занимающие гомологичные локусы в паре гомологичных аутосом, но эти гены несут отпечаток пола родителя, давшего через свою гамету в зиготу хромосому с импринтированным сайтом. Импринтированны-ми могут быть как отдельные сайт или хромосома, так и геном в целом (отцовский или материнский, но не оба одновременно). Импринтиро-ванная генетическая структура (сайт, хромосома, геном) выключается из функции. Так как импринтинг предположительно связывают с метилированием ДНК, его рассматривают как фактор эпигенетической регуляции генетической активности (конкретно подавление этой активности). Последнее не позволяет считать случаи проявления геномной памяти мутациями. Тем не менее рассматриваемый феномен характеризуется отчетливыми фенотипическими изменениями, в том числе патологическими - болезни импринтинга (у человека более 30). При вовлечении у людей в импринтинг целиком генома развиваются истинный пузырный занос (диандрогенные особи - оба генома диплоидных клеток, начиная с зиготы, отцовские) и тератомы (дигиногенные особи - оба генома материнские). В обоих случаях зародыш нежизнеспособен, что заставляет исключить для людей возможность партеногенетического развития (т. е. без оплодотворения яйцеклетки спермием).
Импринтирование критического участка хромосомы 15 (q11.2-q13) дает либо синдром Ангельмана (Энжельмена) - отцовская одноро-дительская дисомия, либо синдром Прадера-Вилли - материнская однородительская дисомия по указанному участку. В первом случае
импринтирован соответствующий сайт материнской хромосомы 15 и, таким образом, генетически активен отцовский, во втором - наоборот.
Феномен комбинативной генотипической изменчивости состоит в образовании различных сочетаний (комбинаций) структур генетического аппарата по уровням его организации - генов (аллелей или аллельных генов), хромосом или их участков, геномов. В функционально-генетическом плане каждое такое сочетание - уникальный по биоинформационному содержанию комплекс. Типичный пример - половое размножение. В гаметогенезе в профазе первого деления мейоза путем рекомбинации (кроссинговер) меняется генный (аллельный) состав гомологичных хромосом. В анафазе этого же деления благодаря независимому расхождению к полюсам клетки негомологичных хромосом отцовского и материнского происхождения в хромосомных наборах дочерних клеток объединяются разные по происхождению и, следовательно, аллельному составу хромосомы, причем от клетки к клетке (фактически от гаметы к гамете) в несовпадающем числовом отношении. При оплодотворении в зиготе на случайной основе комбинируются геномы яйцеклетки и спермия.
Область выражения фенотипической изменчивости, как это следует из названия, - фенотип. Некоторые признаки характеризуются вариабильностью (изменчивостью), в основе которой лежат не геноти-пические изменения на уровне генов, хромосом или генома, а влияния на фенотипические проявления генов факторов среды, прежде всего внешней (3-го порядка, см. п. 4.3.1.1). Такие ненаследственные изменения, отвечающие понятию фенотипической изменчивости, называются модификациями. Классические примеры модификаций - разная в зависимости от температуры окружающей среды окраска шерсти на различных участках тела у кроликов горностаевой породы (рис. 4.1) и разная форма воздушных, плавающих и подводных листьев растения стрелолиста. Соответственно, говорят о модификационной феноти-пической изменчивости. Биологи-эволюционисты подчеркивают роль этой формы биологической изменчивости в процессе видообразования, видя в ней фактор высокой экологической, а также эволюционной пластичности определенных видов животных и растений. С наличием в природе модификаций связывают колебания степени выраженности признаков, описываемые в генетике понятиями экспрессивности и пе-нетрантности (см. п. 4.3.1.1).
Феномен модификационной изменчивости, вариабильность степени выраженности признаков в зависимости от условий среды, создает
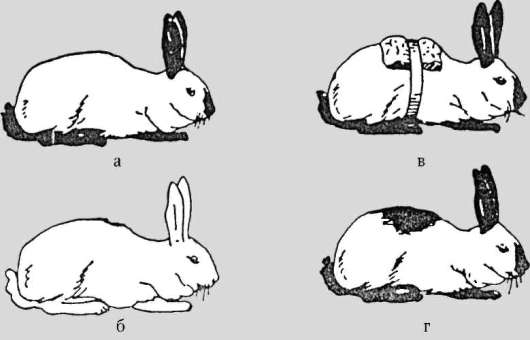
Рис. 4.1. Изменения пигментации шерстного покрова в зависимости от температуры у кроликов горностаевой породы: а - кролик, выращенный при температуре 14-18 °С; б - кролик, выращенный с рождения при температуре более 30 °С; в - кролик с удаленным на спине участком шерсти и помещенным на этом месте пузырем со льдом; г - тот же кролик после того, как на участке с удаленной шерстью и пузырем со льдом шерсть отросла
проблему отношений между геном и соответствующим ему признаком, генотипом и фенотипом. В связи с названной проблемой необходимо остановиться на генетическом понятии «норма реакции». По существу речь идет о характере (норме) реакции конкретного гена или генотипа в целом на определенные условия среды 1-го, 2-го, 3-го порядка (см. п. 4.3.1.1), в которых они реально функционируют. Известны гены с узкой и гены с широкой нормой реакции. Первые дают неизменный фенотипический результат в широком спектре условий, тогда как вторые отличаются значительной вариабельностью феноти-пического результата их генетической активности. Так, гены, определяющие принадлежность человека к группе крови систем АВ0 или резус (Rh), характеризуются узкой нормой реакции. Гены, контролирующие окраску шерсти кроликов горностаевой породы (см. рис. 4.1), - широкой. Даже в отношении генов с узкой нормой реакции существует возможность, пусть редко реализующаяся, возникновения условий, изменяющих фенотипический результат их генетической активности или блокирующих эту активность. В качестве примера приведем из-
вестный еще классической генетике «бомбейский феномен». Речь идет о рождении женщиной с группой крови I (I0I0) ребенка с группой крови IV (IAIB). Очевидно наличие в генотипе матери аллелей IA , IB или обеих одновременно, которые у нее фенотипически не проявились. Объяснение кроется в особенностях генотипической среды женщины, в явлении рецессивного эпистаза - одной из форм взаимодействия не-аллельных генов (см. п. 4.3.3.1).
Наряду с модификационной, выделяют случайную фенотипиче-скую изменчивость - костную мозоль на месте сросшегося перелома. О непрерывной фенотипической изменчивости говорят тогда, когда распределение особей с разной степенью выраженности признака соответствует нормальному. Такое наблюдается, в частности, при полимерном типе полигенного наследования (см. табл. 4.3) количественных признаков, например, роста у людей.
4.2. история представлений об организации и функционировании генетического аппарата
Наследственность и изменчивость как неотъемлемые признаки живых существ обеспечиваются особым материальным субстратом. В процессе развития биологической науки представления о химической природе этого субстрата, его свойствах, принципах организации и функционирования конкретизировались и расширялись.
1865 г., Г. Мендель. Экспериментально обосновал и сформулировал идею о наследственных задатках, их дискретности (корпускулярности), специфичности и аллельном состоянии, наличии доминантных и рецессивных вариантов наследственных задатков (аллелей) или признаков, присутствии в половых клетках только одного аллеля (гаплоидность, чистота гамет) каждого наследственного задатка, а в соматических клетках - двух (диплоидность). Открыл правило независимого комбинирования аллелей разных наследственных задатков у потомка. Открытые Г. Менделем свойства и особенно правила поведения наследственных задатков (менделизм) справедливы для генетики организмов, размножающихся половым путем. В 1909 г. В. Иогансен назвал наследственные задатки Г. Менделя генами (от греч. genos - род, происхождение).
1865 г., Ф. Гальтон . Выходит в свет работа «Наследование таланта и характера», с чего начинается генетика количественных признаков человека (биометрическая генетика). Следствие указанной работы - возникновение «евгеники». В формировании методических
и методологических основ биометрической генетики принимал активное участие К. Пирсон. В 1918 г. Р. Фишер высказал подтвердившееся затем предположение, что в наследовании количественных признаков важная роль принадлежит взаимодополняющему участию многих наследственных задатков (полимерное наследование).
1887 г., А. Вейсман. Выдвинул умозрительную теорию (гипотезу) о наличии бессмертной «зародышевой плазмы» и смертной «сомато-плазмы», в которой предвосхитил современные представления о хромосомах как носителях наследственности (вейсманизм).
1888 г., В. Вальдейер. Предложил термин «хромосома» для обозначения особых ядерных структур.
1900 г., Г. де Фриз , К.Э. Корренс, Э. Чермак . Переоткрыли для науки и медицинской практики закономерности наследования признаков, установленные Г. Менделем в 1865 г.
1901 г., Г. де Фриз. Сформулировал основные положения мутационной теории, предложил термин «мутация».
1902-1907 гг., Т. Бовери , У. Сеттон. Пришли к заключению и представили доказательства, что внутриклеточными носителями генетической программы являются хромосомы.
1902 г., У. Бэтсон. Ввел термины «генотип» и «фенотип».
1902 г., А. Гаррод. Показал аутосомно-рецессивный тип наследования алкаптонурии (выделение с мочой вещества алкаптона, окисляемого кислородом воздуха до соединения темного цвета, которое выпадает в осадок; у генетически нормальных лиц алкаптон метаболи-зирует до воды и углекислоты через стадию образования ацетоуксусной кислоты).
1905 г., Фараби . Показал аутосомно-доминантный тип наследования брахидактилии (короткопалость).
1909 г., В. Иогансен. Предложил называть единицы наследственности (наследственные задатки) генами, совокупность генов организма - генотипом, проявление генов (совокупность признаков и свойств) организма - фенотипом (см. здесь же, выше: 1902 г., У. Бэтсон).
1909 г., К.Э. Корренс, Э. Баур. Открыли явление цитоплазмати-ческой (нехромосомной) наследственности.
1910-1925 гг., Т. Морган. Обобщив результаты исследований возглавляемого им коллектива, сформулировал основные положения хромосомной теории наследственности (морганизм). Нобелевская премия 1933 г. «За открытия, касающиеся роли хромосом в наследственности».
1926 г., Х.Дж. Мёллер. Обнаружил мутагенное действие рентгеновых лучей. Нобелевская премия 1946 г. «За открытие возникновения мутаций под действием рентгеновых лучей».
1926 г. С.С. Четвериков . Отметил значение генетических процессов в популяциях организмов для эволюции.
1932 г., С .Н. Давиденков. Определил принципиальные направления профилактики наследственных болезней (борьба с возникновением новых мутаций, медико-генетический совет в семьях, медико-социальная охрана лиц, наследственно предрасположенных к определенной патологии). Заложил основы медико-генетического консультирования (МГК), т. е. использования генетических знаний в интересах практического здравоохранения.
1944 г., О.Т. Эйвери. Установил химическую природу вещества наследственности - ДНК.
Начало 1950-х годов, Б. Мак-Клинток. Открыла МГЭ («прыгающие» гены, транспозоны). Нобелевская премия 1983 г. «За открытие подвижных элементов генома».
1953 г., Дж. Уотсон . Ф. Крик, М. Уилкинс . Представили доказательства организации ДНК в виде спирали из двух комплементарных друг другу молекул. Нобелевская премия 1962 г. «За открытие молекулярной структуры нуклеиновых кислот и ее значения в передаче информации в живой материи». В 1928 г. Н.К. Кольцов, предвосхитив положения современной генетики и молекулярной биологии, сформулировал предположение о матричном принципе репродукции хромосом.
1955-1966 гг., Х.Г. Корана, М.У. Ниренберг. Расшифровали кодовую систему записи генетической информации в молекулах нуклеиновых кислот - генетический код. Нобелевская премия (совместно с Р.У. Холли, который раскрыл роль в биосинтезе белков тРНК) 1968 г. «За расшифровку генетического кода и его функции в синтезе белков».
1956 г, Дж. Тийо, А. Леван. Установили истинное число хромосом в соматических (диплоидных) клетках человека - 46.
1958 г., Ж. Лежен. Открыл трисомию по хромосоме 21 при синдроме Дауна.
1959 г., К.Э. Форд с коллегами, П.А. Джекобс, И.А. Стронг. Установили связь синдромов Шерешевского-Тернера и Клайнфельте-
ра с изменениями числа половых хромосом.
1960 г. (Денвер); 1963 г. (Лондон); 1966 г. (Чикаго). Унифицированы представления по классификации и номенклатуре равномерно окрашенных хромосом человека; 1971 г. (Париж). Уточнена и конкре-
тизирована номенклатура хромосом человека по результатам их дифференциальной (избирательной) окраски, обеспечивающей персональную идентификацию отдельных хромосом; 1978 г. Введена ISCN - An international system for human cytogenenic nomenclature - «путеводитель» по кариотипу человека.
1961 г., Ф. Жакоб, Ж.Л. Моно. Сформулировали гипотезу о переносе генетической информации с ДНК на белок при участии иРНК. Нобелевская премия (совместно с А.М. Львовым) 1965 г. «За открытия, касающиеся генетической регуляции синтеза ферментов и вирусов».
1970-е годы, Р .Д. Робертс и Ф.А. Шарп. Предоставили доказательства интрон-экзонной организации генов эукариот. Нобелевская премия 1993 г. «За открытие расщепления генов». Экспериментальные доказательства делимости гена представлены А.С. Серебровским и Н.П. Дубининым в 1929 г.: мутации в разных частях «сложного» гена achaete scute (редукция щетинок) плодовой мухи приводили к отсутствию разных щетинок.
1989-2001 гг., Дж. Уотсон, Ф. Коллинз, К. Вентер, фирма Salera и Международная организация по изучению генома человека (Human Genome Organization - HUGO), многие участники из разных стран, включая Россию. Начаты и завершены вчерне работы по проекту «Геном человека». Секвенируются геномы других организмов, в том числе возбудителей инфекционных и паразитарных болезней, а также ближайших «эволюционных родственников» человека.
Кроме того:
1937-1953 гг., Дж.У. Бидл, Э.Л. Тейтам, Дж. Ледерберг. Нобелевская премия 1958 г. «За открытия, касающиеся генетической рекомбинации и организации генетического аппарата бактерий».
1955-1957 гг., С. Очоа, А. Корнберг. Нобелевская премия 1959 г. «За открытие механизмов биологического синтеза нуклеиновых кислот вне живого организма».
1909 г., Ф.П. Раус. Нобелевская премия 1966 г. «За открытие онко-генных вирусов».
1940-1980 гг., Р. Леви-Монталчини, С. Коэн. Нобелевская премия 1986 г. «За открытие факторов роста».
1970-е годы, Дж.М. Бишоп, Х.Э. Вармус. Нобелевская премия 1989 г. «За открытие клеточного происхождения ретровирусных онкогенов».
1980-е годы, С.Б. Прузинер. Нобелевская премия 1997 г. «За открытие прионов - новой биологической причины инфекций».
Конец ХХ-начало XXI вв., Э. Блэкберн, К. Грейдер, Д. Шостак.
Нобелевская премия 2009 г. «За открытие механизма защиты хромосом теломерами и теломеразами».
4.3. уровни организации генетического аппарата эукариот
В современном мире жизни материальным носителем свойств наследственности и изменчивости является ДНК, «выигравшая» историко-эволюционное «соревнование» у РНК (см. п. 1.4.5). Этому способствовали ее большая химическая стабильность и особенности ма-кромолекулярной и надмолекулярной организации. ДНК - высокомолекулярное полимерное соединение. Независимая комбинация по длине макромолекул троек из четырех нуклеотидов-мономеров, строящих ДНК, позволяет записать необходимый объем биоинформации, а надмолекулярная организация в виде двойной спирали делает возможным матричный синтез. Он составляет основу тиражирования (репликация ДНК - см. п. 2.4.5.3) биоинформации для передачи в ряду поколений или копирование (транскрипция информационной или матричной РНК - см. п. 2.4.5.5) этой информации для использования в организации процессов жизнедеятельности. Участки макромолекул ДНК могут быть химически модифицированы (например, метилированы), что в процессе эволюции стало механизмом регуляции генетической активности. ДНК образует химические связи с белками, что также было использовано эволюцией для создания тонких механизмов регуляции генетических функций. Напомним, что в эукариотических клетках ДНК присутствует в виде комплекса с гистоновыми (основными по химической характеристике) белками, выполняющими роль ингибиторов генетической активности, а негистоновые (кислые по химической характеристике) белки, ослабляя указанное действие гистонов путем взаимодействия с ними, обусловливают возможность использования биоинформации, присутствующей в ДНК, причем в клетках многоклеточных организмов частями.
Несмотря на химическую стабильность, нуклеотидные последовательности в макромолекулах ДНК могут быть изменены. При этом такие изменения сохраняются в структуре биополимера при его репликации.
Решение задач, которые жизнедеятельность ставит перед эукариоти-ческими клетками, особенно у многоклеточных форм, требует большой точности и надежности биологических механизмов. Возможно, что, по
крайней мере, отчасти в связи с этим, их генетический аппарат (аппарат наследственности и изменчивости) претерпел в эволюции изменения в сторону его усложнения.
В генетическом аппарате эукариотической клетки (эукариотических организмов, включая человека) выделяют три уровня структурной и одновременно функционально-генетической организации: генный, хромосомный и геномный. На каждом из них решаются свои специфические задачи, с одной стороны, наследственности, а с другой, - биологической изменчивости с целью требуемого биоинформационного обеспечения процессов жизнедеятельности, размножения, индивидуального (онтогенез) и исторического (филогенез, эволюция) развития.
Наряду с такими понятиями, как «ген», «хромосома» и «геном», существуют важные генетические понятия «генотип» и «кариотип», имеющие непосредственное отношение к структурно-функциональной организации генетического аппарата эукариот.
Генотип - это совокупность аллелей всех генов или нуклеотидных последовательностей, сайтов ДНК в диплоидном наборе хромосом. Кариотип - это парный (диплоидный) набор хромосом в ядре соматической клетки организмов соответствующего биологического вида. Существует определение кариотипа с элементами конкретизации. В соответствии с этим определением кариотип рассматривается как совокупность признаков хромосомного набора (не только число, но и размер, форма хромосом), характерных для соматических (диплоидных) клеток того или иного вида организмов.
4.3.1. ГЕННЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ГЕНЕТИЧЕСКОГО АППАРАТА. ОПРЕДЕЛЕНИЕ ГЕНА. ПРИЗНАК КАК ГЕНЕТИЧЕСКОЕ ПОНЯТИЕ
Функциональной единицей генетического аппарата, определяющей возможность развития отдельно взятого признака клетки или организма, является ген. Именно так определяет понятие «ген» (по Г. Менделю, наследственный задаток) классическая генетика.
С развитием генетики возник ряд версий определения гена, как правило, конкретизирующего характера. Если определение, вытекающее из научного багажа классической (домолекулярной) генетики, следует рассматривать как функционально-генетическое, то два наиболее заметных определения более позднего времени характеризуются как биохимическое и молекулярно-биологическое.
Биохимический подход в изучении природы элементарных фено-типических признаков, контролируемых напрямую генами, привел в 1950-е годы ХХ в. к заключению, что таковыми являются ферменты. Появился тезис «один ген - один фермент». Соответственно ген стали определять как участок макромолекулы ДНК, контролирующий образование конкретного белка-фермента (биохимическое определение гена).
Согласно молекулярно-биологической версии ген определяется как фрагмент макромолекулы ДНК, в котором содержится информация об аминокислотной последовательности полипептида (экспресси-руемый, транскрибируемый и транслируемый ген) или нуклеотидной последовательности РНК определенного вида, прежде всего рРНК и тРНК (экспрессируемый, транскрибируемый, но не транслируемый ген, см. также п. 2.4.5.5 - транскрибируемые, но не транслируемые 5' и 3' участки транскриптона эукариот). Молекулярно-биологическое определение гена в наибольшей степени соответствует понятию «структурный (смысловой, кодирующий, экспрессируемый) ген».
Молекулярно-генетическая версия определения гена не противоречит ни функционально-генетической (классической), ни биохимической версии. Она лишь указывает на необходимость определенных дополнений. Так, и это сейчас известно, полипептиды, образуемые клеткой или организмом под контролем соответствующих генов, вносят свой вклад в процессы жизнедеятельности и индивидуального развития не только благодаря каталитическим свойствам, но и выполняя регуляторные и сигнальные (транскрипционные и ростовые факторы, цитокины и рецепторы к ним), строительные (коллагены), транспортные (глобины) и другие функции. Семейство транскрибируемых и нетранслируемых генов, кодирующих специфические по решаемым функциональным задачам разновидности РНК (выполняющие, например, конценсусные функции - см. п. 2.4.5.5, малые ядерные РНК) также пополняется.
Проект «Геном человека» показал, что нуклеотидные последовательности, отвечающие молекулярно-биологическому определению гена, составляют не более 5% от суммарного количества ДНК. В настоящее время популярность приобретает термин сайт. Им обозначают нуклеотидную последовательность, занимающую конкретное место в биспирали ДНК определенной хромосомы. В известной мере термин «сайт» можно рассматривать как синоним термина «локус» классической генетики. Таким образом, сайт - это конкретная нуклеотидная последовательность, соответствующая по функционально-генетической
характеристике, прежде всего, структурному гену. Не исключаются, однако, любые другие функции - регуляторная, сервисная, конценсусная.
Пояснения заслуживает редко используемый сейчас термин «ци-строн». По-существу, цистрон - это ген. Чаще термин «цистрон» используют, говоря о прокариотах. Для прокариотических геномов характерна полицистронная организация функциональных генетических единиц, полицистронный формат транскрипции генов и, следовательно, трансляции соответствующих белков (см. п. 2.4.5.6) с единицей транскрипции опероном. Для эукариот типичен моноцистронный формат транскрипции и трансляции (исключением, возможно, являются гомео-зисные гены - см. п. 4.3.3.2). Единицей транскрипции в этом случае является транскриптон (см. п. 2.4.5.5).
Результат генетической активности состоит в определенном феноти-пическом проявлении, т. е. в возникновении признака. Под признаком в генетике понимают единицу морфологической, физиологической, биохимической, иммунологической, клинической и любой другой дискретности клетки (организма) или, иными словами, отдельное качество или свойство, по которому одну клетку (организм) можно отличить от другой (другого). Большинство признаков клетки или организма относится к категории сложных. Сложные признаки для своего оформления требуют синтеза многих веществ, прежде всего белков со специфическими свойствами - ферменты, структурные, сократительные, транспортные, рецепторные. Механизмы формирования сверхсложных морфологических (морфогенезы), физиологических (функциогенезы), поведенческих признаков наиболее полно проявляют себя в ходе индивидуального развития особи.
Процесс реализации генетической информации в фенотип организма в онтогенезе начинается с находящегося под прямым генетическим контролем образования простых белков (полипептиды, протеины). Функциональные свойства последних определяются аминокислотной последовательностью, которая задается последовательностью триплетов нуклеотидов в ДНК соответствующего структурного (смыслового, кодирующего, транскрибируемого и транслируемого, экспрессируемо-го) гена. Таким образом, полипептид, будучи первичным продуктом генетической активности, свойства которого напрямую определяются геном, является элементарным (простым) фенотипическим признаком. В соответствии с приведенным выше определением, элементарные фенотипические признаки организма - это в основном простые белки с каталитической, транспортной, рецепторной и другими
функциями. Особое место в выяснении закономерностей наследования и изменчивости таких признаков, в том числе патологических, принадлежит биохимическим и иммунохимическим методам генетического анализа человека (см. пп. 5.2.2.4 и 5.2.2.5).
Одно время распространение приобрел тезис «один ген - один полипептид». Открытие альтернативного сплайсинга (см. п. 2.4.5.5), генов-«матрешек», когда структурный ген меньшего размера размещается в пределах другого более крупного гена, наличия у одного, обычно крупного, гена нескольких промоторов (гомеозисные гены - см. п. 4.3.3.2) делают этот тезис либо неприемлемым вообще, либо имеющим ограниченное распространение. В научно-педагогической литературе высказывается мнение, что известным в настоящее время генетическим и молекулярно-биологическим фактам не противоречит тезис «один полипептид - один ген».
4.3.1.1. Свойства гена. Среда как генетическое понятие
Ген как функционально-генетическая единица наделен рядом свойств.
