Биология: учебник: в 2 т. / под ред. В. Н. Ярыгина. - 2011. - Т. 2. - 560 с. : ил.
|
|
|
|
Глава 15. АНТРОПОГЕНЕЗ И ДАЛЬНЕЙШАЯ ЭВОЛЮЦИЯ ЧЕЛОВЕКА
15.1. МЕСТО ЧЕЛОВЕКА В СИСТЕМЕ ЖИВОТНОГО МИРА
Неограниченный прогресс в эволюции живой материи проявился в возникновении человека как биосоциального существа. Появление человека с его целенаправленной трудовой деятельностью качественно изменило облик планеты Земля и определило новое направление эволюции природы в целом. Будучи биологическим видом, человек обладает при этом социальной сущностью. Социальная сущность человека определяет его современное состояние и прогнозирует будущее. Биологическое в нем при этом становится его наследством, поэтому эволюция человека на современном этапе идет не столько по биологическим законам, сколько по законам развития человеческого общества. Биологическая эволюция представляет собой биологическую форму движения материи, а эволюция общества - социальную. Естественно, что социальная форма движения материи возникла лишь с появлением человека. Поэтому изучение вопросов антропогенеза - это не только биологическая, но и философская проблема.
Возникновение человека как социального существа происходило, с одной стороны, в связи с его принадлежностью к отряду приматов, обладающих большим количеством прогрессивных черт организации, а с другой - с совпадением во времени нескольких событий, связанных:
• с особенностями географического расселения предковых форм;
• с геологическими процессами;
• с адекватными им изменениями климатических условий на Земле в целом и в зонах их исходного обитания.
Существует мнение, что если бы в мезозойской эре не вымерли динозавры, то пресмыкающиеся до сих пор оставались бы самой прогрессивной группой животных. И не исключено, что наиболее эволюционно продвинутые некрупные хищные ящеры стенихозавры с относитель-
ным объемом головного мозга, превышающим этот показатель у всех остальных известных современных и вымерших пресмыкающихся в 6 раз, могли бы дать начало появлению разумных существ, отличных от человека.
Человек как биологический вид относится к отряду приматов
(рис. 15.1).
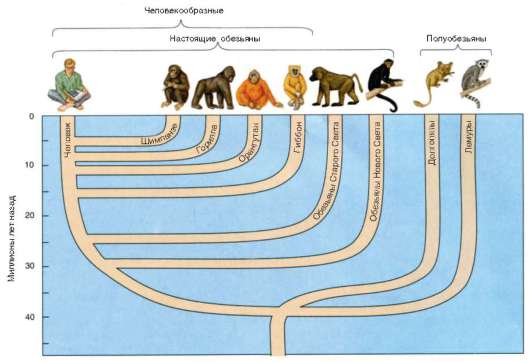
Рис. 15.1. Место человека в отряде Приматы и приблизительное время дивергенции приматов на основные систематические группы
15.2. МЕТОДЫ ИЗУЧЕНИЯ ПРОИСХОЖДЕНИЯ И ЭВОЛЮЦИИ ЧЕЛОВЕКА
Разные этапы эволюции человека требуют и разных подходов к их изучению. Происхождение приматов и их адаптивная радиация - чисто биологическая эволюция. Ее изучают в первую очередь методами палеонтологии и сравнительной морфологии. Так как возникновение рода Homo сопровождается появлением элементов материальной культуры, а к действию элементарных эволюционных факторов подключается действие социального фактора, этот этап антропогенеза изучают также методами археологии. В дальнейшей эволюции рода Homo доминируют
социальные факторы, поэтому методологические подходы к изучению разных этапов антропогенеза различны.
Изучение первых этапов эволюции человека сталкивается с необходимостью датировки палеонтологического материала и элементов материальной культуры. Для определения абсолютного возраста ископаемых останков человека и его предков широко используют физические методы, в частности, радиометрические. С помощью масс-спектрометрии определяют изотопный состав изучаемого объекта и по соотношению элементов с учетом периода полураспада входящих в его состав радиоактивных изотопов выявляют возраст образца. Ископаемый костный материал содержит в своем составе минеральные компоненты и белок коллаген, разрушающийся чрезвычайно медленно. На этом основан коллагеновый метод абсолютной датировки ископаемых остатков: чем меньше коллагена содержится в образце, тем более велика его древность.
В антропологии при изучении происхождения и эволюции человека широко применяют также методы иммунологии, биохимии, молекулярной биологии и цитогенетики. В связи с огромной важностью этих методов остановимся на них подробнее. Для определения прямого родства организмов друг с другом используют иммунологический метод, основанный на изучении иммунных реакций антиген-антитело. Его можно применять для изучения степени родства не только современного человека с человекообразными обезьянами, но и ныне живущих видов с ископаемыми. Для этого следовые количества белка, извлекаемые из костей ископаемых форм, используют для получения антител, которые и применяют в иммунных реакциях с белками современных видов. Из современных человекообразных обезьян к человеку иммунологически наиболее близок шимпанзе, наиболее далеко от человека отстоит орангутан.
Иммунологическим методом было обнаружено, что белки рамапи-тека, человекообразной обезьяны Южной Азии (абсолютный возраст 13 млн лет), более сходны с белками орангутана, чем человека и шимпанзе. Эти данные вместе с результатами морфологических и палеонтологических сопоставлений заставили отказаться от представления о том, что рамапитек является прямым предком человека, и связать его с эволюционной линией орангутана. Из этого следует, что разделение человеческой линии эволюции с африканскими человекообразными обезьянами произошло значительно позже, чем 13 млн лет назад.
Биохимическим методом определяют аминокислотный состав белков, например, гемоглобина, у организмов, находящихся в разной степени родства друг с другом. Естественно, что у близкородственных организмов гомологичные белки имеют большее сходство в аминокислотных последовательностях, чем у организмов, находящихся в более отдаленном родстве (рис. 15.2).
Молекулярно-биологический метод основан на сопоставлении нуклеотидных последовательностей молекул ДНК организмов разной степени родства. Мера сходства двух таксонов соответствует мере их родства. Поэтому организмы, имевшие общего предка в недалеком прошлом, будут более сходными друг с другом, чем имевшие его очень давно. В антропологии чаще используют митохондриальную, а не ядерную ДНК, поскольку она присутствует в клетках в большом количестве копий и некоторое ее количество почти всегда можно обнаружить в ископаемом материале. Секвенирование и сравнение в процессе гибриди-
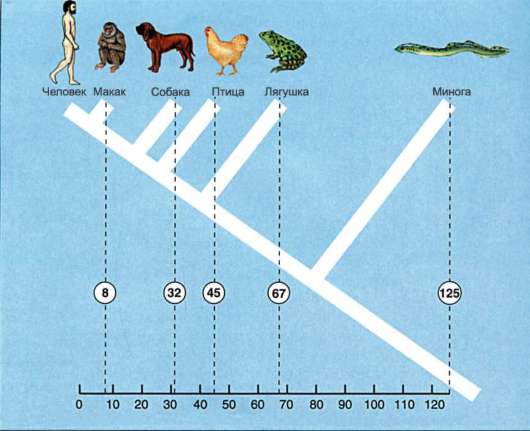
Рис. 15.2. Число отличий аминокислотных остатков в молекулах гемоглобина разных видов позвоночных по сравнению с человеком
зации молекул ДНК разных видов современных приматов, человека и ископаемых предковых форм дает возможность определить степень их родства и примерное время расхождения соответствующих филогенетических ветвей. Естественно, что оценивать степень родства и давность происхождения можно лишь по накоплению нейтральных и полезных мутаций (изменений в нуклеотидных последовательностях) в геноме, так как вредные мутации быстро элиминируются из генофондов.
Расчеты показывают, что при нейтральности мутаций скорость их накопления постоянна только при измерении времени в числе поколений, а не в годах. Так как продолжительность жизни поколений у различных видов различна, то и скорости накопления различий нуклеотидных последовательностей будут более велики у короткоживущих видов по сравнению с долгоживущими. Кроме того, фактическая скорость эволюции может значительно варьировать в различные временные интервалы в различных группах и по разным признакам. В определении применимости методов молекулярной биологии имеет значение возможность возникновения конвергентного сходства молекул, причем вероятность его повышается с увеличением изучаемых временных интервалов. Эволюция генов и белков часто может опережать реальное расхождение популяций, в первую очередь за счет адаптивного генетического полиморфизма. Однако молекулярно-биологические методы применимы для оценки родства и времени дивергенции в качестве приблизительных «молекулярных часов» при сравнении средних скоростей замен нуклеотидов в ДНК в целом и аминокислот во многих белках за длительные интервалы времени. Гибридизация ДНК человека и шимпанзе показала, что момент
дивергенции их эволюционных ветвей наступил 6,5-7 млн лет назад (рис. 15.3).
Биомолекулярный подход - лишь один из путей определения эволюционных расстояний, который работает наряду с классическими методами палеонтологии и антропологии, причем в результатах при этом возможны серьезные расхождения. Так, при изучении скелета человека, обнаруженного на территории Эквадора, по данным радиоу-
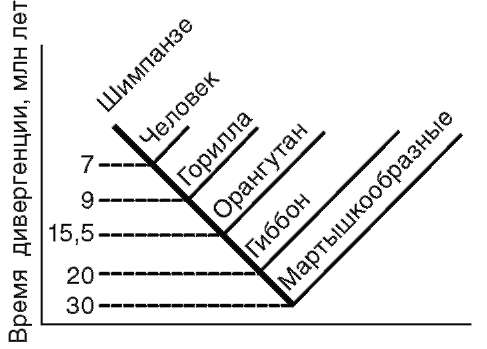
Рис. 15.3. Среднее время дивергенции высших приматов по часам ДНК
глеродного и аминокислотного анализа его возраст оценен в 28 тыс. лет. При использовании же коллагенового анализа возраст того же скелета оказался не более чем 2,5 тыс. лет.
Однако между эволюцией структуры генома в виде накопления генных мутаций и морфофизиологической эволюцией часто нет прямой зависимости (рис. 15.4). Это может быть связано с тем, что в формировании практически всех сложных фенотипических признаков принимают участие различные генные системы. Таким образом, скорость эволюции белков у двух разных родственных видов может быть одинакова, а скорость эволюции в целом, оцененная по комплексу фенотипических признаков, при этом оказывается различной. Сравнение аминокислотных последовательностей белков шимпанзе и человека привело к выводу, что
около 99% их белков абсолютно идентичны. Из этого следует, что и структурные гены человека и шимпанзе сходны в наибольшей степени. С чем же связаны столь значительные морфофизиоло-гические отличия обоих видов? Можно предполагать, что это зависит от различного распределения белков в клетках организма в процессе развития, что, в свою очередь, определяется различиями программы считывания сходной наследственной информации во времени и пространстве.
Интересно, что вследствие вырожденности генетического кода белки у родственных организмов могут нередко отличаться даже меньше, чем ДНК.
Поскольку изучение хромосомного материала возможно только у ныне живущих организмов, применение цитогене-тического метода ограничено современным человеком и человекообразными обезьянами.
