Биология: учебник: в 2 т. / под ред. В. Н. Ярыгина. - 2011. - Т. 2. - 560 с. : ил.
|
|
|
|
Глава 14. ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ ХОРДОВЫХ
Филогенез систем органов хордовых рассмотрен в соответствии с прогрессивным направлением эволюции этого типа животных от подтипа Бесчерепные до класса Млекопитающие. Организация систем органов класса Птицы не описана в связи с тем, что птицы произошли от пресмыкающихся значительно позже млекопитающих и являются боковой ветвью эволюции хордовых.
14.1. НАРУЖНЫЕ ПОКРОВЫ
Покровы любых животных всегда выполняют функцию восприятия внешних раздражений, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции также сопровождается дифференцировкой. Кроме того, характерно расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное - экто- и мезодермальное - происхождение. Из эктодермы развивается эпидермис, из мезодермы - дерма. Для бесчерепных характерна слабая степень дифференцирован-ности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях - дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
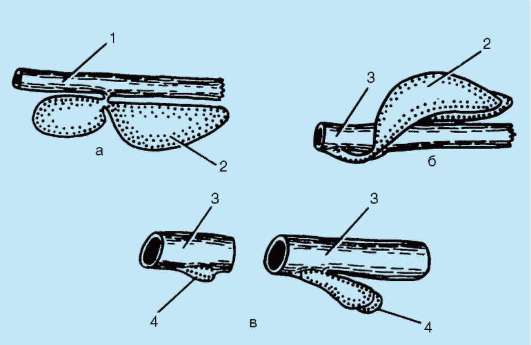
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 14.1). Дентин мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся наружу в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных - стегоцефалов - соответствовала покровам рыб и тоже была покрыта чешуей. Современ-
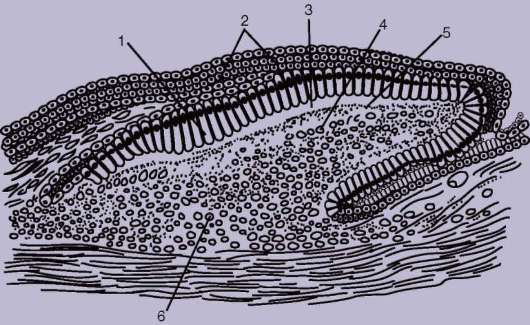
Рис. 14.1. Закладка плакоидной чешуи: 1 - клетки-эмалеобразователи; 2 - эпидермис; 3 - эмаль; 4 - склеробласты-дентинообразователи; 5 - дентин; 6 - сосочек дермы
ные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого числа многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов (см. п. 23.1).
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других - сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие - насекомоядные, грызуны и некоторые другие - наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3-7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 14.2), покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у человека.
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы, основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функ-
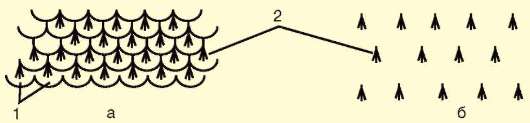
Рис. 14.2. Расположение волос у млекопитающих: а - на хвосте грызунов; б - на других участках тела; 1 - роговые чешуйки; 2 - группы волос, расположенные в шахматном порядке
ции передних конечностей, у многих яйцекладущих и сумчатых - разбросаны по всему телу. Это может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы. У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 14.3). Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает (рис. 14.4). Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна.
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
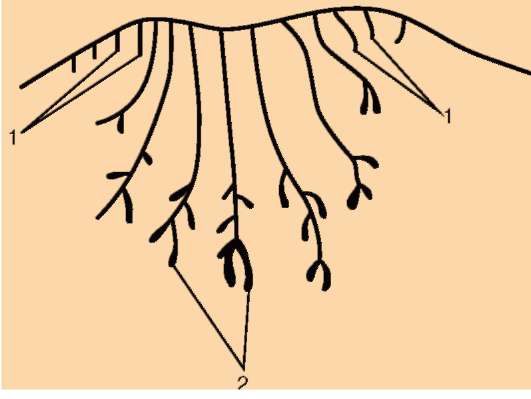
Рис. 14.3. Строение развивающегося соска млекопитающего. Виден постепенный переход от потовых (1) к млечным (2) железам
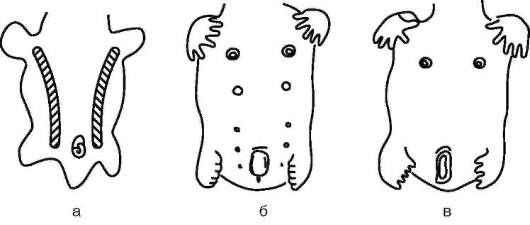
Рис. 14.4. Закладка и развитие млечных желез у зародыша человека: а - зародыш в возрасте 5 нед (видны млечные линии); б - дифференцировка пяти пар сосков; в - зародыш в возрасте 7 нед
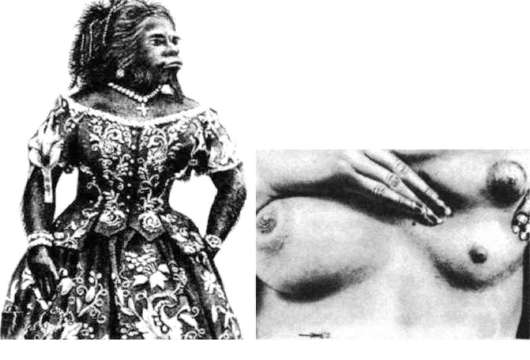
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуй, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития (рис. 14.5): гипертрихоз (повышенное оволосение), полителию (увеличенное число сосков), полимастию (увеличенное число млечных желез). Все они связаны с нарушением редукции избыточного числа этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами - млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Один из самых известных признаков недоношенности новорожденных - повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
а б
Рис. 14.5. Атавистические аномалии развития покровов у человека: а - гипертрихоз; б - полимастия
14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата, претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей.
Рассмотрим в отдельности эволюцию скелета и мышечной системы.
14.2.1. СКЕЛЕТ
У хордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы.
14.2.1.1. Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей - хордом, возникающих на его основе.
У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов (см. п. 13.4). Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб, обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Тело каждого позвонка образуется из остеогенной ткани склеротомов двух соседних сомитов таким образом, что передняя половина каждого позвонка развивается из задней части переднего сомита, а задняя - из передней части заднего. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка (рис. 14.6).
Замещение хорды позвоночным столбом - более мощным органом опоры, имеющим сегментарное строение,- позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией - заменой хрящевой ткани на костную, начиная с костных рыб, а также с дифференцировкой его на отделы.
У рыб только два отдела позвоночника: туловищный и хвостовой. Это связано с перемещением их в воде за счет изгибов тела.
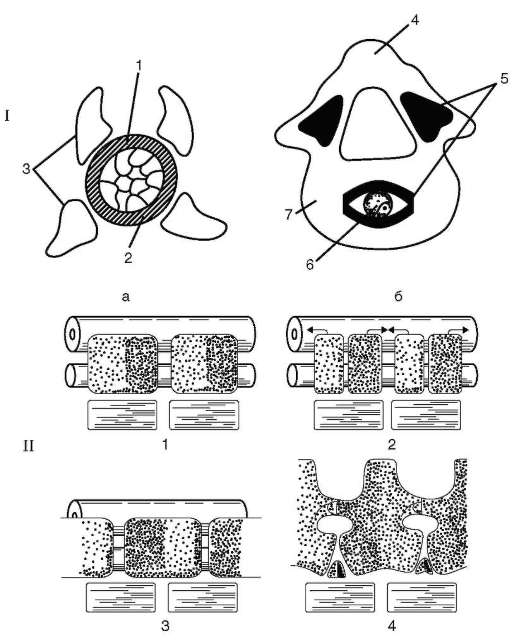
Рис. 14.6. Развитие позвонков у позвоночных: I - фронтальная проекция: а - ранний этап; б - последующая стадия; 1 - хорда; 2 - оболочка хорды; 3 - верхние и нижние позвонковые дуги; 4 - остистый отросток; 5 - зоны окостенения; 6 - рудимент хорды; 7 - хрящевое тело позвонка; II - боковая проекция: 1 - сегменты дорзальной мезодермы; 2 - парные закладки тел позвонков в каждом сегменте; 3 - формирование тел позвонков за счет сращения их зачатков из соседних сегментов; 4 - тела позвонков, сформированные из парных зачатков соседних сегментов
Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй - опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным числом позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5-10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения, - у хвостатых земноводных и пресмыкающихся, поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12-13 пар ребер.
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spina bifida - дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа (рис. 14.7).
Нарушение сращения парных зачатков тел позвонков, развивающихся из двух соседних сомитов, приводит к формированию так называемых hemivertebrae, или полупозвонков.
В возрасте 1,5-3 мес зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8-11 позвонков. Нарушение их редукции
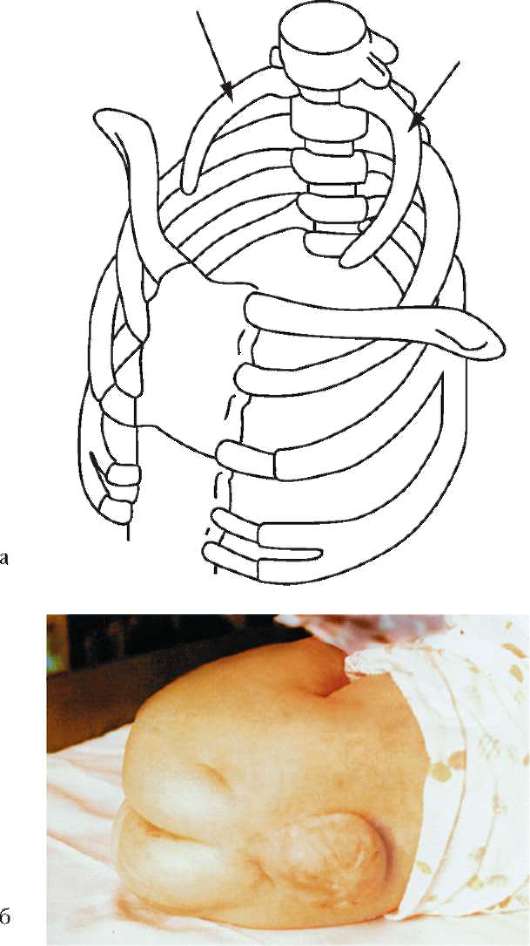
Рис. 14.7. Аномалии развития осевого скелета: а - рудиментарные шейные ребра (показаны стрелками); б - несращение остистых отростков позвонков в поясничной и крестцовой областях. Спинномозговая грыжа (фото)
объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
14.2.1.2. Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой, череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезен-химального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную.
У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.
Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю.
У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Об этом свидетельствует появление большого числа зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с метопическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и поэтому обнаруживаются обычно случайно после смерти.
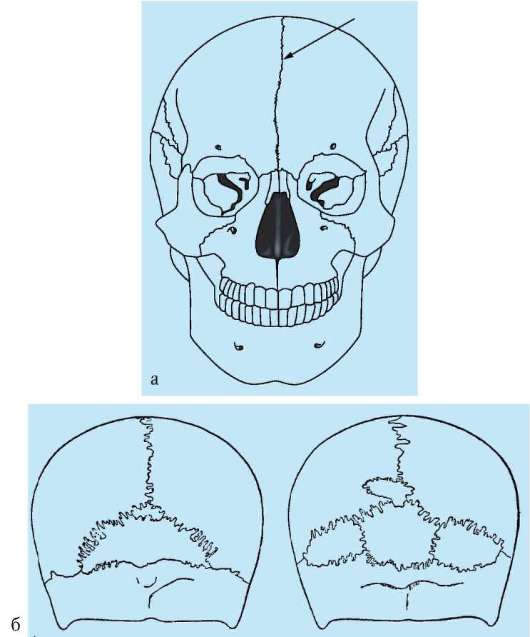
Рис. 14.8. Аномалии развития мозгового черепа человека: а - череп с метопи-ческим швом; б - черепа людей с добавочными костями в затылочной области
Дефект сращения теменных костей мозгового черепа называется cranium bifidum. При этом у новорожденного почти всегда наблюдается грыжа головного мозга.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мигрирующих клеток передней части нервного гребня, то есть из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной ду-
гам взрослых животных. Следующие дуги в числе 4-5 пар выполняют опорную функцию для жабр и называются жаберными (рис. 14.9, а). Предпосылка таких изменений судьбы клеток нервного гребня - их освобождение от участия в формировании сегментарных чувствительных спинальных ганглиев, отсутствующих в головном отделе тела позвоночных. Кроме того, именно у них при образовании переднего отдела нервной трубки формируется особенно обширная масса клеток нервного гребня.
У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1-2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее число висцеральных дуг, чем 6 или 7, а их дифференцировка происходила на фоне олигомеризации.
Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ - первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибу-лярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвески к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным.