Во-первых, он отличается специфичностью действия. Это означает, что конкретный ген обусловливает возможность присутствия в фенотипе клетки (организма) конкретного признака. Известно, однако, немало примеров плейотропии или плейотропного действия гена.
Оно состоит в том, что один структурный ген контролирует образование в организме нескольких или даже многих признаков (рис. 4.2). Достаточно вероятно, что плейотропное действие генов связано с участием продуктов их экспрессии (полипептиды) в целом спектре процессов жизнедеятельности (рис. 4.3).
Наблюдается тенденция расширения области использования исходно генетического понятия плейотропии в биологии. В основе названной тенденции лежат пришедшие, видимо, из химии представления о том, что практически химический (в мире жизни клеточно-биохимический, метаболический) процесс, кроме желательного (для живых форм,
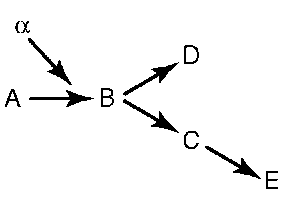
Рис. 4.2. Плейотропный эффект гена. Зависимость формирования нескольких признаков от функций продукта экспрессии гена. Нарушение реакции А-В, катализируемой белком-ферментом, в результате мутации гена ведет к формированию признаков D и Е
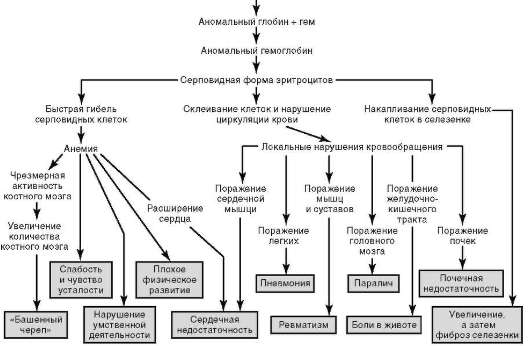
Рис. 4.3. Плейотропный эффект замены одной аминокислоты в β-глобине гемоглобина человека, проявляющийся клинически в виде серповидноклеточной анемии
биологически целесообразного) результата, нередко дает побочные нежелательные эффекты, за которыми закрепилось название параме-таболических. Примером неблагоприятного с биологической точки зрения параметаболического эффекта (альтернативный плейотропный эффект) может служить неизбежное образование активных форм кислорода (с их разрушительным действием на биополимеры и клеточные структуры) в связи с биологически бесспорно целесообразными процессами окислительного фосфорилирования в митохондриях. В па-раметаболических процессах, порождающих в биологических системах антагонистические (разнонаправленные, биологически целесообразные и одновременно и неизбежно биологически вредные) альтернативные плейотропные эффекты, видят, в частности, ведущее звено старения высших многоклеточных животных.
Наряду с плейотропией, при которой одна генотипическая причина дает несколько фенотипических эффектов, существует понятие генетической гетерогенности, когда разные генотипические при-
чины приводят к одному фенотипическому эффекту. Так, заболевание «наследственная полинейропатия Дежерина-Сотта», фенотипически характеризующееся врожденной демиелинизацией периферических нервов, возникает в случае мутаций в разных генах - PMP22 (хромосома 17), Po (хромосома 1), EGR2 (хромосома 10) и PRX (хромосома 19). Конечно, все названные гены имеют отношение к образованию и структурному оформлению миелина и оболочек периферических нервов.
В практике МГК важное место принадлежит понятиям «геноко-пии» и «фенокопии». Феномен генокопирования проявляется в том, что сходный фенотипический результат может быть обусловлен изменениями в разных генах или различными мутациями одного гена (явление генетической гетерогенности, см. здесь же, выше). Феномен фенокопирования состоит в том, что состояние признака, сходное с фенотипическими последствиями определенных мутаций, обусловлено не изменениями наследственных структур, а зависит от условий среды (генотипической или 1-го порядка, 2-го и 3-го порядков, см. здесь же, ниже), в которых происходит реализация генотипа в фенотип.
Во-вторых, ген имеет корпускулярную природу и характеризуется дискретностью действия, т. е. представляет собой в структурном и функционально-генетическом отношении отдельность. В силу дискретности генов возможны, с одной стороны, независимое наследование признаков, а с другой - генные или точковые мутации, затрагивающие в своем фенотипическом выражении отдельно взятые признаки (но: см. явление плейотропии - здесь же, выше). Корпускулярная природа и свойство дискретности генов составляют основу механизма независимого комбинирования признаков родителей в фенотипе потомства, т. е. их независимое друг от друга наследование (см. табл. 4.3).
В-третьих, гены как функционально-генетические единицы характеризуются дозированностью действия. Если фенотипический признак имеет количественное выражение, то его количество обычно пропорционально числу доминантных аллелей гена (см. п. 4.3.1.2). Так, содержание витамина А в исходно триплоидных клетках эндосперма растений пропорционально количеству доминантных аллелей соответствующего гена и убывает в ряду генотипов AAA, ААа, Ааа и ааа. Тем не менее эволюция создала механизм полигенного наследования количественных признаков с участием нескольких или даже многих генов, характеризующихся аддивностью, т. е. суммированием действия (см. п. 4.3.3.1).
Для характеристики степени выраженности признака или вероятности его проявления в фенотипе организма при наличии в генотипе соответствующего гена в генетике используют понятия экспрессивности и пенетрантности. Экспрессивность - это степень выраженности рассматриваемого признака в процентах по отношению к его максимальной выраженности среди всех особей с данным генотипом. Пене-трантность - это доля особей в процентах, у которых рассматриваемый признак проявился хотя бы в незначительной степени по отношению ко всем особям с данным генотипом. Конечно, показатели пенетрантности и экспрессивности генов зависят от разрешающей способности и точности применяемых методов регистрации (детекции) признаков. Вместе с тем на проявление гена в признак оказывают влияние факторы геноти-пической среды, внутренней среды организма и внешней или окружающей организм среды.
Сказанное обращает внимание на то, что в генетике нельзя использовать обобщенное понятие среды. Во-первых, речь может идти о ге-нотипической среде (относительно гена - среда 1-го порядка), т. е. о всей совокупности генов, представленных конкретными аллелями (см. п. 4.3.1.2) в генотипе данной особи. Приведенное определение геноти-пической среды дает возможность остановиться на различиях между такими генетическими понятиями, как «генотип» и «геном». В современной генетике очевидна тенденция относить первое понятие к генетической конституции отдельно взятого организма или индивидуума, тогда как второе - к генетическому «багажу» вида. Во-вторых, это может быть внутренняя среда организма данной особи (относительно гена - среда 2-го порядка; если речь идет о развитии плода, резонно выделять среду 2а и 2б порядков, имея в виду внутреннюю среду развивающегося и материнского организмов соответственно). В-третьих, это может быть окружающая среда жизни данной особи (относительно гена - среда 3-го порядка).
4.3.1.2. Аллельное состояние генов. Формы взаимодействия аллельных генов
Гены характеризуются свойством аллельного состояния. По-существу, аллели гена - это его альтернативные (по фенотипическому проявлению) формы. В классической генетике аллели так и определяли как альтернативные фенотипические состояния известного признака у жизнеспособных особей, имея в виду, что за признаком стоит наслед-
ственный задаток (ген). В настоящее время аллели - это варианты нуклеотидной последовательности участка молекулы ДНК, соответствующего, например, структурному (смысловому, кодирующему, транскрибируемому и транслируемому, экспрессируемому) гену. Количество альтернативных форм (аллелей) от гена к гену варьирует. Минимальное их число равно двум. У широко используемого в фундаментальной и экспериментальной генетике биологического объекта плодовой мухи (дрозофила) ген окраски глаз имеет порядка 1400 аллелей - множественный аллелизм. Ген, определяющий группу крови человека в системе АБО, имеет 3 аллеля (см. п. 4.1.1 и здесь же, ниже), в системе резус (Rh) - 2.
Между аллельными вариантами гена существуют функционально-генетические отношения, определяемые как формы взаимодействия аллельных генов. Типичные или наиболее частые варианты отношений (форм взаимодействия) - доминирование (от лат. dominus - господствующий), рецессивность (от лат. recessus - отступающий), кодомини-рование, неполное доминирование (промежуточное наследование - см. табл. 4.3). Характер межаллельных отношений проявляется в фенотипах диплоидных (эукариотических) организмов, для которых известны состояния гомозиготности и гетерозиготности (по парам аутосом и паре половых хромосом гомогаметного пола, у человека - женский), гемизиготности (по паре половых хромосом гетерогаметного пола, у человека - мужской). У человека это идентифицируется путем анализа появления в ряду поколений потомков того или иного фенотипического варианта признака (метод родословных генетического анализа людей, см. п. 5.2.2.1).
Доминантные признаки воспроизводятся в каждом поколении, т. е. у гомозиготных, гетерозиготных и гемизиготных по соответствующему гену (локусу, сайту ДНК) организмов. Рецессивный вариант признака наблюдается при отсутствии доминантного аллеля (рецессивная гомозигота и гемизигота). Такой вариант обнаруживается не в каждом поколении, а в случае гемизиготности - как правило, только у особей гетерогаметного пола. При кодоминировании у гетерозигот оба аллеля в равной мере участвуют в определении варианта признака, тогда как при неполном доминировании наличие рецессивного алле-ля у гетерозигот препятствует фенотипическому проявлению доминантного аллеля в полном объеме (дозовый эффект, сравни генотипы АА, Аа и аа). Кодоминирование аллелей у людей наблюдается в наследовании групп крови АБО. Группы крови 0(I), A(II), B(III) и AB(IV) определя-
ются геном I, имеющим три аллеля - IA, IB и I0. Аллель I0 относительно аллелей IA и IB проявляет свойство рецессивности. Аллели IA и IB кодо-минантны, чем и объясняется наличие группы крови IAIB или AB(IV). Неполное доминирование наблюдается у людей-гетерозигот по алле-лю серповидноклеточности эритроцитов (мутация ΗΒα2β26 ГлуВал), 60-65% гемоглобина которых имеет нормальную структуру, а 35-40% функционально дефектную, мутантную - в полипептиде β в 6-м положении аминокислота глутаминовая заменена на аминокислоту валин. Такие субъекты жизнеспособны и чувствуют себя комфортно за исключением ситуаций повышенной физической активности, в условиях высокогорья, при полетах на больших высотах или в холодное время года, когда в связи с развитием в организме состояния кислородной недостаточности они ощущают боли в суставах, в области сердца и селезенки.
Свойства доминантности и рецессивности аллелей (признаков) носят относительный характер - «неустойчивая доминантность», что зависит от ряда факторов, природа и механизмы которых не всегда понятны. Такой признак, к примеру, как эпикант («третье веко») проявляет свойство рецессивности у представителей европеоидной (кавказской) расы, но ведет себя как доминантный у представителей монголоидной расы. Отсутствие волос на голове (облысение) проявляет свойства рецессивного признака у женщин и доминантного у мужчин. То, что у женщин один из побочных эффектов применения в терапевтических целях мужского полового гормона тестостерона заключается в потере волос, указывает на участие гормонов в феноти-пическом проявлении этого гена. Факторы относительности свойства доминантности могут иметь как генетическую, так и негенетическую природу. Установленные факторы генетической природы - характер взаимодействия неаллельных генов и локализация аллеля в хромосоме - эффект положения (особенности генотипической среды, см. п. 4.3.1.1). С другой стороны, известно, что характер доминирования зависит от пола и возраста организма (особенности среды 2-го порядка, см. п. 4.3.1.1), а также внешних условий (особенности среды 3-го порядка, см. п. 4.3.1.1).
При множественном аллелизме обычно один аллель серии является рецессивным относительно всех остальных, тогда как другие связаны отношениями «доминантность-рецессивность», «неполное доминирование» или «кодоминирование». В серии аллелей гена окраски глаз дрозофилы абсолютно рецессивным является аллель white (белый), в системе групп крови АВ0 человека - I0. Аллель серии, наиболее распро-
страненный в природе (обычно соответствует нормальному состоянию фенотипического признака), называется «аллель дикого типа».
К сравнительно редким формам взаимодействия аллельных генов относятся межаллельная комплементация (взаимодополнение) и ал-лельное исключение. О межаллельной комплементации говорят тогда, когда у организма, гетерозиготного по двум мутантным аллелям конкретного гена, в фенотипе обнаруживается признак в нормальном, т. е. наиболее часто встречаемом состоянии («дикий тип»). Допустим, что ген D контролирует образование клеткой белка с четвертичной структурой в виде комплекса из нескольких одинаковых полипептидов - мультигомобелковый комплекс. Один мутантный аллель D определяет экспрессию измененного полипептида D', а второй аллель D" - тоже мутантный, но по другому участку гена, определяет экспрессию мутантного полипептида D", причем с другим изменением аминокислотной последовательности. Допускается, что при формировании четвертичной структуры с участием измененных, но по-разному, полипептидов D'" и D" происходит компенсация изменений, и в итоге формируется сложный мультигомобелковый комплекс с нормальной функцией.
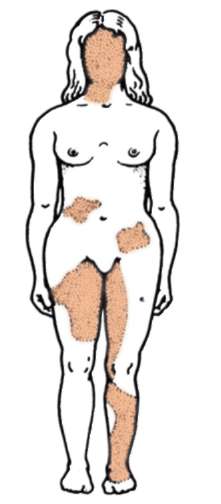
Суть аллельного исключения поясняет пример генетической инактивации одной из хромосом Х у особей гомогаметного пола (у человека - женский, 46ХХ), функционально-генетический смысл которой заключается в компенсации дозы генов соответствующей группы сцепления (хромосомы Х) относительно гетерогаметного пола (у человека - мужской, 46ХY). По хромосоме Х гомогаметный пол представлен особями-«мозаиками» - одна хромосома Х материнского, тогда как другая отцовского происхождения. Так как генетическая инактивация носит относительно хромосом материнского и отцовского происхождения случайный характер, при гетерозиготности организма в одних клетках активен аллель, полученный с хромосомой Х матери, тогда как в других активен аллель, полученный с хромосомой Х отца. Это приводит к фенотипическому мозаицизму (рис. 4.4).
Рис. 4.4. Мозаицизм женского организма (кариотип 46,ХХ) по наличию или отсутствию потовых желез в коже, обусловленный экспрессией нормального или мутантного аллеля гена хромосомы Х. Затемнены участки кожи, лишенные потовых желез, в клетках которых экспрессирует-ся мутантный аллель
4.3.1.3. Изменения нуклеотидных последовательностей ДНК. Генные
Неустраненные и/или неисправленные (см. п. 2.4.5.3-a) изменения химической структуры генов (сайтов, нуклеотидных последовательностей ДНК), воспроизводимые в последующих циклах репликации и проявляющиеся у потомков в виде измененных вариантов признака, называют генными мутациями.
Такие изменения можно подразделить на три группы. Мутации первой группы заключаются в замене одного нуклеотида на другой. Напомним, что нуклеотиды, из которых состоят макромолекулы ДНК, различаются по азотистому основанию, что дает право рассматривать главное событие генных мутаций первой группы как замену одного азотистого основания на другое. На их долю приходится порядка 20% спонтанно (самопроизвольно, без видимой причины) случающихся генных изменений.
Вторая группа мутаций обусловлена сдвигом «рамки считывания», что является следствием изменения числа пар нуклеотидов в пределах гена как в сторону уменьшения (делеция - потеря участка гена), так и увеличения (дупликация - удвоение участка гена). Причиной сдвига «рамки считывания» может стать встраивание (инсерция) в ген участка молекулы ДНК, в частности, вирусной природы (МГЭ, или транспозоны) или из другой хромосомы - транслокация (см.
пп. 1.4.6, 2.4.3.4-д, 2.4.3.4-е, 3.1.4).
мутации
Мутации третьей группы связаны с изменением порядка следования нуклеотидов в пределах гена (инверсия).
Названные варианты генных мутаций в функционально-генетическом отношении отвечают принципу «все или ничего», т. е. мутация либо произошла и проявилась в фенотипе (возможно, через поколения), либо нет. Они случаются как в смысловых (экзоны), так и иных (интроны, области промоторов, энхансеров или сайленсеров, сервисные, регуля-торные или конценсусные сайты, 5' и 3' транскрибируемые, но не транслируемые участки транскриптона) нуклеотидных последовательностях ДНК. При типах мутаций, описанных выше, фенотипические эффекты наблюдаются при изменении в молекуле ДНК одного нуклеотида или в биспирали ДНК 1 п.н. Это позволяет в качестве элементарной единицы мутационного процесса (мутон) считать отдельно взятый нуклеотид или пару нуклеотидов.
Относительно недавно стали выделять еще одну группу генных мутаций на основе общности молекулярного механизма, состоящего в прогрессивном росте числа (экспансия), в основном тринуклеотидных тандемных повторов в регуляторной (нетранслируемой) или смысловой (транслируемой) частях генов. В функционально-генетическом плане эти мутации относят к категории «динамических», поскольку фенотипический эффект они дают после того, как количество повторов достигнет и превысит определенный критический минимум. Состояние, при котором в гене есть нуклеотидные тандемные повторы, но число их меньше критического, рассматривают как «премутацию». Наличие последней делает (ген)аллель-носитель нестабильным, что способствует переходу «премутации» в полную мутацию. Для рассматриваемой категории мутаций характерно явление антиципации, т. е. утяжеления клинических проявлений и более раннего начала заболевания в ряду поколений в пределах одной родословной в связи с ростом числа повторов (нарушение принципа «все или ничего»). Наиболее известны тринуклеотидные повторы, хотя описаны и другие формы, например двенадцатинуклеотидные и даже более. Предположительно к мутациям по типу экспансии тринуклеотидных повторов ведет нарушение функции фермента ДНК-полимеразы в последовательных мейотических и митотических циклах. Элементарной единицей (мутон) мутагенеза такого рода на генном уровне является триплет нуклеотидов или последовательность из 3 п.н. в биспирали ДНК, а не отдельный нуклеотид или 1 п.н. (генные мутации по типу замены нуклеотидов или сдвига «рамки считывания»).
Мутации по типу замены нуклеотидов происходят в силу разных причин. Одна из них заключается в том, что под влиянием определенных химических агентов или без видимой физико-химической причины изменяется азотистое основание нуклеотида, уже включенного в молекулу ДНК. Если такое искажение молекулярной структуры ДНК не устраняется механизмами молекулярной репарации (см. п. 2.4.5.3-а), то в ближайшем цикле репликации к измененному ну-клеотиду присоединится нуклеотид, комплементарный именно ему, а не тому, который занимал соответствующее место в молекуле ДНК до изменения. В итоге возникает новая пара нуклеотидов, что приводит к искажению биоинформации в рассматриваемом участке биспи-рали ДНК. Так, вследствие дезаминирования цитозина цитидиловый нуклеотид в паре Ц-Г превращается в уридиловый, комплементарным которому является адениловый нуклеотид. В силу отмеченного при ближайшей репликации образуется пара У-А (рис. 4.5, I), а при следующей - возникает биспираль с парой А-Т вместо пары Ц-Г. Замена пары Ц-Г на пару А-Т происходит также в том случае, если цитозин оказывается метилированным по 5-му углеродному атому. Дезамини-руемый 5-метилцитозин превращается в тимин (рис. 4.5, II), который в ближайшем репликационном цикле дает пару с аденином. Известны примеры включения в строящуюся цепь ДНК нуклеотида с химически измененным азотистым основанием или его аналогом. Если ошибка не обнаруживается, то участие ошибочно включенного «неправильного» нуклеотида в последующих репликационных циклах приводит к замене в соответствующих участках двойной спирали ДНК нормальной пары нуклеотидов на другую, что сопряжено с искажением биоинформации. Так, к адениловому нуклеотиду материнской цепи ДНК может присоединиться нуклеотид не с тимином, а с 5-бромурацилом (5-БУ). При репликации нуклеотид с 5-БУ обычно присоединяет не аденило-вый, а гуаниловый нуклеотид. В следующем репликационном цикле гуаниловый нуклеотид образует пару с цитидиловым. В итоге пара А-Т заменяется парой Г-Ц (рис. 4.6).
Из приведенных примеров видно, что замены нуклеотидов в ДНК происходят до или в процессе репликации первоначально в одной по-линуклеотидной цепи. Если эти изменения не исправляются, то в ходе последующих репликаций они становятся достоянием обеих полину-клеотидных цепей биспирали ДНК. Из сказанного следует, что важным источником генных мутаций по типу замены нуклеотидов являются нарушения процессов репликации и репарации ДНК.
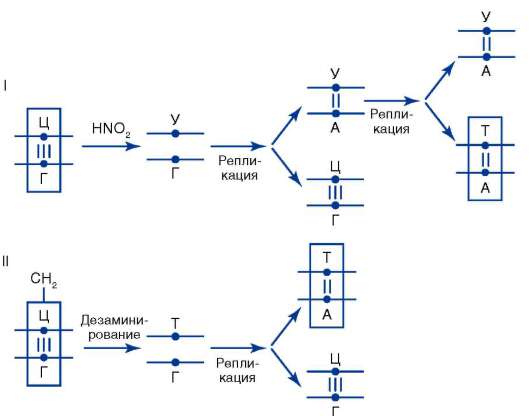
Рис. 4.5. Мутации по типу замены основания (дезаминирование азотистых оснований в молекуле ДНК): I - превращение цитозина в урацил вследствие де-заминирования, замена пары Ц-Г на пару Т-А; II - превращение метилцитозина в тимин, замена пары Ц-Г на пару Т-А
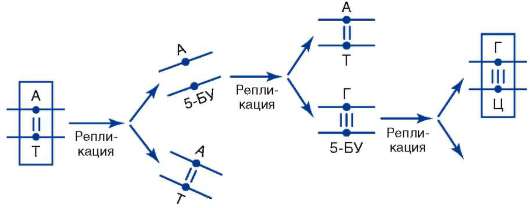
Рис. 4.6. Мутация по типу замены основания (ошибочное включение химического аналога тимина 5-бромурацила при репликации ДНК)
Следствием замены одного нуклеотида в макромолекуле или 1 п.н. в биспирали ДНК является образование нового триплета в нуклеотидной последовательности, кодирующей последовательность аминокислот в полипептиде. В силу вырожденности генетического кода в 25% таких замен возникает триплет-синоним, что не дает изменений аминокислотной последовательности в соответствующем полипептиде. 2-3% замен ведут к образованию триплетов-терминаторов (стоп-кодонов), что фенотипически проявляется в трансляции на мутантных и(м)РНК укороченных полипептидов. Еще один вариант генных мутаций по типу замены нуклеотидов приводит к появлению триплетов, шифрующих другие аминокислоты, которые, однако, характеризуются сходными физико-химическими свойствами, являясь, например, также гидрофобными. Это хотя и ведет к изменению аминокислотного состава полипептида, тем не менее не вызывает резкого изменения его характеристик (см. п. 2.4.5.3-а). Таким образом, генные мутации с полномасштабным фенотипическим эффектом составляют порядка 70-75% всех изменений макромолекулярной структуры ДНК, связанных с нуклеотидными заменами. В качестве примера приведем изменение в гене β-глобина аллеля А («дикий тип») гемоглобина человека на аллель серповидно-клеточности эритроцитов S (мутантный). Фенотипические проявления мутации, ведущее положение среди которых занимает болезнь «серпо-видноклеточная анемия», многообразны (см. рис. 4.3). Суть мутации сводится к замене второго нуклеотида (Т) в триплетах, кодирующих стоящую в β-полипептиде на 6-м месте глутаминовую кислоту (ЦТТ или ЦТЦ), на нуклеотид (А), превращающих их в триплеты, кодирующие аминокислоту валин (ЦАТ или ЦАЦ).
Генные мутации по типу сдвига «рамки считывания» составляют немалую часть спонтанных (самопроизвольных, случающихся без очевидной причины) мутаций. Их число возрастает при действии некоторых химических соединений, в частности акридиновых. Выпадение нуклеотидных пар (делеция) на достаточно протяженных участках макромолекул ДНК типично для мутаций под действием рентгеновских лучей. У плодовой мухи, например, имеется мутация по типу делеции, вызываемая рентгеновским облучением и фенотипически проявляющаяся в изменении окраски глаз, при которой ген, ответственный за цвет глаз, теряет порядка 100 п.н. Действуя на фаг Т4 химическим веществом профлавином, вызывают мутации, состоящие как в выпадении, так и во вставках нуклеотидных пар. При этом фенотипический эффект наблюдается, если в биспирали ДНК появляется или ею теряется всего 1 п.н.