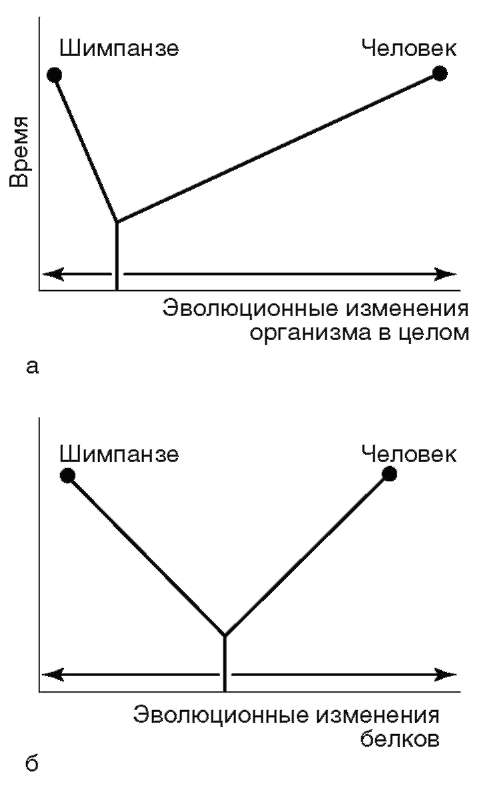
Рис. 15.4. Соотношение скорости морфофизиологической (а) и молекулярной (б) эволюции
Дифференциальная окраска хромосом позволяет не только сопоставлять хромосомы разных видов приматов и человека и изучать хромосомный полиморфизм современного человека, но и решать некоторые вопросы эволюции.
Выяснено, что кариотип человекообразных обезьян отличается по числу хромосом от кариотипа человека на одну пару (23 пары хромосом человека и 24 пары шимпанзе). У человека и шимпанзе практически идентичны 13 пар хромосом. Хромосома 2 человека точно соответствует двум соединенным хромосомам шимпанзе, а остальные хромосомы отличаются друг от друга незначительно. Так, хромосома 5 шимпанзе соответствует такой же хромосоме человека, но небольшой ее перицентрический участок инвертирован на 180° по сравнению с человеческой хромосомой. Инверсии такого рода обнаружены в кариотипах человека и шимпанзе еще в восьми хромосомах. Хромосома 9 человека имеет большие размеры по сравнению с соответствующей хромосомой шимпанзе, а хромосома 12 - несколько короче. Таким образом, наибольшие различия генетического материала человека и человекообразных обезьян касаются не структурных генов, а организации хромосом (см. также п. 4.3.2).
Эти данные вместе с указаниями на сходство белков человека, шимпанзе и других человекообразных обезьян свидетельствуют об их значительной эволюционной близости. Применение FISH-метода изучения хромосом позволяет обнаружить очень высокую степень соответствия физико-химических свойств генетического материала человека и его современных ближайших родственников. Существенные отличия обнаружены, однако, в его пространственной организации в ядрах интерфазных клеток. Действительно, различными оказываются точки прикрепления деконденсированных хромосом к внутренней поверхности ядерной мембраны (см. п. 2.4.3.1), их пространственная организация и взаимное расположение хромосом в ядре клетки, что само по себе может проявляться в различиях интенсивности и последовательности реализации сходной генетической информации.
Использование сравнительно-эмбриологического метода при изучении пренатального и раннего постнатального онтогенеза человекообразных обезьян и человека приводит к выводу о том, что эмбриогенез человека характеризуется выраженными признаками неотении: закладки и ранние этапы развития черепа человека и обезьян протекают практически идентично. Позже наблюдается выраженная аллометрия ростовых процессов (см. п. 8.3.4): у обезьян скорость роста элементов лицевого черепа резко опережает увеличение размеров мозгово-
го, в то время как у человека рост его мозгового отдела происходит с существенно большей скоростью по сравнению с лицевым (рис. 15.5). У человека в постнатальном периоде позже зарастают роднички мозгового черепа, в результате чего оказывается возможным интенсивный
Рис. 15.5. Аллометриче-ский рост черепа в процессе индивидуального развития шимпанзе и человека
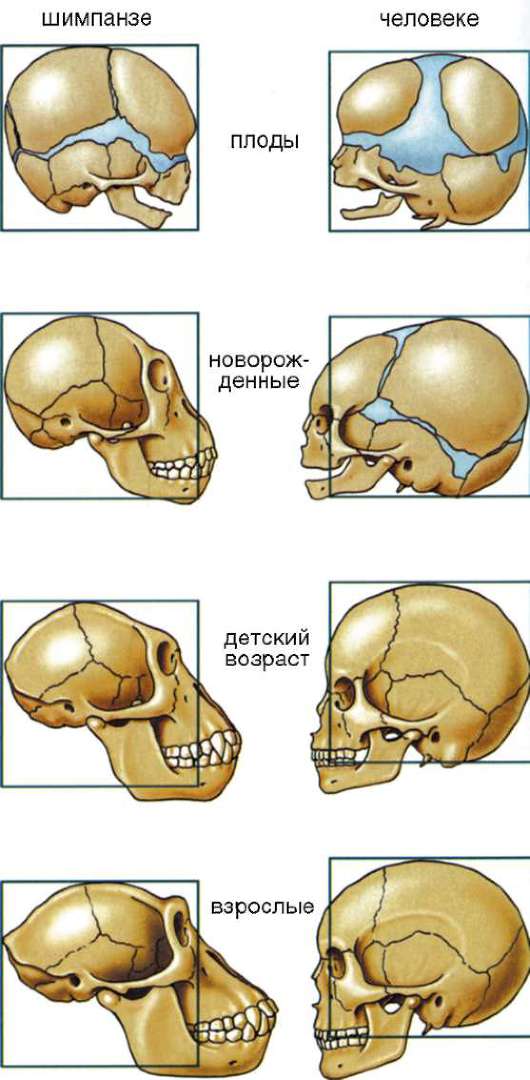
рост головного мозга и в раннем постнатальном периоде, в то время как у обезьян зарастание родничков и увеличение массы мозга приостанавливается значительно раньше. Вероятно, что указанная неравномерность ростовых процессов в формировании элементов черепа и мозга гоминид и их ближайших родственников обусловлена мутациями системных генов, регулирующих интенсивность клеточной пролиферации и перемещения клеток, и возникла уже на довольно ранних этапах дивергенции высших приматов.
Сопоставление кариотипов людей, происходящих из разных популяций, приводит к выводу о полиморфизме хромосом, в первую очередь по размерам гетерохроматиновых участков. Наследуемость индивидуальных вариаций хромосом и их неравномерное распределение в разных популяциях (в частности, расовые различия по размерам длинного плеча Y-хромосомы) делают возможным популяционно-цитогенетический подход в изучении эволюции современного человека.
15.3. ХАРАКТЕРИСТИКА ОТРЯДА ПРИМАТЫ
Отряд Приматы (Primates) включает более 200 видов ныне живущих полуобезьян и обезьян. Возникновение отряда приходится на конец мезозойской (начало - 230 млн лет назад, продолжительность - 163 млн лет) и начало кайнозойской эры (60-70 млн лет назад), когда в результате массового вымирания динозавров на земле оказались незанятыми множество экологических ниш, удобных для существования. Адаптивная радиация млекопитающих привела в это время к биологическому прогрессу этой группы животных, легко приспосабливающихся к разнообразным условиям обитания. В начале кайнозойской эры уже существовало множество видов, родов, семейств и отрядов млекопитающих, среди которых оказались и приматы. Эволюционно они связаны с более примитивным отрядом Насекомоядных. Ключевой адаптацией приматов стали приспособления к древесному образу жизни и передвижение по ветвям не с помощью когтей, как у большинства других древесных животных, а за счет конечностей хватательного типа с противопоставленным большим пальцем. Локтевая кость свободно вращается вокруг лучевой. Эти особенности при освобождении рук от участия в движении - предпосылка к использованию их впоследствии в трудовой деятельности.
Кончики пальцев приматов расширены, уплощены, а кожа, так же, как и кожа ладоней и стоп, покрыта узорами из бороздок и гребешков,
вдоль которых расположены окончания чувствительных нервных волокон - тактильные рецепторы, а также протоки потовых желез. Их секрет обеспечивает более тесный контакт с ветками и предметами, которыми обезьяна манипулирует. Усиление функции осязания кончиками пальцев у них приводит к исчезновению вибрисс - осязательных волосков на лицевой части головы. Это типичный пример органной субституции.
Расположение глазниц в передней части черепа и направленность глаз вперед обеспечивает бинокулярное зрение, необходимое для точной оценки расстояний между предметами и перемещения в трехмерном пространстве. В отличие от большинства других млекопитающих, не способных различать цвета, зрение у большинства приматов цветное. Это может быть связано с ранним освоением предками приматов древесного и дневного образа жизни, позволившего им эффективно сосуществовать с наземными рептилиями и примитивными млекопитающими и сохранить, в отличие от последних, цветное зрение. Действительно, большинство современных млекопитающих не способны различать цвета в связи с упрощением строения сетчатой оболочки глаза. Это обычно связывается с их первично ночным образом жизни, когда различение цветов не имеет смысла. Постоянная температура тела других млекопитающих позволяла им перейти к ночной активности и, таким образом, ослабить конкуренцию с пойкилотермными рептилиями, нуждающимися в более высокой температуре среды.
В развитии органов чувств у приматов ярко проявляется принцип компенсации функций: на фоне высокой степени развития осязательного и зрительного анализаторов существенно снижена роль обоняния. Одной из причин этого явления может быть тот же древесный образ жизни. Действительно, животные, обитающие на поверхности земли, встречаются с гораздо более широким спектром многообразных запахов, чем те, которые большую часть жизни проводят в кронах деревьев. В генотипе большинства млекопитающих, ориентирующихся в пространстве в значительной степени за счет обоняния, имеется более 1000 копий активных генов, ответственных за образование ольфак-торных (обонятельных) рецепторов, связывающих молекулы летучих веществ, и тем самым запускающих каскад сигнальных процессов, обеспечивающих восприятие запахов. У низших обезьян Южной Америки активны около 95% из них. У шимпанзе и гориллы транскрибируется до 50% этих генов, а у человека - не более 30%.
Уменьшение значения обоняния у приматов сопровождается существенной редукцией лицевой части черепа и увеличением размеров его
мозгового отдела. Вес и объем головного мозга обезьян в 2 раза и более превышает эти показатели у других млекопитающих соответствующего размера. Сочетание стереоскопического цветного зрения и хорошо развитого слуха с особенностями передней конечности как органа локомоции, осязания и манипуляции предметами и с увеличением относительного объема головного мозга в основном за счет разрастания коры и тех ее областей, с которыми связаны сложные формы движения, поведения и психики, стало предпосылкой усложнения поведения приматов. Практически все вышеперечисленные морфологические признаки приматов отчетливо видны на рис. 15.6, где изображен представитель полуобезьян, долгопят филиппинский (восточный). Приматы в своем абсолютном большинстве ведут групповой образ жизни; причем в группы входят разновозрастные и разнополые особи. Это способствует развитию сложных взаимоотношений в группах и стало предпосылкой дальнейшего развития мозга
и интеллектуальных способностей. Группой обезьяны легче защищаются от хищников, находят пропитание, мигрируют, охраняют свои территории, перенимают друг у друга полезные особенности поведения, молодые члены группы обучаются у более старых. Между членами одной группы возникают сложные взаимоотношения, поддержание которых требует не только высокой степени развитости морфологии коры полушарий головного мозга, но и ее физиологической пластичности. Сопоставление размеров коры различных видов обезьян с размерами групп, образуемых соответствующими видами, обнаружило прямую зависимость между этими показателями. Это связано с тем, что поддержание социальных отношений с большой группой особей требует одновременно увеличенных размеров и большей структурной и функциональной сложности центральной нервной системы. Групповой образ жизни с

Рис. 15.6. Долгопят восточный (Tarsius spectrum). Видны наиболее характерные признаки приматов: хватательные конечности и бинокулярное зрение
выраженной иерархией в группе требует также развития эффективной системы коммуникации. В результате у обезьян сформировалась наиболее сложная по сравнению с другими млекопитающими мимическая мускулатура, с помощью которой они способны легко выражать эмоции и передавать информацию друг другу.
Большинство приматов экологически пластичны, всеядны и способны легко переходить к использованию разных продуктов питания в разные периоды года и в разных условиях.