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей - челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гио-стильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом - нёбно-квадратным хрящом - срастается полностью с основанием мозгового черепа, и череп становится, таким образом, ау-тостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвески, располагаясь в области первой
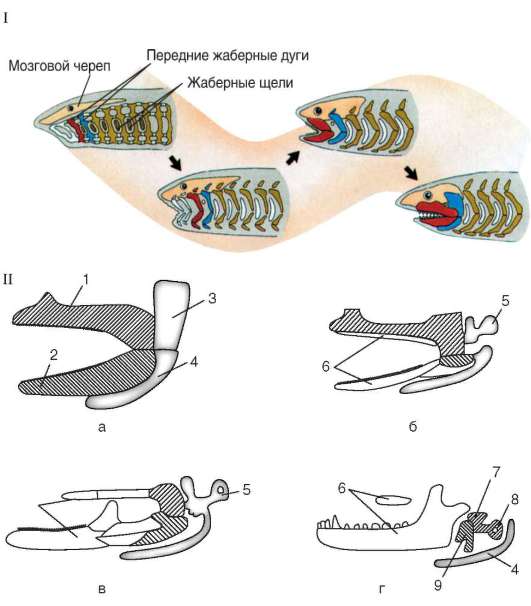
Рис. 14.9. Происхождение и строение висцерального черепа позвоночных: I- передний конец тела гипотетического предка (1) и современной хрящевой рыбы; II - эволюция двух первых висцеральных жаберных дуг позвоночных и происхождение челюстей и слуховых косточек: а - хрящевая рыба; б - земноводное; в - пресмыкающееся; г - млекопитающее: 1 - небно-квадратный хрящ; 2 - меккелев хрящ; 3 - гиомандибулярный хрящ; 4 - гиоид; 5 - столбик; 6 - накладные кости вторичных челюстей; 7 - наковаленка; 8 - стремечко; 9 - молоточек; гомологичные образования обозначены соответствующей штриховкой; в - строение среднего и внутреннего уха: 1 - пресмыкающего; 2 - млекопитающего
жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки - столбика, - передающей звуковые колебания от наружного к внутреннему уху.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения.
Одна слуховая косточка - столбик, характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9, б, в).
Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки - механизм формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки - столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
14.2.1.3. Скелет конечностей
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности - грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных.
Подробнее рассмотрим происхождение и эволюцию парных конечностей .
У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплев-ральными (рис. 14.10). Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль - стабилизацию положения тела
и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.
В результате из передних отделов складок развились грудные, а из задних - брюшные плавники (см. рис. 14.10). Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными парными плавниками (рис. 14.11). За счет слияния оснований хрящевых лучей возникли плечевой и тазовый пояса. Дистальные их участки дифференцировались в скелет свободных конечностей.

Рис. 14.10. Формирование передних и задних парных конечностей из мета-плевральных складок: I-III - гипотетические этапы эволюции

Рис. 14.11. Древняя акулообразная рыба с многочисленными парными конечностями
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое число расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее число их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7-12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными (рис. 14.12, а, б) и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату.
Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания (см. п. 14.3.4) стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего стало происхождение земноводных и всей группы Tetrapoda. Первые их представители - стегоцефалы - обладали семи-, шестиили пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб (см. рис. 14.12, в). В скелете запястья сохранено правильное радиальное расположение костных элементов в 3-4 ряда, в пясти располагается
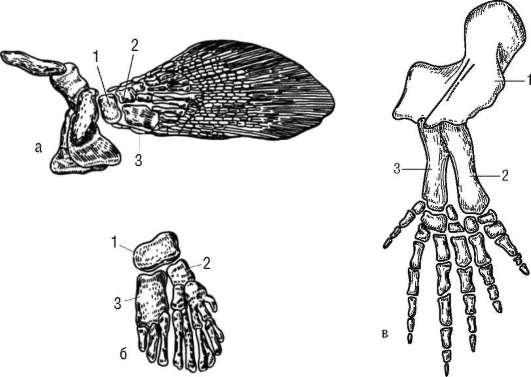
Рис. 14.12. Скелет передней конечности кистеперой рыбы (а), его основание (б) и скелет передней лапы стегоцефала (в): 1 - плечевая кость; 2 - локтевая кость; 3 - лучевая кость
7-5 костей, а далее также радиально лежат фаланги 7-5 пальцев. Интересно, что у всех Тетрапод в развитии передних парных конечностей принимает участие один и тот же ген Tbx5. Он экспрессируется в почках будущих конечностей в раннем эмбриональном развитии, производя специфический белок T-box, регулирующий транскрипцию ряда структурных генов, участвующих в контроле морфогенеза. Дифференцировку задних конечностей контролирует ген Tbx4. Различные формы конечностей таких наземных позвоночных, как пресмыкающиеся, птицы или человек, развиваются в зависимости от того, с какими конкретно генами взаимодействует фактор T-box (рис. 14.13).
У современных земноводных число пальцев в конечностях равно пяти или происходит их олигомеризация до четырех.
Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении числа костей в запястье сначала до трех рядов у амфибий и затем до двух - у пресмыкающихся и млекопитающих. Параллельно уменьшается также и число фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных.
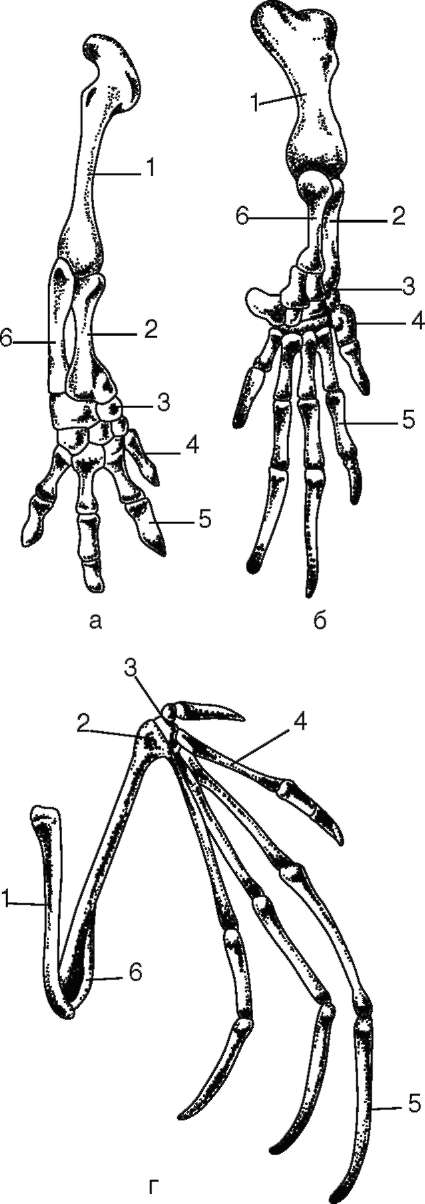
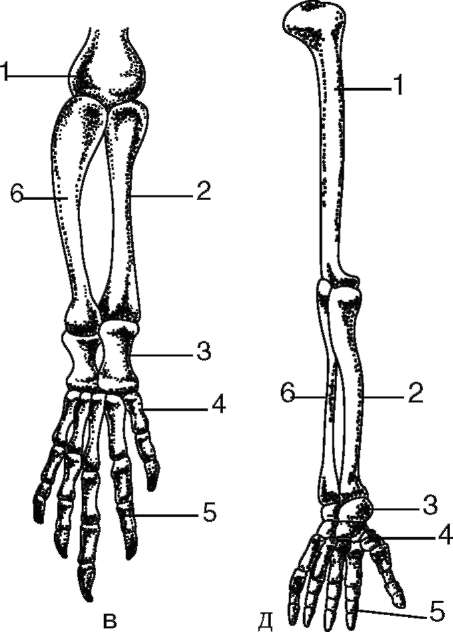
Рис. 14.13. Скелет передней конечности наземных позвоночных: а - лягушка; б - саламандра; в - крокодил; г - летучая мышь; д - человек; 1 - плечевая кость; 2 - лучевая кость; 3 - кости запястья; 4 - кости пясти; 5 - фаланги пальцев; 6 - локтевая кость
Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей.
Многообразие условий обитания, предоставляемых наземным образом жизни, обеспечивает многообразие форм передвижения: прыжки, бег, ползание, полет, рытье, лазание по скалам и деревьям, а при возвращении в водную среду - и плавание. Поэтому у наземных позвоночных можно встретить как почти неограниченное многообразие конечностей, так и их полную вторичную редукцию, причем многие сходные адаптации конечностей в разнообразных средах многократно возникали конвергентно. Однако в процессе онтогенеза у большинства наземных
позвоночных проявляются общие черты в развитии конечностей: закладка их зачатков в виде малодифференцированных складок, формирование в кисти и стопе вначале шести или семи зачатков пальцев, крайние из которых вскоре редуцируются и в дальнейшем развивается только пять (рис. 14.14).
В онтогенезе человека возможны многочисленные нарушения, ведущие к формированию врожденных пороков развития конечностей атавистического плана. Наиболее распространенная аномалией такого рода - полидактилия, или увеличение числа пальцев. Различают несколько форм этой аномалии. Одна и наиболее частая из них - развитие добавочных пальцев на латеральных сторонах ладони перед большим пальцем, либо за мизинцем. Она называется соответственно пре- и постаксиальной. Эта аномалия наследуется как аутосомно-доминантный
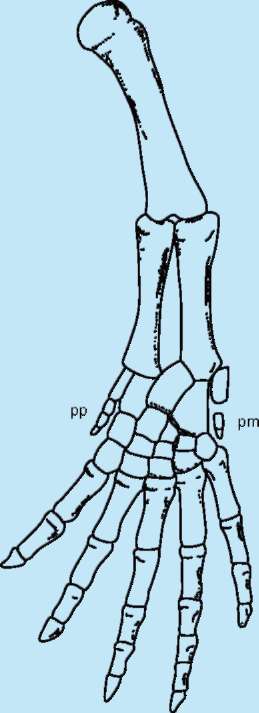
Рис. 14.14. Строение развивающейся конечности позвоночного: pp - prepollex; pm - postminimus - рудиментарные закладки дополнительных пальцев
признак, и является результатом нарушения редукции и дальнейшим развитием закладок дополнительных пальцев, характерных в норме для далеких предковых форм. Интересно, что у классических представителей собак породы бриар (французская овчарка) породным признаком является наличие семи пальцев на задних конечностях, причем оба дополнительных пальца расположены преаксиалярно. В связи с доминантностью наследования этого признака в процессе искусственного отбора он мог быстро закрепиться и широко распространиться у всех особей породы. Другая или аксиальная полидактилия, характеризуется развитием дополнительного среднего пальца. Она связана с мутацией регуляторной последовательности ДНК, контролирующей функцию гомеозисного гена Sonic-hedgehog, определяющего осевое строение конечностей позвоночных. Крайне редкая форма аномалии - полидактилия, сопровождающаяся изодактилией, при которой отсутствует дифференцировка пальцев. Она может сопровождаться синдактилией - полным или неполным сращением пальцев (рис. 14.15 и 14.16). В основе формирования синдактилии лежит нарушение избирательной клеточной гибели в межпальцевых промежутках. Известен также феномен полифалангии, характеризующийся увеличением числа фаланг, обычно большого пальца. В основе его возникновения - развитие трех фаланг в первом пальце, как это в норме наблюдается у пресмыкающихся и земноводных с недифференцированными пальцами конечностей. Двусторонняя полифалангия наследуется аутосомно-доминантно.
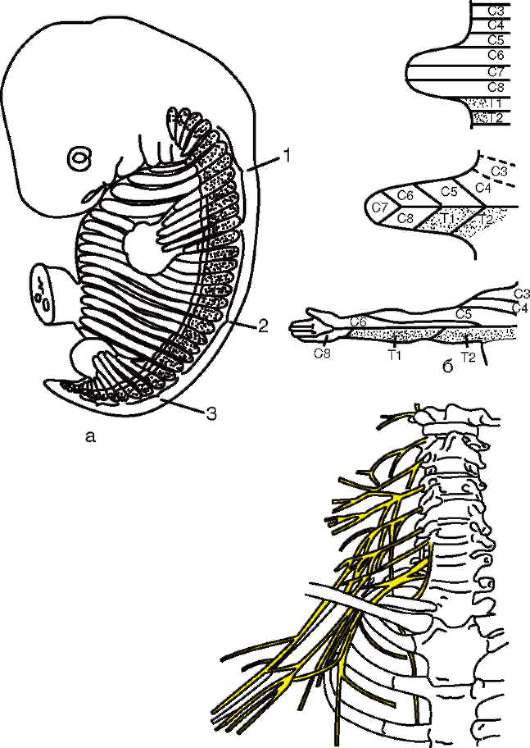
Интересно, что в эмбриогенезе высших позвоночных рекапитулирует не только строение конечностей предков, но и процесс их гетеротопии. Так, у человека верхние конечности закладываются на уровне 3-4-го шейных позвонков, а нижние - на уровне поясничных позвонков. В это же время конечности получают иннервацию из соответствующих от-
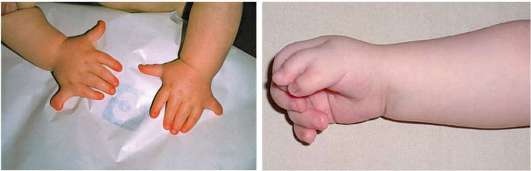
Рис. 14.15. Латеральная (параксиальная) полидактилия у человека
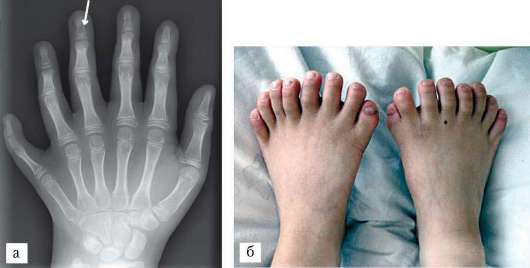
Рис. 14.16. Редкие формы полидактилии у человека: а - аксиальная (стрелкой показан дополнительный средний палец); б - полидактилия, сопровождающаяся изодактилией на нижних конечностях
делов спинного мозга. Гетеротопия конечностей сопровождается формированием шейного, поясничного и крестцового нервных сплетений, нервы которых связаны с одной стороны с теми сегментами спинного мозга, из которых они вырастали в момент формирования конечностей, а с другой - с конечностями, переместившимися на новое место (рис. 14.17; см. также п. 14.2.2.2).