Значительное количество вставок объясняется встраиванием в ДНК МГЭ (транспозонов).
С определенной вероятностью вставки и выпадения п.н. происходят вследствие ошибок рекомбинации, например, при неравном кроссинго-вере (рис. 4.7). Если вследствие неравного кроссинговера в ген встраивается фрагмент псевдогена, говорят о генной конверсии. Такой вариант отражает суть большинства известных мутаций гена фермента 21-гидроксилазы, приводящих к развитию у человека адреногениталь-ного синдрома (врожденная гиперплазия коры надпочечников).
К выпадению или вставкам пар нуклеотидов приводят достаточно частые мутации в сайтах сплайсинга - на границе интронов и экзонов. Эти мутации проявляются в нарушении акта вырезания интронов из пре-и(м)РНК транскрипта. В результате созревающая и(м)РНК лишается части или всего экзона. Еще один возможный вариант - сохранение в и(м)РНК интронной нуклеотидной последовательности.
Считывание информации с ДНК - непрерывный процесс, т. е. ген транскрибируется одним блоком с началом в точке инициации и завершением в точке терминации (транскрибируемые, но не транслируемые 5'- и 3'-участки транскриптона в данном случае в расчет не принимаются, см. п. 2.4.5.5). Учитывая сказанное, а также свойство непере-
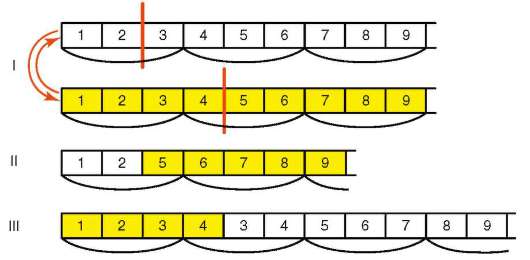
Рис. 4.7. Мутация со сдвигом «рамки считывания» вследствие неравноценного обмена наследственным материалом при внутригенном кроссинговере: I - разрывы аллельных генов в разных участках и обмен фрагментами между ними; II - выпадение 3-й и 4-й пар нуклеотидов, приводящее к сдвигу «рамки считывания»; III - удвоение 3-й и 4-й пар нуклеотидов, сдвиг «рамки считывания»
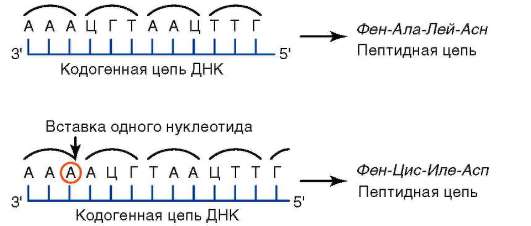
Рис. 4.8. Результат изменения числа нуклеотидных пар в биспирали ДНК. Сдвиг «рамки считывания» вследствие вставки одного нуклеотида в кодогенную цепь нуклеиновой кислоты приводит к изменению аминокислотного состава соответствующего полипептида
крываемости генетического кода (см. п. 2.4.5.2), становится понятным, почему выпадения или вставки нуклеотидных пар, сдвигая «рамку считывания», ведут к изменению содержания генетической информации и, таким образом, представляют собой истинные мутации (рис. 4.8). Иногда следствием сдвига «рамки считывания» становится образование стоп-кодона, что приводит к синтезу укороченного полипетида. Если из биспирали ДНК теряются или в ней появляются дополнительно пары нуклеотидов, количество которых кратно трем, то сдвига «рамки считывания» не происходит (свойство триплетности генетического кода, см. п. 2.4.5.2). На уровне трансляции в таких случаях в образуемых полипептидах соответственно теряются или приобретаются дополнительные аминокислотные остатки.
Мутации по типу изменения положения определенного числа пар нуклеотидов в макромолекуле ДНК происходят вследствие поворота участка нуклеиновой кислоты на 180° (инверсия). Обычно этому предшествует образование соответствующим участком ДНК петли, в пределах которой репликация происходит в направлении, обратном «правильному». На уровне трансляции это проявляется в частичном изменении порядка следования аминокислотных остатков в полипептиде, что меняет его функциональные свойства.
Мутации по типу экспансии нуклеотидных повторов, так же как и другие варианты генных мутаций, случаются как в транслируемых (информативных - экзоны), так и в нетранслируемых (неинформативных - интроны) частях генов, что накладывает свой отпечаток на фенотипи-
ческие проявления. Так, экспансия тринуклеотида (триплета) ЦАГ, кодирующего аминокислоту глутамин, в транслируемой части генов до 40-80 повторов, не нарушая процессов транскрипции и трансляции, приводит к появлению в молекуле полипептида «трека» из соответствующего количества глутаминовых аминокислотных остатков. Такой увеличенный в размерах белок функционально дефектен. Мутации описанного типа лежат в основе развития наследственных нейродегене-ративных патологий, в частности хореи Гентингтона («пляска святого Витта» - одним из ведущих клинических фенотипических проявлений является гиперкинез).
Если мутация локализуется в нетранслируемой части гена, то количество, например, ЦГГ-повторов, соответствующее пороговому значению, исчисляется сотнями и тысячами. Клинико-фенотипические проявления мутаций такого типа разнообразны: синдром Мартина-Белла (ломкая хромосома Х) с классической триадой признаков - олигофрения, дисморфия (нарушения процессов морфогенеза в онтогенезе), макроор-хидизм.
Свои особенности имеют мутации в ДНК митохондрий (мтДНК или хромосома М), что в немалой степени объясняется отличиями в структуре как отдельных генов, так и всего генома названных органелл. Так, митохондриальные гены лишены интронов, а большинство транскрибируемых и(м)РНК лишены 5' и 3' нетранслируемых участков (см. п. 2.4.5.5). В сравнении с ядерным геномом митохондриальный геном характеризуется большей плотностью расположения генов в связи с меньшим содержанием межгенной ДНК. Специфика отличает репликацию мтДНК, которая происходит в два этапа. Триплет АУА в мито-хондриальных и(м)РНК, в отличие от образуемых на ядерных генах, кодирует не изолейцин, а метионин, триплет УГА не выполняет функции стоп-кодона, шифруя аминокислоту триптофан, триплеты АГА и АГГ являются стоп-кодонами.
Митохондриальный геном содержит 22 гена для «собственных» тРНК, 2 - для «собственных» рРНК и 13 - для полипептидов, входящих в 5 надмолекулярных комплексов дыхательных цепей органеллы. Предположительно, 1100-1150 генов, участвующих в биоинформационном обеспечении структуры и функций митохондрий, находятся в ядерном геноме. Соотносительный вклад генов ядерной и митохондри-альной локализации в биоинформационное обеспечение функционирования дыхательных комплексов митохондрий иллюстрирует табл. 4.1. Таких комплексов, представляющих собой мультигетеробелковые об-
разования, пять (I-V). Каждый из них представлен совокупностью белковых субъединиц (полипептидов, простых белков или протеинов). Одна часть субъединиц образуется непосредственно в органеллах под контролем митохондриальных генов (мтДНК), тогда как другая - в цитоплазме клетки под контролем ядерных генов (яДНК). Названные комплексы решают различные специфические задачи в рамках процесса окислительного фосфорилирования (дыхательный обмен, аэробный синтез АТФ, см. п. 2.4.6.1).
Таблица 4.1. Биоинформационное обеспечение функционирования митохон-дриальной дыхательной цепи. Взаимодействие ядерного и митохондриального геномов
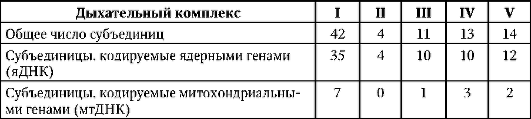
Все митохондрии эукариотической клетки наследуются по материнской линии, т. е. через цитоплазму яйцеклетки. За редким исключением (зрелые эритроциты млекопитающих и человека) каждая клетка содержит десятки и сотни копий мтДНК.
С учетом сказанного, митохондриальные болезни, в патогенезе которых ведущая роль принадлежит мутационным изменениям, принято делить на 3 группы. Это фенотипические патологические проявления, обусловливаемые, во-первых, мутациями ядерных генов, во-вторых, изменениями непосредственно в мтДНК и, в-третьих, нарушением так называемых межгеномных сигнальных эффектов. В первом случае развиваются митохондриальные болезни с аутосомно-доминантным или аутосомно-рецессивным типом наследования (классическое моногенное менделевское наследование). Характерным проявлением поражений соответствующих сайтов ДНК как ядерной, так и митохон-дриальной локализации (делеции, точковые генные мутации, делеции в сочетании с дупликациями) является пониженный уровень энергоснабжения тканей и органов. Нарушения межгеномных сигнальных эффектов связаны с мутациями ядерных генов-регуляторов. Фенотипически они могут проявляться в изменении количества копий митохондриаль-ной ДНК - деплеция (истощение) митохондриального генетического аппарата - и приводить к тканеспецифическим делециям или дуплика-
циям мтДНК. Типичным представляется присутствие в клетке одновременно митохондрий с мутировавшей мтДНК и «генетически здоровых» органелл со случайным от клетки к клетке количественным соотношением, что определяет вариабильность клинической картины. Вклад в фенотипическую изменчивость, особенно в этих случаях, вносит различная чувствительность клеток разных тканей и структур организма к кислородной недостаточности.
Относительно высокая «поражаемость» мтДНК объясняется тем, что в процессе наработки энергии в органелле (т. е. в непосредственном окружении ДНК органеллы) закономерно образуются АФК (см. п. 2.4.8), а также тем, что митохондрии отличаются более низкой, в сравнении с клеточными ядрами, эффективностью механизмов репарации макромолекул нуклеиновой кислоты в случае их повреждения.
Представления о мутационном процессе (мутагенез), которые были сформулированы в основном классической (домолекулярной) генетикой, уточняются, дополняются и переосмысливаются в свете научных данных современной (в том числе молекулярной) генетики, геномики, протеомики и метаболомики, цитомики и клеточной биологии (см. п. 1.1). Действительно, изменения нуклеотидных последовательностей ДНК происходят не только в биоинформационных участках (экзоны) смысловых транскрибируемых и транслируемых генов, но и в области интронов, промоторов и энхансеров, в сайтах, кодирующих транскрипционные и ростовые факторы, цитокины и рецепторы к ним, в участках, представленных в геномах избыточной ДНК и нуклеотидными повторами разного формата, имеющими различную макромолекулярную организацию (см. п. 2.4.3.4-в, 2.4.3.4-д, 2.4.5.5). В связи с появлением нового семейства «динамических» генных мутаций приходится вносить уточнения в представления об элементарной единице мутагенеза (мутон) на генном уровне структурно-функциональной организации генетического аппарата эукариот (см. здесь же, выше).
В соответствии с представлениями классической генетики частота генных (точковых) спонтанных (самопроизвольных, случающихся без видимой причины - см. п. 4.3.1.4) мутаций у всех живых форм составляет в среднем 10-5-10-7 изменений на один локус нуклеиновой кислоты (в другой редакции - на одну гамету) за поколение. Приводимые значения носят ориентировочный характер. Известно, например, что в геномах живых форм, включая человека, имеются локусы (сайты, нуклеотидные последовательности ДНК), различающиеся по интенсивности спонтанного мутагенеза на 1-3 порядка. В связи с этим предлагается считать,
что у людей частота спонтанно возникающих генных мутаций выражается цифрой 10-11 для наиболее устойчивых участков генома, тогда как для высокомутабильных сайтов («горячие» точки мутагенеза) - 10-4 изменений на локус (гамету) за поколение.
4.3.1.4. Функционально-генетическая классификация генных мутаций
Генные мутации классифицируют по ряду оснований.
Большинство изменений макромолекулярной структуры генов фено-типически неблагоприятно (классификация по влиянию на жизнеспособность и/или плодовитость особей) - вредные генные мутации. Среди них выделяют летальные и полулетальные мутации. Первые несовместимы с жизнью в принципе, вторые ограничивают жизнеспособность организма настолько, что он, как правило, не способен достичь возраста половой (репродуктивной, биологической) зрелости, принять участие в размножении и, таким образом, передать свои гены (аллели) организмам следующего поколения.
Закономерен вопрос, почему вновь возникающие мутации обычно вредны. Здесь не следует забывать, что структурно-функциональная организация геномов клеток и организмов носит системный характер. С одной стороны, мутационные изменения закономерны, т. е. они происходят у всех живых форм без исключения регулярно с частотой в среднем 10-5-10-7 мутаций на один локус за поколение. С другой - мутационные изменения случайны в том смысле, что практически невозможно предсказать, когда, какой ген и с какими биоинформационными (фенотипическими) последствиями мутирует. Важно, однако, то, что мутируют гены, встроенные в систему функционально взаимодействующих и взаимовлияющих генов (нуклеотидных последовательностей, сайтов ДНК). В таких условиях каждая мутация, чтобы не нести в себе неблагоприятные фенотипические последствия, должна с момента своего возникновения удовлетворять «правилам», по которым существует система генома (см. п. 4.3.1.1, генотипическая или среда 1-го порядка).
Редко случаются изменения генов с благоприятными фенотипиче-скими последствиями - полезные генные мутации. Известны нейтральные генные мутации, не сказывающиеся на жизнеспособности и репродуктивном потенциале.
Большинство вновь возникающих мутаций (классификация по проявлению в гетерозиготном состоянии) дает рецессивный аллель, кото-
рый, будучи по своим фенотипическим последствиям обычно вредным, у диплоидных эукариот на некоторое время укрыт от действия естественного отбора в гетерозиготах. Предположительно именно это сыграло ведущую роль в формировании резерва наследственной изменчивости. Реже аллели, образующиеся вследствие мутации, проявляют свойства доминантности или кодоминирования (см. п. 4.3.1.2).
Генетики начала и середины ХХ в. выделяли спонтанные (самопроизвольные, случающиеся без видимой причины) и индуцированные (вызываемые факторами известной природы - химические соединения, ионизирующее излучение, биологические агенты, в частности вирусы) мутации - классификация по происхождению. На настоящий момент актуальность приведенной классификации, с одной стороны, несколько снизилась в связи с тем, что многое стало известно о природе факторов спонтанного мутагенеза - активные формы кислорода, ионизирующее излучение космического происхождения, внутриклеточные тепловые колебания. С другой стороны, в связи с появлением значительного количества производимых промышленностью мутагенов, в число которых входят удобрения, инсектициды и пестициды, лекарства, средства борьбы с бытовыми насекомыми и другие химические вещества, а также все более широко используемые в быту приборы и устройства, эксплуатация которых связана с электромагнитными и другого рода излучениями, осознается необходимость мониторинга присутствия и концентрации в среде жизни людей факторов, индуцирующих мутагенез.
Различают также мутации прямые (классификация по направлению), которые переводят аллель «дикого типа» в мутантный аллель, и обратные (реверсии), возвращающие мутантный аллель в алелль «дикого типа», биохимические, морфологические, физиологические, поведенческие и др. (классификация по фенотипическому проявлению), цитоплазматические (митохондриальные, в клетках растений - также пластидные) и ядерные (классификация по локализации в клетке изменяемого генетического материала).
Принципиально деление мутаций на генеративные, случающиеся в половых клетках, и соматические, затрагивающие генетический аппарат соматических клеток (классификация по месту возникновения и характеру наследования). Мутации различного ранга (генные, хромосомные, геномные), возникающие в соматических клетках, наследуются потомками этих клеток, что делает организм генотипическим мозаи-ком, т. е. особью со смешанными клеточными популяциями, которые содержат как генетически нормальные, так и мутировавшие клетки.
Хотя классическая генетика, учитывавшая интересы биологов-эволюционистов, а также в связи с задачами МГК, в большей мере ориентировалась на генеративные мутации, в настоящее время именно соматические мутации представляют приоритетный интерес, например, для онкологии (см. п. 3.1.4).
4.3.1.5. Биологическое значение генного уровня организации генетического аппарата
Свойство дискретности генетического материала (см. п. 4.2, Г. Мендель) подразумевает делимость этого материала на отдельности (корпускулярный характер) - гены, которые служат элементарными функционально-генетическими единицами, т. е. обеспечивают возможность наследования и изменений признаков организма порознь, их независимое комбинирование в фенотипах особей.
Благодаря наличию генного уровня стал возможен научно-экспериментальный анализ закономерностей наследования и изменения отдельных признаков и их ассоциаций, была вскрыта химическая природа и описаны макромолекулярные и надмолекулярные свойства непосредственного носителя генетических функций - ДНК (см. п. 4.2, О.Т. Эйвери; Дж. Уотсон, Ф. Крик и М. Уилкинс; Х.Г. Корана и М.У. Ни-ренберг), установлен ряд законов наследственности и изменчивости, сформулированы представления о гено(аллело)фондах популяций организмов, понято значение их изменений для процесса исторического развития (эволюция), в частности видообразования (см. п. 4.2, С.С. Четвериков).
4.3.2. ХРОМОСОМНЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ГЕНЕТИЧЕСКОГО АППАРАТА
На хромосомном уровне структурно-функциональной организации генетического аппарата решается ряд специфических, только этому уровню присущих задач проявления свойств наследственности и изменчивости в процессах жизнедеятельности, в размножении, в индивидуальном и историческом развитии эукариот. Во-первых, хромосомный принцип построения генетического материала решает задачу разделения возросшего при переходе к эукариотическому типу клеточной организации количества ДНК на отдельные части с целью оптимизации репликации ДНК и передачи без количественных и качественных потерь биоинформации в ряду поколений клеток и организ-
мов, размножающихся половым путем (см. митоз и мейоз). Во-вторых, распределение генов (сайтов, нуклеотидных последовательностей ДНК разной функционально-генетической направленности) между хромосомами создает предпосылки для их сцепленного наследования (см. п. 4.3.2.1). В-третьих, в эволюции переход к распределению ДНК клетки между хромосомами совпадает с изменением формы молекул ДНК с кольцевой на линейную. Это создает новые возможности в решении задачи регуляции генетических функций. Уместно вспомнить механизм «спирализация-деспирализация» не только целых геномов, но отдельных хромосом (дозовая компенсация по генам хромосомы Х гомогамет-ного пола относительно гетерогаметного пола, у человека - женского), а также их участков (см. п. 2.4.3.4-в). Линейность молекул ДНК способствует тому, что генетический материал эукариотической клетки исходно существует в репрессированном состоянии благодаря комплексу ДНК с белками щелочного характера (гистоны). То, что названный комплекс имеет нуклеосомную организацию, создает дополнительные возможности тонкой регуляции генетических функций ДНК. В-четвертых, речь идет об использовании биоинформации частями благодаря механизмам регуляции с участием белков кислого характера - негистоновых, к семейству которых относится большинство транскрипционных факторов. Новейшие данные свидетельствуют о большой роли в регуляции генетических функций на макромолекулярном уровне белок-белковых взаимодействий. На этой основе функционирует, в частности, инициа-торный мультигетеробелковый комплекс, запускающий с промотора транскриптона процесс транскрипции генов в эукариотических клетках (см. п. 2.4.5.5). В-пятых, важная роль в функционировании структурных (экспрессируемых, транскрибируемых и транслируемых) генов, принадлежит закономерному пространственному взаиморасположению хромосом в клеточном ядре. Предположительно механизмом, который ведет к этому, является взаимодействие теломерных (см. п. 2.4.3.4-г) участков хромосом со структурами ядерного матрикса (см. п. 2.4.3.2) и ядерной ламиной (см. п. 2.4.3.1).
Специфический вклад хромосомного уровня структурно-функциональной организации генетического аппарата в изменчивость заключается, с одной стороны, в том, что независимая комбинация негомологичных хромосом отцовского и материнского происхождения в анафазе первого деления мейоза при образовании половых клеток представляет собой эффективный механизм комбинативной геноти-пической изменчивости. С другой стороны, уместно вспомнить, что
диплоидный набор у человека представлен 46 хромосомами, тогда как у шимпанзе - 48 хромосомами. Сравнительный анализ кариотипов показывает, что хромосома 2 людей произошла, видимо, в результате слияния двух мелких акроцентрических хромосом обезьяноподобного предка. Во всяком случае два плеча хромосомы 2 человека соответствуют по нуклеотидным последовательностям двум разным хромосомам современных человекообразных обезьян: 12 и 13 - шимпанзе, 13 и 14 - гориллы и орангутан(г)а. Человеческая хромосома 9 длиннее соответствующей хромосомы шимпанзе, а хромосома 12 короче, на хромосомах 1 и 18 людей имеются протяженные инверсии в сравнении с одноименными хромосомами шимпанзе. Зарегистрирован также ряд перицентрических инверсий (хромосомы 4, 5, 12 и 17), не меняющих генный состав хромосом (групп сцепления), но, возможно, создающих эффект положения. Наибольшие различия между человеком и шимпанзе касаются не структурных генов, а хромосом.
Хромосомные перестройки (мутации) вследствие нарушения мейо-за способны сразу привести к репродуктивной изоляции, которая согласно современным представлениям является необходимым условием процесса видообразования.
4.3.2.1. Хромосомная теория наследственности. Основные положения
Хромосомы как особые ядерные структуры открыты во второй половине XIX в. (см. п. 4.2, В. Вальдейер). В это же время было высказано предположение о том, что они имеют отношение к явлению наследственности (см. п. 4.2, А. Вейсман). В начале XX в. предположение А. Вейсма-на нашло убедительное подтверждение (см. п. 4.2, Т. Бовери, У. Сеттон), а генетика как наука благодаря результатам исследований Т.Г. Моргана и его коллег, обобщенным в виде хромосомной теории наследственности, обогатилась новыми сведениями о закономерностях наследования признаков (см. п. 4.2, Т.Г. Морган).
Согласно этой теории, гены («наследственные задатки», по Г. Менделю) расположены в ядерных структурах - хромосомах (но: см. митохондриальный геном). Каждая хромосома характеризуется уникальным и постоянным генным составом. В хромосоме гены располагаются друг за другом линейно. Гены одной хромосомы нередко передаются (наследуются) в ряду поколений сцеплено друг с другом, что послужило основанием рассматривать каждую хромосому как отдельную группу сцепления генов. Сила сцепления, проявляющаяся
в возможности генов хромосомы наследоваться совместно, для разных генов группы сцепления варьирует, и тем меньше, чем больше расстояние между ними в хромосоме.
Если иметь в виду, что при образовании половых клеток в профазе первого деления мейоза гомологичные хромосомы обмениваются участками (рекомбинация путем кроссинговера), причем частота таких обменов для каждой пары генов (локусов) - величина постоянная и может быть представлена в процентах половых клеток с признаками прошедшего кроссинговера (кроссоверные гаметы, см. п. 4.3.5.2), указанное генетическое явление можно использовать для определения порядка расположения генов по длине хромосомы, а также расстояния между генами, т. е. для составления генетических карт хромосом. Расстояние между парой генов на генетической карте выражается в морганидах - М (в зарубежной генетической литературе более часто используют термин сантиморганиды - сМ). Если два гена расположены в хромосоме на расстоянии в одну морганиду (сантиморганиду), это означает, что кроссинговер между ними случается (т. е. эти гены рекомбинируют) в 1% делений клетки. Гены одной группы сцепления, если они располагаются в хромосоме на расстоянии в 50 морганид (сан-тиморганид) и более, наследуются независимо друг от друга. Благодаря новым методическим возможностям в настоящее время осуществляется составление физических карт хромосом, характеризующихся в сравнении с генетическими картами большей разрешающей способностью. Расстояние между анализируемыми сайтами ДНК (генами) при физическом картировании выражается в парах нуклеотидов. Генетическое расстояние в 1 М(сМ) соответствует физической дистанции примерно в 1 млн п.н. или 1 мегабазе - Мб (Mb). Составление полных физических карт хромосом является главной целью проекта «Геном человека».
Хромосомная теория не противоречит представлениям о корпускулярной природе генов и вытекающим из этого правилам (закономерностям) независимого наследования признаков, установленным Г. Менделем. Вместе с тем она указывает на существование в природе совместного или сцепленного наследования, вытекающего из факта расположения генов в хромосомах.