Плодовитость приматов чрезвычайно низка - обычно рождается один детеныш, беспомощный и нуждающийся в уходе на протяжении нескольких лет. Поэтому тесная связь между матерью и потомством сохраняется надолго. Это способствует проявлению еще одной очень характерной особенности поведения приматов - выраженного инстинкта подражания, который обеспечивает быстрое овладение потомством навыками поведения, адекватного меняющейся ситуации, и возможность передачи социального опыта в ряду поколений.
43 млн лет назад в геном предков обезьян встроился вирус, экспрессия генов которого приводит к образованию белков, входящих в состав его оболочки. Эти белки разрушают мембрану клетки-хозяина, облегчая проникновение вируса в нее. У высших приматов гены вирусного происхождения (Syncytin 1, Syncytin 2, Envv 1, Envv 2 и EnvPb 1) экспрессируют-ся в трофобласте и инициируют слияние клеток плаценты, разрушая их наружную мембрану, в результате чего образуется синцитиотрофобласт, и плацента становится гемохориальной. Такая плацента обеспечивает оптимальные условия транспорта веществ между организмом матери и плода. Кроме того, она защищает плод как от действия иммунной системы матери, так и от вирусов экзогенной природы.
Перечисленные черты отряда приматов стали необходимыми пре-адаптациями к возникновению человека как социального вида при появлении комплекса благоприятных условий.
В отряде приматов наиболее эволюционно продвинуто надсемейство гоминид, или человекообразных обезьян, к которому относятся как современные гиббоны, орангутаны, гориллы и шимпанзе, так и человек.
От других обезьян гоминиды отличаются наиболее сложным строением головного мозга, отсутствием хвоста, строением плечевого сустава, позволяющего максимальную свободу вращения передних конечностей и передвижения за счет брахиации - перебрасывания тела в подвешенном состоянии на руках в вертикальном положении. Для них характерно редкое оволосение тела, сложное поведение, позднее половое созрева-
ние и беременность, по длительности сопоставимая с беременностью у человека.
Возникновение человека именно в отряде Приматы не случайно, так как этот отряд обладает наибольшим количеством ароморфных черт строения и физиологии, необходимых для перехода на принципиально новый, более высокий уровень организации по сравнению с другими группами животного мира.
15.4. АДАПТИВНАЯ РАДИАЦИЯ ПРИМАТОВ И ОСВОЕНИЕ ИМИ ОТКРЫТЫХ ПРОСТРАНСТВ
Начиная с середины кайнозойской эры (25-6 млн лет назад) климат на Земле становился все более неустойчивым. Периоды потепления сменялись периодами значительного похолодания, причем каждое последующее похолодание было более отчетливым по сравнению с предыдущим.
Похолодания обычно сопровождаются образованием ледников на полюсах Земли в результате конденсации паров воды из атмосферы. При этом в приэкваториальных зонах климат становится более сухим. Это отражается и на изменениях биогеоценозов: зоны распространения влажных тропических лесов уменьшаются в размерах, а саванны, степи и полупустыни расширяются. Этим объясняется и периодическое изменение видового состава экологических систем. Во влажных тропических лесах биологический прогресс наблюдается обычно среди некрупных млекопитающих, обитающих на деревьях, а на открытых местностях процветают более крупные наземные формы и мелкие, ведущие роющий образ жизни.
В середине кайнозоя процесс бурной адаптивной радиации затронул и отряд приматов, в рамках которого выделилось надсемейство церко-питековых, питающихся листьями, цветами и насекомыми - низших узконосых обезьян - предков павианов, макак и мартышек, и гоминои-дов, или человекообразных обезьян - более крупных форм, питающихся преимущественно плодами деревьев и животной пищей. Уменьшение размеров тропических лесов привело к сокращению кормовой базы обезьян. Мелкие, подвижные, быстро размножающиеся церкопитеко-вые приматы в условиях усиления конкуренции стали вытеснять более крупных человекообразных обезьян из оставшихся лесных экосистем.
Первые представители человекообразных обезьян (надсемейство Hominoidea) появились в Африке около 25 млн лет назад. 16-17 млн лет
назад ареал гоминоидов расширился за счет появления сухопутной перемычки между Африкой и Евразией. Через нее они заселили также Южную Европу и Азию. В Азии представители этой группы дожили до наших дней (орангутаны и гиббоны), а в Европе вымерли около 7 млн лет назад. В Африке в этот период также произошел биологический регресс этой группы, до настоящего времени дожили лишь три африканских вида - горилла, шимпанзе и карликовый шимпанзе (бонобо), но именно в этой части ареала гоминоидов возникли первые представители семейства гоминид (Hominidae), ведущие к отделению человеческой линии в генеалогическом древе приматов.
7-6 млн лет назад к такому фактору среды, как похолодание, присоединился еще один - начавшееся образование Восточно-Африканской рифтовой системы - гигантского разлома земной коры с образованием горных хребтов, располагающихся меридианально, и системы Великих Восточно-Африканских озер: Рудольфа, Альберта, Виктории, Танганьики, Эдуарда, Ньяса, Мвару и др. Хребты отделили Восточную Африку от Западной и перекрыли проникновение влажных западных ветров в восточном направлении, сделав Восточную Африку еще более сухой и изолировав местных гоминид в саваннах и полупустынях.
Горообразовательные процессы обычно сопровождаются вулканической деятельностью: выбросом в атмосферу радиоактивных элементов и других мутагенных факторов химической природы, резкими температурными перепадами, низкочастотными механическими и звуковыми колебаниями, вызывающими состояние стресса. Частота разного рода мутаций у организмов, обитающих в данных условиях, при этом резко возрастает. Видимо, именно поэтому перечисленные Восточно-Африканские озера характеризуются уникальной ихтиофауной: такого огромного видового и родового многообразия эндемичных рыб из семейства Цихлид не встречается более нигде за пределами данной области. Возможно, что повышенный мутагенный фон наряду с изменениями климатических условий, ставших малоблагоприятными для обитания большинства приматов, поставили их в это время на грань вымирания, но жесткий естественный отбор на фоне высокого уровня организации наиболее продвинутых форм позволил им выжить, размножиться и вновь расселиться.
15.5. ОСОБЕННОСТИ ПОВЕДЕНИЯ И ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКООБРАЗНЫХ ПРИМАТОВ КАК ПРЕДПОСЫЛКА К АНТРОПОГЕНЕЗУ
Человекообразные обезьяны - горилла, орангутан и особенно шимпанзе наиболее близки к человеку и характеризуются очень сложным поведением, способностью к обучению и даже самосознанием: они узнают свое отражение в зеркале, способны обучаться и использовать язык глухонемых при общении с экспериментатором и друг с другом, передавать друг другу ложную информацию в корыстных целях, проявлять внешнее дружелюбие для того, чтобы на время замаскировать агрессивные намерения. Человекообразные обезьяны способны не только использовать простейшие орудия труда, но даже и изготавливать их: они очищают от листьев и коры веточки для извлечения муравьев и термитов из их жилищ; передвигаясь по стволам, покрытым острыми шипами, собирают пучки мягких листьев и используют их в качестве грубых варежек.
В сообществах крупных человекообразных обезьян, часто даже в пределах одной популяции, навыки примитивной трудовой деятельности как культурные традиции передаются из поколения в поколение за счет взаимного обучения. Поэтому способы использования и изготовления орудий труда одного и того же назначения в разных группах обезьян, не контактирующих друг с другом, часто оказываются различными: различны методы раскалывания орехов, ловли насекомых, добывания меда диких пчел и т.д. Это свидетельствует о негенетических механизмах передачи соответствующих поведенческих особенностей приматов в отличие от других групп млекопитающих, генетически наследующих большинство видовых форм поведения.
Еще одна особенность, делающая поведение человекообразных обезьян близким к человеческому, проявляется в том, что они способны к выражению дружеских чувств, что особенно часто наблюдается между взрослыми самцами внутри естественной группировки. Одновременно для них характерна и необыкновенная жестокость по отношению к представителям других групп своего вида. Тактика охраны своей кормовой территории от соседей у них проявляется в осторожности наблюдения за противником и в использовании фактора неожиданности при нападении и атаке лишь в том случае, когда они уверены в численном превосходстве и победе.
Сопоставление звуковой коммуникации современных высших приматов и человеческой речи приводит к выводу о том, что у обезьян нет морфофизиологических предпосылок к ее возникновению по многим причинам: высокое расположение гортани, отсутствие подбородочного выступа, отсутствие в височной и теменных долях центров речи.
Но главная причина отсутствия речи у наших далеких родственников в том, что в ней нет и не было необходимости. Действительно, для эффективного выживания в природной среде обезьянам нет необходимости в речевой коммуникации; все, что они хотят сообщить друг другу, может быть сообщено и сообщается без помощи членораздельной речи. Поэтому речь в отряде приматов появилась только тогда, когда реализовалось иное - социальное направление эволюции жизни на Земле, связанное с антропогенезом.
15.6. ПРОИСХОЖДЕНИЕ СЕМЕЙСТВА ГОМИНИД
Появление семейства Гоминид (Hominidae) на Земле связано в первую очередь с переходом к бипедальной локомоции, то есть к передвижению на задних конечностях. Преимущества прямохождения перед движением на четырех конечностях могут быть различны. Так, экспериментально показано, что двуногая походка человека со средней скоростью энергетически выгоднее четвероногой походки других млекопитающих. Кроме того, известно, что наиболее быстро бегающие четвероногие млекопитающие обладают низкой выносливостью к долгому бегу. Так, гепард, развивающий скорость бега более 100 км/ч, выдерживает эту скорость не более нескольких минут. Такая скорость бега недостижима для человека, однако он обладает недостижимой для гепарда выносливостью к длительному бегу (например, на марафонскую дистанцию).
Вертикальное положение тела организмов небольших или средних размеров с короткой шеей, обитающих в открытых ландшафтах, сопряжено с необходимостью издалека заметить хищника и вовремя отреагировать на его появление. Оно также облегчает поиск пищи, воды и ориентировку на местности. Поэтому наземные виды кенгуру, тушканчики и ряд насекомоядных млекопитающих перемещаются по земле исключительно на задних конечностях, а многие степные грызуны - суслики, песчанки, сурки и мелкие хищники постоянно принимают вертикальную позу (рис. 15.7, а). Среди современных низших приматов перемещение на задних конечностях прыжками хорошо известно у ряда видов лемуров Мадагаскара. Прямохождение встречается даже среди
современных пресмыкающихся: австралийская плащеносная ящерица передвигается по деревьям на четырех конечностях, а по земле бегает на задних ногах. Бипедия многократно параллельно и конвергентно возникала в процессе эволюции в разных классах Позвоночных, а весь класс Птицы представлен исключительно двуногими формами. В мезозойской эре огромное число видов пресмыкающихся, господствовавших на Земле, были прямоходящими (рис. 15.7, б).
Прямохождение гоминид, обитавших на открытой местности, на фоне вышеперечисленных преимуществ, могло быть связано также с необходимостью решения и других экологических задач - перенос беспомощных детенышей и продуктов питания на большие расстояния при миграциях, существенное снижение вероятности перегрева тела по сравнению с перемещением на четырех ногах; когда большая поверхность тела подвергается действию солнечного излучения, использование

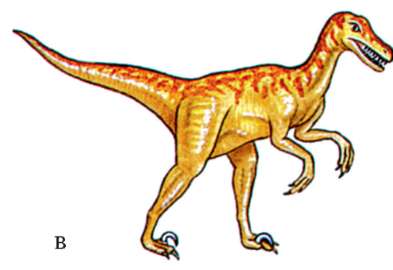
Рис. 15.7. Вертикальная поза и прямохож-дение в разных систематических группах позвоночных: а - мелкий африканский хищник Suricata suricata; б - динозавр Af-rovenator, живший в Африке 130 млн лет назад; в - динозавр Velociraptor
передних конечностей для сигнальной коммуникации между членами группы.