Серьезный порок развития - нарушение гетеротопии пояса верхних конечностей из шейной области на уровень 1-2-го грудных позвонков. Эту аномалию называют болезнью Шпренгеля или врожденным высоким стоянием лопатки (рис. 14.18). Она выражается в том, что плечевой пояс с одной либо с двух сторон находится выше нормального положения на несколько сантиметров. В связи с тем, что такое нарушение часто сопровождается аномалиями ребер, грудного отдела позвоночника и деформацией лопаток, следует думать, что механизмы его возникновения - не только нарушение перемещения зародышевых структур, но и обусловленное этим нарушение морфогенетических корреляций (см. п. 13.4).
Сравнительно-анатомический обзор эволюции скелета хордовых свидетельствует о том, что скелет человека полностью гомологичен опорному аппарату предковых и родственных форм. Поэтому многие пороки его развития у человека можно объяснить родством млекопи-
в
Рис. 14.17. Формирование передних конечностей, их гетеротопия и иннервация в онтогенезе человека: а - врастание шейных миотомов в формирующуюся переднюю конечность зародыша; б - развитие кожной иннервации руки; в - расположение шейного и плечевого сплетений, участвующих в иннервации руки: 1 - шейные миотомы; 2 - грудные миотомы; 3 - поясничные миотомы; буквами C и T, обозначены шейные и грудные сегменты
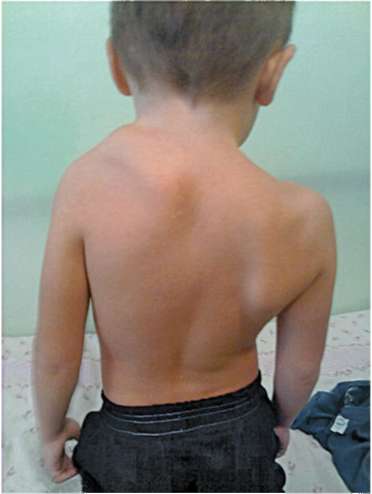
Рис. 14.18. Болезнь Шпренгеля (пояснения в тексте)
тающих с пресмыкающимися, земноводными и рыбами. Однако в процессе антропогенеза появились такие особенности скелета, которые характерны лишь для человека и связаны с его пря-мохождением и трудовой деятельностью. К ним относят:
• изменения стопы, переставшей выполнять хватательную функцию, выражающиеся в потере способности к противопоставлению большого пальца и появлению ее сводов, служащих для амортизации при ходьбе;
• изменения позвоночного столба - его S-образный изгиб, обеспечивающий пластичность движений в вертикальном положении;
• изменения черепа - резкое уменьшение его лицевой части и увеличение
мозговой, смещение большого затылочного отверстия кпереди, увеличение сосцевидного отростка и сглаживание затылочного рельефа, к которому прикрепляются мышцы шеи и выйная связка;
• специализация верхних (передних) конечностей как органа труда и выключение их из локомоции. Так, на руки человека приходится от 7 до 9% массы тела, а на ноги - 32-38%, в то время как у африканских человекообразных обезьян передние конечности составляют 14-16%, а задние - около 18%;
• появление подбородочного выступа нижней челюсти в связи с развитием членораздельной речи.
Перечисленные особенности имеют характер анаболий и девиаций.
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к пря-мохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохож-дению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формиру-
ются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Фактически это анаболии, возникшие в ходе филогенеза приматов. Поэтому атавистические аномалии скелета, связанные с задержками развития признаков, характерных только для человека, встречаются чаще всего. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.
14.2.2. МЫШЕЧНАЯ СИСТЕМА
У представителей типа Хордовые мускулатура подразделяется по характеру развития и иннервации на соматическую и висцеральную.
Соматическая мускулатура развивается из миотомов и иннерви-руется нервами, волокна которых выходят из спинного мозга в составе брюшных (передних) корешков спинномозговых нервов. Висцеральная мускулатура развивается из других участков мезодермы и иннер-вируется нервами вегетативной нервной системы. Вся соматическая мускулатура поперечнополосатая, а висцеральная может быть как поперечнополосатой, так и гладкой (рис. 14.19).
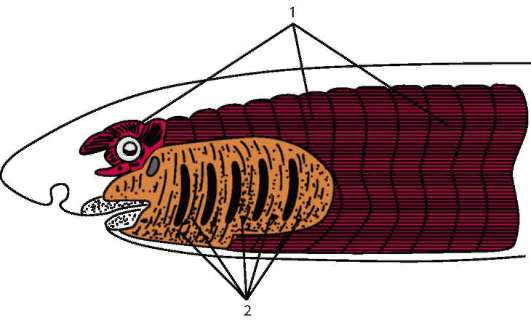
Рис. 14.19. Соматическая и висцеральная мускулатура позвоночных: 1 - соматическая мускулатура, развивающаяся из миотомов; 2 - висцеральная мускулатура жаберной области
14.2.2.1. Висцеральная мускулатура
Наиболее существенные изменения претерпела висцеральная мускулатура, связанная с висцеральными дугами передней части пищеварительной трубки. У низших позвоночных большая часть этой мускулатуры представлена общим сжимателем висцерального аппарата - m. constrictor superficialis, покрывающим всю область жаберных дуг со всех сторон. В области челюстной дуги эта мышца иннервируется тройничным нервом (V), в области подъязычной дуги - лицевым (VII), в области первой жаберной дуги - языкоглоточным (IX), наконец, ее часть, лежащая каудальнее,- блуждающим нервом (X). В связи с этим все производные соответствующих висцеральных дуг и мышц, связанных с ними, иннервируются впоследствии у всех позвоночных перечисленными нервами.
В передней части сжимателя обособляется крупная мускульная масса, обслуживающая челюстной аппарат. Позади висцерального аппарата дифференцируется трапециевидная мышца m. trapezius, отдельными пучками прикрепляющаяся к последним жаберным дугам и переднему краю спинного отдела плечевого пояса. Часть поверхностного сжима-теля в области подъязычной дуги у пресмыкающихся разрастается, охватывает шею снизу и с боков и образует сжиматель шеи m. sphincter colli. У млекопитающих эта мышца делится на два слоя: глубокий и поверхностный. Глубокий сохраняет прежнее название, а поверхностный называется platysma myoides и располагается подкожно. Эти две мышцы разрастаются на всю область головы и дают начало сложной системе лицевой подкожной мускулатуры, которую у приматов и человека называют мимической. Поэтому вся мимическая мускулатура иннерви-руется так же, как и мышца, из которой она происходит, - лицевым нервом.
Мышцы собственно жаберного аппарата с утратой жаберного дыхания редуцируются, но отдельные их элементы сохраняются в виде мышц подъязычного аппарата, глотки и гортани. Трапециевидная мышца полностью теряет связь с жаберным аппаратом и становится исключительно мышцей плечевого пояса. Часть ее у млекопитающих отходит от сосцевидного отростка черепа и прикрепляется к ключице и грудине, обособляется - грудино-ключично-сосцевидная мышца m. sternocleidomastoideus. Иннервирующая эту мышцу задняя часть блуждающего нерва становится самостоятельным черепным нервом X пары, добавочным - n. accessorius.
Основные этапы филогенеза висцеральной мускулатуры жаберной области рекапитулируют в эмбриогенезе млекопитающих, в том числе человека. Знание этих рекапитуляций позволяет объяснить сложность иннервации мышц лица и шеи, объединенных с ними общностью происхождения.
14.2.2.2. Соматическая мускулатура
14.2.2.2-а. Мускулатура головы
У всех позвоночных животных в процессе эмбриогенеза нижние концы миотомов образуют выросты в вентральном направлении, охватывающие полость тела снаружи и срастающиеся по центральной линии на брюшной стороне. Таким образом, закладки соматической мускулатуры становятся сегментарными не только на дорсальной стороне в связи с сегментацией сомитов, но и на вентральной. В миотомах и в их вентральных отростках идет образование продольных мускульных волокон.
Миотомы, лежащие на головном конце тела, распадаются на мезенхиму и образуют зачатки отдельных мышц. Из первого миотома головы формируются верхняя внутренняя и нижняя прямая и нижняя косая мышцы глаза, иннервируемые глазодвигательным нервом n. oculomotorius (III пара). Из второго миотома - верхняя косая мышца, иннервируемая блоковым нервом n. trochlearis (IV пара); а из третьего - наружная прямая мышца, получающая иннервацию от отводящего нерва n. abducens (VI пара).
Задние миотомы головы, образующие мощные вентральные отростки, распространяющиеся вокруг висцеральной мускулатуры глоточной области, образуют подъязычную мускулатуру, которая у рыб слабо дифференцирована, а начиная с земноводных распадается на m. sternohyoideus, m. omohyoideus и m. geniohyoideus. У наземных позвоночных за счет последней мышцы формируется собственная мускулатура языка - m. genioglossus и m. hyoglossus. Вся подъязычная мускулатура иннервируется подъязычным нервом n. hypoglossus, который у амниот становится типичным черепным нервом.
14.2.2.2-6. Мускулатура туловища и конечностей
У бесчерепных, а также у рыб вся мускулатура туловища состоит из ряда мышечных сегментов, или миомеров, правой и левой сторон, которые вместе образуют так называемые боковые мышцы. Каждый миомер развивается из миотома одного сомита и иннервируется перво-
начально двигательной ветвью одного спинномозгового нерва. Мио-меры отделены друг от друга миосептами - соединительнотканными перегородками. Такой же перегородкой, идущей продольно, боковая мышца разделена на спинную и брюшную мышцы.
Уже у рыб направление пучков мышечных волокон в миомерах начинает изменяться на разной глубине мышечного слоя. Эта диффе-ренцировка значительно более выражена у наземных позвоночных и приводит у них к постепенному обособлению различных слоев брюшной и спинной мускулатуры. В результате этого возникают сложные группы мышц, от первоначальной четкой метамерии которых остаются только следы в виде глубоких мышц спины и шеи, связывающих друг с другом соседние позвонки. Значение спинной мускулатуры наземных позвоночных снижается в связи с передвижением большинства из них с помощью конечностей, а брюшные мышцы претерпевают смену функций: первоначально принимающие участие в перемещениях тела, они у пресмыкающихся и млекопитающих служат для изменения объема грудной и брюшной полостей в процессе дыхания.
Мускулатура парных плавников рыб закладывается в виде ряда мускульных почек, вырастающих от вентральных концов миотомов. Каждая из этих почек подразделяется на два мышечных зачатка, врастающих в основание закладки плавника с его спинной и брюшной сторон. Первые функционально становятся мышцами, отводящими плавник, вторые - приводящими. У наземных позвоночных из мышечного зачатка, гомологичного отводящей мышце плавника, развивается группа разгибателей пятипалой конечности, а из зачатка ее антагонистов - группа сгибателей. В пределах каждой группы идет дифференцировка на поверхностные и глубокие мышечные пучки, становящиеся самостоятельными мышцами. В целом мышцы наземных позвоночных, гомологичные мышцам плавников рыб, образуют первичную мускулатуру конечностей. Она иннервируется нервами плечевого и пояснично-крестцового сплетений, образованных в процессе перемещения поясов конечностей в ходе эмбриогенеза (см. п. 14.2.1).
При дальнейшей дифференцировке миотомов грудной клетки развивается группа мышц, приводящая в движение сам плечевой пояс, или вторичная мускулатура. К ней относят широчайшую мышцу спины, большую и малую грудные, а также зубчатую мышцы. Они иннервируют-ся непосредственно спинномозговыми нервами сегментов спинного мозга, расположенных каудальнее тех его участков, которые осуществляют иннервацию первичной мускулатуры. Задняя пара конечностей не имеет
вторичной мускулатуры в связи с тем, что гетеротопия тазового пояса по отношению к позвоночнику в процессе эволюции менее выражена.
Изменение среды обитания и характера движений позвоночных привело к усилению и обособлению большого числа мышц, обслуживающих конечности, и к относительной редукции собственно мышц туловища. Такие мышцы, как грудные, широчайшая мышца спины и трапециевидная, в значительной мере покрывают туловищную мускулатуру и даже частично вытесняют ее функционально.
14.3. ПИЩЕВАРИТЕЛЬНАЯ И ДЫХАТЕЛЬНАЯ СИСТЕМЫ
Уникальная особенность организации хордовых - филогенетическая, эмбриогенетическая, а также функциональная связь пищеварительной и дыхательной систем. Действительно, только у хордовых дыхательная система развивается на базе пищеварительной и на первых этапах эволюции функционирует совместно с ней. Так, у ланцетника, сохранившего в значительной степени черты организации предковых форм, специализированных органов дыхания нет, а дыхательную функцию выполняет глотка - передняя часть пищеварительной трубки, пронизанная сквозными отверстиями - жаберными щелями, главной функцией которых является фильтрация воды. Челюстного аппарата у ланцетника нет, и активно питаться он не может. Поэтому источник его питания - взвесь органических частиц в воде, которые задерживаются жаберными перегородками, прилипая к слизи, обильно выделяемой железистыми клетками глотки. Постоянный ток свежей воды через глотку способствует газообмену в кровеносных сосудах, расположенных в жаберных перегородках. На капилляры эти сосуды не распадаются, что свидетельствует о второстепенности дыхательной функции глотки.
У более высокоорганизованных хордовых, начиная с рыб, пищеварительная и дыхательная функции осуществляются специализированными системами, объединенными анатомически общей полостью рта и глотки, а также развитием из общего энтодермального зачатка.