При этом необходимо различать сцепленное наследование как таковое и наследование, сцепленное с полом. В первом случае речь идет о совместной передаче в процессе полового размножения от родителей потомству генов, располагающихся в одной группе сцепления (в хромосоме, в том числе в аутосоме). Во втором случае речь идет о наследо-
вании потомством генов, располагающихся в половых хромосомах, у человека - Х и Y.
4.3.2.2. Изменения структурной организации хромосом. Хромосомные мутации
Несмотря на эволюционно отработанный механизм сохранения постоянной физико-химической и морфологической организации хромосом в ряду клеточных поколений, эта организация может изменяться. В основе изменения структуры хромосом, как правило, лежат первоначальные изменения их целостности - разрывы, приводящие к разного рода перестройкам. Хромосомные перестройки называются хромосомными мутациями или хромосомными аберрациями.
С одной стороны, разрывы происходят закономерно в мейозе в связи с кроссинговером и сопровождаются обменом взаимосоответствующими участками между гомологичными хромосомами. Нарушения хода кроссинговера, приводящие к обмену количественно неравнозначными участками наследственного материала (ДНК), приводит к образованию новых по генному составу групп сцепления, характеризующихся либо утратой (делеция), либо удвоением (дупликация) определенных сайтов (нуклеотидных последовательностей, генов). С другой стороны, разрывы хромосом могут вызываться воздействием на них мутагенов. Наиболее часто в роли мутагенов выступают физические факторы (ионизирующие излучения), химические соединения, вирусы. Иногда нарушение структурной целостности хромосомы сопровождается поворотом участка между двумя разрывами на 180° с последующим встраиванием этого участка в хромосому - инверсия. В зависимости от того, включает ли инвертируемый участок центромеру или нет, различают соответственно перицентрические и парацентрические инверсии. Если участок, отделившийся от хромосомы вследствие ее разрыва, лишен центромеры, он может быть утрачен клеткой при очередном митозе. Нередко, однако, такой участок прикрепляется к другой хромосоме - транслокация. Часто две поврежденные негомологичные хромосомы обмениваются отделившимися от них участками - реци-прокная транслокация. Если оторвавшийся участок присоединяется к своей же хромосоме, но в новом месте, говорят о транспозиции (рис. 4.9). Известны примеры транслокаций целых хромосом. Так, синдром Дауна имеет несколько цитогенетических форм. У одной части пациентов с этим синдромом определяются три отдельных хромосомы 21,
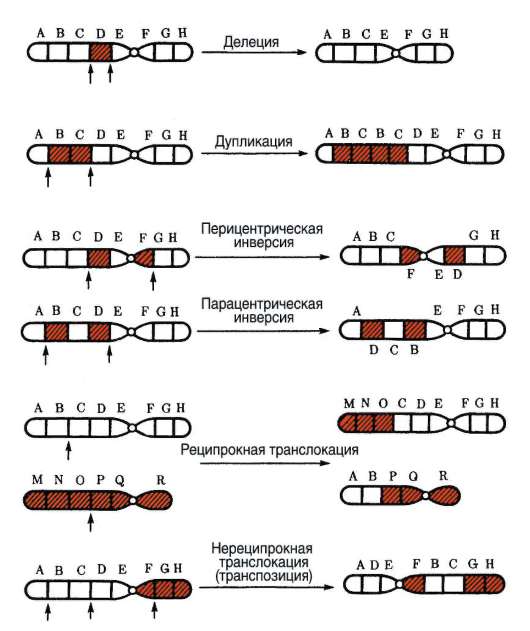
Рис. 4.9. Виды хромосомных перестроек
у другой части «лишняя» хромосома 21 транслоцирована на другую хромосому (такая хромосома приобретает необычно большие размеры и изменяет форму, см. рис. 4.24).
Очевидно, что инверсии и транслокации ведут к изменению локализации соответствующих нуклеотидных последовательностей (генов, сайтов).
Хромосомные аберрации (мутации, перестройки) обычно проявляются в изменении морфологии хромосом, что можно наблюдать с помощью микроскопа (цитогенетический метод генетического анализа). Метацентрические хромосомы становятся субметацентрическими и/или акроцентрическими и, наоборот, возникают кольцевые и полицентрические хромосомы (рис. 4.10, 4.11). Особая категория хромосомных мутаций - аберрации, связанные с центрическим слиянием или разделением хромосом. В таких случаях две негомологичные хромосомы «объединяются» в одну - робертсоновская транслокация, или из одной хромосомы образуются две самостоятельных (рис. 4.12). При мутациях описанного типа появляются хромосомы с новой морфологией, может изменяться число хромосом в кариотипе.
Хромосомные мутации обычно сопровождаются изменениями в генетической программе, наследуемой дочерними клетками после деления материнской. При делециях и дупликациях нарушается количество соответствующих сайтов (генов) в сторону уменьшения или увеличения, тогда как при инверсиях, транспозициях и транслокациях меняют-
Рис. 4.10. Изменение формы хромосом вследствие перицентрических инверсий
Рис. 4.11. Образование кольцевых (I) и полицентрических (II) хромосом
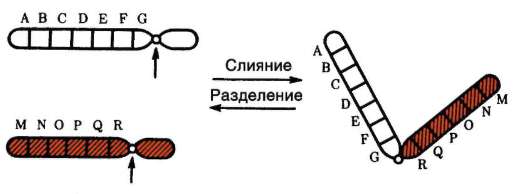
Рис. 4.12. Хромосомные перестройки, связанные с центрическим слиянием или разделением хромосом. Являются причиной изменения числа хромосом в ка-риотипе
ся либо условия и, таким образом, характер функционирования в связи с изменением взаиморасположения нуклеотидных последовательностей (генов, сайтов) в хромосоме, либо состав групп сцепления. Чаще структурные перестройки хромосом соматических клеток сказываются
на их жизнеспособности отрицательно (соматические хромосомные
мутации). Нередко такие перестройки указывают на возможность ма-лигнизации. Серьезные последствия имеют хромосомные аберрации в клетках-предшественницах половых клеток (генеративные хромосомные мутации), что нередко сопровождается нарушением конъюгации гомологичных хромосом и их нерасхождением в дочерние клетки в мейозе. Делеции и дупликации участка одной из гомологичных хромосом сопровождаются при конъюгации образованием гомологом петли с количественно неравноценным наследственным материалом (рис. 4.13). Реципрокные транслокации между двумя негомологичными хромосомами приводят при конъюгации к возникновению не бивалента, а квадривалента с образованием благодаря взаимному притягиванию гомологичных участков, расположенных в разных хромосомах, фигуры креста (рис. 4.14). Участие в реципрокных транслокациях не двух, а большего числа хромосом с возникновением уже не квадривалента, а поливалента приводит к формированию при конъюгации более сложных структур (рис. 4.15). При инверсиях бивалент, возникающий в профазе I мейоза, образует петлю, включающую взаимно инвертированный участок (рис. 4.16).
Конъюгация и последующее расхождение структур, образованных измененными хромосомами, способствуют появлению новых хромосомных перестроек. В результате гаметы, получая неполноценный наследственный материал, не способны обеспечить нормальное развитие особи нового поколения.
Несмотря на неблагоприятные в целом последствия генеративных хромосомных мутаций, в тех случаях, когда они оказываются совместимыми с развитием и жизнью организма, такие мутации через эволюцию
Рис. 4.13. Петля, образующаяся при конъюгации гомологичных хромосом, которые несут неравноценный наследственный материал в соответствующих участках вследствие хромосомной аберрации
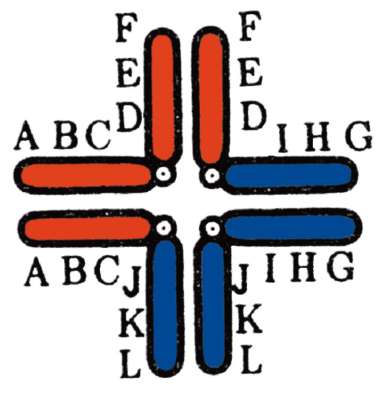
Рис. 4.14. Образование при конъюгации квадривалента из двух пар хромосом, несущих ре-ципрокную транслокацию
Рис. 4.15. Образование при конъюгации поливалента шестью парами хромосом, участвующих в реципрокных транслокациях: I - конъюгация между парой хромосом, не несущих транслокацию; II - поливалент, образуемый шестью парами хромосом, участвующих в транслокации
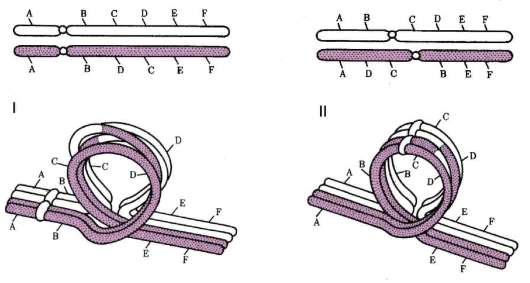
Рис. 4.16. Конъюгация хромосом при инверсиях: I - парацентрическая инверсия в одном из гомологов; II - перицентрическая инверсия в одном из гомологов
структуры хромосом эффективно способствуют биологической эволюции (видообразованию). Даже делеции, если они незначительны по размерам, сохраняются в гетерозиготном состоянии в ряду поколений. Менее вредны, в сравнении с делециями, дупликации, хотя, если увеличение количества наследственного материала значительно (10% и более), организм, как правило, нежизнеспособен. Робертсоновские транслокации обычно совместимы с жизнью в силу того, что они не связаны с изменениями количества наследственного материала. Это, видимо, было «использовано» в интересах эволюции. О вероятности этого говорят различия числа хромосом в клетках организмов близкородственных видов, объясняемые слиянием или разделением хромосом. Так, у разных видов плодовых мух (дрозофила) количество хромосом в гаплоидных наборах варьирует от 3 до 6. О возможной роли хромосомных перестроек на уровне обезьяноподобного предка в эволюции человека см. п. 4.3.2.
4.3.2.3. Биологическое значение хромосомного уровня организации генетического аппарата
Хромосомный уровень структурно-функциональной организации генетического аппарата эукариотической клетки вносит свой специфический вклад в функционирование генов, характер наследования признаков потомками, регуляцию генетической активности.
Хромосомы как комплексы генов (сайтов, нуклеотидных последовательностей ДНК) представляют собой эволюционно сложившиеся структуры, обязательные для клеток всех особей данного вида. Немаловажная роль в особенностях функционирования генов принадлежит их взаимному расположению в хромосомах (в группах сцепления). Локализация гена в той или иной хромосоме, в частности в аутосоме, половых хромосомах Х или Y, хромосоме М, определяет закономерности (правила) наследования соответствующего признака. Принадлежность генов к одной хромосоме обусловливает то более, то менее выраженный сцепленный характер наследования контролируемых ими признаков, тогда как расстояние между генами в хромосоме влияет на частоту рекомбинации указанных признаков в фенотипе потомства (см. п. 4.3.2.1). Напротив, локализация генов в разных хромосомах или в одной хромосоме, но на расстоянии в 50 морганид (сантиморганид) и более составляет основу независимого наследования признаков.
Вклад хромосомного уровня структурно-функциональной организации генетического аппарата эукариот в регуляцию генетической
активности ДНК - см. п. 4.3.2. Устойчивое комплексирование ДНК с основными белками-гистонами делает ее недоступной для взаимодействия с белками - обязательными участниками процесса транскрипции, например с РНК-полимеразой, транскрипционными факторами, энхан-серами. Предварительная декомпактизация хроматина также, видимо, является условием транскрипции биоинформации с соответствующего участка ДНК.
Возникновение в эволюции генетического аппарата при переходе к эукариотическому типу клеточной организации хромосомного уровня связано с возросшим количеством генетического материала. Распределение ядерной ДНК между ограниченным числом структур - хромосом - обусловливает упорядоченную пространственную организацию всей массы генетического материала по группам сцепления. Относительная самостоятельность хромосом в процессах редупликации и распределения между дочерними клетками в митозе и мейозе позволяет закономерно воспроизводить и передавать в ряду поколений без количественных потерь и качественных изменений (за редкими исключениями - см. п. 4.1.1, генотипическая изменчивость) значительные объемы жизненно важной биоинформации. Распределение генов (сайтов, конкретных по функционально-генетическим характеристикам нуклеотидных последовательностей ДНК) по группам сцепления делает возможной рекомбинацию генетического материала и, следовательно, соответствующей биоинформации, заключенной в гомологичных и негомологичных хромосомах, при мейозе и оплодотворении. Такая рекомбинация существенно увеличивает уровень генотипической комбинативной изменчивости, что само по себе является важным эволюционным фактором, обеспечивающим в каждом поколении наличие биоинформационно разнообразного наследственного материала и, следовательно, фенотипических вариантов для естественного отбора.
4.3.3. ГЕНОМНЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ГЕНЕТИЧЕСКОГО АППАРАТА
Геном определяется как совокупность наследственного материала (генов, сайтов, нуклеотидных последовательностей ДНК) в гаплоидном наборе хромосом клеток организмов соответствующего биологического вида. Геном видоспецифичен, поскольку он отождествляется с полным набором генов, необходимых для формирования видовых характеристик организмов в процессе индивидуального развития (онтогенез).
Размер генома в определенной степени коррелирует с положением группы, к которой принадлежит соответствующий вид, в иерархической системе мира жизни. Так, размер генома прокариот не превышает 8х106 п.н., у дрожжей (одноклеточные эукариоты) этот размер равен 1,35х107 п.н., т. е. на порядок выше, у шпорцевой лягушки (Xenopus laevis), так же как у высших позвоночных (млекопитающие), на 2 порядка выше, чем у дрожжей, - 109 п.н. (см. также п. 1.5). Данные по геному человека см. в п. 2.4.3.4-д.
Большинство видов высших организмов диплоидны, т. е. в их соматических клетках одновременно представлено два генома. Один из них материнского, второй - отцовского происхождения (см. п. 4.3, генотип). В природе известны виды, представители которых способны размножаться партеногенезом - при отсутствии оплодотворения. В таких случаях, благодаря включению особого механизма, соматические клетки развивающегося зародыша диплоидны. Обычно оба генома, присутствующие в их ядрах, имеют материнское происхождение. В экспериментальных условиях получены зиготы, имеющие цитоплазму яйцеклетки и два генома отцовского происхождения. На вопрос о возможности партеногенетического развития человека в настоящее время наука дает отрицательный ответ. Дело в том, что при отсутствии материнского генома (диандрогенез) образуются только провизорные структуры, но не внутренняя клеточная масса, клетки которой, собственно, и дают новый организм. Наоборот, при отсутствии отцовского генома (дигиногенез) провизорные структуры не образуются. Формируется лишь внутренняя клеточная масса. В обоих случаях зародыш нежизнеспособен.
Для некоторых видов организмов на временной (в онтогенезе) основе возможны гаплоидные особи. Так, у определенных видов насекомых пол определяется диили гаплоидностью особи. У пчел и муравьев, например, самцы, развивающиеся из неоплодотворенных яйцеклеток, первично гаплоидны. В дальнейшем их соматические клетки становятся диплоидными. При определенных внешних воздействиях на яйцеклетки некоторых видов организмов (низкие температуры, икра тритонов) в эксперименте удавалось получать гаплоидных животных, отличавшихся, однако, пониженной жизнеспособностью. В исключительных случаях гаплоидные особи тритонов появляются в природных условиях, возможно, вследствие резких температурных колебаний в период оплодотворения.
Отмечено, в частности у человека, появление на свет полиплоидных (три- и тетраплоидных, т. е. 3n и 4n соответственно) организмов, которые отличались сниженной жизнеспособностью: ни один из новорожденных полиплоидов человека не прожил более двух недель после рождения. Описаны, в том числе среди взрослых людей, особи-мозаики, в органах и тканях которых, наряду с диплоидными клетками, присутствуют клетки более высокой плоидности.
В приведенных выше примерах обращает на себя внимание факт изменения в клетках числа полных геномов. Это обстоятельство - веский аргумент в пользу выделения у эукариот, наряду с генным и хромосомным, также геномного уровня организации генетического аппарата.
Значение указанного уровня для наследственности состоит, во-первых, в том, что геном представляет собой не случайный, а сбалансированный качественно (по биоинформационному наполнению) и количественно (по дозам структурных генов и сайтов ДНК с другими функциями) объем генетической информации. Будучи эволюционно «проработанной» и отобранной, эта биоинформация необходима и одновременно достаточна для обеспечения присущего особям конкретного биологического вида типа индивидуального развития (онтогенез) и обмена веществ (метаболизм). Во-вторых, эффективное решение названной задачи допускает наличие некодирующих (в том числе транскрибируемых, но не транслируемых) нуклеотидных последовательностей с приобретением ими регуляторных, сервисных, конценсусных и иных функций. В-третьих, оказалась возможной кластерная организация генов, контролирующих жизненно важные клеточные и организменные функции - с одной стороны, кластеры генов рРНК, тРНК, гистонов, с другой, кластеры α- и β-глобиновых генов. Кластерная организация генов в приведенных примерах позволяет решать разные функционально-генетические задачи (см. п. 2.4.3.3 и 4.3.3.2). В-четвертых, отдельные хромосомы являются группами сцепления генов. Учитывая корпускулярную природу генов, это обусловливает как независимое, так и сцепленное (частично или полностью) наследование, а также различные комбинации (сочетания) аллелей и признаков. Следствием последнего является формирование в каждом поколении уникальных по аллельно-му составу генотипов и, следовательно, появление различающихся по фенотипам потомков.
Присутствие в геномах эукариот избыточной ДНК, возможно, было использовано в процессе исторического развития (эволюция) для создания в случае необходимости «новых» структурных генов (транс-
крибируемых и транслируемых, экспрессируемых сайтов ДНК) или нуклеотидных последовательностей с регуляторной, сервисной и иными функциями. С геномным уровнем организации генетического аппарата эукариот связывают также переход к возможности избирательной транскрипции генов по времени (период онтогенеза), месту (тип клеток) и объему. В связи с отмеченным вспомним о смене форм β-полипептида гемоглобина в ряду «эмбрион-плод-родившийся человек». При этом происходит не только смена активного (экспрессируемого, транскрибируемого и транслируемого) гена β-глобинового кластера (см. п. 2.4.3.4-д), но и типа клеток, в которых соответствующий ген экспрес-сируется: стенка желточного мешка эмбриона, печень плода, красный костный мозг родившегося человека. Молекула гемоглобина образована полипептидами α и β, которые находятся в строго определенном количественном соотношении: на каждые 2 полипептида α приходится 2 полипептида β. Очевидно, что транскрипция генов названных полипептидов должна быть в количественном плане согласована, что и имеет место.
4.3.3.1. Формы взаимодействия неаллельных генов
Большинство признаков и свойств живого существа являются сложными. Их развитие почти всегда представляет собой результат взаимодействия ряда генов, причем неаллельных, т. е. располагающихся на разных хромосомах или на одной хромосоме, но в разных локусах. Классической (домолекулярный и догеномный периоды) генетикой были определены такие формы взаимодействия неаллельных генов, как полимерия (полимерное наследование), комплементарное взаимодействие и эпистаз - рецессивный и доминантный. Формы взаимодействия аллельных генов - см. п. 4.3.1.2.
Развитие количественных признаков нередко обусловлено полимерными генами или полигенами - системой неаллельных генов, в равной мере влияющих на формирование соответствующего признака и характеризующихся аддитивным (суммирующим) действием. Соответственно степень выраженности признака зависит от совокупной дозы доминантных аллелей всех полимерных генов системы. С известной долей осторожности можно сказать, что полимерное наследование - один из генетических механизмов, оказывающих влияние на экспрессивность количественного признака. По полимерному типу у людей наследуется, в частности, интенсивность окраски кожных покровов, зависящая от количества откладываемого в цитоплазме пигментных клеток (меланоци-
ты) черного пигмента меланина. По общепринятому мнению, в геноме человека за пигментацию кожи ответственна система из 4 генов. Учитывая диплоидность соматических клеток, каждый из этих генов представлен в генотипе двойной дозой. Так как они участвуют в определении степени развития одного признака, их обозначают одной буквой, но с разными цифровыми индексами: Р1, Р2, Р3 и Р4 - доминантные аллели или р1, р2, р3, р4 - рецессивные аллели. Наличие в генотипе индивидуума восьми доминантных аллелей рассматриваемых полигенов (Р1Р1 Р2Р2 Р3Р3 Р4Р4) обусловливает максимальную пигментацию кожных покровов, отличающую африканских негров. Гомозиготы по рецессивным аллелям четырех генов характеризуются минимальным уровнем пигментации, отличающим представителей европеоидной расы. Большее или меньшее число доминантных аллелей - от 8 до 0 - определяет разную интенсивность окраски кожных покровов (рис. 4.17). Полигенное наследование характеризует у человека такие признаки, как рост и масса тела, возможно, интеллект. В последнем случае предполагается соучастие большого количества генов - несколько десятков или более.
Если формирование сложного признака требует совокупного действия (взаимодействия) аллелей конкретных неаллельных генов, то, очевидно, он будет проявляться в фенотипе лишь тех особей, которые в генотипах имеют именно требуемую комбинацию аллелей. По существу, речь идет о том, что в результате совокупного действия возникает новое качество, которого не было бы при отсутствии соответствующего взаимодействия. Указанное взаимодействие получило название комплементарного. В качестве примера рассмотрим процесс формирова-
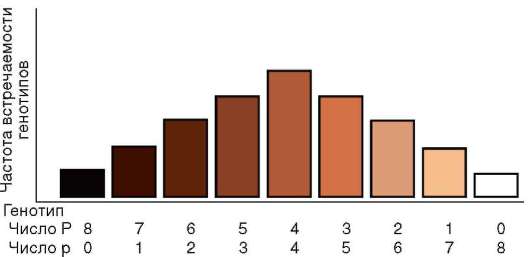
Рис. 4.17. Зависимость пигментации кожных покровов у человека от числа в генотипе доминантных аллелей системы полигенов Р
ния половой принадлежности у людей. Известно, что у вида H. sapiens комплекс половых признаков определяется сочетанием в кариотипе половых хромосом (гетерохромосом) Х и Y. Установлено, однако, что для развития мужского фенотипа недостаточно нуклеотидных последовательностей ДНК хромосомы Y, определяющих дифференцировку половых желез по мужскому типу и образование гормона тестостерона. Необходим также продукт экспрессии другого гена (находится на хромосоме Х), обусловливающего образование белка-рецептора к тестостерону и располагающегося в клеточных оболочках. Отсутствие синтеза рецептора, например, вследствие мутации делает ткани-мишени невосприимчивыми к мужскому половому гормону. В результате развивается в целом женский фенотип, однако такой человек лишен детородной функции и наделен рядом физических (метаболические и функциональные параметры скелетной мускулатуры) и характерологических (решительность, импульсивность в действиях) черт, более присущих мужскому полу. Описанное состояние известно как синдром тестикуляр-ной феминизации Морриса (рис. 4.18).