Существует и еще одна, более экзотическая версия объяснения феномена прямохождения, заключающаяся в допущении того, что пред-ковые формы человека обитали в саваннах на берегах крупных рек и озер, добывая пищу в водной среде: рыбу, земноводных, моллюсков и ракообразных, перемещаясь в основном не вплавь, а в вертикальном положении по дну водоемов, собирая со дна и вылавливая руками объекты охоты на поверхности и в толще воды. Этой версией одновременно можно объяснить, почему среди всех приматов только у человека редуцирован волосяной покров на теле, наиболее выражена подкожная жировая клетчатка, отложение большого количества жировой ткани в области молочных желез женщин, наибольшее количество сальных желез на единицу площади кожи, а в состав их секрета входит ненасыщенный углеводород сквален из группы ациклических тритерпенов, обнаруживающийся почти исключительно в секрете сальных желез водных и околоводных млекопитающих. Кроме того, человек - единственный примат, способный произвольно задерживать дыхание, что позволяет ему не только плавать, но и нырять.
Все же любые попытки объяснить вертикальную осанку гоминид исходят из изначально древесного образа жизни их предков и перемещения за счет брахиации, которая и представляет собой прямохождение, но не на задних, а на передних конечностях в подвешенном состоянии, а также за счет вертикального лазания по стволам и толстым ветвям деревьев.
Для человекообразных обезьян, в отличие от низших, более характерно питание плодами и другими съедобными частями растений на деревьях при срывании их передними конечностями с верхних веток, стоя в вертикальном положении на нижних ветках или на земле. В то же время мартышкообразные приматы более адаптированы собирать продукты питания под ветками, на которых они находятся, и поэтому они редко принимают вертикальное положение.
Из этого следует, что у предков гоминид, еще живших на деревьях, мог возникнуть комплекс адаптаций к вертикальной осанке, который был успешно использован и усовершенствован ими при переходе к наземному образу жизни.
Однако, каким бы образом ни возникала двуногая осанка среди пред-ковых форм человека, важно то, что руки предков человека освободились от локомоции и стали использоваться как органы труда. Сложные
движения кистью и пальцами требуют сложной регуляции со стороны центральной нервной системы. Поэтому прогрессивное развитие передней конечности как органа манипуляции предметами сопровождается скоординированными структурно-функциональными эволюционными преобразованиями двигательных и ассоциативных зон коры больших полушарий головного мозга. Морфофункциональные предпосылки этих координаций у приматов существуют с глубокой древности и также связаны с их первично древесным образом жизни).
15.7. ЗНАЧЕНИЕ ИЗМЕНЕНИЙ ГЕНОМА
В ПРОИСХОЖДЕНИИ И ДАЛЬНЕЙШЕЙ
ЭВОЛЮЦИИ ЧЕЛОВЕКА
Существенная часть генетических различий человека и высших приматов не имеет большого значения. Так, гены, отвечающие за структуру гемоглобина у человека и шимпанзе, имеют некоторые отличия, но они никак не сказываются на свойствах самого белка гемоглобина. Ряд известных особенностей генома человека трактуют как изменения, влияющие на рост отдельных частей головного мозга. Существенным своеобразием характеризуется регуляторная область гена, кодирующего белок продинорфин, предшественник нейропептидов - гормонов, связанных с регуляцией эмоционального состояния и поведения, формирования социальных связей, способностей к обучению и запоминанию. Интересно, что кодирующая часть гена продинорфина у человека осталась неизменной, поэтому как сам продинорфин, так и эндорфин, образующийся из него, у шимпанзе и человека идентичны. Изменения же регуляторной части гена приводят к изменению количества синтезируемого продукта в разных ситуациях в зависимости от тех или иных стимулов. Изменения регуляторного участка продинорфинового гена происходили не только в процессе антропогенеза, но и позже, при интенсивном расселении человека из Африки на другие континенты и в разные природно-климатические зоны. Этим, в частности, можно объяснить формирование отличающихся друг от друга моделей поведения и культурных традиций народов, принадлежащих к разным этническим группам и подвергающихся действию разнообразных экологических факторов. Однако те несколько точечных мутаций регуляторного участка гена продинорфина, которые отличают Homo sapiens от его предков-обезьян, сохраняются у всех представителей современного человека в неизменном виде.
Сопоставление последовательностей нуклеотидов в геномах человека и шимпанзе показало, что отличия касаются около 35 млн пар нукле-отидов, что составляет около 1% генома. Однако у человека выявлено 6 регионов, не встречающихся у шимпанзе. Так, ген, отвечающий за синтез малой регуляторной РНК, транскрибирующийся во время закладки коры полушарий мозга человека, имеет 18 отличий от соответствующего гена шимпанзе. Ген FOX p2, встречающийся у певчих птиц и многих млекопитающих, обладает определенной спецификой строения у человека. Его мутантные формы у человека приводят к нарушению речи.
Характерна для человека также инактивация гена MYH 16, активного у других приматов. Этот ген у млекопитающих принимает участие в регуляции развития жевательной мускулатуры. Вероятно, он в значительной степени утратил свою роль при переходе предков человека к употреблению менее грубой растительной и животной пищи. Ослабление жевательной функции в процессе антропогенеза способствовало интенсификации моторики более мелких мышц, обеспечивающих более тонкие движения губ, языка, мягкого нёба и глоточной области, принимающих участие в артикуляции речи.
Но не все изменения молекул ДНК имеют одинаковое значение в прогрессивной эволюции человека. С помощью современной вычислительной техники удалось выявить 49 областей генома, которые в процессе антропогенеза изменялись примерно в 70 раз быстрее других. Они были названы HAR-областями (human accelerated regions - области ускоренного развития человека). В наиболее сильно отличающейся области изменения затронули 18 пар нуклеотидов из 118. Изучение этой части генома позволило установить, что она содержит 2 гена - HAR1F и HAR1R). Точно определить значение этих генов трудно, но имеются основания считать, что они связаны с регуляцией развития коры головного мозга. То, что этот участок генома существует только у человека, свидетельствует о его большой значимости как в происхождении человека, так и в его индивидуальном развитии, обеспечивая формирование чисто человеческих особенностей организации мозга.
В эмбриогенезе человека экспрессия этих нуклеотидных последовательностей начинается на 7-й неделе развития зародыша, а заканчивается на 12-й неделе. Именно в этот период морфогенез головного мозга происходит наиболее интенсивно.
В целом фенотипические характеристики человека и обезьян отличаются друг то друга гораздо сильнее, чем генотипические. Это может быть объяснено, в частности, различным распределением белков в клет-
ках зародыша или плода в процессе развития в разные его периоды и в разных тканях и органах, что в свою очередь определяется спецификой активности регуляторных генов. Дальнейшее изучение геномов человека и его ближайших родственников должно привести к более глубокому пониманию механизмов антропогенеза.
Изучение ДНК современных людей дает возможность не только определить родство представителей разных популяций, но и проследить их историческую судьбу.
Несмотря на то что основные принципы организации генома каждого вида, включая человека, являются общими, у разных индивидов отдельные участки его могут весьма сильно отличаться.
По этим участкам можно проводить идентификацию человека, определять степень родства отдельных людей и популяций. В группах людей, общих по происхождению, сходство таких варьирующих участков ДНК велико по сравнению с неродственными группами. Таким образом, сопоставляя степень различий последовательностей нуклеотидов ДНК людей, населяющих разные географические зоны, можно определить степень их родства, а исходя из данных о скорости накопления генных мутаций, - и время, прошедшее с момента разделения популяций в процессе их перемещения из области их происхождения.
Наиболее удобный объект для таких исследований - молекулы ми-тохондриальной ДНК. Во-первых, в каждой клетке она встречается в количестве нескольких сот копий и лучше, чем ядерная ДНК, сохраняется в биологических образцах. Во-вторых, она не подвергается рекомбинации, так как передается только по материнской линии. На основе анализа митохондриальной ДНК современных людей разного этнического и географического происхождения было построено филогенетическое древо человечества. Оказалось, что все варианты митохондриальной ДНК человека произошли от митохондриальной ДНК одной женщины, жившей в Восточной Африке.
У-хромосома обладает свойствами, сходными с митохондриальной ДНК - она также не подвергается рекомбинации, а передается только по мужской линии. Исследование истории человечества по ДНК У-хромосомы также привело к выводу об африканском происхождении современного человека. Время появления Homo sapiens датируется обоими методами в интервале 135-185 тыс. лет назад.
Методом анализа митохондриальной ДНК было получено подтверждение выводов о том, что неандертальцы не были предками Homo sapiens, а одновременно сосуществовали с ним на протяжении не-
скольких десятков тысяч лет и вымерли. Однако, долгое сосуществование неандертальцев на одной территории с представителями близкого вида - современного человека - не исключало возможности их отдельных скрещиваний. Это было доказано работами немецких антропологов 2010 г. Выяснилось, что в генетическом материале 1-4% современных европейцев и жителей Ближнего Востока существует некоторое количество нуклеотидных последовательностей, которые могут представлять собой фрагменты ДНК неандертальцев. Характерно, что в африканских человеческих популяциях людей такие нуклеотидные последовательности не обнаруживаются.
15.8. ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ ГОМИНИД
И ПРОИСХОЖДЕНИЕ ЧЕЛОВЕКА
Если на протяжении почти всего ХХ в. древнейшими гоминидами считались представители рода австралопитеков (Australopithecus), то на рубеже третьего тысячелетия были обнаружены ископаемые остатки многочисленных гоминид более раннего происхождения, но уже с явными признаками прямохождения и с прогрессивным строением черепа, зубов и конечностей
Наиболее древним известным африканским представителем семейства гоминид является сахельантроп чадский - Sahelanthropos tschadensis, (рис. 15-8), обладавший мозаикой примитивных и прогрессивных черт организации, но главные его особенности - явные признаки прямохож-дения. Исторический возраст этого организма - 7-6 млн лет. Археологические раскопки на территории обнаружения останков этого примата показали, что он жил на берегу пресного водоема в засушливой безлесной местности. Зная, что момент дивергенции эволюционной линии, ведущей с одной стороны к современным человекообразным обезьянам, а с другой - к предкам человека, именно около 7 млн лет назад, можно предположить, что общие предки человека и современных высших го-минид внешне были похожи на сахельантропа.
Примерно таким же возрастом датируются находки другого гомини-да - Orrorin tugenensis, обитавшего на территории современной Кении в районе Великого Африканского рифта. Судя по строению скелета, он мог одинаково хорошо передвигаться как на задних конечностях по земле, так и лазать по деревьям. Строение его зубов близко к человеческому.
Во временном интервале 5,8-4,4 млн лет назад в Восточной Африке обитали представители другого рода гоминид - ардипитеки
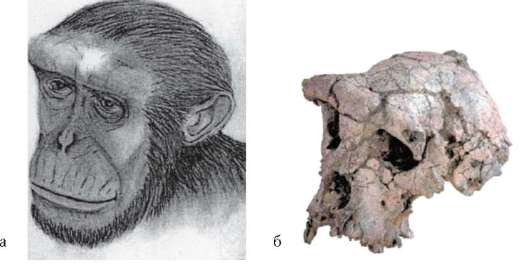
Рис. 15.8. Сахельантроп чадский (а) (Sahelanthropos tschadensis) и его череп (б)
(Ardipithecus). Они жили в условиях более высокой влажности и были адаптированы как к вертикальному лазанию, так и к прямохождению.
В период 4,2-2,5 млн лет назад в области той же Восточной Африки наблюдается расцвет нового рода высших приматов - австралопитеков, которые широко расселяются по африканскому континенту. Строение их скелетов свидетельствует об их бипедальной локомоции. Изученные ископаемые формы австралопитеков относят к четырем или пяти видам, наиболее древний из которых - A. anamensis. В настоящее время предполагают, что этот вид представяет собой переходную форму между прогрессивными ардипитеками и более поздними австралопитеками.