Тесная связь обеих систем в филогенезе определяется в первую очередь их топографическими и динамическими координациями, а развитие в онтогенезе - морфогенетическими и эргонтическими корреляциями (рис. 14.20). Пищеварительная и дыхательная системы хордовых в эмбриогенезе закладываются вначале в виде прямой трубки, подразделяющейся на три участка. Переднюю ее часть, начинающуюся ротовым отверстием и заканчивающуюся переходом в глотку, называют
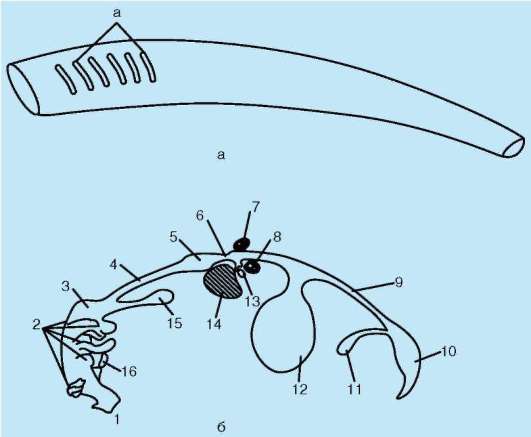
Рис. 14.20. Кишечная трубка позвоночных (а) и ее дифференцировка в эмбриогенезе человека (б): а - жаберные щели в глотке; 1 - ротовая полость; 2 - 1-5-й глоточные карманы; 3 - глотка; 4 - пищевод; 5 - желудок; 6 - двенадцатиперстная кишка; 7 - дорсальная поджелудочная железа; 8 - вентральная поджелудочная железа; 9 - тонкая кишка; 10 - клоака; 11 - мочевой пузырь; 12 - желточный мешок; 13 - желчный пузырь; 14 - печень; 15 - легкие; 16 - закладка щитовидной железы
stomodeum. Слизистая оболочка, выстилающая этот участок, эктодер-мального происхождения и в развитии связана с кожным эпидермисом и его производными (см. п. 14.1). Средняя часть трубки (кишки) начинается глоткой и заканчивается в том месте, где ее энтодермальная слизистая оболочка контактирует с эктодермальной слизистой оболочкой задней кишки, или proctodeum.
14.3.1. РОТОВАЯ ПОЛОСТЬ
Рассмотрим вначале эволюцию ротовой полости и ее производных. У бесчерепных ротовая полость окружена предротовой воронкой
со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками - губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком.
Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое выполняет функцию твердого нёба.
Наружные ноздри большинства рыб не сообщаются с внешней средой и выполняют только обонятельную функцию. В группе так называемых хоановых рыб и земноводных появляются внутренние ноздри - хоаны, открывающиеся в ротовую полость непосредственно позади альвеолярной дуги верхней челюсти.
Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо, полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания (рис. 14.21, а - в).
До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба - порок развития, известный под названием волчья пасть (рис. 14.21, г, д). Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии - 0,4 случая на 1000 рождений.
Зубы позвоночных связаны по происхождению с плакоидной чешуей хрящевых рыб (см. п. 14.1), у которых наблюдается непрерывный переход от типичных чешуй к зубам в ротовой полости (рис. 14.22). Зубы первоначально расположены во много рядов и
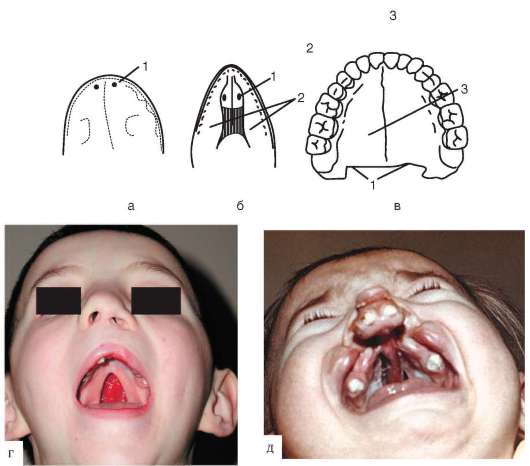
Рис. 14.21. Крыша ротовой полости у позвоночных: а - лягушка; б - ящерица; в - человек; г - неполная расщелина твердого нёба у человека; д - полная двусторонняя расщелина твердого нёба
покрывают всю слизистую оболочку ротовой полости, располагаясь у многих рыб даже на языке. У ряда земноводных зубы также расположены не только на альвеолярной дуге, но и на других костях, например на сошнике. У пресмыкающихся обнаруживается только один ряд зубов, причем так же, как у земноводных и рыб, дифферен-цировка их отсутствует. Такую зубную систему, в которой все зубы одинаковы, называют гомодонтной. У животных перечисленных классов зубы, как и плакоидные чешуи, могут многократно выпадать, сменяясь новыми поколениями. Многократную смену зубов называют полифиодонтизмом.
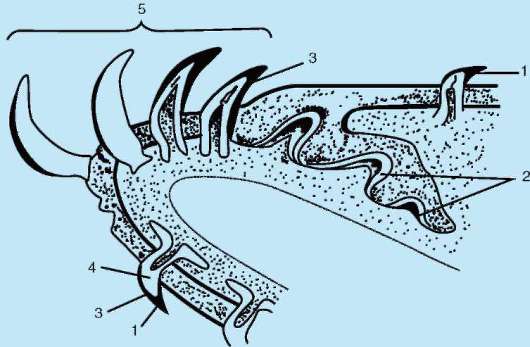
Рис. 14.22. Переход от плакоидной чешуи к зубам по краю ротового отверстия акулы: 1 - плакоидная чешуя; 2 - закладка новых зубов; 3 - эмаль; 4 - дентин; 5 - зубы
Зубы млекопитающих дифференцированы на резцы, клыки и коренные. Они выполняют различные функции. Такую зубную систему называют гетеродонтной. Если резцы, и в особенности клыки, млекопитающих еще весьма сходны с коническими зубами предков, то наибольшим эволюционным преобразованиям подверглись коренные зубы. Впервые они возникли у зверозубых ящеров начала мезозойской эры за счет расширения оснований зубов и появления дополнительных бугорков, а затем и увеличения жевательных поверхностей при их сглаживании. Общее число зубов у млекопитающих уменьшается; так, у высших приматов всего 32 зуба. У некоторых млекопитающих, например, неполнозубых, зубов существенно меньше. Зубы располагаются только на альвеолярных дугах челюстей, в ячейках. Основание зуба сужается, образуя корень.
Зубы человека по сравнению с другими приматами уменьшены в размерах, особенно клыки. Моляры имеют четырехбугорчатое строение. Зубная дуга округлой формы. В связи с дифференцировкой зубов увеличилась продолжительность их функционирования, в результате чего в онтогенезе сменяются только два их поколения: молочные и постоянные. Это явление называют дифиодонтизмом. В редких случаях и
у человека в пожилом возрасте может наблюдаться прорезывание отдельных зубов третьего поколения.
У человека возможны атавистические аномалии зубной системы, связанные с нарушениями как дифференцировки зубов, так и с их числом. Редкая аномалия - гомодонтная зубная система, в которой все зубы имеют коническую форму. Более часто встречается такая патология, как трехбугорчатое строение коренных зубов. Нередко встречается прорезывание сверхкомплектных зубов в ряду или за его пределами, иногда даже на твердом нёбе (рис. 14.23). Это свидетельствует о том, что у человека возможно образование большего числа зубных зачатков, чем 32, как это в норме встречается у низших млекопитающих и представителей более отдаленных классов позвоночных. Свидетельство тенденции к дальнейшему уменьшению числа зубов у человека - то, что нередко последние коренные зубы, так называемые «зубы мудрости», вообще не прорезываются, а если и прорезываются, то это происходит поздно - до 25 лет. Кроме того, эти зубы имеют явно рудиментарный характер, уменьшены в размерах и часто слабо дифференцированы.
На дне ротовой полости позвоночных располагается непарный выступ - язык, который у рыб представляет собой складку слизистой оболочки, лишенную мышц. Его движения осуществляются вместе с челюстями и подъязычной костью. У наземных позвоночных в языке располагаются мышцы, и они определяют его собственную подвижность. У пресмыкающихся и млекопитающих язык закладывается из трех зачатков: одного непарного и двух парных, лежащих кпереди от первого.
Рис. 14.23. Прорезывание сверхкомплектных зубов при клейдокраниальной дисплазии
Парные зачатки позже срастаются и дают начало телу языка. У большинства рептилий это срастание неполное, и язык раздвоен на конце. Весьма редким пороком развития языка у человека является раздвоенность его конца как результат несращения парных зачатков в эмбриогенезе.
Слюнные железы у рыб, заглатывающих пищу вместе с водой, отсутствуют и появляются только при переходе на сушу. Земноводные, обитающие на земле, имеют несколько слюнных желез, выделяющих только слизь. Слюна пресмыкающихся содержит уже и пищеварительные ферменты, а у некоторых змей - и токсины (см. п. 23.1). У млекопитающих слюнные железы многочисленны: это и мелкие - зубные, щечные, нёбные, язычные, гомологичные железам земноводных и пресмыкающихся, и крупные - подъязычные, подчелюстные и околоушные. Подъязычная и подчелюстная железы - результат дифферен-цировки подъязычной железы пресмыкающихся, околоушная - новое приобретение млекопитающих.
14.3.2. ГЛОТКА
Глотка - орган, выполняющий у всех хордовых две функции: дыхательную и пищеварительную. У ланцетника она пронизана множеством жаберных щелей (более 150 пар). У рыб 5-7 жаберных щелей закладываются как слепые парные выросты глотки - жаберные мешки. Навстречу им выпячиваются кожные покровы - жаберные карманы. В месте их соприкосновения происходит прорыв тканей эктодермы кожи и энтодермы глотки, и возникают сквозные жаберные щели.
У личинок земноводных в глотке образуется четыре пары жаберных щелей, а у пресмыкающихся прорыв их происходит только в эмбриональном развитии; вскоре после этого они зарастают. У млекопитающих в эмбриогенезе начинается закладка глоточных мешков и жаберных карманов, которые в норме никогда не прорываются и не образуют жаберных щелей (рис. 14.24).
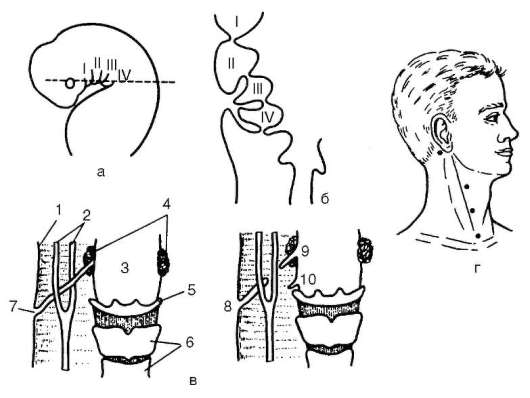
При нарушении эмбриогенеза во время закладки зачатков жаберных щелей они могут прорываться и даже сохраняться в постэмбриональном периоде у млекопитающих и человека. Эту аномалию называют латеральными свищами шеи. Они открываются на коже боковой поверхности шеи, а другим концом впадают в глотку. Если их полость имеет связь только с глоткой или открывается только на поверхности кожи, эти образования называются соответственно внутренними или
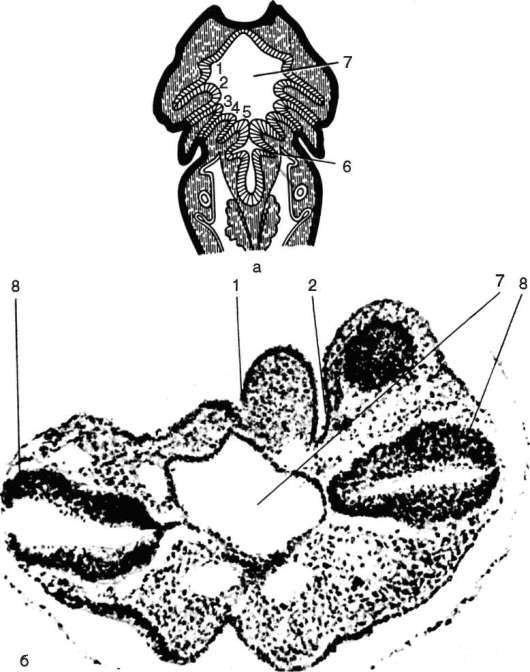
Рис. 14.24. Закладка жаберных щелей у позвоночных: а - фронтальный срез личинки тритона; б - срез через глотку зародыша мыши (головной конец зародыша сильно изогнут, поэтому на фотографии видны два среза одной нервной трубки); 1-5 - жаберные карманы и жаберные щели; 6 - зачаток легкого; 7 - полость глотки; 8 - нервная трубка
наружными шейными синусами. Чаще встречается феномен под названием латеральные кисты шеи. Эта аномалия связана с сохранением в постнатальном развитии эмбрионального материала глоточных жаберных мешков или эктодермальных жаберных карманов, не связанных ни с полостью глотки, ни с внешней средой. Участки этих образований могут заполняться жидкостью, увеличиваться в размерах и обладают склонностью к малигнизации (рис. 14.25). Необходимы своевременная диагностика этих образований и их хирургическое удаление.
Первая жаберная щель у всех наземных позвоночных, начиная с земноводных, в процессе формирования превращается в евстахиеву трубу, барабанную полость и наружный слуховой проход. Располагаясь топографически в зоне редукции первичных челюстей и подъязычной висцеральной дуги, преобразующихся в слуховые косточки, она меняет свои функции, включаясь в систему слухового анализатора (см.
п. 14.2.1).
Рис. 14-25. Развитие области глотки и ее аномалии у человека: а - зародыш в возрасте 5 нед; б - тот же зародыш (срез через область глотки по пунктирной линии, изображенной на рис. а); в - схема латеральных свищей шеи; г - проекции наиболее часто встречающихся латеральных шейных свищей; I-IV -жаберные дуги; 1 - кожа; 2 - сонная артерия; 3 - глотка; 4 - нёбные миндалины; 5 - подъязычная кость; 6 - гортань; 7 - сквозной шейный свищ; 8-10 - несквозные свищи (внутренние и наружный шейные синусы)
14.3.3. СРЕДНЯЯ И ЗАДНЯЯ КИШКА
Кишечная трубка претерпевает в ходе филогенеза следующие прогрессивные преобразования: увеличение общей длины, дифференци-ровку на отделы и образование крупных многоклеточных желез.