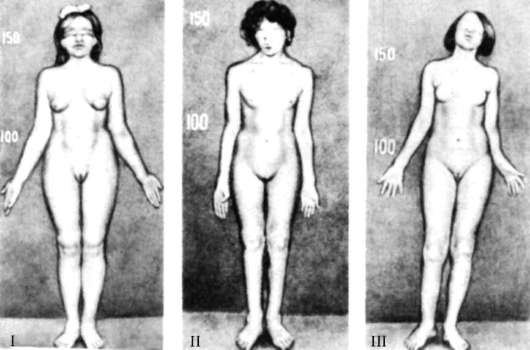
Рис. 4.18. Тестикулярная феминизация - синдром Морриса: I - кариотип 46XY (семенники удалены в детском возрасте); II - евнухоидная форма, кариотип 46XY (отсутствие молочных желез, вторичного оволосения, естественного влагалища); III - кариотип 46XY
Еще одна форма взаимодействия неаллельных генов - эпистаз. В качестве примера рассмотрим ситуацию, связанную с формированием принадлежности человека к определенной группе крови в системе АВ0 (см. также пп. 4.1.1 и 4.3.1.2). Отвечающий за развитие рассматриваемого признака ген I в виде аллелей IA и IB обеспечивает образование организмом белков с антигенными свойствами А и В, располагающихся на поверхности эритроцитов. Для того чтобы названные антигены были синтезированы, требуется вещество-предшественник, образование которого детерминируется геном Н. Доминантный аллель гена Н присутствует в генотипах подавляющего большинства людей (генотипы НН или Hh). У редких, но все-таки встречающихся среди людей рецессивных гомозигот (hh) вещество-предшественник не образуется, вследствие чего у таких лиц, даже при наличии в генотипах аллелей IA и/ или IB, антигены А и В не синтезируются, и по фенотипу они относятся к субъектам с группой крови I, а не II, III или IV. Приведенный пример известен под названием «бомбейский феномен» и соответствует генетическому понятию рецессивного эпистаза. С позиций рецессивного эпистаза следует рассматривать случаи альбинизма у людей, когда даже у представителей негроидной расы, которые имеют максимальное число доминантных аллелей в системе полигенов, определяющих интенсивность пигментации кожных покровов, радужной оболочки глаз и волос, рождаются альбиносы с полным отсутствием в клетках организма пигмента меланина (рис. 4.19), который не может образоваться в принципе. Генетика располагает также примерами, когда некий ген, например В, обусловливает развитие соответствующего признака только в том случае, если другой (неаллельный гену В) ген, например А, представлен в генотипе двумя рецессивными аллелями (аа). При наличии доминантного аллеля названного гена (генотипы АА или Аа) признак В не формируется. Приве-

Рис. 4.19. Пример рождения альбиноса в семье негра и негритянки (разно-яйцовые близнецы - девочка и мальчик, - один из которых альбинос)
денная ситуация соответствует генетическому понятию доминантного эпистаза.
С известной долей осторожности можно предполагать, что от наличия эпистатической формы взаимодействия неаллельных генов зависит такое свойство, как пенетрантность.
Как уже отмечалось, рассмотренные формы взаимодействия неал-лельных генов в виде полимерного наследования, комплементарного взаимодействия и эпистаза описаны представителями классической (до-молекулярной) генетики. Они были открыты путем анализа вариантов наследования определенных фенотипических признаков потомством и относятся, по существу, к генному уровню организации генетического аппарата эукариот. Расшифровка химической природы генетического материала (ДНК) и непосредственная работа с ним методами молекулярной генетики (см. п. 5.2.2.3-в), осознание системного принципа функционирования аппарата наследственности и биологической изменчивости, появление в генетическом обиходе понятия «геном» (см. п. 4.3.3.2) способствовали расширению представлений о формах взаимодействия различных нуклеотидных последовательностей или сайтов ДНК в процессе формирования фенотипа особи. Вспомним, к примеру, о взаимодействии элементов промотора и сайтов с энхансерной функцией (см. п. 2.4.5.5). Особое внимание в настоящее время уделяется нуклео-тидным последовательностям, выполняющим регуляторные функции относительно структурных (экспрессируемые, информативные, транскрибируемые и транслируемые) генов.
4.3.3.2. Функционально-генетическая характеристика нуклеотидных последовательностей ДНК (сайтов, генов)
В классической (домолекулярной) функциональной генетике, когда методы исследования закономерностей наследования признаков и генетических законов ограничивались фактически гибридологическим (растения и животные), генеалогическим и близнецовым (человек) методами, а представления о материальной природе генов носили гипотетический характер, в функционально-генетическом плане речь могла идти исключительно о структурных генах, отличительной характеристикой которых было наличие соответствующих им признаков.
В современной генетической номенклатуре к категории структурных генов относят две разновидности нуклеотидных последовательностей (сайтов) ДНК. С одной стороны, это последовательности, которые транскрибируются с образованием и(м)РНК и затем транслируются
с образованием простых белков (полипептидов, протеинов). С другой стороны, к категории структурных генов причисляют также последовательности, которые транскрибируются с образованием определенных видов РНК - рибосомных и транспортных, необходимых для организации в клетках процесса биосинтеза белков. Ни рРНК, ни тРНК не транслируются. Таким образом, среди структурных генов выделяют транскрибируемые и транслируемые, а также транскрибируемые, но не транслируемые. Оказалось, однако, что на долю таких последовательностей приходится не более 5% общего количества ДНК генома (см. п. 2.4.3.4-д).
Достижения генетики последних десятилетий позволяют связать некоторую часть структурных генов (в частности, из числа транскрибируемых и транслируемых нуклеотидных последовательностей ДНК) с важнейшими событиями индивидуального развития и процессами жизнедеятельности, выделяя их в отдельные субсемейства. Так, выделяют структурные гены, напрямую связанные с обеспечением эмбриогенеза многоклеточных животных. К примеру, на ранних стадиях эмбрионального развития под генетическим контролем устанавливается своеобразная координатная сетка, позволяющая клеткам развивающегося организма определить себя в пространстве. Благодаря последовательно активируемым группам генов (материнские гены, gap и pair-rule гены, гены сегментарной полярности) у плодовых мух (дрозофилы) определяется головной и хвостовой концы, а тело зародыша подразделяется в переднезаднем направлении на фиксированное число сегментов, клетки которых дадут в дальнейшем конкретные структуры. Благодаря соответствующему генетическому контролю определяется также спинно-брюшное направление. В рассматриваемом отношении интересны гомеозисные гены, которые располагаются кластерами линейно друг за другом. Они контролируют развитие в требуемом направлении определенных клеточных популяций. При этом последовательность генов в кластере соответствует пространственному взаиморасположению возникающих в процессе развития структур организма - проксимодистальное и дорсовентральное направления.
Эмбриогенез в реальном пространственно-временном воплощении представляет собой единый процесс. Тем не менее выделение в этом процессе определенных стадий (дробление, бластула, гаструла, первичный органогенез или закладка осевых органов, вторичные или локальные органогенезы) оправдано хотя бы тем, что, как оказалось, они находятся под специальным генетическим контролем. Так, у мыши обнаружен
локус Т, представленный 117 аллелями. Аллель t проявляет свойство рецессивности относительно всех остальных аллелей. У гомозигот по некоторым аллелям локуса T эмбриональное развитие проходит нормально лишь до определенной стадии. По достижении этой стадии зародыш прекращает развитие и гибнет. Нежизнеспособны гомозиготы: t12t12 (не образуется трофобласт, гибель на стадии 16 бластомеров - мору-ла), tw73tw73 (в связи с функциональной несостоятельностью поверхности клеток хориона зародыш не имплантируется в стенку матки, гибель на стадии 64 бластомеров - бластоциста), t0t0 (не образуется амнион), tw5tw5 (возникнув, зародышевая эктодерма погибает), t9t9 (зародыш нежизнеспособен, так как нет мезодермы: не образуется первичная полоска - дефект эмбриональной индукции), tw1tw1 (дефект нейруляции - гибнут клетки вентральных отделов нервной трубки и закладки головного мозга). У гомозигот TT нервная трубка коротка и не достигает заднего конца тела зародыша, в связи с чем нарушается ход вторичных (локальных) органогенезов, и зародыш гибнет. Детальна также рецессивная гомозигота tt. Типичный фенотипический признак мышей с генотипом Tt - короткие хвосты.
У эукариот обнаружены структурные гены, которые можно выделить в субсемейство, члены которого решают задачи экологического характера или же обеспечивают соответствующий образ существования (например, паразитический). Выше говорилось (см. п. 4.3.3), что у человека на разных этапах онтогенеза синтезируются разные полипептиды α- и β-глобинов, благодаря чему образуются гемоглобины - эмбриональный, плодный и родившегося человека. Необходимость последовательной смены в онтогенезе названных вариантов гемоглобина диктуется изменениями условий снабжения тканей кислородом, т. е. экологическими обстоятельствами.
Паразитам высших животных, в том числе млекопитающих, отличающихся наличием эффективной иммунной системы, приходится решать специфическую задачу ухода от надзорной функции последней. Один из способов - особый генетический механизм. Так, в геноме внутриклеточного паразита человека, возбудителя африканского трипаносомоза (сонная болезнь) жгутикового простейшего трипаносомы (Trypanosoma brucei gambiense) имеется «кассета» генов, контролирующих образование белка клеточной оболочки паразита и имеющих некоторые отличия в нуклеотидных последовательностях. Уход трипаносом от противостояния иммунной системы организма-хозяина связан с регулярной сменой активного гена «кассеты». Результат заключается в
появлении в клеточной оболочке «нового» варианта белка-антигена, что заставляет иммунную систему выстраивать свои отношения с паразитом после каждой такой смены заново. Некоторые внутриклеточные паразиты человека (возбудитель трехдневной малярии Plasmodium vivax, возбудитель американского или городского трипаносомоза, известного так же, как болезнь Чагаса, Trypanosoma cruzi - см. п. 2.4.2) используют в качестве «ворот» для проникновения в клетки хозяина белки плазмолеммы, образуя под генетическим контролем соответствующие молекулы-лиганды.
В качестве самостоятельного субсемейства следует, по-видимому, выделить гены «социального» контроля. Речь здесь может идти о протоонкогенах (см. п. 3.1.4), которые обусловливают образование ферментов протеинкиназ, факторов роста и клеточных рецепторов к этим факторам, участвующих в регуляции клеточной пролиферации. Ряд синтезируемых под контролем названных генов веществ принимает участие в запуске и реализации генетически запрограммированной клеточной гибели - апоптоза (см. п. 3.1.2). Напомним, что механизм апоп-тоза сложился в эволюции в немалой степени в связи с необходимостью контроля клеточного состава многоклеточного организма и направлен в том числе против клеток, вышедших из-под влияния общеорганиз-менных регуляторных систем, например, вставших на путь онкотранс-формации (см. п. 2.2 - концепция «клеточного государства» Р. Вирхова;
см. п. 3.1.4).
Существует достаточно многичисленное субсемейство нуклеотидных последовательностей ДНК, которые принимают участие в регуляции генетической активности структурных (в частности, транскрибируемых и транслируемых) генов. В этом плане предположительно речь может идти о некоторых З'-транскрибируемых, но не транслируемых последовательностях ДНК транскриптона (см. п. 2.4.5.5), например, тех, с которыми связано образование в молекулах и(м)РНК поли(А) «хвоста». Действительно, дезаденилирование названных и(м)РНК используется, видимо, как механизм регуляции времени полураспада, т. е. продолжительности существования информационных РНК в цитоплазме. Регуляторная функция через образование контролируемых ими белков характеризует сайты ДНК, обусловливающие образование транскрипционных факторов (см. п. 2.4.5.5).
Определенные нуклеотидные последовательности ДНК используются для контроля важных общеклеточных функций. Так, теломер-ной ДНК приписывают роль, с одной стороны, таймера (биологические
часы), указывающего предел числа отпущенных клеткам многоклеточного организма делений - эффект Хейфлика, а с другой - эта ДНК предположительно обеспечивает путем взаимодействия с ядерной ламиной и внутриядерным матриксом упорядоченное расположение хромосом в объеме интерфазного ядра (см. п. 2.4.3.4-г).
Среди функций нуклеотидных последовательностей ДНК называют также сервисную и конценсусную.
В качестве примера сайтов с сервисной функцией укажем 5'-транскрибируемые, но не транслируемые последовательности области промотора транскриптона (см. п. 2.4.5.5), которые создают условия для инициации процесса транскрипции биологической информации с ДНК. Эта же функция выполняется, по-видимому, сайтами ДНК, контролирующими образование белков пререпликативного и репликативного комплексов, обеспечивающих самоудвоение биспирали ДНК (см. п. 2.4.5.3). Без определенных белков, образуемых клетками под контролем соответствующих нуклеотидных последовательностей ДНК (генов), выполняющих, видимо, также сервисную функцию, невозможен транспорт пре-и(м)РНК из ядра в цитоплазму в форме ядерных информосом (см.
п. 2.4.5.5).
Конценсусная функция приписывается нуклеотидным последовательностям на стыке интронов и экзонов, а также транскрибируемым, но не транслируемым сайтам, контролирующим образование малых ядерных и ядрышковых РНК - участниц процессинга пре-РНК транскриптов в качестве обязательного компонента сплайсосом (см.
п. 2.4.5.5).
Транскрибируемым, но не транслируемым нуклеотидным последовательностям ДНК 5' и 3' концов транскриптона, с которыми связано наличие в и(м)РНК соответственно колпачка (кэпа) и уже упоминавшегося поли(А) «хвоста», приписывают защитную функцию - предохранение и(м)РНК от разрушительного действия ферментов нуклеаз.
На настоящий момент для большей части нуклеотидных последовательностей ядерной ДНК функция однозначно не определена и здесь в обозримом будущем следует ожидать важных открытий (область интересов функциональной геномики).
4.3.3.3. Геномный уровень и биологическая изменчивость. Геномные мутации
Геномные мутации представлены двумя группами изменений в генетическом аппарате эукариот. Во-первых, это изменение числа геномов на клетку в сравнении с диплоидным или удвоенным как в сторону уменьшения до одинарного - гаплоидия, так и в сторону увеличения до трехкратного, четырехкратного и далее - полиплоидия. Примеры обоих вариантов геномных мутаций такого рода, а также их влияние на жизнеспособность особей-мутантов - см. п. 4.3.3. Эти примеры относятся к миру животных. В мире растений негативные последствия мутаций, связанных с изменением количества геномов, не столь очевидны. Напротив, селекционеры нередко индуцируют такие мутации, получая путем отбора среди мутантов новые сорта. Так, если сравнить количество генетического материала (ДНК) в клетках культивируемых сортов злаковых, например пшеницы, легко убедиться в различиях, которые в абсолютном выражении кратны величине, соответствующей одинарному (гаплоидному) набору хромосом, т. е. геному.
Геномные мутации, состоящие в изменении числа геномов, приводят к изменению дозы всех структурных генов и нуклеотидных последовательностей с другими (регуляторными, сервисными, конценсусными) функциями. Из приведенных примеров следует, что для индивидуального развития и жизнедеятельности неблагоприятны в одинаковой мере как уменьшение (гаплоидия), так и увеличение (полиплоидия) дозы генов в сравнении с удвоенной (диплоидия).
Во-вторых, к геномным мутациям относят изменения числа отдельных хромосом - анэуплоидия. Среди мутаций такого рода выделяют 2 варианта. С одной стороны, это уменьшение числа гомологичных хромосом конкретной пары в диплоидном наборе с двух до одной - мо-носомия, с другой - увеличение числа до трех и более - трисомия, полисомия. Геномные мутации, заключающиеся в изменении числа отдельных хромосом, приводят к нарушению генного баланса по той или иной группе сцепления (см. п. 4.3.2.1). Достоверно установлено, что полные моносомии по аутосомам нежизнеспособны. Во всяком случае среди родившихся людей организмы-моносомики по аутосомам не обнаружены. Напротив, жизнеспособные особи-моносомики по половым хромосомам известны. Так, у людей с синдромом Шерешевского-Тернера (рис. 4.20) утрачена одна из половых хромосом - кариотип 45ХО. Именно по хромосомам Х и Y возможна полисомия. В частности, среди людей обнаружены три-, тетра-, пентасомики по хромосоме Х -
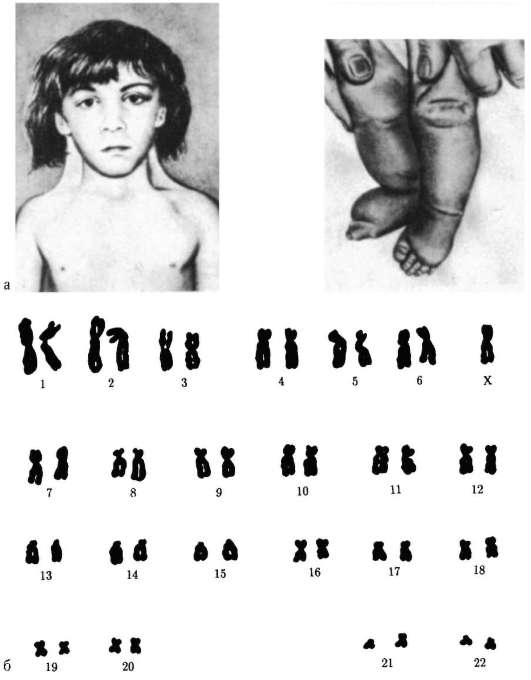
Рис. 4.20. Синдром моносомии Х (ХО-синдром, синдром Шерешевского-Тернера): а - внешний вид больной; б - кариотип женщины с синдромом ХО: I - выраженная трапециевидная шейная складка, широкая грудная клетка, широко расставленные и слабо развитые соски молочных желез; II - характерные лимфатические отеки на ногах

Рис. 4.21. Кариотип женщины с синдромом трисомии Х
кариотипы 47ХХХ (рис. 4.21), 48ХХХХ, 49ХХХХХ. Обнаружены субъекты с увеличенным числом хромосом Y (кариотипы 47ХYY, 48ХYYY) - синдром Клайнфельтера. Названный синдром воспроизводится и при ка-риотипе XXY (рис. 4.22). Встречаются кариотипы 48ХХХY и 48ХХYY, а также мозаики, в организме которых часть клеток имеет моносомию по хромосоме Х, тогда как часть клеток отличается обычным мужским кариотипом - 45Х0/46ХY). Лица с кариотипами, приведенными выше и имеющими отклонения от нормального, характеризуются более или менее выраженным нарушением здоровья, в частности полового.
Известны организмы-трисомики по аутосомам. Лица с синдромом Дауна, например, характеризуются трисомией по хромосоме 21. «Лишняя» хромосома 21 у таких пациентов может существовать самостоятельно (рис. 4.23) или же быть транслоцированной на другую хромосому
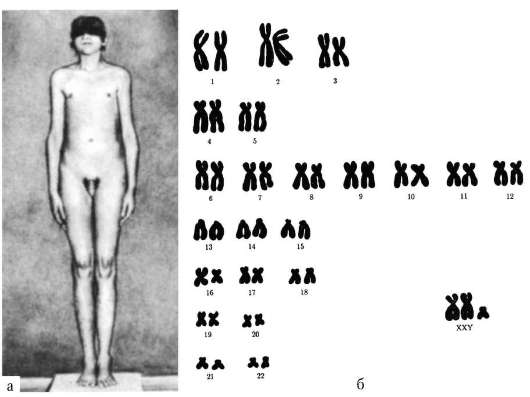
Рис. 4.22. Синдром Клайнфельтера: а - внешний вид больного (характерен высокий рост, непропорционально длинные конечности); б - кариотип больного (XXY)
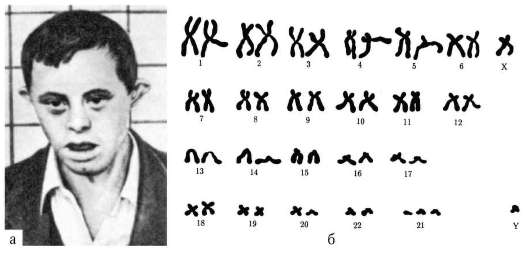
Рис. 4.23. Синдром трисомии 21 (синдром Дауна): а - внешний вид больного; б - кариотип больного
(рис. 4.24). Молекулярно-цитогенетические и клинико-генетические исследования последних десятилетий показали, что хромосома 21 имеет «критический» участок (q22.3). Увеличение дозы сайтов этого участка до трех, собственно, и дает развитие синдрома Дауна (частичная трисо-мия по хромосоме 21). Предположительно ключевую роль в развитии типичной для названного синдрома умственной отсталости играет увеличение дозы гена фермента супероксиддисмутазы, располагающегося в критическом участке q22.3. Напомним, что этот фермент - важный участник внутриклеточных антиоксидантных механизмов, обеспечивающих снижение негативных последствий образования АФК (свободные радикалы), см. п. 2.4.8. Установлено, что у человека, наряду с трисоми-ей 21, совместимы с жизнью трисомии по хромосомам 8 (рис. 4.25), 13 (синдром Патау - рис. 4.26) и 18 (синдром Эдвардса - рис. 4.27), тогда как полные трисомии по хромосомам 1, 5, 6, 11 и 19 приводят к гибели эмбриона на ранних стадиях развития. Трисомия по хромосоме 16 обнаруживается только в материале абортусов.
Цитологические механизмы геномных мутаций связаны с нарушением гаметогенеза (мейоза): с нерасхождением или утратой геномов (полиплоидия, гаплоидия), а также отдельных хромосом (анэуплоидия). Нерасхождение в мейозе геномов приводит к образованию диплоидных гамет. При оплодотворении таких гамет гаплоидной гаметой возникают триплоидные особи. Рождение тетраплоидных организмов имеет в своей основе более сложные нарушения мейоза или же события на стадии первых дроблений зиготы.
Из практики селекционной работы известны примеры образования жизнеспособных и дающих потомство гетерополиплоидных растений. В частности, речь идет о межвидовом гибриде капусты и редьки (Г.Д. Кар-печенко). В гаплоидном геноме гамет этого гибрида объединены геномы (хромосомы) родителей - капусты и редьки. Названный гибрид не нашел применения в сельском хозяйстве, поскольку в противоречии с ожидаемым - надземная часть капусты и подземная редьки, и перспектива получать с одной площади земли одновременно урожай двух разных овощей - он имел надземную часть редьки и подземную капусты.
К первичной гаплоидности может привести развитие особи из яйцеклетки без оплодотворения (партеногенез).
К анэуплоидиям ведет нерасхождение хромосом в анафазе первого деления мейоза. При этом одна из будущих гаплоидных половых клеток несет «лишнюю» хромосому, тогда как вторая лишена этой хромосомы.
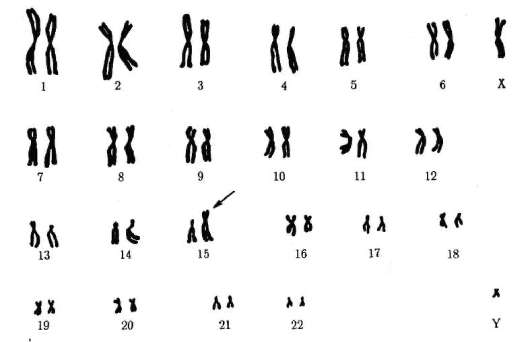
Рис. 4.24. Кариотип при транслокационном синдроме Дауна. Одна хромосома 21 присоединена к хромосоме 15 - указано стрелкой
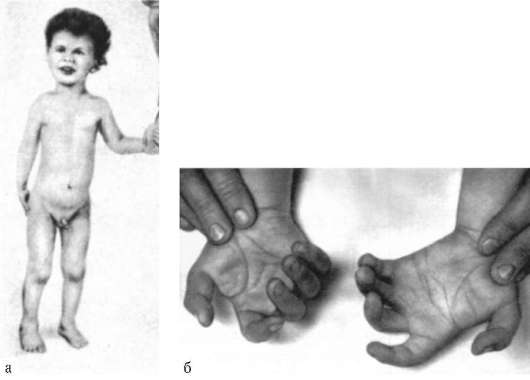
Рис. 4.25. Синдром трисомии 8: а - внешний вид больного; б - контрактуры в межфаланговых суставах кистей
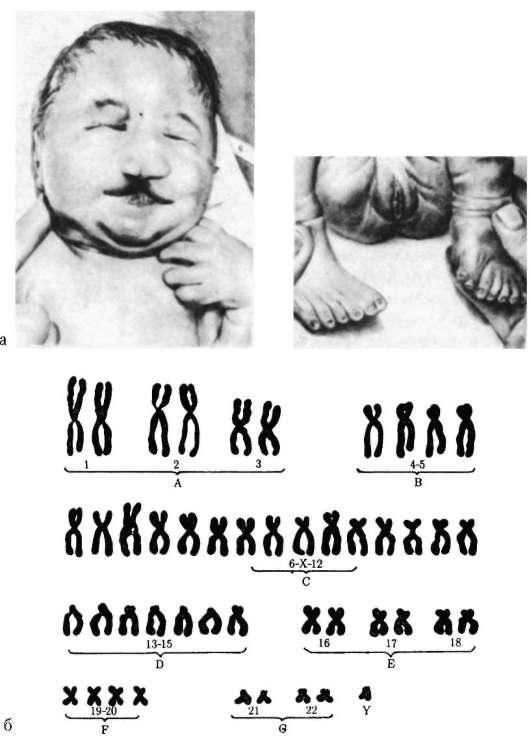
Рис. 4.26. Синдром трисомии 13 (синдром Патау): а - внешний вид больного; б - кариотип больного трисомией в группе D
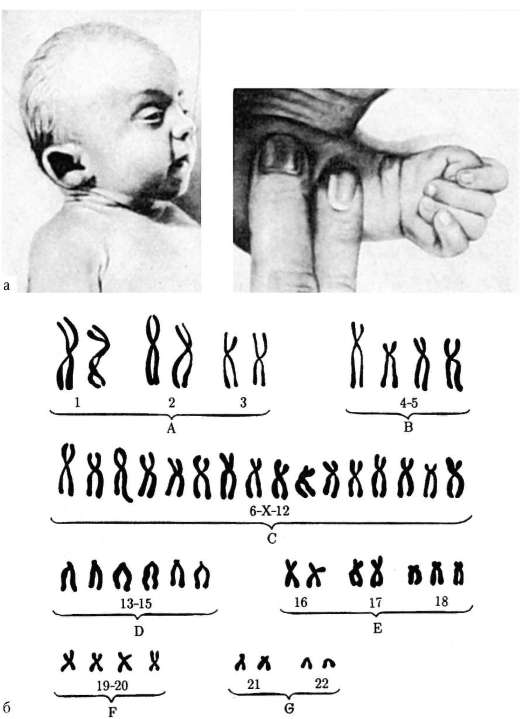
Рис. 4.27. Синдром трисомии 18 (синдром Эдвардса): а - внешний вид больного; б - кариотип больного трисомией в группе Е
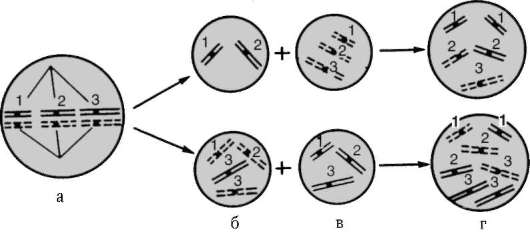
Рис. 4.28. Нарушение расхождения отдельных бивалентов (1, 2, 3) как причина анэуплоидий: а - метафаза I мейоза; б - образование аномальных гамет вследствие нарушения расхождения 3-го бивалента в анафазе I мейоза; в - оплодотворение аномальной гаметы нормальной гаметой особи другого пола; г - образование зигот с анэуплоидными кариотипами (моносомия и три-сомия по хромосоме 3, соответственно сверху и снизу)
Участие таких гамет в оплодотворении приводит соответственно к три-сомиям и моносомиям (рис. 4.28).