Australopithecus afarensis, другой вид австралопитеков, известен по остаткам более 300 особей. К этому виду, в том числе, относится известная Люси. Он сохранил в себе много признаков обезьян, но главное отличие от них - прямохождение. Объем головного мозга мало отличается от обезьян (рис. 15.9, а, б).
3-2,5 млн лет назад широко расселился по Африке, достигнув ее южной области, A. africanus. По сравнению с предыдущим видом он имеет более округлый череп, больший объем мозга (до 550 см3), более уплощенное лицо, все зубы, более близкие человеческим и расположенные в виде широкой дуги, как у человека, клыки малых размеров.
Есть мнение, что этот вид питался в основном животной пищей, причем скорее всего трупами животных, убитых более крупными специа-

Рис. 15.9. а - Австралопитек афарский (Australopithecus afarensis), реконструкция; б - череп австралопитека афарского; в - парантроп (Paranthropus boisei) реконструкция; г - череп парантропа
лизированными хищниками, такими как львы и леопарды. На многих костях крупных травоядных животных, найденных вместе с останками австралопитеков, обнаруживаются следы их преднамеренного раскалывания. Потребление пищи животного происхождения могло привести к прогрессивному развитию ряда морфологических и физиологических признаков этих существ, а умение извлекать головной и костный мозг путем раскалывания костей убитых животных при помощи камней могло лежать в основе формирования первых трудовых навыков. Большое число черепов животных, мозгом которых они питались, расколото однотипно и с левой стороны, что свидетельствует, с одной стороны, о том, что австралопитеки были, в основном, правшами, а с другой - о том, что первые трудовые навыки могли передаваться социальным путем, то есть в процессе обучения.
2,5 млн лет назад существовал еще один вид австралопитеков - A. garhi. Этот вид в настоящее время многие антропологи считают пред-ковой формой человека умелого - Я. habilis, хотя утверждать это с высокой вероятностью невозможно.
Интересно, что одновременно с разными видами австралопитеков сосуществовали несколько близких им видов так называемых парантропов (Paranthropus - P. robustus, P. boisei), представлявших собой своеобразную ветвь двуногих человекообразных форм, эволюционировавших в сторону усиления звериных черт. У всех представителей этой группы го-минид существовал костный гребень в области шва теменных костей черепа для прикрепления мощных жевательных мышц, коренные зубы их очень массивны, лицо сильно уплощено, а лоб при этом невелик. Объем головного мозга около 500 см3, сильно выражены надбровные дуги. Некоторые формы этой группы, видимо, умели изготавливать простейшие орудия труда. Мощный челюстной аппарат парантропов свидетельствует об использовании ими грубой растительной пищи (рис. 15.9, в, г).
В целом парантропы оказались тупиковой ветвью эволюции и вымерли около 1 млн лет назад. Появление и эволюция практически одновременно нескольких родов и видов гоминид свидетельствуют о том, что и в этой группе организмов на определенных этапах ее существования проявляется адаптивная радиация форм жизни, ведущая к биологическому прогрессу и способствующая эффективному отбору наиболее продвинутых видов.
В 1964 г. по находкам, сделанным в ущелье Олдувай в Танзании, был выделен вид Homo habilis, или человек умелый, имеющий абсолютный возраст 2-1,7 млн лет (рис. 15.10, а, б). Его отличительные черты - дву-
ногость, в целом прогрессивное строение кисти, зубной системы, объем мозговой коробки от 540 до 700 см3, что примерно в полтора раза превышает объем мозга австралопитеков. На внутренней поверхности черепа обнаруживаются признаки прогрессивных нейроморфологических изменений, определяющиеся по отпечаткам головного мозга: выраженная асимметрия полушарий и развитие двух речевых центров как условие для возникновения членораздельной речи. Большой палец стопы не отведен в сторону. Это свидетельствует о том, что морфологические перестройки, связанные с прямохождением, у него полностью завершились. Вместе с останками H. habilis найдены орудия труда со следами целенаправленной обработки, свидетельствующие о ранних формах трудовой деятельности. Эти грубо обработанные, заостренные с одной стороны каменные рубила получили название орудий олдувайской культуры
(рис. 15.10, в).
Перечисленные признаки, ведущим из которых является прогрессивное развитие мозга, характеризуют организм уже с иной морфо-функциональной организацией по сравнению с австралопитеками. Эти признаки считаются специфичными для рода Homo.
Близкие к Homo habilis виды человека существовали одновременно с ним, их останки обнаружены как в Восточной Африке - H. rudolfensis, так и в Западном Закавказье - H. georgicus. Археологический возраст обоих - 1,8 млн лет. Орудия труда этих представителей гоминид были очень похожи на орудия олдувайского типа. Особенно интересен второй вид своим местообитанием - это самая древняя находка гоминид за пределами Африки, свидетельствующая о том, что расселение людей из места их первоначального расселения произошло более чем на миллион лет раньше, чем считалось до этого открытия (рис. 15.11, а). Следующий, более прогрессивный вид ископаемого человека африканского происхождения - Homo ergaster (человек работающий). Возраст его - 1,9-1,6 млн лет. Объем его мозга около 900 см3, кости черепа тонкие, а зубы уменьшены в размерах (рис. 15.11, б). Но самое главное то, что он создавал орудия труда нового типа - обоюдоострые рубила и стал использовать огонь. Кроме того, по сравнению с H. habilis, у H. ergaster увеличились размеры тела, что было связано, вероятно, с возрастанием доли животной пищи: применяя новые орудия труда, он мог охотиться эффективнее, а также более успешно конкурировать с животными, питающимися падалью.
С этим видом ископаемого человека или с близкими к нему формами в настоящее время ряд исследователей связывают происхождение двух
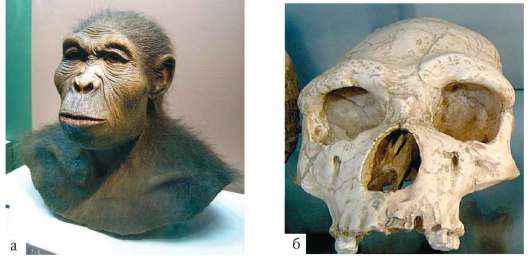

Рис. 15.10. а - человек умелый (Homohabilis) (реконструкция); б - череп человека умелого; в - орудия олдувайской культуры

Рис. 15.11. а - Homogeorgicus (реконструкция); б - череп H. ergaster
эволюционных ветвей, ведущих как к появлению нескольких форм вымерших представителей рода Homo, так и к возникновению современного человека.
Homo erectus, или человек прямоходящий (рис. 15.12), появился примерно 1,6 млн лет назад, существовал очень долгое время и вымер, как было обнаружено недавно, около 50 тыс. лет назад. Последние ископаемые останки этого человека найдены на острове Ява. Широкое использование огня, продвинутая по сравнению с более ранними формами человека материальная культура и ярко выраженная социальность позволили ему быстро и эффективно расселиться почти по всей территории Африки и южной Евразии и освоить обширный ареал, разнообразный в природно-климатическом отношении.
Действительно, орудия труда H. erectus более прогрессивны, чем у H. habilis, а масса мозга (от 800 до 1000 г) явно превышает минимальную массу (750 г), при которой возможно существование речи. Наличие при этом речевых центров, возникших впервые у человека умелого, предполагает и развитие второй сигнальной системы.
Выделяют три группы H. erectus, обитавших в Европе, Азии и Африке. Долгое время древнейшими архантропами считались азиатские представители из Индонезии и Восточного Китая - питекантроп и синантроп. Однако находки последних лет на территории Израиля (1982) и Кении (1984), датирующиеся соответственно 2,0 и 1,6 млн лет, сопровождающиеся элементами материальной культуры и признаками использования огня, показали, что арогенная эволюция гоминид происходила на африканском континенте и на Ближнем Востоке. Это позволило связать происхождение H. erectus с восточно-африканскими предковы-ми формами.
Наличие многочисленных находок этого вида древностью 1,50,1 млн лет в отдаленных от Африки регионах - в Юго-Восточной и Восточной Азии, в Центральной Европе и даже на Британских островах - свидетельствуют об активных адап-
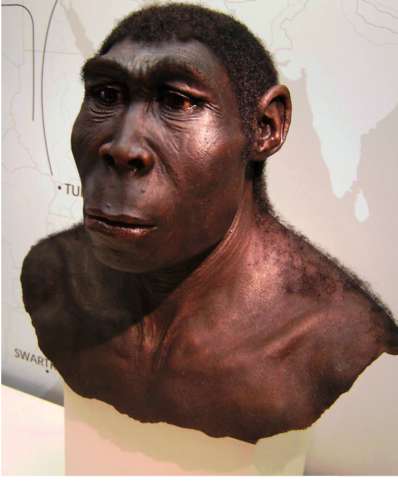
Рис. 15.12. Homo erectus (реконструкция)
тациях их к разнообразным условиям существования. В связи с тем, что небольшое различие ископаемых останков Я. erectus не соответствует значительному разнообразию природно-климатических условий указанных территорий, можно заключить, что в формировании этих адаптации значительную роль играли наряду с факторами биологической эволюции также и социальные факторы (рис. 15.13).
В Индонезии на острове Флорес (малые Зондские острова) недавно были обнаружены костные останки ранее неизвестного карликового вида людей, живших еще 38-18 тыс. лет назад. Предполагается, что этот вид (Homo floresiensis) - боковая ветвь изолированных островных популяций Я. erectus. Обнаружение на этом же острове каменных орудий Я. erectus возрастом 850 тыс. лет свидетельствует о том, что он освоил этот остров достаточно давно. В условиях долгой островной изоляции потомки Я. erectus, вероятно, уменьшились в размерах и так видоизменились, что их пришлось описать как отдельный вид (рис. 15.14). Рост их был менее метра, а объем мозга - 380 см3, как у современных
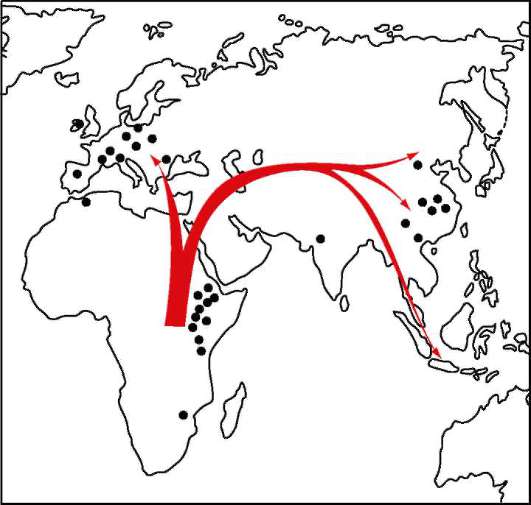
Рис. 15.13. Места обнаружения ископаемых останков человека прямоходящего и пути его миграции
шимпанзе. Тело не было покрыто волосяным покровом, челюсти сильно выступали вперед. Они владели огнем, изготавливали совершенные каменные орудия и охотились на крупных животных. Вымирание их связывается с заселением острова более прогрессивными формами человека. Вероятно, заселение островов Зондского архипелага именно современным человеком привело к быстрому исчезновению последних популяций H. floresiensis. Существование в прошлом такой экзотической формы человека свидетельствует о широких адаптивных возможностях рода Homo и делает возможным находки других популяций и видов архаичного человека, сильно уклоняющихся от известных вымерших видов и современных популяций человека.
Наиболее прогрессивные популяции Я. erectus африканского происхождения мигрировали на север, в Европу и, по-видимому, эволюционировали в новый вид - Homo antecessor, что в переводе означает предок (рис. 15.15). Лицевая часть его черепа близка по строению к черепу современного человека. Представители этого вида охотились на крупных животных. Вероятно, среди этой группы людей был распространен и каннибализм. Объем их мозга составлял около 1000 см3. Возраст этого вида - 780-800 тыс. лет. Многие антропологи считают, что этот вид мог быть общим предком как неандертальского, так и современного человека.