Если длина этого отдела пищеварительной трубки ланцетника составляет примерно 1/3 длины тела, то у млекопитающих возможно превышение ее длины по отношению к телу в 10 раз и более. Этим достигается удлинение времени контакта пищевых частиц с ферментами пищеварения и увеличение всасывающей поверхности. На органном уровне это сопровождается появлением складчатости слизистой оболочки кишки, возникновением ворсинок и крипт. Кишечник ланцетника абсолютно не дифференцирован. В средней его части расположен печеночный вырост - простое слепое выпячивание брюшной стенки кишки, выстланное железистыми клетками. Функционально этот вырост представляет собой зачаточную печень. Он является единственной многоклеточной железой пищеварительной системы ланцетника.
У рыб за глоткой следует короткий пищевод, затем желудок, слабо от него отграниченный. В кишечнике выделяются тонкий и толстый отделы, последний открывается анусом во внешнюю среду. В отличие от ланцетника у рыб хорошо развита печень, снабженная желчным пузырем. Поджелудочная железа у различных рыб построена по-разному. В некоторых случаях она представлена отдельными мелкими дольками в стенке кишечника, в мезентерии (брыжейке), рассеяна в ткани печени. Нередко уже у рыб она представляет собой компактный орган, включающий в себя как экзокринную часть, которая ответственна за синтез пищеварительных ферментов, так и эндокринную, которая выделяет гормоны, регулирующие углеводный обмен. Таким образом, у рыб наблюдаются все основные стадии эволюции сложной многоклеточной железы, структура и функции которой впоследствии принципиально практически не изменяются.
Единственное отличие пищеварительной трубки земноводных от трубки рыб - ее удлинение и впадение толстой кишки в клоаку.
Существенная особенность пищеварительной трубки пресмыкающихся - возникновение в ней слепой кишки. У большинства из них она зачаточна, но у некоторых развита весьма существенно. Слепая кишка - важное эволюционное приобретение пресмыкающихся, позволяющее им расширить рацион питания и использовать растительную пищу,
переваривание которой сложно и требует участия симбиотических простейших и бактерий. Слепая кишка особенно богата микрофлорой, под действием которой осуществляются процессы брожения, позволяющие наиболее полно использовать пластические вещества растительного происхождения.
У млекопитающих за счет дальнейшей дифференцировки пищеварительного тракта, и в частности больших размеров слепой кишки, резко увеличивается разнообразие используемых продуктов питания и возникает пищевая специализация на травоядность и плотоядность; всеядность встречается редко, в частности в отряде приматов.
Задняя кишка плацентарных млекопитающих дифференцирована, клоака отсутствует и прямая кишка заканчивается анусом.
В онтогенезе человека рекапитулируют основные стадии филогенеза кишечной трубки и ее производных. Из пороков развития, связанных с задержкой эмбриогенеза, известны гипоплазии всей пищеварительной системы, укорочение кишки и недоразвитие любых ее отделов, а также печени и поджелудочной железы. В зависимости от степени недоразвития тяжесть аномалии может быть как незначительной, так и несовместимой с жизнью.
Клинический интерес представляет гетеротопия тканей поджелудочной железы в стенке тонкого кишечника или желудка. Иногда гетеротопированные фрагменты могут симулировать опухолевую трансформацию слизистой оболочки. По данным патологоанатомов, частота этой аномалии - от 0,6 до 5%. Механизм гетеротопии - нарушение клеточной миграции зачатков железы из стенки кишечной трубки.
Таков же механизм формирования другой редкой аномалии - гете-ротопии печеночной ткани в стенку желудка или кишечника.
С недоразвитием и задержкой дифференцировки связана и такая аномалия, как персистирование клоаки, при которой мочеполовые пути и прямая кишка объединены. При нормальном развитии после 8-й недели эмбриогенеза клоака должна полностью дифференцироваться на прямую кишку, мочевыделительные и половые протоки.
14.3.4. ОРГАНЫ ДЫХАНИЯ
Жабры - наиболее ранние специализированные органы дыхания, появляющиеся среди хордовых впервые у рыб. Они представляют собой тонкие складки слизистой оболочки глотки, лежащие на жаберных
дугах, снабжаемые венозной кровью через жаберные артерии и распадающиеся здесь на капилляры.
Позади последней жаберной дуги у кистеперых рыб за счет выпячивания вентральной стороны глотки формируется парное образование - плавательный пузырь, который выполняет в первую очередь гидростатические функции, уравновешивая тело рыбы в толще воды. Связь между ним и глоткой, имеющаяся в эмбриогенезе, не прерывается и позже, поэтому воздух при попадании в глотку может беспрепятственно проходить в плавательный пузырь. Эта морфофизиологическая особенность при изменении условий существования кистеперых рыб стала предпосылкой преобразования плавательного пузыря в легкие (рис. 14.26).
У земноводных, связанных по происхождению с кистеперы-ми рыбами, в личиночном состоянии функционируют жабры, а во взрослом - органами дыхания становятся легкие. В связи с отсутствием грудной клетки и диафрагмы воздух в них попадает из ротовой полости за счет глотательных движений, осуществляющихся подбородочно-подъязычной мышцей. Материал жаберных дуг, следующих за подъязычной дугой, частично редуцируясь, входит в со-
Рис. 14.26. Плавательный пузырь рыб (а - костной; б - кистеперой) и развитие легких у человека (в - ранние стадии): 1 - средняя кишка; 2 -плавательный пузырь; 3 - глотка; 4 - развивающиеся легкие
став хрящей гортани, которая, появляясь впервые у земноводных, является первым органом, относящимся к нижним дыхательным путям. Легкие начинаются непосредственно от гортани. Они крупноячеисты и имеют относительно малую дыхательную поверхность, в связи с чем газообмен в большей степени осуществляется через кожные покровы (см. п. 14.1).
Пресмыкающиеся, вышедшие на сушу окончательно, имеют как верхние дыхательные пути (не полностью отграниченную от ротовой полости полость носа), так и нижние - гортань, трахею и бронхи. Их легкие мелкоячеисты, содержат многочисленные внутренние перегородки и обладают большой дыхательной поверхностью. Впервые появляется диафрагма, которая в дыхании принимает скорее пассивное участие, так как она либо лишена мышечных элементов, либо разделяет грудную и брюшную полости не полностью. Механизм дыхания основан на сокращении межреберных мышц, приводящих в движение грудную клетку.
У млекопитающих дыхательные пути, как и у предков, выстланы мерцательным эпителием. Они полностью отделены от пищеварительной системы и только перекрещиваются с ней в глотке. Бронхи многократно разветвляются, вплоть до бронхиол, ведущих в альвеолы - легочные пузырьки, имеющие в сумме огромную площадь поверхности (у человека до 90 м2). Основной мышцей, изменяющей объем грудной полости, становится диафрагма.
В эмбриогенезе человека отражается первоначальное единство пищеварительной и дыхательной систем. На этом основано формирование большой группы врожденных пороков развития пищевода и трахеи типа эзофаготрахеальных свищей (рис. 14.27). Нарушением альвеолярной дифференцировки легочной ткани могут быть объяснены дизонтогенетические бронхолегочные кисты - округлые полости в легких, отграниченные от окружающих тканей примитивно построенной стенкой недифференцированного бронха, а также кистозная гипоплазия (недоразвитие) легких (рис. 14.28). При этой патологии недоразвитыми оказываются целые доли легкого, которые представляют собой многочисленные полости, связанные с крупными бронхами и имеющие малую площадь поверхности. Газообмен в таких участках легких резко ослаблен. Широко известны также гипоплазии диафрагмы от небольших дефектов в ее куполе до полной аплазии. Последняя аномалия несовместима с жизнью и встречается обычно вместе с другими множественными пороками развития.
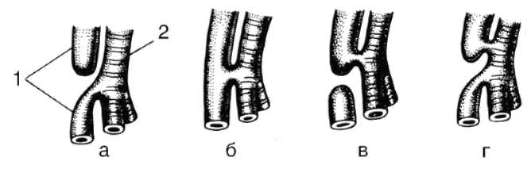
Рис. 14.27. Различные формы эзофаготрахеальных свищей (а - г): 1 - пищевод; 2 - трахея
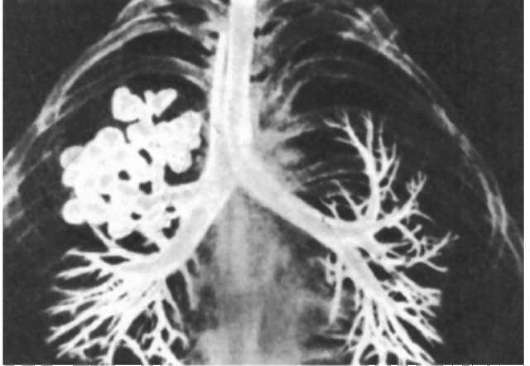
Рис. 14.28. Кистозная гипоплазия правого легкого. Контрастная рентгенограмма
14.4. КРОВЕНОСНАЯ СИСТЕМА
Необходимое условие существования высокоорганизованных крупных многоклеточных организмов - наличие жидкой подвижной внутренней среды, которая обеспечивает интеграцию организма в целостную систему, выполняя транспортные функции. Эти функции являются основными для кровеносной системы. Конкретная функция кровеносной системы зависит от того, что она транспортирует: питательные вещества, кислород, углекислый газ, другие продукты диссимиляции или гормоны. Кровеносная система всех хордовых замкнутая и состоит из двух основных артериальных сосудов: брюшной и спинной аорт. По брюшной аорте венозная кровь продвигается кпереди, обогащается кислородом в органах дыхания, а по спинной - кзади. Из спинной аор-
ты кровь через систему капилляров возвращается по венам в брюшную аорту. Брюшная аорта или ее часть, периодически сокращаясь, проталкивает кровь по сосудам (рис. 14.29).
14.4.1. ЭВОЛЮЦИЯ ОБЩЕГО ПЛАНА СТРОЕНИЯ КРОВЕНОСНОЙ СИСТЕМЫ ХОРДОВЫХ
У ланцетника кровеносная система наиболее проста (рис. 14.30, а). Круг кровообращения один. По брюшной аорте венозная кровь поступает в приносящие жаберные артерии, число которых соответствует числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
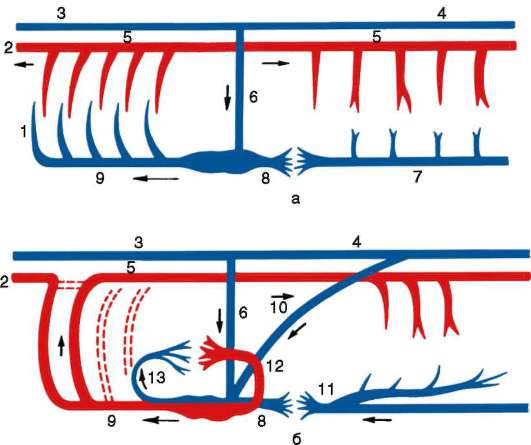
Рис. 14.29. Строение кровеносной системы водных (а) и наземных (б) позвоночных: 1 - жаберные артерии; 2 - сонная артерия; 3 - передняя кардинальная вена; 4 - задняя кардинальная вена; 5 - спинная аорта; 6 - кювьеров проток; 7 - подкишечная вена; 8 - печеночная вена; 9 - брюшная аорта; 10 - задняя (нижняя) полая вена; 11 - воротная вена печени; 12 - легочная вена; 13 - легочная артерия; стрелками указано направление движения крови
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к переднему концу тела, так и назад. Передние ветви этих двух сосудов называются сонными артериями. На уровне заднего конца глотки задние ветви, срастаясь, образуют спинную аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и распадающиеся на капилляры.
После тканевого газообмена кровь поступает в парные передние или задние кардинальные вены, расположенные симметрично. Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток. Оба кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной системы венозная кровь оттекает по воротной вене печени в печеночный вырост, где формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд - печеночную вену, по которой кровь поступает в брюшную аорту.
Таким образом, несмотря на простоту кровеносной системы в целом, уже у ланцетника имеются основные магистральные артерии, характерные для позвоночных, в том числе для человека: это брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень легочной артерии; спинная аорта, становящаяся позже собственно аортой, и сонные артерии. Основные вены, имеющиеся у ланцетника, также сохраняются у более высокоорганизованных животных. Так, передние кардинальные вены станут позже яремными венами, правый кювьеров проток преобразуется в верхнюю полую вену, а левый, сильно редуцировавшись, - в коронарный синус сердца. Для того чтобы понять, как это происходит, необходимо сопоставить кровеносные системы всех классов позвоночных животных.
Активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризации их артериальных жаберных дуг, в конечном счете, до четырех пар в них отмечается высокая степень дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное сердце, состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным аппаратом. В остальном кровеносная система рыб соответствует строению ее у ланцетника (рис. 14.30, б).
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга кровообращения. Соответственно
этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное, но, как и у рыб, от правой половины единственного желудочка начинается единственный сосуд - артериальный конус, разветвляющийся последовательно на три пары сосудов: кожно-легочные артерии, дуги аорты и сонные артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое - малого с артериальной кровью. При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту, снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и, находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными венами, а кювьеровы потоки, в которые они впадают вместе с подключичными венами, - передними полыми венами (рис. 14.30, в).