4.3.3.4. Биологическое значение геномного уровня организации генетического аппарата
Геном, объединяя всю совокупность хромосомных генов (сайтов, нуклеотидных последовательностей ДНК), представляет собой эво-люционно сложившуюся структуру, которая отличается относительно большей стабильностью в сравнении со специфическими структурами генного и хромосомного уровней - генами и хромосомами. На геномном уровне, т.е. в геноме, функционирует система сбалансированных (взаимосоответствующих) по дозам и объединенных сложными функциональными взаимосвязями генов, которая представляет собой нечто большее, чем простая совокупность отдельных единиц наследственности. Именно поэтому результатом функционирования генома как единого целого является формирование фенотипа организма соответствующего биологического вида во всем многообразии его (организма) характеристик на всем протяжении индивидуального развития. Таким образом, поддержание постоянства организации генетического аппарата на геномном уровне имеет первостепенное значение для обеспечения нормального развития особи и воспроизведения в ходе этого развития в первую очередь видовых характеристик.
В то же время рекомбинация единиц наследственности в генотипах особей, например, в связи с мейозом и оплодотворением обусловливает генетическое разнообразие, что имеет особое эволюционное значение. Мутации, реализующиеся на геномном уровне через изменение характера межгенных взаимодействий, в частности в области регуляторных нуклеотидных последовательностей ДНК, отличаются широким плейо-тропным действием. Количественные изменения доз, а также транслокации и транспозиции генов влияют на экспрессию генов, а включение в геном чужеродной (другого вида организмов) биологической информации при горизонтальном переносе нуклеотидных последовательностей может оказаться эволюционно перспективным, став основной причиной ускорения темпов эволюционного процесса на отдельных этапах исторического развития живых форм на Земле.
4.3.4. ПОНЯТИЕ О КАРИОТИПЕ
Генетические явления, характеризующие наследственность и биологическую изменчивость, биологи достаточно давно связывают с особыми ядерными образованиями - хромосомами (см. п. 4.2, А. Вейсман; В. Вальдейер; Т. Бовери и У. Сеттон; Т. Морган), которые с полными основаниями рассматриваются в качестве структур, в которых размещаются гены (см. п. 4.3.2.1). В истории генетики как науки на протяжении длительного времени при отсутствии реальных знаний о материальном носителе свойств наследственности и изменчивости и благодаря опережающему развитию микроскопической техники хромосомы были фактически единственным объектом для непосредственных наблюдений. Это привело к появлению цитогенетического метода генетического анализа, которому и сейчас принадлежит важное место, в частности, в практике МГК, а также особого понятия - кариотип (см. п. 4.3).
Кариотип - это диплоидный набор хромосом (2n), свойственный соматическим клеткам организмов данного вида, представляющий собой видоспецифический сложный признак и характеризующийся определенным числом, строением и генным составом хромосом (рис. 4.29). В таблице 4.2 приведено число хромосом в ядрах соматических клеток некоторых видов животных.
Если число хромосом в одинарном гаплоидном наборе хромосом половых клеток обозначить n, то формула кариотипа будет выглядеть как 2n. Значение n обычно различно у разных видов. Таким образом,
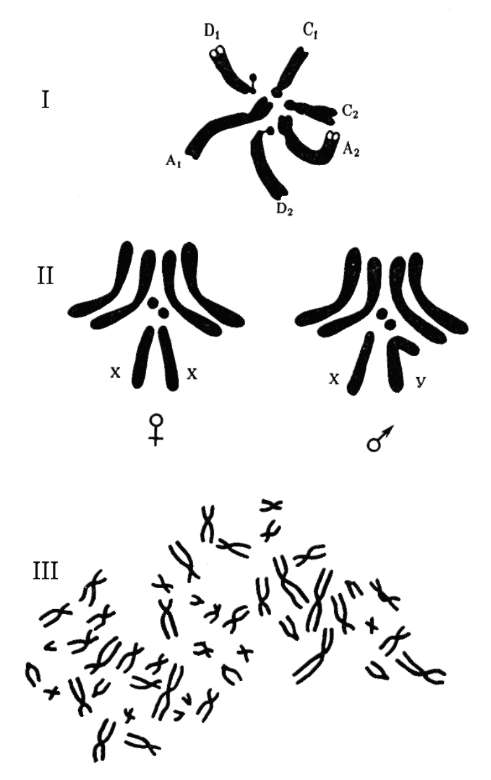
Рис. 4.29. Кариотипы организмов различных видов: I - скерда; II - дрозофила; III - человек
Таблица 4.2. Диплоидное число хромосом (кариотип) в соматических клетках разных видов животных
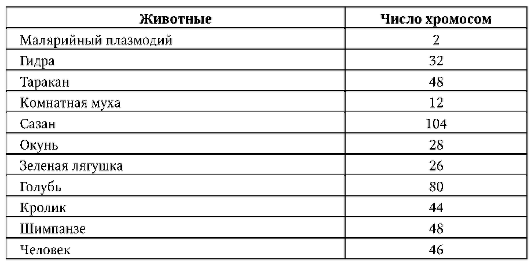
гаплоидное количество хромосом в гаметах людей равно 23 (n = 23), а диплоидное, соответствующее кариотипу, - 46 (2n = 46).
Каждая хромосома представлена в кариотипе парой гомологов. Одна из гомологичных хромосом пары унаследована от отца, другая - от матери через половые клетки родителей, принявшие участие в оплодотворении. Генный состав пары гомологичных хромосом одинаков. Вместе с тем один и тот же ген в гомологах может быть представлен разными его альтернативными формами или аллелями (аллельными генами). Учитывая известные отношения между аллелями в виде доминантности и рецессивности, а также присутствие в гомологичных хромосомах одинаковых, либо доминантных, либо рецессивных аллелей или же разных аллелей (доминантного и рецессивного), возможны состояния доминантной гомозиготности, рецессивной гомозиготно-сти и гетерозиготности (см. также п. 4.3.1.2).
В кариотипах строго гомологичными хромосомами (аутосомы) представлены все пары, кроме одной (гетерохромосомы или половые хромосомы). В клетках пара половых хромосом у особей одного пола (гомогаметный пол, у человека - женский) представлена двумя одинаковыми хромосомами (у человека - ХХ), тогда как у другого (гетеро-гаметный пол, у человека - мужской) двумя разными хромосомами (у человека - ХY). В первом случае генный состав пары половых хромосом совпадает. Поэтому в зависимости от совпадения или несовпадения в двух хромосомах Х аллелей соответствующих генов воспроизводятся известные состояния доминантной или рецессивной гомозиготности и гетерозиготности. Большинство генов разных половых хромосом особей гетерогаметного пола различны. В связи с этим возможно состояние гемизиготности, когда у особей гетерогаметного пола (у людей мужской - ХY), ген хромосомы Х, не имея гомолога в хромосоме У, присутствует в кариотипе в единственном экземпляре. Такой ген обязательно проявит себя в фенотипе, даже если он представлен рецессивным алле-лем. Существуют виды, у которых самки и самцы различаются числом гетерохромосом, соответственно ХХ и ХО.
Подчеркнем еще раз, что характеристика кариотипа как видового признака не может ограничиваться исключительно количеством хромосом. Из материалов табл. 4.2, например, следует, что одинаковое число хромосом может быть в кариотипах двух далеко отстоящих друг от друга в эволюционном плане видов - таракана и шимпанзе (см. также
п. 4.3).
Рис. 4.30. Хромосома человека, дифференциальная окраска (схема): р - короткое плечо хромосомы; q - длинное плечо хромосомы; цифрами обозначены сегменты
На настоящий момент разработаны и применяются методы избирательной (дифференциальной) окраски метафазных хромосом, которые выявляют типичную (неповторимую, уникальную) для отдельных хромосом поперечную исчерченность, что дает возможность их безошибочной идентификации (рис. 4.30). Введение в цитогенетическую практику методов дифференциальной окраски позволило перейти от Денверской классификации хромосом человека к более информативной и, следовательно, более удобной для врачей-генетиков Парижской классификации (см. п. 4.2).
Важнейшая функционально-генетическая характеристика хромосом и кариотипа в целом - то, что они в клеточном цикле редуплицируются (относительно использования терминов «репликация» и «редупликация», см. п. 2.4.5.3) путем самокопирования.
4.3.5. КЛЕТОЧНЫЕ МЕХАНИЗМЫ, ОПРЕДЕЛЯЮЩИЕ ТИПЫ НАСЛЕДОВАНИЯ ПРИЗНАКОВ, КОНТРОЛИРУЕМЫХ ЯДЕРНЫМИ ГЕНАМИ
Начальная (стартовая) биоинформация, на основе которой в процессе индивидуального развития формируется фенотип эукариотического организма, находится главным образом (90%) в ядерных структурах клеток - хромосомах. Некоторое (порядка 10%) количество наследственного материала (ДНК) содержится в цитоплазматических клеточных структурах - в частности, в митохондриях (мтДНК, хромосома М).
Ядерные и цитоплазматические структуры, содержащие ДНК, в процессе клеточного деления (митоз, мейоз) распределяются между дочерними клетками по-разному. Это касается не только соматических клеток, но также гамет. В связи с отмеченным, передача ядерных и цитоплаз-матических генов потомству происходит по разным правилам, что обусловливает особенности наследования соответствующих признаков. Сказанное нашло отражение в том, что наряду с ядерной хромосомной наследственностью известна цитоплазматическая наследственность (см. п. 4.2; К.Э. Корренс и Э. Баур).
Гены (сайты, нуклеотидные последовательности ДНК), связанные с ядерными хромосомами, закономерно распределяются между дочерними клетками благодаря механизму митоза, который обеспечивает постоянство кариотипа в ряду клеточных поколений. Мейоз и оплодотворение обеспечивают постоянство кариотипа в ряду поколений организмов, размножающихся половым путем. Поведение хромосом в митозе, мейозе и при оплодотворении лежит в основе наблюдаемых в природе типов наследования признаков, контролируемых ядерными генами (см. табл. 4.3).
Таблица 4.3. Основные типы наследования признаков, контролируемых ядерными генами
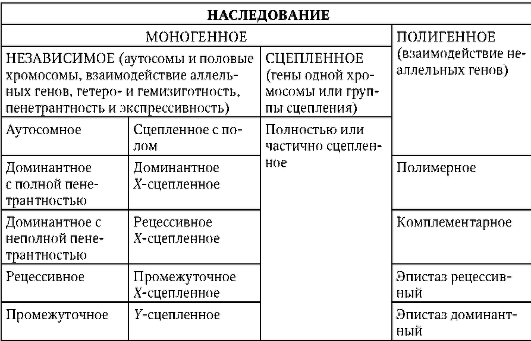
4.3.5.1. Моногенное независимое наследование: аутосомное и сцепленное с полом
Так как кариотип организма - диплоидный набор хромосом, гены аутосом и хромосомы Х (гомогаметный пол, у людей женский - 46ХХ) представлены аллельными парами. Аллельные гены, занимая гомологичные локусы в гомологичных хромосомах (аутосомах или хромосоме Х), проявляя свойства доминантности, рецессивности и другие (см. п. 4.3.1.2) и взаимодействуя между собой, определяют основные типы моногенного независимого наследования признаков (см. табл. 4.3) - аутосомно-доминантный, аутосомно-рецессивный и, если речь идет о гетерозиготах, промежуточный, когда ни один из двух аллелей не проявляет относительно другого аллеля свойства абсолютной доминантности. Неполная пенетрантность доминантного аллеля гена, контролирующего развитие анализируемого признака, вносит свои особенности в характеристики аутосомно-доминантного наследования (рис. 4.31).
Так как хромосома Х присутствует в кариотипе каждой особи независимо от пола, признаки, контролируемые ее генами, формируются в фенотипах и мужских, и женских особей. Если это представители гомогаметного пола, имеющие в кариотипе две хромосомы Х, то благодаря возможности гомозиготности (доминантной или рецессивной), гетерозиготности или ситуации с неполной доминантностью выделяются принципиально те же типы моногенного независимого наследования - доминантное, рецессивное и промежуточное. Однако чтобы подчеркнуть факт принадлежности гена к половой хромосоме Х, говорят об Х-сцепленном наследовании (в отличие от аутосомного). По-иному обстоят дела у особей гетерогаметного пола, кариотипы которых несут одну хромосому Х, имеющую в паре половых хромосом хромосому Y. Половые хромосомы Х и Y отличаются отсутствием полной гомологии локусов (рис. 4.32). В связи с этим по многим генам хромосомы Х особи гетерогаметного пола являются гемизиготами, в силу чего ген хромосомы Х проявляет себя в фенотипе такой особи всегда, вне зависимости от того, представлен ли он доминантным или рецессивным аллелем. Этим объясняются некоторые особенности рецессивного Х-сцепленного типа наследования (рис. 4.33).
Наличие хромосомы Y исключительно в кариотипах особей гетеро-гаметного пола дает основания к выделению Y-сцепленного типа наследования в качестве самостоятельного и объясняет его особенности.
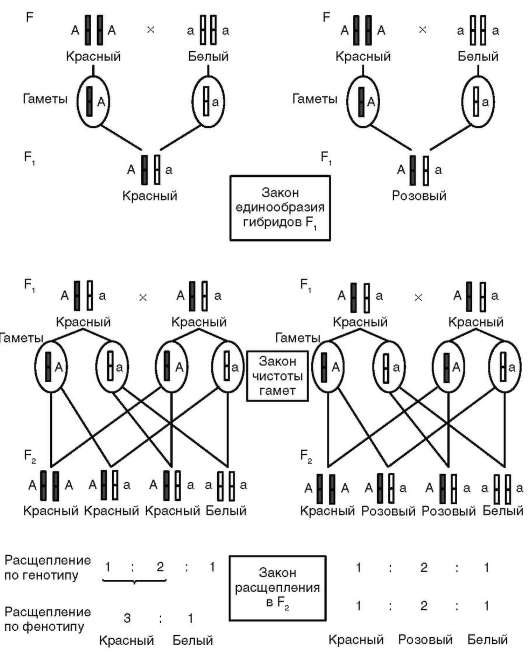
Рис. 4.31. Аутосомный тип наследования признака: I - при полном доминировании аллелей (цвет лепестков у гороха); II - при неполном доминировании - промежуточное наследование (цвет лепестков у ночной красавицы)
Рис. 4.32. Схема гомологичных и негомологичных локусов половых хромосом человека: I - хромосома Х: заштрихованы локусы, отсутствующие в хромосоме Y (красно-зеленая цветовая слепота, гемофилия А и др.); II - хромосома Y: заштрихованы локусы, отсутствующие в хромосоме Х (гены, участвующие в развитии фенотипа по мужскому типу, перепонки между пальцами)
Главная из этих особенностей - то, что передача в ряду поколений признаков, контролируемых генами хромосомы Y, если эти гены не имеют гомологов в хромосоме Х, происходит только по линии особей гетерога-метного пола - у людей мужского пола (рис. 4.34).
4.3.5.2. Еще раз о независимом наследовании. Соотносительное наследование нескольких признаков. Сцепленное наследование
Выше рассмотрены типы наследования отдельных признаков. Однако фенотип организма определяется совокупностью многих признаков, за развитие которых отвечают разные гены, принадлежащие одной, занимая в ней различные локусы, или разным группам сцепления (хромосомам). При этом обнаруживаются признаки, которые наследуются как независимо друг относительно друга, так и совместно (сцеплено).
Закон независимого наследования признаков сформулирован Г. Менделем по результатам наблюдений за наследованием одновременно двух признаков - цвета и формы горошин. Названный закон утверждает, что различные признаки, контролируемые неаллельными генами
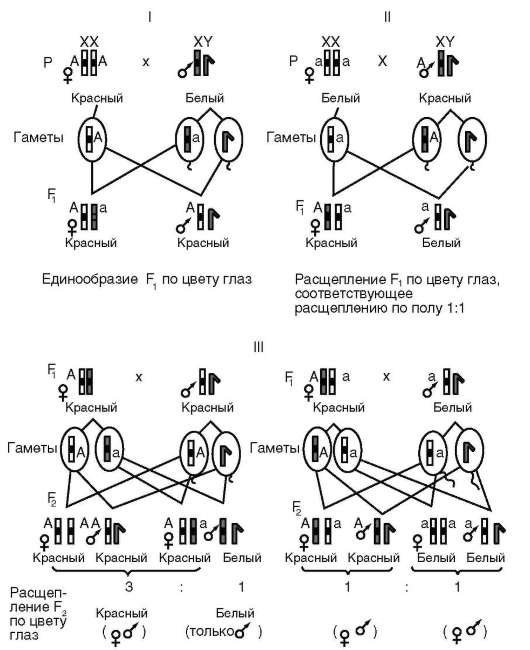
Рис. 4.33. Наследование, сцепленное с полом (окраска глаз у дрозофилы):
I - сочетание половых хромосом в кариотипе представителей разного пола;
II - гомогаметный пол образует один тип гамет, гетерогаметный - два; III - представители гомогаметного пола получают хромосомы Х от обоих родителей, представители гетерогаметного пола получают единственную хромосому Х от гомогаметного родителя, а хромосому Y - от гетерогаметного родителя. Окрашены отцовские хромосомы
Рис. 4.34. Обоснование особенностей Х-сцепленного и У-сцепленного типов наследования поведением половых хромосом при образовании гамет особями гомо- и гетерогаметного пола и при оплодотворении: I - сочетание половых хромосом в кариотипах представителей разных полов; II - гомогаметный пол образует один тип гамет, гетерогаметный два; III - представители гомогаметно-го пола получают хромосомы от обоих родителей; представители гетерогамет-ного пола получают хромосому Х от гомогаметного родителя, а хромосому У от гетерогаметного родителя; это справедливо для генов, расположенных в негомологичных локусах хромосом Х и У; окрашены отцовские хромосомы
(наследственными задатками), передаются от родителей потомству независимо друг от друга и обнаруживаются в фенотипах потомков во всех возможных сочетаниях (рис. 4.35).
Очевидно, что этому закону должны подчиняться признаки, развитие которых контролируют неаллельные гены, находящиеся в разных (негомологичных) хромосомах. В таком случае независимый характер наследования двух признаков или более объясняется поведением негомологичных хромосом в мейозе. Названные хромосомы образуют со своими гомологами в первом делении мейоза пары (биваленты). В анафазе I мейоза гомологи каждой пары расходятся к полюсам делящейся клетки независимо от гомологов других пар. В результате гаплоидные хромосомные наборы будущих гамет на полюсах представлены случайными сочетаниями отцовских и материнских хромосом. Следовательно, различные гаметы содержат разные комбинации отцовских и материнских аллелей неаллельных генов.
Разнообразие вариантов образуемых гамет зависит от степени ге-терозиготности организма и описывается формулой 2n, где n - число
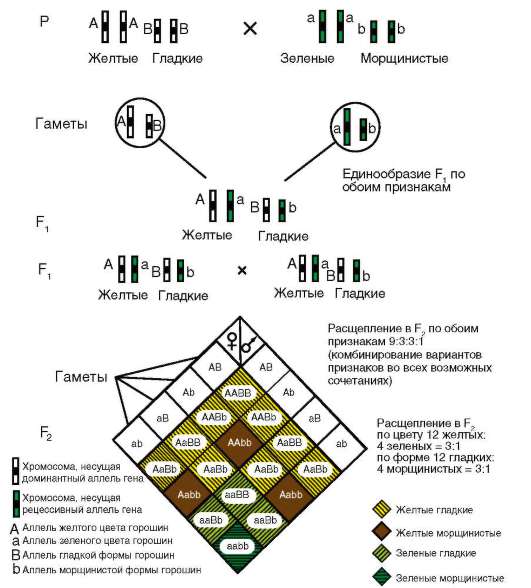
Рис. 4.35. Независимое наследование двух признаков (цвет и форма горошин)
локусов в гетерозиготном состоянии. В связи с этим дигетерозиготные гибриды первого поколения (F1), рождаемые в скрещиваниях гомозиготных, по доминантным (один родитель - АВ) и рецессивным (второй родитель - ав) аллелям анализируемых генов, проявляя в силу гетеро-зиготности и свойства доминантности-рецессивности аллелей (АаВв) фенотипическое единообразие по обоим наблюдаемым признакам, образуют четыре варианта гамет, причем с равной вероятностью каждо-
го из вариантов. При скрещивании гибридов F1 между собой благодаря всем возможным комбинациям аллелей, присутствующих в гаметах указанных четырех вариантов, в случае независимого наследования двух признаков (дигибридное скрещивание) среди гибридов второго поколения (F2) обнаруживается четыре фенотипические группы (в сравнении с F1 происходит расщепление по фенотипу) потомков в отношении 9:3:3:1. Анализ потомства F2 отдельно по каждой из двух наблюдаемых пар альтернативных признаков - цвет (зеленый или желтый) и форма (гладкая или морщинистая) горошин - выявляет наличие по каждой паре двух фенотипических групп (расщепление по фенотипу) потомков в отношении 3:1 (моногибридное скрещивание).