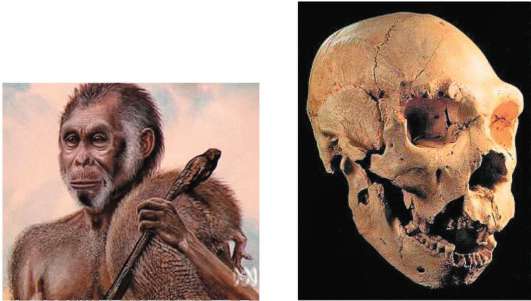
Рис. 15.14. Homo floresiensis
Рис. 15.15. Череп Homo antecessor
Более совершенная форма человека описана по материалам, обнаруженным в Европе, в качестве вида H. heidelbergensis, жившего 800200 тыс. лет назад. Интересно, что этот вид изготавливал и использовал новый тип орудий - метательные копья с заостренным концом и с центром тяжести, расположенным так же, как у современных изделий этого же предназначения.
Считается, что гейдельбергский человек, связанный по происхождению с Я. antecessor, дал начало той ветви эволюции человека, которая ведет к возникновению неандертальцев. Неандертальцы (Яото neanderthalensis) широко расселялись по Европе и Западной Азии на протяжении длительного периода от 200 до 28 тыс. лет назад (рис. 15.16), но за пределы этой зоны не вышли. Археологические находки последних

Рис. 15.16. а - гейдельбергский человек (реконструкция); б - неандерталец (реконструкция); в - приблизительный ареал расселения Я. neanderthalensis
лет дают повод судить о недооценке интеллектуальных возможностей неандертальцев. На всех стоянках обнаружены следы костров и обгоревшие кости животных, что свидетельствует об использовании огня для приготовления пищи. Орудия труда их гораздо совершеннее, чем у более ранних форм и часто не отличаются от орудий труда, которые изготавливались ранними формами людей современного физического типа (рис. 15.17). Масса мозга неандертальцев около 1500 г, причем особое развитие получили отделы, связанные с логическим мышлением. Имеются данные о ритуальных захоронениях неандертальцев на территории Ближнего Востока. Костные останки неандертальца из Сен-Сезер (Франция) были найдены вместе с орудиями труда, свойственными верхнепалеолитическому человеку, что свидетельствует об отсутствии резкой интеллектуальной грани между неандертальцем и современным человеком. Размеры и форма ротовой полости, строение зубов, челюстей и положение подъязычной кости у неандертальцев почти ничем не отличаются от соответствующих органов артикуляции современного
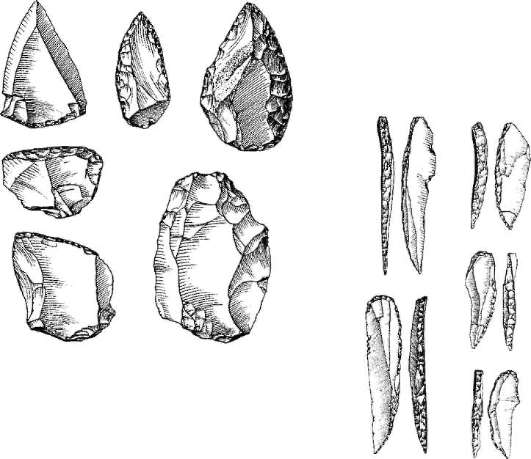
Рис. 15.17. Орудия труда, которые изготавливали как неандертальцы, так и ранние формы современных людей
человека. Объем и соотношение долей мозга их также отличаются незначительно. Из этого следует, что неандертальцы были способны к развитию речи, но неизвестно, использовали ли они эту возможность.
Многочисленные находки неандертальцев и современного человека, Homo sapiens, на территории Европы, датирующиеся 37-25 тыс. лет, свидетельствуют о сосуществовании обоих видов в течение многих тысячелетий.
В тот же период представители Homo sapiens обитали уже не только в Европе и Африке, но и в отдаленных районах Азии (острове Тайвань, острове Окинава) и даже начали заселять Америку. Эти данные указывают на необычайно быстрый процесс расселения современного человека, что может быть доказательством «взрывного», скачкообразного характера антропогенеза в этот период как в биологическом, так и в социальном смысле. А Homo neanderthalensis в виде ископаемых остатков не обнаруживается позже рубежа в 28 тыс. лет.
С возникновением человека современного физического типа роль биологических факторов в его эволюции свелась к минимуму, уступив место социальной эволюции. Об этом отчетливо свидетельствует отсутствие существенных морфологических различий между ископаемым человеком, жившим 30-25 тыс. лет назад, и нашим современником.
Существует несколько версий, объясняющих вымирание неандертальцев. Одна из них исходит из того, что по сравнению с современным человеком тело неандертальцев более массивно, конечности существенно короче, а трубчатые кости значительно толще. Такая конституция требует больших затрат энергии при передвижении и поэтому менее выгодна по сравнению с более грацильными пропорциями тела человека современного физического типа. Однако именно такое телосложение является выгодной адаптацией к обитанию в условиях холодного климата Европы по сравнению с африканской прародиной человека. Поэтому эта версия кажется сомнительной.
Другая версия основана на экосистемных причинах: современный человек, мигрировав из Африки на европейскую территорию, давно освоенную неандертальцами, мог принести с собой африканские виды возбудителей и переносчиков инфекционных и паразитарных заболеваний, с которыми европейские популяции человека ранее никогда не встречались. Это могло приводить к вспышкам катастрофических эпидемий болезней среди неандертальцев, не имевших к ним иммунитета. Эта версия кажется достаточно правдоподобной, так как в истории человечества известны примеры подобного рода. Так, в эпоху Великих
географических открытий контакты европейцев с племенами людей, обитавших на изолированных островах Океании и на территории Южной и Северной Америки, нередко приводили к массовой гибели местного населения. Так, на острове Тасмания уже в XIX в. не осталось ни одного местного жителя, несмотря на то, что европейские иммигранты не ставили перед собой задачи полного уничтожения тасманийцев: все аборигенные жители острова вымерли от занесенных европейцами инфекционных заболеваний, с которыми ни они сами, ни их предки ранее никогда не встречались.
Не исключено также, что на определенном этапе социального развития современного человека в Европе технология изготовления и качество производимых им орудий труда стали существенно прогрессивнее неандертальских. В результате их трудовая деятельность стала более эффективной, и неандертальцы постепенно уступили место более успешным представителям рода.
Какую-то роль в исчезновении неандертальцев могло сыграть и их физическое уничтожение современным человеком в процессе конкуренции за жизненные ресурсы, но достоверных палеонтологических данных об их столкновениях и массовой гибели пока не имеется.
Несмотря на весьма существенные различия неандертальцев и человека современного физического типа, оба вида даже на ранних этапах их эволюции, несомненно, характеризовались выраженной социальностью, степень развития которой со временем нарастала. И не исключено, что если бы современный человек не возник в свое время, а неандертальцы продолжали бы эволюционировать как социальный вид, то современная цивилизация могла бы быть создана ими. Вполне возможно, что она выглядела бы несколько иначе.
Археологические, а также молекулярно-генетические данные, основанные на изучении нуклеотидных последовательностей мито-хондриальных ДНК и ДНК Y-хромосом современных людей разного географического и этнического происхождения привели к выводу о том, что древняя прародина человека современного физического типа - та же зона Северо-Восточной Африки, где и произошло возникновение первых представителей рода Hото. Время дивергенции эволюционных ветвей, ведущих к неандертальцам и современному человеку, по этим данным, соответствует примерно 500 млн лет. Различия их нуклеотид-ных последовательностей укладываются в рамки межвидовых, и поэтому гибридизация двух сосуществовавших во времени и пространстве форм человека едва ли могла быть успешной. Поэтому не подтвержда-
ется и мнение о том, что неандертальский человек мог быть ассимилирован современным за счет скрещивания.
Разнообразие митохондриальных ДНК современного человека очень мало и это может быть объяснено тем, что все существующее в настоящее время человечество имеет общее происхождение от небольшой предковой популяции размерами не более 10 тыс. особей, находящихся в более или менее близком родстве друг с другом по материнской линии (рис. 15.18).
Наиболее архаичные формы Homo sapiens в соответствии с этими данными появились 160-130 тыс. лет тому назад. Основные пути миграции ранних форм Н. sapiens и их расселение по Земле показаны на рис. 15.19.
На рис. 15.20 показано время существования известных гоминид и предположительные эволюционные связи между ними.
15.9. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВА
С момента возникновения Н. sapiens социальное в человеке стало его сущностью и биологическая эволюция видоизменялась, проявляясь в возникновении широкого генетического полиморфизма (см. п. 12.3).
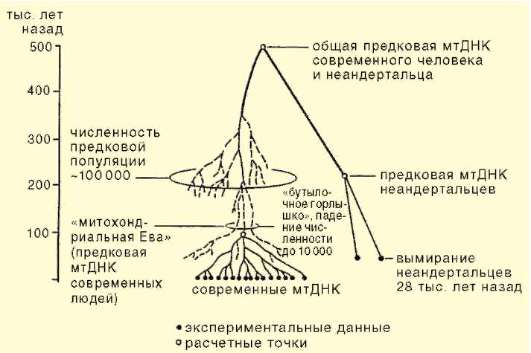
Рис. 15.18. Схема эволюции линий митохондриальной ДНК современного человека и неандертальцев
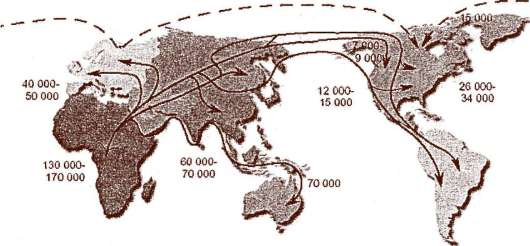
Рис. 15.19. Основные пути и время миграции человека современного физического типа по территории Земли. Цифрами указано время миграции в тысячах лет тому назад, установленное по генетическим и археологическим данным
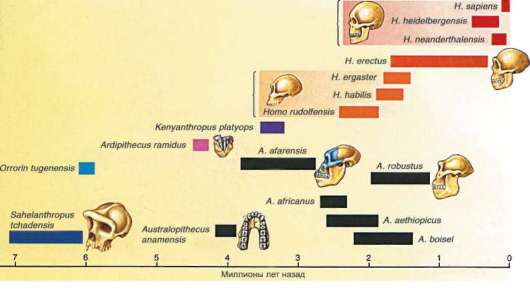
Рис. 15.20. Время существования ископаемых гоминид и их предполагаемые генетические связи с современным человеком
Генетическое разнообразие на уровне генов и в меньшей степени хромосом обеспечивает разнообразие генотипов особей. Разнообразные генотипы по-разному проявляются в меняющихся условиях среды, давая огромное фенотипическое многообразие людей.
В основе морфофизиологического полиморфизма человечества лежат полиморфизм наследственного материала на уровне генома и мо-дификационная изменчивость. Эти факторы обеспечивают не только индивидуальное морфофизиологическое многообразие, но и внутривидовую групповую дифференциацию человечества на расы и адаптивные экологические типы.
15.9.1. РАСЫ И РАСОГЕНЕЗ
На протяжении длительного времени в антропологии господствовали представления о значимости расовой дифференцировки человечества и о большой роли естественного отбора в формировании основных расовых признаков. Применение методов молекулярной антропологии в значительной степени изменило представление о расах и расогенезе.
Человеческие расы - это большие популяции людей, характеризующиеся устойчивыми комплексами морфофизиологических признаков преимущественно адаптивной природы.
Морфологические и в меньшей степени физиологические признаки дают возможность выделить внутри человечества три основные большие расы: европеидную, австрало-негроидную и монголоидную.