В кровеносной системе пресмыкающихся возникают следующие прогрессивные изменения: в желудочке их сердца имеется неполная перегородка, затрудняющая смешение крови, поступающей из правого и левого предсердий; от сердца отходит не один, а три сосуда, образовавшихся в результате разделения артериального ствола. Из левой половины желудочка начинается правая дуга аорты, несущая артериальную кровь, а
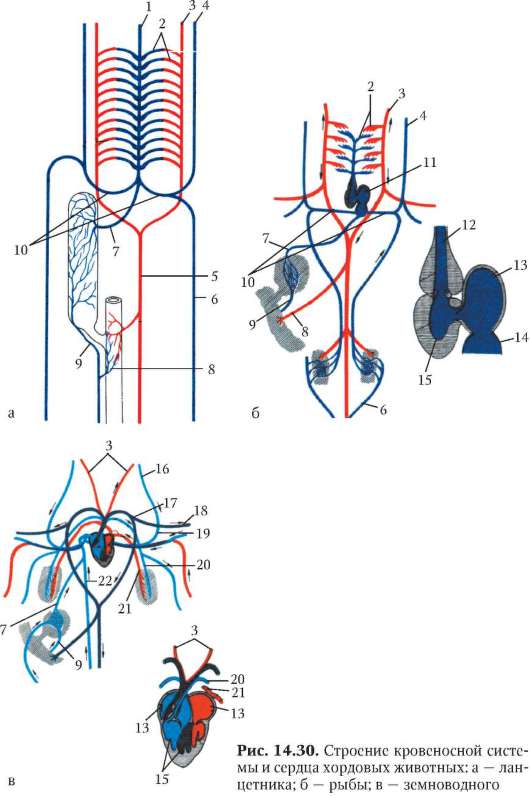
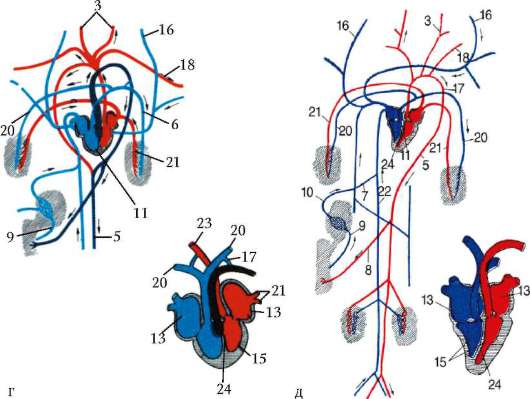
Рис. 14.30. Продолжение. г - пресмыкающегося; д - млекопитающего; 1 - брюшная аорта; 2 - жаберные артерии; 3 - сонная артерия; 4 - передняя кардинальная вена; 5 - спинная аорта; 6 - задняя кардинальная вена; 7 - печеночная вена; 8 - чревная артерия; 9 - воротная вена печени; 10 - кювьеровы протоки; 11 - сердце; 12 - артериальный конус; 13 - предсердие; 14 - венозный синус; 15 - желудочек; 16 - яремная вена; 17 - левая дуга аорты; 18 - подключичная артерия; 19 - передняя полая вена; 20 - легочная артерия; 21 - легочная вена; 22 - задняя полая вена; 23 - правая дуга аорты; 24 - межжелудочковая перегородка
из правой - легочная артерия с венозной кровью. Из середины желудочка, в области неполной перегородки, начинается левая дуга аорты со смешанной кровью. Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи и пищевода в спинную аорту, кровь в которой смешанная, но более богата кислородом, чем у земноводных, в связи с тем, что до слияния сосудов только по левой дуге течет смешанная кровь. Кроме того, сонные и подключичные артерии с обеих сторон берут начало от правой дуги аорты, в результате чего артериальной кровью снабжается не только голова, но и передние конечности. В связи с появлением шеи сердце располагается еще более каудально, чем у земноводных. Венозная система пресмыкающихся принципиально не отличается от системы вен земноводных (рис. 14.30, г).
Прогрессивные изменения кровеносной системы млекопитающих сводятся к полному разделению венозного и артериального кровотоков. Это достигается, во-первых, завершенной четырехкамерностью сердца и, во-вторых, редукцией правой дуги аорты и сохранением только левой, начинающейся от левого желудочка. В результате все органы млекопитающих снабжаются артериальной кровью. В венах большого круга кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь только от миокарда, а непарная и полунепарная вены - рудименты задних кардинальных вен, имеют существенное значение в основном в случаях формирования обходных путей венозного оттока через кава-кавальные анастомозы, формируемые ими (рис. 14.30, д).
В эмбриональном развитии млекопитающих, в том числе человека, рекапитулируют закладки сердца и основных кровеносных сосудов предковых классов.
Сердце закладывается в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехка-мерным сердцем (рис. 14.31). Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала, чем у рептилий. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка
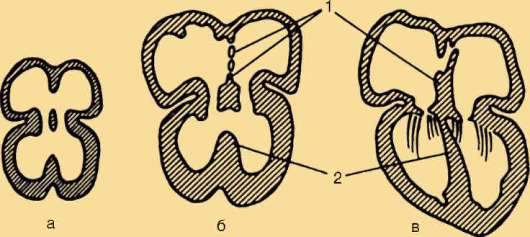
Рис. 14.31. Развитие перегородок сердца у зародыша человека в возрасте 30 сут (а), 35 сут (б) и 2 мес (в): 1 - межпредсердная; 2- межжелудочковая
является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.
Интересно, что место закладки и положение сердца в филогенетическом ряду позвоночных полностью рекапитулируют у млекопитающих, в том числе человека. Так, закладка сердца у человека осуществляется на 20-е сутки эмбриогенеза, как у всех позвоночных, в задней части головы. Позже за счет изменения пропорций тела, появления шейной области, смещения легких в грудную полость осуществляется и перемещение сердца в переднее средостение.
Нарушения развития сердца могут выражаться как в возникновении аномалий строения, так и места его положения. Возможно сохранение к моменту рождения двухкамерного сердца. Этот порок совершенно не совместим с жизнью.
Чаще встречаются дефекты межпредсердной перегородки (1 случай на 1000 рождений), межжелудочковой перегородки (2,5-5 случаев на 1000 рождений), вплоть до трехкамерного сердца с одним общим желудочком (рис. 14.31, в). Известен и такой порок, как шейная эктопия сердца, при которой оно находится в шейной области. Этот порок связывают с задержкой сердца в области его первоначальной закладки. В большинстве случаев при шейной эктопии сердца его дифференци-ровка сильно нарушена. При этом ребенок обычно погибает сразу после рождения. Перечисленные пороки сердца наиболее часто встречаются не в изолированном виде, а в комплексе с другими аномалиями сердца, сосудов, а нередко и других органов. Это свидетельствует о том, что в морфогенезе сердца большое значение имеют онтогенетические корреляции (см. п. 13.4). Состояние больных при таких пороках зависит от того, насколько сильно нарушается гемодинамика и осуществляется смешение крови в кровеносном русле.
14.4.2. ФИЛОГЕНЕЗ АРТЕРИАЛЬНЫХ ЖАБЕРНЫХ ДУГ
В связи с тем что основные артериальные сосуды у млекопитающих формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных (рис. 14.32). В эмбриогенезе абсолютного большинства позвоночных закладывается шесть
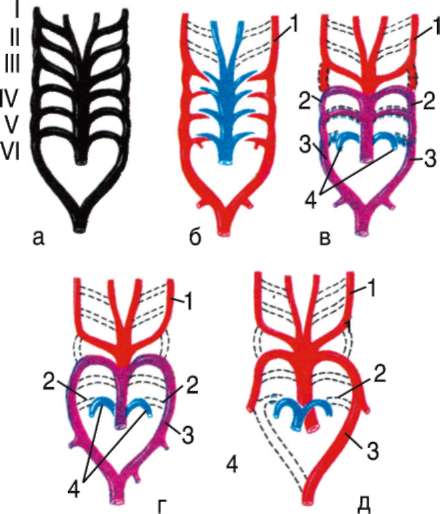
Рис. 14.32. Эволюция артериальных жаберных дуг позвоночных: а - закладка в эмбриогенезе; б - жаберные дуги рыб; в - жаберные дуги хвостатых земноводных; г- жаберные дуги пресмыкающихся; д - жаберные дуги млекопитающих; I-VI - жаберные дуги; 1 - сонные артерии; 2 - боталлов проток; 3 - артериальные дуги большого круга кровообращения; 4 - легочные артерии
пар артериальных жаберных дуг, соответствующих шести парам висцеральных дуг черепа. Поскольку две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты - основными сосудами большого круга кровообращения.
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент - плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных,
кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытий детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты 4-й пары. При этом обе дуги, так же, как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытий) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5-1,2 случая на 1000 новорожденных) - персистирование артериального, или боталлова, протока (рис. 14.33, а), представляющего собой часть корня спинной аорты между 4-й и 6-й парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития - персистирование первичного эмбрионального ствола, в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке (рис. 14.33, б). Такой порок обычно приводит к смерти ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов - от-хождение аорты от правого желудочка, а легочного ствола - от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен. Если обе они впадают в правое предсердие, аномалия клинически не проявляется.
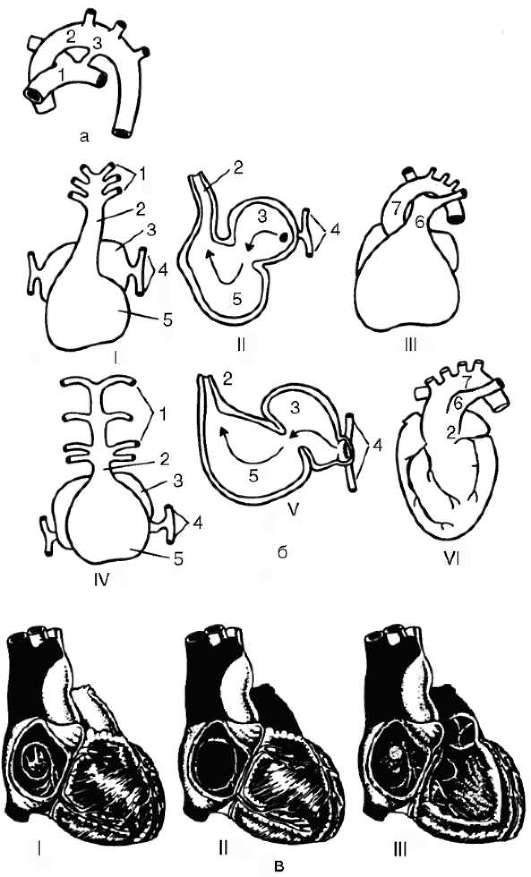
Рис. 14.33. Врожденные пороки развития сердца и сосудов у человека: а - пер-систирование артериального (боталлова) протока у человека: 1 - легочный ствол; 2 - левая дуга аорты; 3 - артериальный (боталлов) проток; б - онто-филогенетические основы формирования пороков развития сердца и отходящих от него сосудов у человека; I - сердце зародыша человека в возрасте 5 нед; II - то же, в разрезе; III - разделение артериального конуса у зародыша человека в возрасте 6 нед; IV - сердце рыбы; V - то же, в разрезе; VI - персистиро-вание неразделенного артериального конуса у человека: 1 - артериальные жаберные дуги; 2 - артериальный конус; 3 - предсердие; 4 - кардинальные вены; 5 - желудочек; 6 - общая легочная артерия; 7 - левая дуга аорты; в - аномалии межпредсердной (I, II) и межжелудочковой (III) перегородок
При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия - неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, рудименты задних кардинальных вен.
В случае постепенного сужения просвета нижней полой вены за счет различных причин, в частности, сдавления ее растущей опухолью, венозный отток из нижней части тела осуществляется через непарную и полунепарную вены. Это пример рудиментарных структур, сохраняющихся в современном организме не только потому, что их формирование заложено в генетическую программу развития, но и потому, что в критических ситуациях они могут быть полезны для выживания организма.
14.5. МОЧЕПОЛОВАЯ СИСТЕМА
Выделительная и половая системы в целом выполняют разные функции. Однако их рассматривают в едином комплексе в связи с единством эмбрионального развития и первичной функциональной связью со вторичной полостью тела - целомом. В эмбриогенезе закладка мочеполовой системы - нефрогонотом - формируется в области ножки сомита в непосредственном контакте с целомом. Половые железы у всех хордовых животных располагаются в целоме. Продукты диссимиляции у всех целомических животных, в том числе и низших хордовых, также поступают в целом. Наиболее простой путь выведения во внешнюю среду как половых клеток, так и продуктов диссимиляции - через общий канал, начинающийся воронкой в целоме и заканчивающийся выделительной порой на покровах. Таковы взаимоотношения половых желез и выделительных каналов у многих беспозвоночных и, вероятно, у древних предков хордовых (рис. 14.34).
14.5.1. ЭВОЛЮЦИЯ ПОЧКИ
Органами выделения позвоночных являются почки - парные компактные органы, структурно-функциональная единица которых
представлена нефроном. В наиболее примитивном виде это - воронка, открывающаяся в целом и соединенная с выделительным канальцем, который впадает в общий выводной проток - мочеточник. В филогенезе позвоночных почка прошла три этапа эволюции: предпочка - головная, или пронефрос; первичная почка - туловищная, или мезонефрос, и вторичная почка - тазовая, или метанефрос.
Предпочка полностью развивается и функционирует как самостоятельный орган у личинок рыб и земноводных. Она находится на переднем конце тела, состоит из 2-12 нефронов, воронки которых открыты в целом, а выводные канальцы впадают в пронефрический канал, который соединен с клоакой. Предпочка имеет сегментарное строение. Продукты диссимиляции фильтруются в целом из кровеносных сосудов, которые поблизости от нефронов формируют клубочки (рис. 14.35, а).
У взрослых рыб и земноводных кзади от предпочек, в туловищных сегментах тела, формируются первичные почки, содержащие до нескольких сотен нефронов. В ходе онтогенеза число нефронов увеличивается за счет их почкования друг от друга с последующей диффе-ренцировкой. Они вступают в связь с кровеносной системой, формируя капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, в которых располагаются сосудистые клубочки, благодаря чему продукты диссимиляции могут поступать из крови непосредственно в нефрон. Некоторые нефроны первичной почки сохраняют связь с целомом через воронки, другие - утрачивают ее (рис. 14.35, б, в).