В опытах Г. Менделя наследственная конституция гибридов F1 (АаВв) устанавливалась путем анализа фенотипов потомства F2, получаемого при самоопылении растений-родителей (F1). Эта же задача решается, если применяется так называемое анализирующее скрещивание. Оно заключается в скрещивании организма, генотип которого необходимо установить, с организмом-гомозиготой по рецессивному(ым) аллелю(ям) соответствующего(их) гена(ов) - рис. 4.36 (моногибридное анализирующее скрещивание) и рис. 4.37 (дигибридное анализирующее скрещивание). Так как гомозиготные родители образуют один тип гамет: аа-а (моногибридное скрещивание), ааЪЪ-аЪ (дигибридное скрещивание), ааЪЪсс-аЪс (тригибридное скрещивание) и т.д., - при анализирующем скрещивании количество разных фенотипов потомков зависит от числа типов гамет организма с доминантным фенотипом. Если последний гомозиготен по анализируемым генам, то он тоже образует только один тип гамет, и потомство в анализирующем скрещивании отличается фенотипическим единообразием, так как все потомки имеют доминантный фенотип (см. рис. 4.35, I). Если организм, генотип которого необходимо установить, гетерозиготен по одному гену, он образует два типа гамет, в силу чего при анализирующем скрещивании рождают-
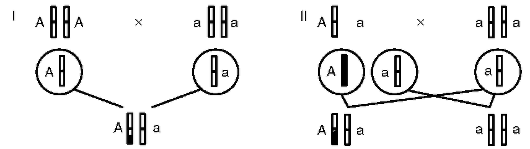
Рис. 4.36. Анализирующее моногибридное скрещивание. Объяснения в тексте
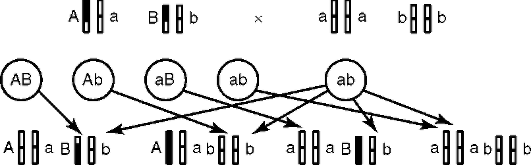
Рис. 4.37. Анализирующее дигибридное скрещивание. Объяснения в тексте
ся потомки двух разных фенотипов - один с доминантным и второй с рецессивным признаком (см. рис. 4.35, II). Дигетерозиготный организм дает при анализирующем скрещивании потомство с четырьмя разными фенотипами (см. рис. 4.37).
Признаки, контролируемые генами, находящимися в одной хромосоме, могут демонстрировать как независимый, если соответствующие локусы расположены относительно далеко - 50 морганид (сантиморга-нид) и более, так и сцепленный тип наследования. В такой ситуации признаки передаются потомству всегда или в определенном проценте случаев совместно. Подозрение в том, что феномен сцепленного наследования существует, возникло, когда было обнаружено, что результаты анализирующего скрещивания гибридов F1 у дрозофил иногда отличались от ожидаемых, если исходить из исключительно независимого соотносительного наследования нескольких признаков (Т.Г. Морган). Конкретно у потомков в таких скрещиваниях вместо ожидаемого свободного комбинирования фенотипических признаков, контролируемых аллельными парами разных генов, наблюдали тенденцию к наследованию преимущественно родительских сочетаний признаков. Как уже отмечалось, в основе сцепленного соотносительного наследования признаков лежит расположение соответствующих генов в одной хромосоме. Именно это обстоятельство привело к тому, что каждая хромосома стала рассматриваться как отдельная группа сцепления. На рисунке 4.38 представлены результаты наследования признаков окраски тела и формы (длины) крыльев у дрозофилы, а также цитологическое обоснование этих результатов. При анализирующем скрещивании самцов из F1 появлялось всего два фенотипических вида потомков, сходных с родительскими формами по сочетанию вариантов анализируемых признаков (серая окраска тела и нормальные крылья; черная окраска тела и короткие крылья) в соотношении 1:1. Это указывает на образо-
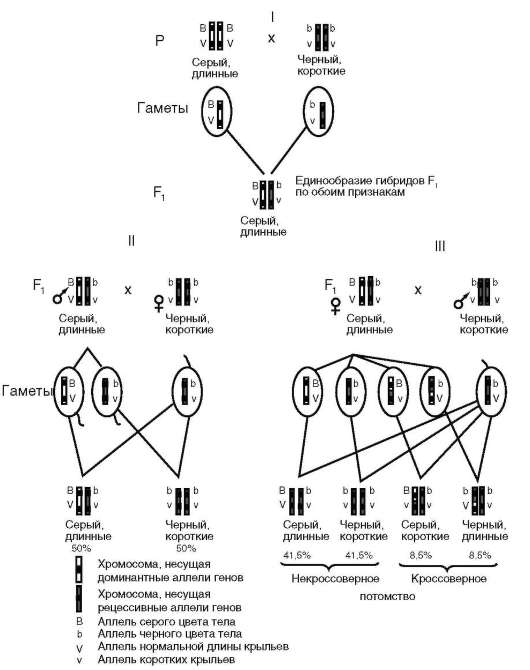
Рис. 4.38. Сцепленное наследование признаков (окраска тела и длина крыльев) у дрозофилы: I - скрещивание чистых линий; II и III - анализирующие скрещивания, соответственно самцов и самок из F1
вание (с одинаковой вероятностью) самцами F1 всего двух типов гамет, в которые попадают исходные (родительские) сочетания аллелей генов, контролирующих развитие названных признаков - окраска тела (В и b) и форма или длина крыльев (V и v). При анализирующем скрещивании самок F1 обнаруживалось четыре фенотипических варианта потомков, практически со всеми возможными сочетаниями признаков. При этом потомки с родительским сочетанием признаков составили 83%. У 17% потомков - кроссоверное потомство - обнаруживались иные комбинации признаков (серая окраска тела и короткие крылья; черная окраска тела и нормальные крылья). Видно, что в указанных скрещиваниях проявляется склонность к сцепленному наследованию либо доминантных, либо рецессивных признаков (83%). Частичное (17%) нарушение сцепления наследования признаков потомками объясняется процессом кроссинговера - обменом гомологичными участками между гомологичными хромосомами в профазе I мейоза.
Сцепленное соотносительное наследование двух признаков или более не следует путать с наследованием, сцепленным с полом. Напомним, что наследование, сцепленное с полом, выделяется на основании одного критерия - локализации соответствующего гена в половой хромосоме (Х или У).
Анализ соотносительного наследования сочетаний других признаков, прежде всего у плодовых мух дрозофил, показал, что процент кроссоверного потомства для каждой пары признаков всегда один и тот же, но различается для разных пар. Это послужило основанием для заключения о том, что гены («наследственные задатки», по Г. Менделю) располагаются в хромосомах в линейном порядке, причем разные (негомологичные) хромосомы представляют собой группы сцепления определенных генов. Напротив, гомологичные хромосомы - это группы сцепления одних и тех же генов, которые, однако, в гомологах могут быть представлены разными аллелями. В профазе I мейоза гомологи каждой пары конъюгируют, т. е. сближаются с точным противостоянием гомологичных локусов (другими словами, аллелей соответствующего гена). Затем благодаря кроссинговеру хромосомы в парах гомологов могут обменяться гомологичными участками. Если обмениваемые участки гомологичных хромосом представлены разными аллелями гена, занимающего локус, обмен приводит к изменению аллельного состава каждой из гомологичных хромосом (один из факторов генотипической комбинативной изменчивости). Следовательно, потомок, получив в результате акта оплодотворения хромосому с измененным аллельным на-
Рис. 4.39. Схема, поясняющая низкий процент кроссоверных гамет для двух генов. Плюсом обозначены клетки-предшественницы гамет, в которых на участке между двумя избранными локусами произошел кроссинговер; красным цветом обозначены кроссоверные гаметы
бором, будет отличаться от родителя определенными фенотипическими особенностями.
Частота, с которой происходит обмен на участке хромосомы между двумя конкретными генами, зависит от расстояния между ними (правило Т. Моргана). Другими словами, при увеличении расстояния между генами одной хромосомы (группы сцепления) вероятность кроссинго-вера между ними растет, однако даже в случае осуществления обмена между генами данной пары во всех клетках-предшественницах половых клеток процент кроссоверных гамет не превышает 50. Это происходит потому, что в акте кроссинговера участвуют две хроматиды из четырех, имеющихся в каждом биваленте (рис. 4.39). С увеличением расстояния между генами в группах сцепления растет вероятность того, что на соответствующем участке одновременно произойдет несколько кроссин-говеров. Так как каждый второй перекрест (кроссинговер) приводит к восстановлению в хромосоме прежнего сочетания аллелей, то с ростом расстояния между генами количество кроссоверных гамет может не увеличиваться, а уменьшаться. Следовательно, процент кроссоверных гамет представляет собой адекватный показатель расстояния между двумя генами, только если они находятся на достаточно близком расстоянии, когда исключается вероятность второго кроссинговера.
Различают неполное (частичное) и полное сцепление. Полное сцепление (фактически отсутствие кроссинговера) может быть видовой характеристикой гаметогенеза у представителей одного из полов, например у самцов дрозофилы.
Применение анализирующего скрещивания дает возможность выяснить не только генный состав отдельных групп сцепления (хромосом), но также установить расстояние между генами.
4.3.5.3. Еще раз о наследовании признаков, развитие которых обусловлено взаимодействием неаллельных генов
На характер наследования в ряду поколений особей сложных фе-нотипических признаков определенное влияние оказывает взаимодействие неаллельных генов. Различные комбинации аллелей таких генов могут привести к появлению нового варианта признака, к исчезновению признака, к изменению формы его проявления у потомков. Взаимодействующие неаллельные гены, сами по себе, могут наследоваться друг относительно друга независимо или сцеплено. Это влияет на частоту, с которой у потомства появляются комбинации аллелей, соответствующие тому или иному фенотипическому результату известных со времен классической генетики вариантов взаимодействия неаллельных генов - полимерия, комплементарность, эпистаз (см. п. 4.3.3.1).
Ниже будут рассмотрены закономерности наследования признаков в случае независимого наследования взаимодействующих неаллельных генов.
43.53-а. Наследование при полимерном взаимодействии неаллельных генов
Когда состояние признака определяется совокупным действием нескольких неаллельных генов, каждый из которых в силу диплоидности эукариот представлен в генотипе соматических клеток парой аллелей, причем действие этих генов характеризуется аддитивным (добавляющим, прибавляющим, кумулятивным: от англ.: additive - помогающий нарастить) фенотипическим эффектом, в поколениях организмов встречаются особи с разной степенью выраженности наблюдаемого признака, что зависит от суммарной дозы аллелей соответствующих генов.
Выше рассмотрен пример полигенного полимерного наследования такого признака, как степень пигментации кожных покровов человека, в развитии которого принимают участие четыре неаллельных гена (четыре пары аллелей). В развитие того, о чем в связи с этим примером уже говорилось (см. п. 4.3.3.1), отметим, что в браке мулата и мулатки, гетерозиготных по всем четырем генам (генотипы родителей одинаковы - Р1р1Р2р2Р3р3Р4р4) и вследствие этого образующих каждый 24=16 типов гамет, среди детей 1/256 часть характеризуется вероятностью иметь максимально пигментированную кожу, еще 1/256 часть - минимально пигментированную кожу, тогда как остальные - это дети с промежуточными значениями интенсивности пигментации кожных покровов (см. рис. 4.17).
Известны примеры, когда доминантные и рецессивные аллели полигенов обусловливают развитие разных вариантов признака. Так, в генотипе растения «пастушья сумка» присутствуют два гена, совместно контролирующих такой признак, как форма стручка. При скрещивании дигетерозигот по указанным генам (рис. 4.40) среди потомков наблюдается расщепление по фенотипу в отношении 15:1, где 15/16 потомства имеет от одного до четырех доминантных аллелей соответствующей пары генов (один фенотип), а 1/16 не имеет в генотипе доминантных аллелей вообще (другой фенотип).
43.53-6. Наследование при комплементарном взаимодействии неаллельных генов
Если формирование фенотипического признака требует комплементарного (взаимодополняющего: от англ.: complementary - дополнительный, добавочный) действия определенных аллелей неаллельных генов, то такой признак может появиться только у тех особей, в генотипе которых имеется требуемая комбинация, в частности, доминантных аллелей взаимодействующих генов. Другими словами, признак в определенном своем состоянии воспроизводится в фенотипе исключительно при наличии в генотипе доминантных аллелей соответствующих генов и не воспроизводится при отсутствии доминантного аллеля хотя бы одного из них. В таком случае при скрещивании дигетерозиготных особей между собой анализируемый признак обнаруживается у части потомков - 9/16, тогда как у остальных - 7/16 он отсутствует (рис. 4.41).
Известны также ситуации, когда каждый из взаимодействующих в формате комплементарности неаллельных генов при отсутствии доминантного аллеля другого гена дает свой вариант признака, тогда как, будучи оба представленными в генотипе доминантными аллелями, они, взаимодействуя, дают другой вариант признака (рис. 4.42).
У людей два неаллельных гена, контролирующие отложение в волосах черного и красного пигментов, что обусловливает известные варианты естественного цвета волос, при определенных сочетаниях аллелей дают новый признак - особый блеск волос (глянцевитые и/или лоснящиеся волосы).
43.53-в. Наследование при эпистатическом взаимодействии неаллельных генов
Вспомним, что в случае эпистаза один из генов (В) проявляет себя фенотипически только при отсутствии в генотипе определенного алле-ля другого гена (А). Строго говоря, эпистатическое взаимодействие не-
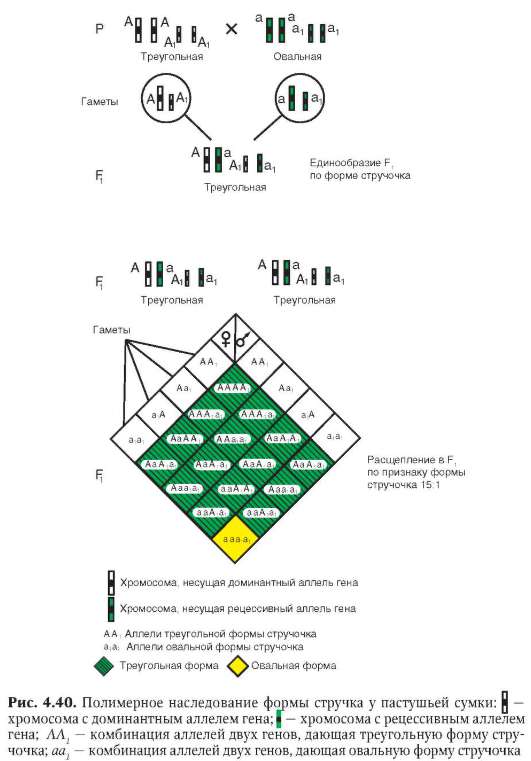
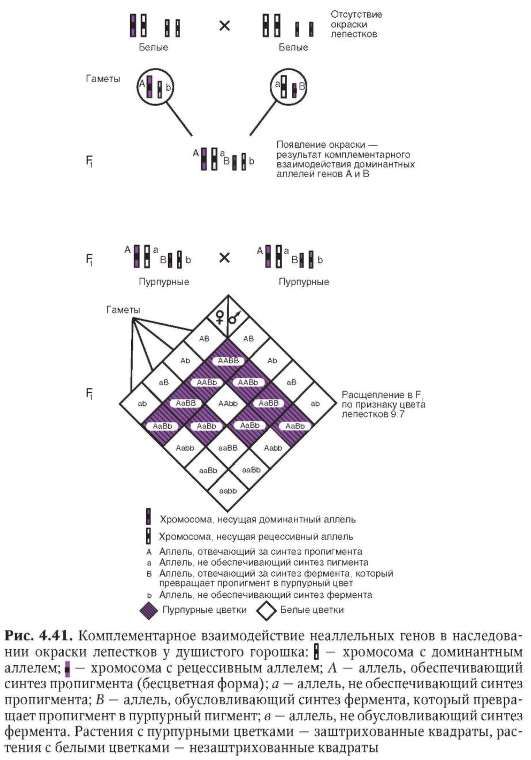
Рис. 4.42. Комплементарное взаимодействие неаллельных генов в наследовании формы гребня у кур: I - A?bb - розовидная; II - aaB? - гороховидная; III - A?B? - ореховидная; IV - aabb - листовидная
аллельных генов можно рассматривать как вариант комплементарного взаимодействия неаллельных генов. Действительно, речь, по существу, идет о том, что и при эпистазе в части соответствующих признаков фенотипы особей зависят от конкретного сочетания в их генотипах аллелей неаллельных генов. Соответственно, расщепление по фенотипу среди потомков от скрещивания дигетерозигот по генам анализируемых признаков может быть различным.
При доминантном эпистазе, когда доминантный аллель одного гена (генотипы - АА или Аа) препятствует фенотипическому проявлению любого из аллелей (генотипы - BB, Bb или bb) другого гена, расщепление по фенотипу может быть в отношении 12:3:1 или 13:3 (рис. 4.43). При рецессивном эпистазе ген, определяющий развитие какого-либо признака (В), не проявляет себя в фенотипе у рецессивных гомозигот (генотипы - аа) по другому гену (А). Расщепление по фенотипу среди потомства от скрещивания дигетерозигот даст соотношение 9:3:4 (рис. 4.44).
Примером рецессивного эпистаза у людей является «бомбейский феномен», рассмотренный ранее (см. п. 4.3.3.1).
Приведенные выше соотношения особей с разными фенотипами среди потомства (расщепление по фенотипу), получаемого в скрещиваниях гетерозигот или в анализирующих скрещиваниях, в ситуациях, когда имеет место взаимодействие неаллельных генов, так же как в случаях моноили дигибридного скрещивания при моногенном независимом и других типах наследования, носят вероятностный характер. Эти соот-
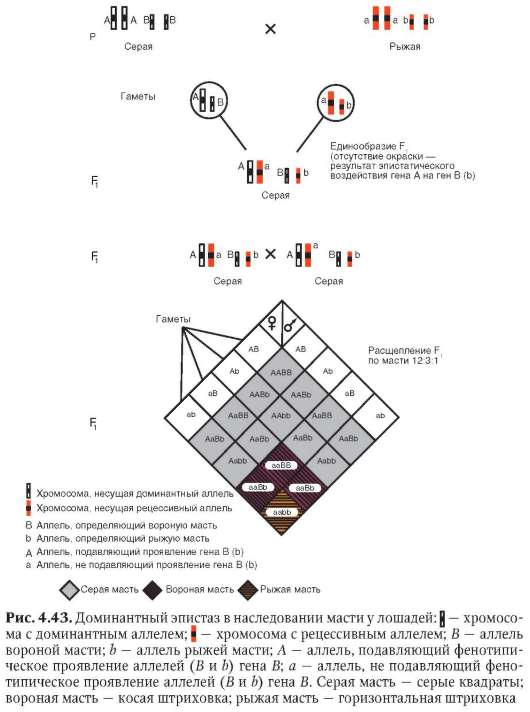
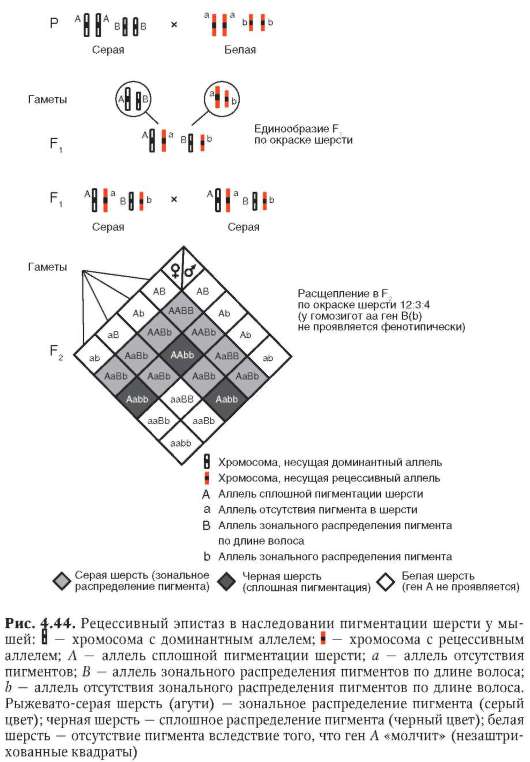
ношения регистрируются исследователем только, если в гаметогенезе образуются все возможные генотипические варианты гамет, которые в равной мере жизнеспособны, вследствие оплодотворения возникают все возможные при данном генотипическом ассортименте гамет гено-типические варианты зигот, и все развивающиеся из этих зигот особи одинаково жизнеспособны. При этом потомство должно быть многочисленным, что делает результаты статистической обработки материала свободными от случайных искажений. При выполнении перечисленных условий гибридологический метод генетического анализа, разработанный Г. Менделем, является надежным инструментом познания закономерностей наследования признаков (у растений и у животных, исключая мир человека).
4.3.6. НАСЛЕДОВАНИЕ ПРИЗНАКОВ, ОБУСЛОВЛИВАЕМОЕ ВНЕЯДЕРНЫМИ ГЕНАМИ. ЦИТОПЛАЗМАТИЧЕСКАЯ НАСЛЕДСТВЕННОСТЬ
Порядка 10% ДНК эукариотической клетки связано не с ядерными структурами (хромосомами), а с внутриклеточными образованиями преимущественно цитоплазматической локализации, в частности с митохондриями и пластидами (растительные клетки). Внеядерные (внехромосомные) генетические элементы также участвуют в формировании фенотипических признаков в процессе индивидуального развития и жизнедеятельности организма. Вместе с тем наследование признаков, контролируемых генетическими элементами внеядерной локализации (цитоплазматическими генами, цитоплазматической ДНК), характеризуется своими особенностями - цитоплазматическая наследственность. Оно не следует правилам наследования признаков (см. табл. 4.3), установленным Г. Менделем - неменделевское наследование - и вытекающим из закономерного поведения хромосом при митозе, мейозе и оплодотворении (см. п. 4.3.2 и 4.3.2.1). Так как организм, развивающийся из зиготы, образующейся в результате оплодотворения, получает цитоплазматические структуры, в частности митохондрии, исключительно от яйцеклетки, цитоплазматическое наследование соответствующих признаков осуществляется по материнской линии.
Цитоплазматический тип наследования характерен для признака «пестрые листья» у некоторых растений. Наличие названного признака обусловлено мутацией в ДНК хлоропластов, фенотипически проявляющейся в нарушении образования зеленого пигмента хлорофилла. Раз-
множение в растительных клетках нормальных (зеленых) и мутантных (бесцветных) пластид с последующим их случайным распределением между дочерними клетками приводит к появлению единичных клеток, совершенно лишенных окрашенных хлоропластов. Потомство (клон) этих клеток образует обесцвеченные зоны в листьях. Фенотип потомков по анализируемому признаку зависит от фенотипа материнского растения. У растения с зелеными листьями потомство нормальное. У растения с бесцветными листьями потомство имеет такой же фенотип - листья неокрашены. У материнского растения с пестрыми листьями потомки могут иметь все возможные фенотипы по указанному признаку - листья от полностью зеленых до полностью бесцветных, а также всевозможные переходные (промежуточные) формы (рис. 4.45). При этом
Рис. 4.45. Наследование признака «пестрые листья» у ночной красавицы: а - зеленые листья; б - пестрые листья; в - белые листья; I, II, III - результаты скрещивания разных материнских растений (а, б, в) с разными отцовскими
состояние листьев растений-потомков не зависит от фенотипа листьев отцовского растения.
Цитоплазматический тип наследования признаков, обусловленный митохондриальной ДНК (мтДНК, хромосома М), характеризуется своими особенностями прежде всего в связи с тем, что принципиальные структурно-функциональные параметры указанных органелл либо находятся под генетическим контролем ядерных генов, либо требуют адекватного взаимодействия элементов ядерного и митохондриального геномов (см., например, п. 4.3.1.3). В современной медицинской генетике существует самостоятельный раздел - митохондриальные наследственные болезни.
4.3.7. ФЕНОТИП ОРГАНИЗМА. РОЛЬ НАСЛЕДСТВЕННОСТИ И СРЕДЫ В ФОРМИРОВАНИИ ФЕНОТИПА
Любой организм характеризуется наличием генотипа и фенотипа (см. п. 1.3), что связано с двумя обстоятельствами, необходимыми для земной белково-нуклеиновой жизни. Во-первых, существование живых форм возможно лишь в их взаимодействии со средой обитания - источником энергии и «строительных» (пластических) материалов. Во-вторых, эволюционно отобранная видоспецифичная биологически целесообразная, т.е. гарантирующая выживание и размножение, информация передается в ряду поколений. Это происходит в мире жизни обязательно, так как смена поколений - необходимое условие эволюции. Задача организации отношений со средой жизни в каждом поколении решается благодаря наличию фенотипа. Задача наработки новой, сохранения и передачи в ряду поколений видоспецифичной биологической (генетической) информации решается благодаря наличию генотипа. Напомним, что генотип эукариотического организма - это совокупность всех генов или, что в функционально-генетическом плане более точно, аллелей структурных (экспрессируемых, кодирующих аминокислотные последовательности полипептидов) генов, а также сайтов (нуклеотид-ных последовательностей) ДНК с другими функциями в диплоидном наборе хромосом. Фенотип организма - совокупность признаков, свойств и качеств отдельно взятой особи конкретного биологического вида. Благодаря этим признакам, свойствам и качествам особь осуществляет необходимое для жизнедеятельности организмов данного вида взаимодействие со средой обитания.