Европеоиды имеют светлую или смуглую кожу, прямые или волнистые волосы, узкий выступающий нос, тонкие губы и развитый волосяной покров на лице и теле. У монголоидов кожа также может быть как светлой, так и темной, волосы обычно прямые, жесткие, темно пигментированные, косой разрез глаз и эпикант («третье веко»). Негроиды характеризуются темной кожей, курчавыми или волнистыми волосами, толстыми губами и широким, слегка выступающим носом. Имеются отличия рас и по некоторым физиологическим и биохимическим показателям: интенсивность потоотделения с единицы площади кожи у негроидов выше, чем у европеоидов, средние показатели уровня холестерина в плазме крови наиболее велики у европеоидов.
В рамках каждой большой расы выделяются отдельные антропологические типы с устойчивыми комплексами признаков, называющиеся малыми расами. Существует три основных подхода к классификации рас: без учета их происхождения, с учетом происхождения и родства и
на основе популяционной концепции. В соответствии с первым подходом три большие расы включают в себя 22 малые, причем между большими расами располагаются по две переходные малые. Схема расовой классификации изображается при этом в виде круга (рис. 15.21). Несмотря на то что при такой классификации не учитывается происхождение рас, само существование малых переходных рас, сочетающих в себе одновременно признаки двух больших рас (эфиопская, южносибирская, уральская и т.д.), свидетельствует, с одной стороны, о динамизме расовых комплексов признаков, а с другой - об условности членения человечества даже на большие расы.
Гибридизация ДНК между большими выборками представителей малых рас в рамках одной большой показала высокую степень гомологии
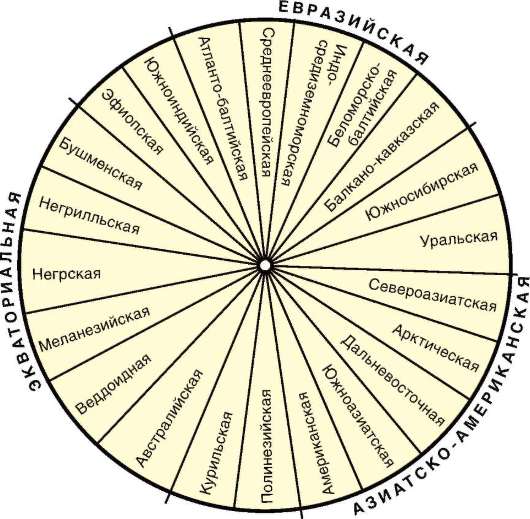
Рис. 15.21. Расовая классификация человечества без учета происхождения рас
нуклеотидных последовательностей. Гибридизация ДНК представителей пар разных больших рас выявляет их относительную отдаленность друг от друга. Изучение гомологии нуклеотидных последовательностей западных европеоидов и представителей малой уральской расы и центрально-азиатских монголоидов с той же самой уральской расой дает среднее значение. Эти данные свидетельствуют о том, что переходные малые расы совмещают в себе не только морфологические признаки в соответствии с их промежуточным положением, но оказываются промежуточными и в отношении генетическом. Из этого следует, что они либо гибридогенны, либо сохранили в своей организации более древние черты, характерные для этапа существования человечества, предшествующего формированию больших рас.
Классификация с учетом происхождения рас изображается в виде эволюционного древа с коротким общим стволом и расходящимися от него ветвями (рис. 15.22). В основе таких классификаций лежит обнаружение черт архаизма и эволюционной продвинутости отдельных рас, в соответствии с чем разные большие и малые расы занимают разное положение на ветвях такого древа. Выявление архаичных и прогрессивных черт среди морфологических признаков носит субъективный характер, благодаря чему схемы расовых классификаций такого рода очень многообразны. Но самый большой недостаток подхода к классификации рас такого рода -
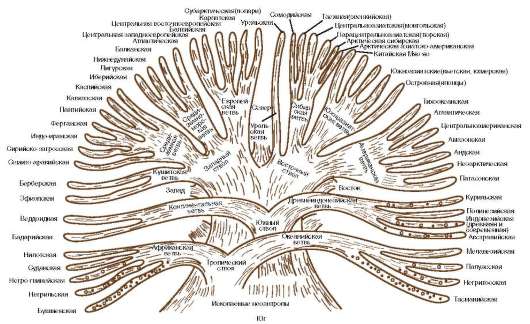
Рис. 15.22. Расовая классификация человечества с учетом происхождения
попытка расположить расы на разных уровнях эволюционного древа, т.е. признание их биологической неравноценности. На рис. 15.23 изображена другая схема расовой классификации, учитывающая возможность генетического смешения ветвей различных расовых типов в прошлом. Она чрезвычайно сложна и построена на основе антропометрических и других внешних признаков без учета генетических характеристик. Разветвления, перекрещивания и слияния различных ветвей в схеме не везде подтверждаются данными истории и археологии.
Кроме того, данные палеоантропологических исследований показывают, что вплоть до верхнего палеолита на территориях, обитаемых людьми, практически нигде не сформировались расовые типы человека, с которыми были бы генетически связаны современные большие расы. Это подтверждает анализ верхнепалеолитических находок скелетов людей современного физического типа из сунгирских погребений (Россия, Владимирская обл.), живших приблизительно 26 тыс. лет назад. Все черепа, принадлежащие им, характеризуются мозаичным сочетанием расовых признаков и не могут быть отнесены ни к одной из современных больших рас. Этим данным соответствует и описание ископаемого скелета из Южной Калифорнии, пролежавшего в земле 21,5 тыс. лет и характеризующегося отсутствием выраженных монголоидных черт, несмотря на то что аборигенным населением Америки являются монголоиды.
Только более поздние мезолитические находки свидетельствуют о формировании у человека расовых признаков. Так, известны более поздние мезолитические черепа с территории Северной Африки возрастом 10-8 тыс. лет с явными признаками не просто негроидной, а малой эфиопской расы. Сходные данные получены на территории Европы и в других регионах. Все это указывает на то, что процесс формирования расовых признаков - довольно поздний, идущий параллельно в разных регионах на рубеже верхнего палеолита-мезолита на фоне исходной разнородности морфофизиологических признаков у человека современного физического типа.
Первичное появление на протяжении эволюции признаков малых, а не больших рас позволяет сделать вывод о том, что европеоидная, монголоидная и негроидная расы имеют мозаичное происхождение и представляют собой крупные популяции, объединенные не столько общностью происхождения, сколько климато-географическими характеристиками условий существования и адаптивностью большинства основных признаков.
Это заключение хорошо согласуется с популяционной концепцией рас. Суть ее заключается в следующем. Если принять, что большие расы
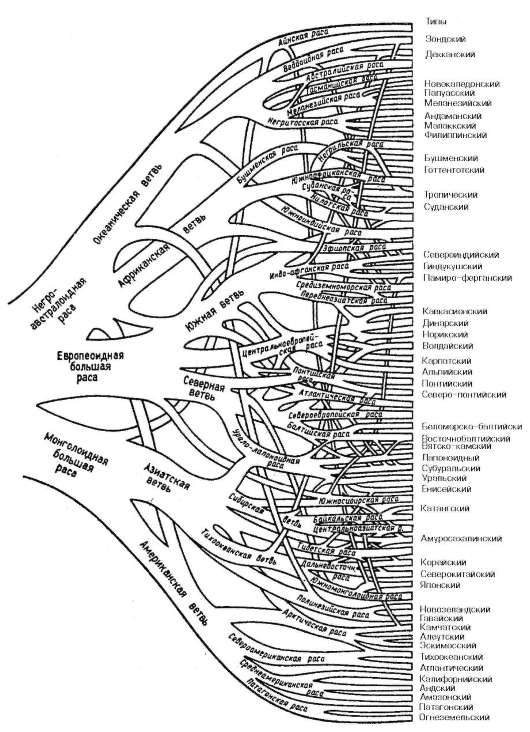
Рис. 15.23. Расовая классификация человечества. Переплетение и слияние ветвей отражают процессы смешения в различные исторические периоды
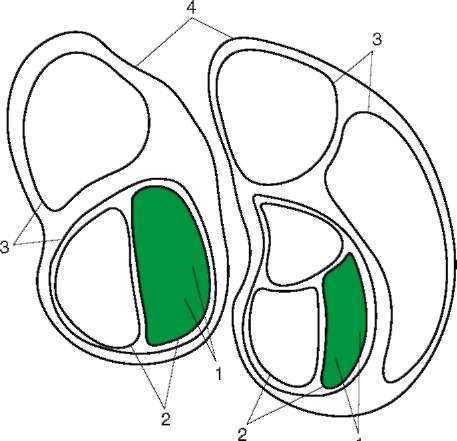
человека представляют собой огромные популяции, то малые расы - субпопуляции больших, локальные естественные общности людей внутри которых - конкретные этнические образования (нации, народности) - являются меньшими популяциями. Если предположить при этом, намеренно упрощая ситуацию, что этносы не разделяются на элементарные популяции, и считать их просто состоящими из конкретных особей, то получится сложная структура, включающая в себя четыре уровня иерархии: большие расы, малые расы, этносы, индивидуумы (рис. 15.24).
Рис. 15.24. Расы как выражение генетического полиморфизма человечества: 1 - отдельные индивидуумы; 2 - этносы; 3 - малые расы; 4 - большие расы
На основании исследований распределения различных групп крови и белков в популяциях человека произведено сравнение доли каждого из четырех уровней меж- и внутрипопуляционных различий в общем объеме генетического полиморфизма человека по этим признакам (табл. 15.1):
Таким образом, от тотального генетического полиморфизма человечества расовые признаки составляют только 8%, в то время как основная доля генетического разнообразия определяется многообразием отдель-
Таблица 15.1. Доля генетического разнообразия в зависимости от уровня по-пуляционных различий
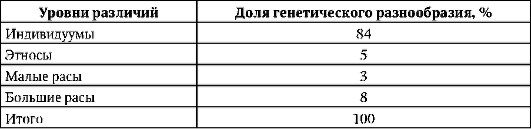
ных индивидуумов. Иными словами, немец может быть генетически гораздо ближе к полинезийцу, чем к другому немцу, живущему в соседней квартире. Изучение геногеографии популяций человека показало, что географическое распределение частот аллелей генов групп крови системы AB0, MN, Lutheran, Duffy, Diego и др., а также различных форм ферментов и иммуноглобулинов не соответствует ареалам расселения ни одной из рас. Так, по группам крови АВ0 и MN жители Европы оказываются ближе к африканцам, в то время как по системе иммуноглобулинов они ближе к монголоидам Азии. Сходные результаты получены и в отношении распределения в популяциях вариантов митохондриальной ДНК.
Эти данные свидетельствуют о том, что биохимический полиморфизм человека эволюционно возник раньше и развивался дольше по сравнению с возникновением комплексов расовых признаков. Из этого следует, что расы не представляют собой особых изолированных групп людей, характеризующихся наборами специфических генов. Расовые же характеристики - не более чем отдельные проявления общего генетического полиморфизма, выражающегося в первую очередь в сложных морфологических признаках. Некоторые из них адаптивны, другие сформировались на основе коррелятивной изменчивости, но все они касаются лишь ряда второстепенных особенностей (цвета кожи, волос, глаз и т.д.) и не затрагивают таких общечеловеческих признаков, как морфология головного мозга, а также строение и функции руки как органа труда.
На основании определения числа аллелей, свойственных той или иной группе организмов, возможно определение генетического расстояния между ними. Эта величина для больших рас человека составляет 0,03. Она гораздо ниже цифр, характерных для истинных подвидов (0,17-0,22), и еще незначительнее по сравнению с межвидовым расстоянием (0,5-0,6 и более). В животном мире генетическое расстояние, равное 0,03, соответствует обычно генетическим отличиям местных популяций друг от друга. Все эти данные свидетельствуют о том, что понятие расы условно, второстепенно и не позволяет подводить под иерархическую классификацию рас глубокую биологическую, а значит, и социальную базу.