Выделительные канальцы удлиняются, и в них осуществляется обратное всасывание в кровь воды, глюкозы и других веществ, в связи с
чем концентрация продуктов диссимиляции в моче повышается. Однако воды с мочой теряется много, поэтому животные, обладающие такой почкой, могут обитать только в водной или влажной среде. Первичная почка сохраняет признаки метамерного строения.
У пресмыкающихся и млекопитающих возникают вторичные почки. Они закладываются в тазовом отделе тела и содержат сотни тысяч нефронов наиболее совершенного строения. У новорожденного ребенка в почке их насчитывается около 1 млн. Они образуются за счет многократного ветвления развивающихся нефронов. Нефроны не имеют воронки и, таким образом, теряют полностью связь с целомом. Каналец нефрона удлиняется, теснее контактирует с кровеносной системой, а у млекопитающих дифференцируется на проксимальный и дистальный участки, между которыми появляется еще и так называемая петля Ген-ле (рис. 14.35, г).
Такое строение нефрона обеспечивает не только полноценную фильтрацию плазмы крови в капсуле, но и, что более важно, эффективное обратное всасывание в кровь воды, глюкозы, гормонов, солей и других необходимых организму веществ. В результате концентрация продуктов диссимиляции в моче, выделяемой вторичными почками, велика, а само ее количество - мало. У человека, например, за сутки в капсулах нефронов обеих почек фильтруется около 150 л плазмы крови, а мочи выделяется около 2 л. Это позволяет животным, обладающим вторичными почками, быть более независимыми от водной среды и заселять засушливые области. У пресмыкающихся вторичные почки на протяжении всей жизни сохраняются на месте их первоначальной закладки - в тазовой области. В них прослеживаются черты первичного метамер-ного строения.
Почки млекопитающих располагаются в поясничной области, и у большинства из них внешняя сегментация не выражена. В онтогенезе человека обнаруживается выраженная рекапитуляция в развитии почки: закладывание вначале про-, затем мезо-, а позже метанефроса. Последний развивается в тазовой области, а затем за счет различий в скоростях роста позвоночника, таза и органов брюшной полости перемещается в поясничную область. У пятинедельного зародыша можно обнаружить сосуществование предпочки, первичной, а также зачатков вторичной почки (рис. 14.36).
На начальных этапах развития почка человека сегментирована. Позже ее поверхность сглаживается и метамерность сохраняется лишь во внутреннем строении в виде почечных пирамид. Пороки развития по-
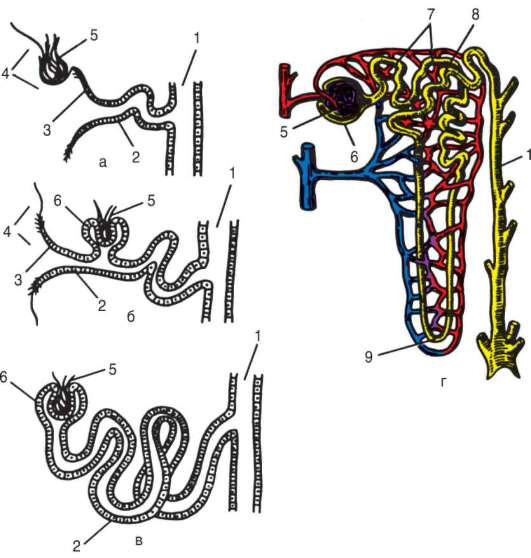
Рис. 14.35. Эволюция нефрона: а - предпочка; б, в - первичная почка; г - вторичная почка; 1 - собирательная трубочка; 2 - выделительный каналец; 3 - нефростом; 4 - целом; 5 - капиллярный клубочек; 6 - капсула; 7, 8 - извитой каналец; 9 - петля нефрона
чек у человека, основанные на их филогенезе, многообразны. Сохранение мезонефроса и одностороннее отсутствие вторичной почки описаны пока только у мышей, хотя в принципе такая аномалия возможна и у человека. Относительно часто встречается сегментированная вторичная почка, имеющая один или даже несколько мочеточников; возможно и полное ее удвоение. Часто наблюдается тазовое расположение почки, связанное с нарушением ее перемещения на 2-4-м месяцах зародышевого развития (рис. 14.37).
Рис. 14.36. Пятинедельный зародыш человека с тремя поколениями почек: 1 - предпочка; 2 - первичная почка; 3 - вторичная почка
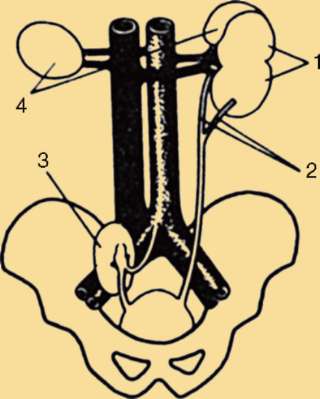
Рис. 14.37. Онтофилогенетически обусловленные пороки развития почек: 1 - удвоение почки; 2 - удвоенный мочеточник; 3 - тазовая эктопия почки; 4 - надпочечники
14.5.2. ЭВОЛЮЦИЯ ПОЛОВЫХ ЖЕЛЕЗ
Половые железы у всех позвоночных развиваются в виде парных складок части нефрогонотома в области ножки сомита. Половые складки вдаются в полость тела и оказываются подвешенными на брыжейке. Первичные половые клетки (см. также п. 6.5.3) обособляются у зародышей очень рано - уже на стадии гаструляции. Вначале они обнаруживаются в составе презумптивной эктодермы головного конца эмбриона, затем попадают в энтодерму, откуда активно перемещаются в половые складки. Здесь дифференцирующийся эпителий половой железы, включающий в себя первичные половые клетки, объединяется с соединительнотканной стромой в виде шнуров. Такая гонада индифферентна в половом отношении и может развиваться в дальнейшем как в семенник, так и в яичник в зависимости от генетических и эпигенетических факторов дифференцировки пола (см. также п. 4.3.7).
У хрящевых рыб строение половых желез самок и самцов в значительной степени сходно. У всех остальных позвоночных яичник всегда имеет фолликулярное строение, т.е. содержит пузырьки - фолликулы, в каждом из которых находится одна будущая яйцеклетка. При созрева-
нии яйцеклетки стенка фолликула разрывается, и она поступает вначале в брюшную полость, а затем в яйцевод. Семенники содержат семенные трубочки, которые соединены с семявыносящими каналами, по которым зрелые сперматозоиды выходят во внешнюю среду.
Индифферентность развивающейся половой железы позвоночных называют первичным гермафродитизмом. Он эволюционно связан, вероятно, с гермафродитизмом древних предков позвоночных. Доказательством этого может служить наличие у наиболее примитивных современных позвоночных - круглоротых - таких половых желез, передняя часть которых является яичником, а задняя - семенником. У самок некоторых земноводных на протяжении всей жизни сохраняется рудиментарный участок индифферентной половой железы. Зародыш человека до определенного возраста имеет недифференцированные половые железы, которые в зависимости от различных факторов (см. п. 6.1.2) становятся либо семенниками, либо яичниками. Нарушение их дифференцировки может привести к возникновению ovotestis, в котором сочетаются элементы семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2% мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма - неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте.
14.5.3. ЭВОЛЮЦИЯ МОЧЕПОЛОВЫХ ПРОТОКОВ
В эмбриогенезе всех позвоночных при развитии предпочки вдоль тела, от головного конца к клоаке, закладывается канал, по которому продукты диссимиляции из нефронов поступают во внешнюю среду. Это пронефрический канал. При развитии первичной почки этот ка-
нал либо расщепляется на два канала, идущих параллельно, либо второй канал образуется в продольном утолщении стенки первого. Один из них - вольфов - вступает в связь с нефронами первичной почки. Другой - мюллеров - срастается передним концом с одним из не-фронов предпочки и образует яйцевод, открывающийся передним концом в целом широкой воронкой, а задним - впадающий в клоаку.
Вне зависимости от пола у всех позвоночных обязательно формируются как вольфов, так и мюллеров каналы, однако судьба их различна как у разных полов, так и у представителей разных классов. У самок рыб и земноводных вольфов канал всегда выполняет функцию мочеточника, а мюллеров - яйцевода. У самцов мюллеров канал редуцируется и обе функции - половую и выделительную - выполняет вольфов канал. Семенные канальцы при этом впадают в почку, а сперматозоиды при оплодотворении поступают в воду вместе с мочой.
У пресмыкающихся и млекопитающих большая часть вольфова канала не принимает участия в выведении мочи и только его наиболее каудаль-ная часть в области впадения в клоаку образует выпячивание, становящееся мочеточником вторичной почки. Сам же вольфов канал у самцов выполняет функцию семяизвергательного канала. Мюллеров канал у них подвергается редукции. У самок вольфов канал редуцируется (за исключением его каудальной части, формирующей мочеточник), а мюллеров - становится яйцеводом (рис. 14.38). У плацентарных млекопитающих мюллеров канал дифференцируется на собственно яйцевод, матку и влагалище. Будучи парным образованием, как и все элементы половой системы, мюллеров канал сохраняет парность строения у яйцекладущих и частично у сумчатых млекопитающих, у которых имеется два влагалища, две матки и два яйцевода. В дальнейшей эволюции происходит срастание мюллеровых каналов с образованием одного влагалища и матки, которая может быть либо двойной, как у многих грызунов, либо двураздельной, как у хищных; либо двурогой, как у насекомоядных и китообразных, либо простой, как у приматов, в том числе человека.
Соответственно дифференцировкам мюллерова канала самок у самцов пресмыкающихся и млекопитающих развиваются копулятивные органы. У большинства пресмыкающихся, а также у сумчатых млекопитающих они парные. У плацентарных с одним влагалищем копуля-тивный орган непарный, но в его развитии обнаруживается срастание парных зачатков.
В эмбриогенезе человека закладываются парные вольфовы и мюл-леровы каналы. Позже в зависимости от пола происходит их редукция.
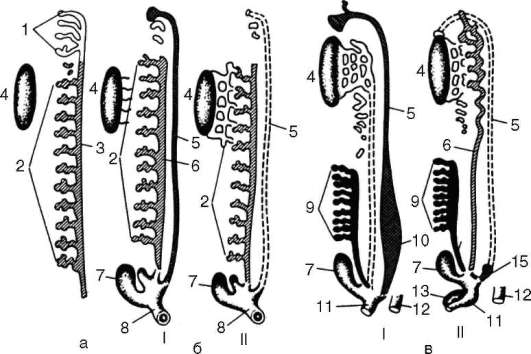
Рис. 14.38. Эволюция почки и мочеполовых каналов: а - нейтральное зародышевое состояние; б - анамнии; в - амниоты; I - самки; II - самцы; 1 - пред-почка; 2 - первичная почка; 3 - канал предпочки; 4 - половая железа; 5 - мюллеров канал; 6 - вольфов канал; 7 - мочевой пузырь; 8 - клоака; 9 - вторичная почка; 10 - матка; 11 - мочеполовой синус; 12 - задняя кишка; 13 - половой член; 14 - мочеточник вторичной почки; 15 - мужская «маточка»
Рудимент мюллерова канала у мужчин располагается в предстательной железе и называется мужской маточкой - utriculus masculinus. Канальцы передней части первичной почки у них вступают в связь с семенниками и преобразуются в придаток семенника - эпидидимис. У плодов женского пола возможно нарушение редукции вольфовых каналов, которые располагаются по бокам от влагалища. Эта аномалия опасна возможностью образования кист и злокачественного перерождения. Распространенными пороками развития являются также различные формы удвоения матки (1 случай на 1000 перинатальных вскрытий). Они развиваются как результат нарушения срастания мюл-леровых каналов. В случае полного срастания мюллеровых каналов, но при нарушении редукции клеточного материала их стенок внутри матки возникает аномалия, называемая разделенная матка. Нарушение срастания парных зачатков полового члена в эмбриогенезе человека может привести к формированию такого порока развития, как его удвоение (рис. 14.39). Основной клеточный механизм возникновения описанных
Рис. 14.39. Аномалии матки и полового члена у человека: а-г - различные формы удвоения матки и влагалища; д - разделенная матка; е - удвоение полового члена
пороков развития - нарушение избирательной клеточной адгезии и дифференцировки в процессе морфогенеза.
14.6. ИНТЕГРИРУЮЩИЕ СИСТЕМЫ
У большинства многоклеточных животных, в том числе и у позвоночных, существует две системы регуляции функций, интегрирующие организм в целостную систему, - нервная и эндокринная. Нервная система осуществляет быстрое реагирование организма на изменения условий среды посредством рефлексов. Эндокринная система с помощью гормонов обеспечивает более медленные, но и более стойкие приспособительные реакции. В связи с тесным взаимодействием обеих регуляторных систем в их филогенезе проявляются тесные динамические координации, а в онтогенезе - эргонтические корреляции.
14.6.1. ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы. Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными.
Действительно, у наиболее примитивного представителя хордовых - ланцетника - центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования - глазки Гессе. Кроме того, основные дистантные органы чувств - зрения, обоняния и слуха - образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки.
В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри - невроцелем. У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок (рис. 14.40). Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.
У всех позвоночных центральная нервная система - производное нервной трубки, передний конец которой становится головным мозгом, а задний - спинным. Образование головного мозга называют кефализацией. Она связана с усилением двигательной активности
позвоночных и необходимостью постоянного анализа раздражений, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных - обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координаций между обонятельными рецепторами и передним мозгом, зрительными - и средним, слуховыми - и задним. Видимо, поэтому головной мозг
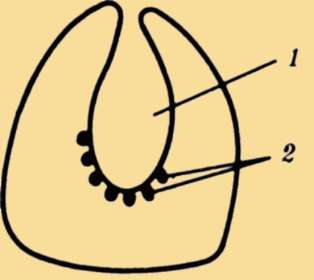
Рис. 14.40. Нервная трубка ланцетника: 1 - невроцель; 2 - глазки Гессе
всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей - переднего, среднего и заднего - и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей.