В фенотипе биоинформация представлена в ее актуализированной, т. е. участвующей в процессах жизнеобеспечения непосредственно,
форме - прежде всего в виде ферментов, транспортных, структурных и других функциональных разновидностей белков. Биоинформация, представленная в генотипе, в обеспечении процессов жизнедеятельности прямо не участвует. Перевод биоинформации из «потенциальной» в «действующую» форму связан с формированием на основе определенного генотипа соответствующего этому генотипу фенотипа. Этот процесс осуществляется при активном участии и модифицирующем влиянии среды в ее широком понимании (см. п. 4.3.1.1).
Рассматривая соотносительную роль наследственности (генотип) и факторов среды (эпигенетических, внегенетических факторов) в оформлении фенотипа особи, следует исходить из сути такого генетического понятия, как норма реакции (см. п. 4.1.1), представлений о системном принципе организации и функционирования генома (см. п. 4.3.3.4) и генотипа (см. п. 4.3.1.1), что выражается в зависимости развития отдельных фенотипических признаков (см. п. 4.3.1) и фенотипа в целом не только от наличия соответствующих генов с присущими им свойствами (см. п. 4.3.1.1), но и от конкретных форм взаимодействия аллельных (см. п. 4.3.1.2) и неаллельных (см. п. 4.3.3.1) генов.
В отличие от классической (домолекулярной) генетики, современная генетика располагает сведениями о том, что многие сайты ДНК, не кодируя аминокислотные последовательности полипептидов, выполняют регуляторные, конценсусные, сервисные и другие функции (см. п. 4.3.3.2). Такие сайты участвуют в процессе формирования фенотипа, влияя на параметры транскрипции и трансляции структурных генов (см. пп. 2.4.5.5-а, 4.3.3.2), пост(после)транскрипционные (см. п. 2.4.5.5) и пост(после)трансляционные (см. п. 2.4.5.6) процессы. В генотипе особи присутствуют также гены, не определяющие развитие конкретных фе-нотипических признаков. В их функцию входит организация своеобразных «координатных сеток» или морфогенетических полей. Эти «сетки» или поля содержат позиционную информацию, благодаря чему клетки определяют свое положение в строящемся организме и, таким образом, осуществляют необходимую для оформления биологически зрелого или дефинитивного фенотипа траекторию развития (см. п. 4.3.3.2). Экзон/интронная организация (см. п. 2.4.5.5) и другие молекулярно-генетические особенности структуры генов эукариот, например наличие у гена нескольких промоторов, дают различный или несовпадающий по степени выраженности фенотипический результат мутаций в разных участках одного и того же гена (см. п. 5.2.2.3-в, пример с муковисцидо-зом), явления генокопирования и фенокопирования делают отношения между генотипом и фенотипом еще более сложными.
По мере накопления новых знаний, особенно в области функциональной геномики (см. пп. 1.1 и 2.4.3.4-д), при условии геномной паспортизации (см. предисловие, геномное тестирование или портретирование) населения представления о закономерностях оформления фенотипа организма на основе определенного генотипа будут приобретать все большую определенность. Учитывая интересы практического здравоохранения, развитие биомедицинской науки в названном направлении в высшей степени желательно. С одной стороны, это важно постольку, поскольку современная медицина располагает методами лечения и/или предотвращения (профилактики) развития нежелательного «проблемного» фенотипа при некоторых формах наследственной патологии. В указанных условиях возрастает значение точной диагностики генетического «дефекта». Так, наряду с классической фенилкетонури-ей I типа (см. п. 5.2.2.8, мутация гена с локализацией на длинном плече хромосомы 12, приводящая к функциональному дефициту фермента фенилаланин-4-гидроксилазы, участвующего в обмене соответствующей аминокислоты, поступающей в организм с пищей), известен ряд генотипических вариантов, дающих сходный патологический фенотип (явление генокопирования). Речь, в частности, идет о мутации гена с локализацией на коротком плече хромосомы 4, приводящей к дефициту фермента дигидроптеридинредуктазы - атипичная фенилкетонурия II типа. Известно, что ведение детей с неблагоприятным генотипом на безфенилаланиновой диете эффективно препятствует оформлению патологического фенотипа в случаях фенилкетонурии I типа, но практически не предотвращает развитие тяжелых неврологических нарушений (умственная отсталость, вплоть до идиотии) при фенилкетонурии II типа. С другой стороны, ускоренными темпами идет накопление знаний о генотипических основах мультифакториальных болезней, в развитии которых значительное место занимает наследственная предрасположенность (см. п. 5.2.2.8). И в этом секторе практической медицины эффективность лечебно-превентивных (профилактических) мероприятий зависит от точной информации о генетической конституции индивидуума (см. п. 5.2.2.3-в, г).
Важная роль в развитии фенотипа принадлежит факторам среды. Наряду с факторами генотипической среды (см. п. 4.3.1.1, среда 1-го порядка), о которых речь шла выше, свой вклад в оформление фенотипа вносят факторы внутренней среды (см. п. 4.3.1.1, среда 2-го порядка, в которой на период внутриутробного развития организма целесообразно выделять внутреннюю среду его самого - 2а и внутреннюю среду мате-
ринского организма, вынашивающего плод, - 2б) и, наконец, факторы внешней среды (см. п. 4.3.1.1, среда 3-го порядка).
Можно заключить, что переход «потенциальной» биоинформации генотипа в «актуализированную» действующую биоинформацию фенотипа - сложный процесс. Это обстоятельство при отсутствии знаний о химической природе вещества наследственности, функционально-генетическом многообразии нуклеотидных последовательностей ДНК и тонкой структуре генов, о сути пост(после)транскрипционных и пост(после)трансляционных событий, о механизмах влияния на функциональную активность генов факторов среды 2а и 2б порядка (сигнальные молекулы в форме транскрипционных и ростовых факторов, гормонов, цитокинов и других биологически активных веществ-регуляторов, определяющих поведение клеток, а также узнающие эти молекулы клеточные рецепторы, молекулы-участницы внутриклеточных сигнальных путей) способствовало появлению в классической генетике, опиравшейся практически исключительно на результаты анализа закономерностей наследования признаков, таких важных, в том числе для практики МГК (см. п. 5.2.2.8), связывающих генотип и фенотип организма генетических понятий, как «пенетрантность», «экспрессивность» и «генетическая гетерогенность» (см. п. 4.3.1.1), «широкая» или «узкая» норма реакции (см. п. 4.1.1). Только сейчас, благодаря успехам молекулярной генетики и клеточной биологии, появляется возможность представить себе механизмы, составляющие основу перечисленных генетических феноменов (см. п. 4.3.3.1).
Использование генотипической биоинформации в целях структурно-функционального обеспечения процессов жизнедеятельности путем ее перевода в фенотипическую биоинформацию осуществляется постоянно на всем протяжении жизни особи. Вместе с тем наиболее энергично это происходит в связи с формированием дефинитивного (состояние биологической, а для человека и социальной, зрелости) фенотипа организма. Для млекопитающих, в том числе человека, - это внутриутробный и ранний постнатальный периоды онтогенеза.
4.3.7.1. Участие генетических и внегенетических (средовых, эпигенетических) факторов в развитии фенотипических признаков пола особи
На ранних стадиях развития земной жизни размножение живых форм происходило бесполым путем. И сейчас бесполым способом размножаются прокариоты. Типичный пример бесполого размножения у эукари-
от - митотическое деление клетки. Бесполое размножение встречается у современных эукариотических организмов среди простейших одноклеточных форм, в том числе ведущих паразитический образ жизни (шизогония у малярийного плазмодия), а также среди низкоорганизованных многоклеточных (гидры, кольчатые черви). Бесполое размножение путем образования почек, стеблевых и корневых клубней, луковиц характерно для растений. В процессе исторического развития живых форм возник половой процесс, представляющий собой способ увеличить биоинформационное разнообразие потомства и таким образом расширить возможности действия естественного отбора. Представление о половом процессе дает явление конъюгации, например, у инфузорий. Оно заключается во временном соединении двух особей с целью обмена (рекомбинации) наследственным материалом. В результате образуются организмы, генетически отличные от каждого из участников конъюгации. Затем такие особи размножаются бесполым путем. Поскольку при конъюгации количество инфузорий не увеличивается, говорить о размножении в прямом смысле нет оснований. Можно заключить, что первоначально половой процесс не решал задачи размножения. Порядка 3 млрд лет назад в ходе эволюции возникает половое размножение, типичные черты которого - образование половых клеток или гамет (рис. 4.46) и оплодотворение. Соответственно жизненный цикл представителей видов, размножающихся половым путем, представлен двумя фазами (рис. 4.47) - диплофазой (половозрелая особь, соматические клетки, характеризующиеся диплоидным или 2n числом хромосом и 2с количеством ДНК) и гаплофазой (гаметы, характеризующиеся гапло-
Рис. 4.46. Половые клетки: а - яйцеклетка; б - сперматозоиды: 1 - цитоплазма; 2 - ядро; 3 - хроматин ядра; 4 - шейка; 5 - жгутик; 6 - головка
Рис. 4.47. Фазы жизненного цикла многоклеточных животных
идным или n числом хромосом и с количеством ДНК). В основе процесса образования половых клеток (гаметогенез) лежит особая форма клеточного деления - мейоз, обеспечивающая доведение числа хромосом до гаплоидного. Одновременно первое деление мейоза, благодаря кроссинговеру в профазе и независимому расхождению негомологичных хромосом материнского и отцовского происхождения в анафазе (см. п. 4.3.4), служит эффективным инструментом комбинативной ге-нотипической изменчивости (см. п. 4.1.1). Оплодотворение, еще один инструмент комбинативной генотипической изменчивости, заключается в слиянии двух половых клеток с восстановлением типичного для соматических клеток особей соответствующего вида диплоидного числа хромосом (см. п. 4.3.4, кариотип).
Для видов, размножающихся половым путем, типичен половой диморфизм (рис. 4.48), который заключается в наличии различающихся по фенотипу женских и мужских особей или самок и самцов.
Половой диморфиз как биологическое явление проявляет себя в патологии людей. В истории здравоохранения особенности биологии женщин определили возникновение в свое время отдельной медицинской дисциплины - гинекологии. На наших глазах укрепляются позиции медицинской дисциплины, которая своим возникновением обязана особенностям биологии мужчин, - андрологии. Многое из области интересов гинекологии, с одной стороны, и андрологии, с другой, связано со специфической ролью, которую играют женщины и мужчины в осу-
Рис. 4.48. Половой диморфизм у людей. Характерны различия: 1 - по кариоти-пу и главному половому гормону; 2 - структуре волос и характеру оволосения; 3 - строению гортани; 4 - развитию молочных желез; 5 - развитию мускулатуры; 6 - строению половых органов; 7 -распределению жировой ткани; 8 - показателям роста длинных трубчатых костей. Половой диморфизм проявляется также на уровне гамет (см. рис. 4.47)
ществлении детородной функции. Вместе с тем медицина располагает фактами, свидетельствующими, например, о различиях (при одинаковой дозировке) в терапевтическом эффекте, спектре и выраженности побочных реакций, а также об особенностях фармакодинамики и фармако-кинетики (параметры, характеризующие метаболизм и, следовательно, «судьбу» медицинского препарата в организме больного) ряда лекарственных средств в зависимости от того, идет ли речь о пациенте или о пациентке. Злокачественные новообразования одной органной локализации чаще встречаются у женщин, тогда как другой - у мужчин. При этом речь не идет об органах репродуктивной системы (молочная железа, матка, яичники - у женщин, предстательная железа - у мужчин). Различия в средней продолжительности жизни мужчин и женщин - реальность, хотя конкретные цифры колеблются от популяции к популяции. Так, в США (1979) и Франции (1980) они превышали 8 лет, в Греции (1981) составили 4,5 года, в Болгарии и Японии (1981) были равны 5,5 годам - все в пользу женщин.
Современная живая природа дает примеры различных способов определения пола. При прогамном способе, например, пол организма определяется особенностями структуры яйцеклетки, которая была оплодотворена сперматозоидом. Так, у коловраток крупные яйцеклетки дают самок, более мелкие - самцов. При эпигамном способе мужской или женский пол определяется факторами внешней среды, например, температурой в кладке яиц: у многих видов черепах при температуре ниже 27 °С развиваются только самцы, свыше 30 °С - только самки, в интервале 27-30 °С - самцы и самки. У большинства видов животных, размножающихся половым путем, в процессе исторического развития закрепились разные варианты «надежного» генотипического способа определения пола. При эусингамном варианте (пчелы, муравьи), самцы первично гаплоидны, поскольку они развиваются из неопло-дотворенных яйцеклеток, тогда как самки диплоидны. Напомним, что в процессе развития соматические клетки самцов таких животных становятся диплоидными. У плодовых мух наблюдается «балансовый» вариант генотипического способа, при котором пол определяется отношением числа хромосом Х к числу наборов аутосом (А). Если указанное отношение равно единице (ХХ/2А), развивается самка, при значении отношения 0,5 (XY/2A) - самец, особи с кариотипом ХХ/3А (отношение меньше единицы, но больше 0,5) или XY/ЗА (отношение меньше 0,5) - интерсексы. Исследования, выполненные также на плодовых мухах, заставляют думать, что генотипический механизм формирования ком-
плекса фенотипических признаков женского или мужского пола у них более сложен. Так, у дрозофил на хромосоме 3 обнаружен локус tra или t с геном, изменяющим пол организма в сторону мужского вне зависимости от значений отношения числа хромосом X и числа наборов аутосом. Особи как с генотипом XY/2Att, так и с генотипом XX/2Att - фенотипи-чески самцы, однако первые плодовиты (образуют сперматозоиды), а вторые стерильны. Можно заключить, что для сперматогенеза хромосома Y необходима.
Предположительно с точки зрения интересов эволюционного процесса оптимален хромосомный вариант, получивший повсеместное распространение среди высокоорганизованных многоклеточных животных (амниоты - птицы, млекопитающие, включая человека), однако встречающийся у анамниа (земноводные) и среди членистоногих (некоторые виды клопов). Для этого варианта генотипического способа характерно, что один из полов (гомогаметный) образует одинаковые гаметы, тогда как второй (гетерогаметный) - разные. У млекопитающих гомогаметны женские особи, имеющие в кариотипе пару одинаковых половых хромосом (XX), а гетерогаметны - мужские особи, имеющие в кариотипе пару разных половых хромосом (XY). У земноводных и птиц гомогаметны мужские особи (пара одинаковых половых хромосом - ZZ), тогда как гетерогаметны женские особи (пара разных половых хромосом - ZW). Легко видеть, что при таком варианте генотипического способа определения один из полов (гомогаметный) по паре половых хромосом характеризуется как гомозиготный, тогда как второй (гетеро-гаметный) - как гетерозиготный.
Предположительно в генотипическом определении пола у парамеций принимает участие один локус, т. е. задействован моногенный механизм. При этом есть основания думать, что у парамеций особи одного пола по указанному локусу гомозиготны, а другого - гетерозиготны.
Хромосомный (или более редкий моногенный, см. пример с парамециями) вариант определения пола оптимален для процесса эволюции потому, что он, благодаря гомогаметности и гетерогаметности полов и отношению разнополых особей в период активного размножения 1:1 (у человека таковым отношение между мужскими и женскими особями становится в юношеском возрасте, хотя среди новорожденных на 100 девочек приходится в среднем 106 мальчиков), обеспечивает:
• максимальную вероятность встречи разнополых особей в целях репродукции;
• наиболее высокий уровень разнообразия генетической информации родителей, привлекаемой для создания генотипов потомков в каждом очередном поколении;
• поддержание оптимальной численности особей в популяциях. Известны виды, у которых гетерогаметный пол представлен особями
с парой разных половых хромосом - тогда как особи гомогаметного пола имеют одну половую хромосому - Х0. Встречаются также иные варианты. Так, у клопов из рода Protenor самки - ХХ, самцы - Х0. Напомним, что среди людей лица с моносомией по паре половых хромосом (ХО) характеризуются патологическим фенотипом, в целом сдвинутым в женскую сторону (синдром Шерешевского-Тернера).
Наличие в человеке биологического, социального и духовного начал (см. предисловие) объясняет, почему проблема пола, отнесенная к людям, имеет много аспектов (табл. 4.4).
Таблица 4.4. Пол человека: биосоциальные факторы определения пола у людей - генетика и среда
У млекопитающих, в том числе у человека, эмбриональная закладка половых желез (гонады) в виде парной структуры (половой валик) поначалу не имеет признаков дифференциации по мужскому (семенник) или женскому (яичник) типу, т.е. является индифферентной (бипо-тенциальной). Направление дифференциации зависит от комбинации пары половых хромосом в зиготе, т.е. от того, была ли оплодотворена яйцеклетка сперматозоидом с хромосомой Х или с хромосомой Y. В присутствии в кариотипе зиготы хромосомы Y (комбинация пары половых хромосом XY) развитие происходит по мужскому типу, что связано с расположением на указанной хромосоме гена SRY (Sex determining
Region Y, Ур11.31-32). Названный ген контролирует образование транскрипционного фактора, который благодаря сродству к промоторам активирует гены, необходимые для развития семенника. Поэтому ген SRY называют также TDF (Testis Determining Factor). У человека экспрессия гена SRY (TDF) начинается на стадии зиготы. На хромосоме Y расположены также гены AZF (Azoospermia Factor, Уq11) и H-Y антигена. Первый участвует в регуляции сперматогенеза: его мутации ведут к снижению продукции сперматозоидов вплоть до полного подавления. Фе-нотипически это проявляется в олигоспермии или азооспермии, т. е. в недостаточном количестве или полном отсутствии спермиев в семенной жидкости. Второй обусловливает синтез белков клеточных оболочек. В настоящее время его участие в генетическом контроле развития феноти-пических признаков по мужскому типу оспаривается.
В развитии эмбриональной закладки гонад по мужскому типу участвует ряд аутосомных генов. Среди них гены АМН или MIS (Anti-Mullarian Hormone или Mullerian Inhibiting Substance, 19р13.2-3), SOX9 (Sox-related HMG-box-containing gene, расположен на хромосоме 17) и WT1 ( Wilm s Tumore-associated gene 1, расположен на хромосоме 11). Экспрессия двух последних генов происходит в клетках закладки половых желез на индифферентной стадии. Мыши-мутанты с «выключенным» геном WT1 нежизнеспособны: у них не развиваются гонады и почки. На хромосоме Х в зоне DSS (Dosage-Sensitive Sex reversal) расположен ген DAX1 или АНС (Adrenal Hypoplasia Congenita), который репрессируется (подавляется) в условиях активации гена SRY, т. е. в случае начала развития гонад по мужскому типу. При отсутствии активности гена SRY ген DAX1 активно функционирует, что необходимо для развития гонад по женскому типу (яичники). В настоящее время названная функция гена DAX1 подвергается сомнению. Ген АМН ответствен за редукцию мюл-леровых протоков, не нужных для развития мужской репродуктивной системы, так как семявыносящие пути и ряд других структур образуются из вольфовых протоков.
На 6-й неделе внутриутробного развития гонады дифференцированы по полу. Если к названному сроку беременности развитие гонад по мужскому типу не началось, «по умолчанию» развитие комплекса половых признаков сдвигается в направлении женского фенотипа. Описанные события, ведущая роль в которых принадлежит, видимо, ге-нотипическим факторам (наличие в генотипе соответствующих генов и их взаимодействие, особенности генотипической среды или среды 1-го порядка), составляют содержание периода первичной детерминации
пола развивающегося организма. Главный результат этого этапа состоит в дифференциации первоначально индифферентной эмбриональной закладки половых желез в семенники или яичники.
Начиная с 7-й недели внутриутробного развития, когда гонады приобретают структуру либо семенника, либо яичника, начинается период вторичной детерминации пола. В этом периоде главную роль играют гормоны. Так как гормоны являются сигнальными молекулами, клеточные оболочки должны иметь молекулы-рецепторы, специфически узнающие гормон и запускающие соответствующие внутриклеточные сигнальные пути. Образование гормонов и рецепторов находится под генетическим контролем.
Главный мужской половой или андрогенный гормон тестостерон образуется в семенниках клетками Лейдига. Для развития полноценного фенотипического комплекса мужского типа необходима также активность гена АМН, контролирующего продукцию андрогенного гормона, подавляющего развитие мюллеровых протоков. Активация названного гена обусловлена продуктом активности гена SRY. Под влиянием тестостерона из вольфовых протоков образуются мужские внутренние половые органы, такие, как сямявыносящие канальцы, индуцируется развитие семенных пузырьков и придатка яичка (эпидидимис), а также формирование на основе мочеполового синуса таких наружных половых органов, как простата, половой член, мошонка. Действие тестостерона требует наличия в клеточных оболочках белка-рецептора, образование которого контролирует ген AR (Xq11). Оба названных выше андроген-ных гормона необходимы для развития по мужскому типу (маскулинизирующее действие) экстрагенитальных органов и тканей-мишеней, что обусловливает половой диморфизм центральной нервной системы, мускулатуры, пропорций и размеров тела, внутренних органов, метаболизма и т.д.
При нарушении образования в организме андрогенных гормонов или рецепторов к ним наблюдаются отклонения в развитии фенотипическо-го комплекса мужского пола (сдвиг в сторону фенотипического комплекса женского пола) разной степени - от гипоспадии (относительно низкое расположение мочеиспускательного канала) легкой степени и/ или крипторхизма (неопущение яичка в мошонку) до оформления выраженного женского фенотипа (синдром тестикулярной феминизации или Морриса). Причиной этих отклонений могут быть как мутации соответствующих генов или нарушение межгенных взаимодействий (генотипическая среда или среда 1-го порядка), в том числе в формате
явления генокопирования, так и изменения в ходе морфогенетических процессов в связи с особенностями среды 2а (внутренняя среда развивающегося организма) и 2б (внутренняя среда организма женщины, вынашивающей ребенка), в том числе спровоцированными условиями внешней среды или среды 3-го порядка - явление фенокопирования. В период вторичной детерминации пола по мужскому варианту определенную роль играют и женские половые гормоны - эстрогены. Так, они необходимы для созревания костной ткани, а также обеспечивают некоторые качественные характеристики сперматозоидов.
На настоящий момент можно думать, что количество генов, вовлеченных в развитие и обеспечение функций яичек и простаты, составляет не менее 1200, яичника - 500, матки - 1800. В таких условиях трудно говорить о моногенном принципе генотипического обеспечения развития и функционирования фенотипического комплекса признаков пола. Именно поэтому в настоящее время принцип генетической регуляции половой дифференцировки у человека нередко определяют как сетевой, подразумевая под этим, что действие многих генов-участников взаимосвязано и взаимообусловлено.
Наряду с фенотипическими признаками, связанными с мужским или женским полом непосредственно, выделяют также признаки, зависящие от пола и ограничиваемые полом. В данном случае речь может идти, например, о генах раннего облысения, которые у мужчин ведут себя как доминантные, а у женщин как рецессивные. Известно, что не только у коров, но и у быков (нельзя исключить, что и у мужчин) есть гены, контролирующие возможность продукции молока, причем определенной жирности. У быков эти гены, однако, не экспрессируются.
Нельзя также забывать о том, что существует Х- и Y-сцепленное наследование признаков.
Вопросы для самоконтроля
1. В чем состоят свойства наследственности и изменчивости на разных уровнях организации жизни?
2. Перечислите формы изменчивости и дайте их краткую характеристику?
3. Является ли ген структурной или функциональной единицей генетического материала? Каковы свойства гена?
4. Что такое аллели генов, и каков механизм их возникновения?
5. Перечислите уровни организации генетического материала. В чем биологическое значение каждого уровня?
6. Что такое «кариотип», и чем он характеризуется?
7. Какие типы наследования существуют? Какие виды взаимодействия генов реализуются при каждом типе наследования?
8. Что такое «фенотип»? Какие наследственные и средовые факторы принимают участие в его формировании?
9. Что такое «пол»? Каковы уровни его формирования у человека?
10. Какие могут быть нарушения формирования пола у человека?