15.9.2. АДАПТИВНЫЕ ЭКОЛОГИЧЕСКИЕ ТИПЫ ЧЕЛОВЕКА
Человечество, заселившее уже около 15 тыс. лет назад все более или менее благоприятные для жизни природно-географические зоны, встретилось с необходимостью адаптироваться к самым разнообразным условиям существования. Адаптации человека к среде, как уже указывалось, проявляются в основном на социальном уровне, однако человечество на
ранних этапах эволюции подвергалось непосредственному действию биотических и абиотических экологических факторов в значительно большей степени по сравнению с современной эрой научно-технического прогресса. Комплексы таких факторов имели разнонаправленное действие на человеческие популяции. В результате в разных климато-географических зонах сформировались разнообразные адаптивные типы людей.
Адаптивный тип представляет собой норму биологической реакции на комплекс условий окружающей среды и проявляется в развитии морфофункциональных, биохимических и иммунологических признаков, обеспечивающих оптимальную приспособленность к данным условиям обитания.
В комплексы признаков адаптивных типов из разных географических зон входят общие и специфические элементы. К первым относят, например, показатели костно-мускульной массы тела, количество иммунных белков сыворотки крови человека. Такие элементы повышают общую сопротивляемость организма к неблагоприятным условиям среды. Специфические элементы отличаются разнообразием и тесно связаны с преобладающими условиями в данном месте обитания - гипоксией, жарким или холодным климатом. Именно их сочетание служит основанием к выделению адаптивных типов: арктического, тропического, зоны умеренного климата, высокогорного, пустынь и др.
Разберем особенности условий жизни человеческих популяций в различных климато-географических зонах и адаптивные типы людей, сформировавшиеся в них.
Условия обитания в Арктике характеризуются постоянно низкими температурами воздуха, скудной растительностью, богатством животного мира и сезонной периодичностью поступления продуктов питания: растительная пища доступна только в короткие летние месяцы, а животная - в периоды нереста рыбы, гнездования птиц, размножения оленей и морского зверя. Продукты питания богаты витаминами, белками, жирами и микроэлементами, но бедны углеводами растительного происхождения. Воздух и почва в Заполярье содержат очень мало микроорганизмов, низкие температуры препятствуют сохранению цист патогенных простейших, яиц и личинок гельминтов.
Из перечисленных факторов наибольшее влияние на формирование комплекса признаков арктического адаптивного типа оказали, по-видимому, холодный климат и преимущественно животная пища. Арктическому комплексу признаков свойственны относительно сильное развитие костно-мускульного компонента тела, большие размеры грудной клетки, высокий уровень гемоглобина, относительно большое
пространство, занимаемое костным мозгом, повышенное содержание минеральных веществ в костях, высокое содержание в крови белков, холестерина, повышенная способность окислять жиры. Среди аборигенов Арктики почти не встречаются лица с астеническим телосложением. Практически стерильные условия окружающей среды определяют относительно низкий уровень иммунитета
В целом арктический тип характеризуется усиленным энергетическим обменом, который отличается стабильностью показателей в условиях переохлаждения. Имеют свои особенности и механизмы терморегуляции. Так, при одинаковой степени охлаждения у канадских индейцев резко падает температура кожи, но уровень обмена веществ меняется незначительно, а у пришлого белого населения наблюдается меньшая степень снижения кожной температуры, но появляется сильная дрожь, т.е. интенсифицируется обмен.
В тропиках и субтропиках располагается чуть ли не большая часть Ойкумены. Этот регион отличается в целом большим количеством тепла и влаги и сглаженностью сезонных колебаний условий обитания. Вместе с тем благодаря особенностям рельефа наблюдается значительная контрастность распределения тепла и влаги - массивы влажных лесов нередко соседствуют с засушливыми плато, обширными равнинами и редколесьем. В экваториальной и субэкваториальной областях сосредоточены огромные количества растительной биомассы. Влажные тропические леса относительно бедны животными, тогда как в саваннах животный мир разнообразен и включает крупных стадных животных, издавна используемых человеком как объект охоты. Почва, воздух и вода содержат большое количество микроорганизмов, яиц гельминтов и цист патогенных простейших. Богатство и разнообразие животного мира обеспечивает существование огромного множества промежуточных и окончательных хозяев гельминтов и переносчиков возбудителей трансмиссивных заболеваний. К преобладающим экологическим факторам, под влиянием которых формировался комплекс признаков тропического адаптивного типа, относят жаркий влажный климат и рацион с относительно низким содержанием животного белка.
В тропической области наблюдается исключительно широкая вариабельность групп населения в расовом, этническом и экономическом отношениях. Это проявляется в поразительном размахе изменчивости, например по соматическим признакам. Тем не менее преобладающие экологические факторы, особенно климатический, способствовали образованию определенного комплекса морфофизиологических призна-
ков обитателей тропиков и субтропиков. К характерным признакам тропического типа относят удлиненную форму тела, сниженную мышечную массу, относительное уменьшение массы тела при увеличении длины конечностей, уменьшение окружности грудной клетки, более интенсивное потоотделение за счет повышенного количества потовых желез на 1 см2 кожи, низкие показатели основного обмена и синтеза жиров, сниженную концентрацию холестерина в крови (рис. 15.25).
Антропологическое изучение современных обитателей зоны умеренного климата под углом зрения формирования биологических механизмов адаптации к природным условиям затруднено, так как значительная часть людей проживает в промышленно развитых странах с большой долей городского населения. Результаты наблюдения позволяют, однако, судить о том, что и в этом случае в процессе исторического развития человеческих популяций сформировался комплекс признаков, соответствующий особому адаптивному типу умеренного пояса.
По соматическим показателям, уровню основного обмена население умеренного пояса занимает промежуточное положение между коренными жителями арктического и тропического регионов. Это соответствует условиям биогеографической среды в зоне умеренного климата. Для нее характерны неравномерное распределение районов, отличающихся по количеству тепла и влаги, типу растительности (от сухих степей и полу-
Рис. 15.25. Адаптивные типы человека и большие расы.

пустынь до тайги), богатству животного мира. Вместе с тем температура и влажность воздуха здесь не достигают экстремальных величин, хорошо выражен сезонный ритм биоклиматических условий.
Условия высокогорья для человека во многих отношениях экстремальны. Их характеризуют низкое атмосферное давление, сниженное парциальное давление кислорода, холод, относительное однообразие пищи. Основным экологическим фактором формирования горного адаптивного типа стала, по-видимому, гипоксия. У жителей высокогорья независимо от климатической зоны, расовой и этнической принадлежности наблюдаются повышенный уровень основного обмена, относительное удлинение длинных трубчатых костей скелета, расширение грудной клетки, повышение кислородной емкости крови за счет увеличения числа эритроцитов, содержания гемоглобина и относительной легкости его перехода в оксигемоглобин.
15.9.3. ПРОИСХОЖДЕНИЕ АДАПТИВНЫХ ЭКОЛОГИЧЕСКИХ ТИПОВ
Человечество возникло в тропической зоне Африки, и поэтому следует полагать, что наиболее древним и исходным для остальных экологических типов человека является тропический тип. Об этом свидетельствует и наибольший полиморфизм морфофункциональных признаков человека в рамках этого типа на территории Центральной и Северо-Восточной Африки. В связи с этим уместно обратиться к учению Н.И. Вавилова о центрах происхождения и многообразия культурных растений, в соответствии с которым именно в зонах первоначального вовлечения биологических видов в социальную среду наблюдается наиболее выраженный наследственный полиморфизм местных популяций по большому числу признаков.
Об этом свидетельствует и то, что именно в тропической Африке в непосредственной близости друг от друга, т.е. под действием сходных экологических факторов, обитают наиболее низкорослые и наиболее высокорослые племена, например пигмеи, готтентоты и бушмены, с одной стороны, и масаи - с другой. Пределы изменчивости африканских популяций, например, по росту и массе тела таковы, что в них укладываются все известные на Земле человеческие популяции. Основные же черты тропического типа в Африке, отмеченные выше, остаются неизменными. Они же характеризуют монголоидные популяции Индокитая, Малайского архипелага и некоторые группы индейских племен зоны влажных тропических лесов Центральной и Южной Америки.
Это указывает на то, что тропический адаптивный тип развивается в результате асинхронного параллелизма в эволюции человека, причем вна-
чале на африканском континенте, а потом и в других областях (см. п. 3.13). Отсюда следует и еще один вывод: адаптивный тип формируется на фоне расогенеза и вне зависимости от него. В процессе адаптогенеза в популяциях человека можно проследить и проявление закона гомологических рядов (см. п. 13.3.5). Это выражается, например, в том, что в бассейне Меконга в Индокитае и на острове Суматра имеются пигмеоидные популяции, по антропометрическим признакам соответствующие африканским пигмеям.
Экологический тип умеренного пояса сформировался на базе исходного генетического и фенотипического полиморфизма тропического типа при расселении популяций человека в умеренных зонах Евразии и позже - Северной Америки. Он оформился в рамках двух больших рас: европеоидной и монголоидной.
При заселении человеком арктической зоны в Евразии и Северной Америке произошло формирование арктического типа. Независимо от него среди индейцев Южной Патагонии и Огненной Земли в приантаркти-ческой зоне Южной Америки возникли популяции индейцев, по основному комплексу признаков соответствующие арктическому типу. Это еще один убедительный пример параллелизма эволюции человеческих популяций и реализации закона гомологических рядов, а также доказательство вторич-ности адаптивных типов по отношению к большим расам человечества.
Формирование горного адаптивного типа иллюстрирует общие закономерности адаптогенеза, отмеченные выше. Этот тип также развился независимо от расовой и этнической принадлежности популяций - среди европеоидов Альп, Кавказа, Памира и Гималаев, а также в монголоидных популяциях Тибета, Тянь-Шаня и Анд. В связи с тем, что высокогорья заселялись человеком в последнюю очередь, горный экологический тип является по происхождению самым молодым. Интересно, что, несмотря на особенно выраженную расовую и этническую разнородность этого типа, комплекс основных признаков его монолитен. Однако в отличие от других типов, вероятно, именно горный проявляется в основном только на фенотипическом уровне и не имеет наследственной природы. Об этом свидетельствует то, что число эритроцитов в крови и объем грудной клетки людей, переселяющихся в условия высокогорья и обратно, могут меняться на протяжении жизни одного поколения.
Следовательно, адаптивные типы человека не только отражают его прошлое, но формируются и в настоящем, а разные типы имеют и разную по длительности историю.
В настоящее время пока еще на фенотипическом уровне идет формирование адаптивного типа человека городской среды, который характеризуется широкой лабильностью психических реакций, обе-
спечивающих способность относительно легко переживать состояния практически постоянного стресса, вызванного высокой плотностью населения и рядом других морфофизиологических особенностей, оптимальных для жизни в специфических условиях города (см. п. 17.3.2).
Формирование экологических типов человека в значительной степени обеспечило всесветное расселение людей. Меняющаяся среда обитания под действием антропогенных факторов ставит перед популяциями человека новые задачи, решаемые и сегодня за счет не только социальных, но и биологических адаптации. Схему эволюционных взаимоотношений больших рас и адаптивных типов человека см. на рис. 15.26.

Вопросы для самоконтроля
1. По каким признакам человек относится к отряду Приматы?
2. Какие методы применяются для изучения происхождения и дальнейшей эволюции человека?
3. Каковы основные механизмы антропогенеза?
4. Какой континент является древней родиной человечества?
5. Какие методы используются для изучения происхождения и эволюции человека?
6. Охарактеризуйте основные этапы антропогенеза.
7. Является ли неандертальский человек непосредственным предком человека разумного? Ответ обоснуйте.
8. Внутривидовая дифференцировка человечества: расы и антропологические типы.
9. Концепции расовой классификации человечества.
10. Адаптивные типы человека и их происхождение.