Головной мозг современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге это спинномозговой канал, а в головном - желудочки мозга. Ткань мозга состоит из серого вещества (скопления нервных клеток) и белого (отростков нервных клеток).
Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, центральный орган эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтио-псидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр
зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг - наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегра-тивного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней - archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих - маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой - neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерно также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних - со слуховым. Очень хорошо развит мозжечок (рис. 14.41).
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференциров-ка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных, происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити - filum terminate. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост - cauda equina.
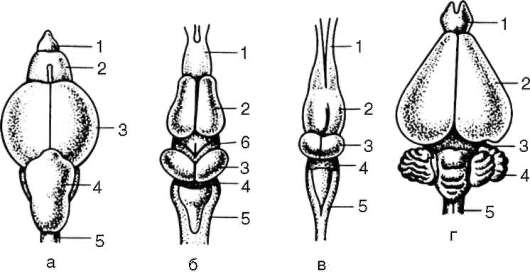
Рис. 14.41. Эволюция головного мозга позвоночных: а - рыба; б - земноводное; в - пресмыкающееся; г - млекопитающее; 1 - обонятельные доли; 2 - передний мозг; 3 - средний мозг; 4 - мозжечок; 5 - продолговатый мозг; 6 - промежуточный мозг
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.42): передний (I), средний (II) и задний (III). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (1) и промежуточный (2) мозг - telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг - mesencephalon, a задний - в задний (4) мозг - metencephalon - и продолговатый (5) мозг - medulla oblongata.
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека - уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут особенно быстро. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том
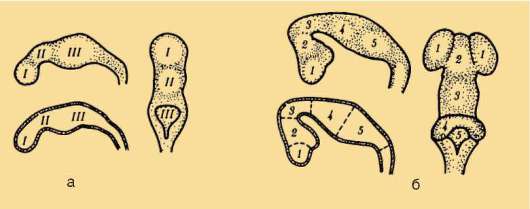
Рис. 14.42. Мозговые пузыри в эмбриогенезе мозга человека: а - стадия трех мозговых пузырей; б - стадия пяти мозговых пузырей (остальные пояснения в тексте)
числе центров письменной и устной речи и других, характерных только для человека (см. п. 15.3).
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз, или плати-неврию, - отсутствие замыкания нервной трубки (рис. 14.43). Такая аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга - прозэнцефалия - выражается в нарушении морфогенеза мозга, при котором полушария оказываются неразделенными, а кора - недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью и часто встречается у мертворожденных при
различных хромосомных и генных синдромах. Клеточные механизмы формирования этого порока - комплексные нарушения как клеточной пролиферации, так и миграции, дифференцировки и формирования межклеточных контактов, а генетические - в первую очередь нарушение генного баланса.
Нарушения дифференцировки коры - агирия (отсутствие извилин) и олигогирия с пахигирией
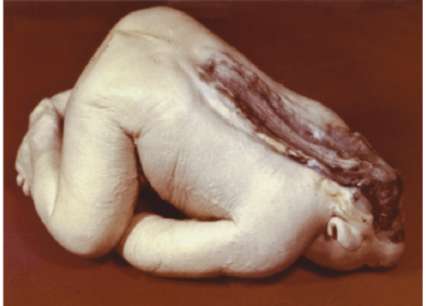
Рис. 14.43. Рахисхиз (платиневрия)
(малое количество утолщенных извилин) - сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей при этом умирают в течение первого года жизни.
14.6.2. ЭНДОКРИННАЯ СИСТЕМА
Эндокринная система наряду с нервной является ведущим аппаратом интеграции многоклеточного организма, обеспечивая гуморальную регуляцию функций органов. Эта регуляция осуществляется гормонами - биологически активными веществами разной химической природы, выделяемыми железами внутренней секреции. Действие гормонов строго специфично: разные гормоны действуют на разные органы, вызывая определенные изменения их функционирования. Железы внутренней секреции не имеют протоков и выделяют гормоны непосредственно в кровь, что облегчает их транспорт к органам-мишеням. Клетки органов-мишеней на своих мембранах имеют специфические рецепторы, с которыми связываются гормоны, вызывая определенные изменения их метаболизма. Гуморальная регуляция эволюционно возникла значительно раньше нервной, поскольку она более проста и не требует развития таких сложных структур, как нервная система. Действительно, даже у самых примитивных многоклеточных животных - губок - некоторые клетки способны к секреции таких известных медиаторов нервной системы и гормонов более высокоорганизованных форм животного мира, как серо-тонин и ацетилхолин. У губок они служат для согласования деятельности отдельных частей тела. В организме иглокожих и брюхоногих моллюсков, а также низших хордовых обнаруживается инсулин. У большинства исследованных беспозвоночных найдены также стероидные гормоны, секретирующиеся клетками половых желез и ряда других органов. Вероятно, их происхождение очень древнее, так как они обнаружены даже у грибов и у многих видов растений.
Обсуждая вопрос о филогенезе эндокринной системы хордовых, необходимо рассмотреть происхождение и эволюцию как гормонов, так и самих желез внутренней секреции.
14.6.2.1. Гормоны
Химическая структура гормонов многообразна. Это могут быть белки (инсулин, пептидные нейрогормоны), стероиды (половые гормоны), продукты метаболизма отдельных аминокислот (тироксин, адреналин).
Они могут иметь и другое строение. Это свидетельствует о том, что гормоны разнообразны и по происхождению. Главное, что их объединяет, - способность специфически изменять клеточный метаболизм при контакте с цитоплазматической мембраной.
Предполагают, что уже древние одноклеточные организмы использовали биологически активные вещества для межклеточных коммуникаций. Некоторые вещества такого рода, обладающие регуляторными функциями, могли действовать как на отдельные клетки простейших, так позже и на клетки многоклеточных организмов. Впоследствии они и стали выполнять функции гормонов. Интересно, что в прогрессивной эволюции гуморальной регуляции структура самих гормонов может и не меняться. Доказательство этого - обнаружение таких известных гормонов, как адреналин, норадреналин и некоторые другие, в клетках простейших и низших растений, где они выполняют функции регуляторов клеточного деления, движения ресничек и вакуолей.
В эмбриогенезе многоклеточных ряд гормонов выявляется уже в первые часы и дни развития. В процессе дробления они регулируют течение клеточного цикла. Позже - перемещения клеток и образование межклеточных контактов, действуя либо внутри клеток, их продуцирующих, либо на близлежащие клетки. Гормоны приобретают свойства дистантных регуляторов в филогенезе только у трехслойных животных, а в онтогенезе многоклеточных - соответственно на стадии первичного органогенеза.
При неизменности химической структуры функции гормонов могут нередко изменяться. Так, гормон пролактин, выделяющийся у млекопитающих, в том числе человека, гипофизом и регулирующий секрецию молока млечными железами, обнаружен также у рыб, земноводных и птиц. У первых он регулирует выделение кожными железами слизи, которой питаются мальки, у вторых - образование оболочек икринок в яйцеводах, у третьих - некоторые элементы брачного поведения, а также выделение у кормящих родителей «зобного молочка». Как видно из этого примера, эволюция действия гормонов может быть канализированной выполнением одной и той же функции в самом широком смысле. Действительно, все перечисленные функции пролактина у животных, находящихся на разных уровнях организации, имеют отношение к обеспечению успешности размножения.
Функции других гормонов, например, адреналина, могут в филогенетическом ряду тех же позвоночных практически не меняться, обеспечивая у всех регуляцию в первую очередь энергетического обмена.
При этом часто гормоны продолжают выполнять те функции, которые первично проявлялись еще у одноклеточных и низших многоклеточных животных.
Так, адреналин в организме млекопитающих, кроме регуляции энергетического обмена, артериального давления и работы сердца, замедляет прохождение клеток по клеточному циклу и вступление их в митоз. Функция регуляции размножения у одноклеточных была исторически, по-видимому, главной. Из главной эта функция у многоклеточных животных превратилась во второстепенную. Таким образом, на филогенез гормонов, так же, как и на эволюцию органов, распространяются основные закономерности макроэволюционных преобразований биологических структур (см. гл. 13).
14.6.2.2. Железы внутренней секреции
Железы внутренней секреции, как и гормоны, выделяемые ими, имеют разное происхождение, что важно для изучения их эволюции.
Некоторые эндокринные железы связаны по происхождению с эпителиальной выстилкой глотки. К ним относятся щитовидная и паращитовидная железы. Эпифиз развивается как вырост мозга; гипофиз, надпочечники и поджелудочная железа имеют сложное происхождение.
Среди хордовых только у бесчерепных эндокринная система существует в виде отдельных клеток и клеточных комплексов, которые находятся в разных отделах тела, объединенных друг с другом за счет гуморального взаимодействия. У позвоночных в основании промежуточного мозга развивается гипоталамус - нейросекреторное образование, осуществляющее связь между двумя системами интеграции организма (нервной и эндокринной) в единое целое. Вместе с гипофизом гипоталамус образует единую гипоталамо-гипофизарную систему.
Эволюционный предшественник гипоталамуса - так называемый инфундибулярный вырост ланцетника, состоящий из нейросекретор-ных клеток и находящийся на вентральной стороне переднего конца нервной трубки.
Начиная с рыб, гипоталамус дифференцируется на многочисленные ядра, клетки которых с помощью отростков контактируют как с нейронами мозга, так и с клетками гипофиза. Нейросекреторные клетки гипоталамуса выделяют две основные группы гормонов: пептидные и моноаминовые.
Первые - гормоны, влияющие на функции внутренних органов - ва-зопрессин, регулирующий артериальное давление, окситоцин, действующий на мускулатуру матки, и др. Вторая группа гормонов (дофамин, норадреналин, серотонин) регулирует деятельность передней доли гипофиза. Их действие стимулирует или подавляет секрецию гормонов соответствующими гипофизарными клетками.
Подробнее остановимся на эволюции гипофиза. Эта железа состоит из трех долей: передней (аденогипофиза), средней (промежуточной) и задней (нейрогипофиза). Гипофиз соединен с гипоталамусом его выростом - воронкой, через которую проходят отростки нейронов гипоталамуса и кровеносные сосуды, обеспечивающие их гуморальное взаимодействие.
Доли гипофиза имеют разное происхождение. Передняя доля развивается из выпячивания эктодермального эпителия крыши ротовой полости, так называемого кармана Ратке, который растет в сторону промежуточного мозга. Задняя доля развивается из задней части воронки. Клетки, входящие в ее состав, по происхождению являются глиаль-ными. Промежуточная доля - производная от передней (рис. 14.44).
У хрящевых рыб во взрослом состоянии сохраняется первоначальная связь передней доли гипофиза с эпителием ротовой полости. За счет ее задней части формируется также средняя доля. Обе доли вырабаты-
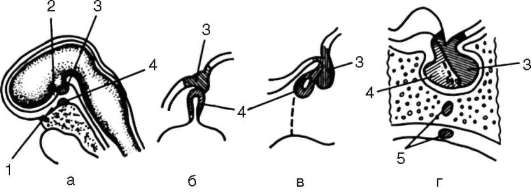
Рис. 14.44. Развитие гипофиза у человека: а - сагиттальный срез 4-недельного зародыша; б - объединение зачатков гипофиза из стенки глотки и основания мозга; в - формирование трех долей гипофиза на 8-й неделе развития; г - сформированный гипофиз: 1 - крыша ротовой полости; 2 - основание мозга; 3 - выпячивание основания мозга (задняя доля гипофиза); 4 - карман Ратке (промежуточная и передняя доли гипофиза); 5 - аномалия развития (добавочные дольки передней доли в стенке глотки и внутри клиновидной кости, оставшиеся в области выпячивания кармана Ратке)
вают гонадотропные гормоны. У костных рыб и личинок земноводных имеются передняя и промежуточная доли, а у взрослых амфибий, переходящих к наземному существованию, появляется также задняя, регулирующая водный обмен. Средняя доля у них перестает выделять гонадотропный гормон, но вырабатывает пролактин.
В связи с наземным образом жизни пресмыкающихся и млекопитающих у них наиболее прогрессивно развивается задняя доля гипофиза, что связано с интенсификацией водного обмена.
Передняя доля вырабатывает соматотропный гормон (гормон роста) и ряд гормонов, регулирующих функции других желез внутренней секреции, а средняя - пролактин и некоторые другие. При этом усиливается дифференцировка и интенсифицируются функции ядер гипоталамуса, находящихся в функциональной связи со всеми долями гипофиза.
У человека в эмбриогенезе развитие гипофиза соответствует основным этапам его эволюции. Очень часто, в 30-40%, у нормальных людей под слизистой оболочкой крыши глотки, в основании клиновидной кости, обнаруживается группа клеток длиной 5-6 мм и шириной 0,5-1 мм, по структуре и функциям соответствующая передней доле гипофиза. Это результат нарушения перемещения клеток при закладке гипофиза в эктодерме ротовой полости в области турецкого седла. Эту аномалию называют эктопией аденогипофиза, она не сопровождается патологическими проявлениями. Более опасно сохранение полости в области кармана Ратке - киста кармана Ратке. Она располагается между передней и промежуточной долями, содержит слизь и в ряде случаев имеет тенденцию к росту и даже к переходу в злокачественное новообразование. Растущие кисты сдавливают гипофиз и вызывают его гипофункцию. Кроме того, возможно сдавление области hiasma opticum, что проявляется в нарастающем сужении полей зрения. Больные нуждаются в оперативном вмешательстве.
Эпифиз, или верхний мозговой придаток, - эндокринная железа, связанная по происхождению с органом зрения. Его строение у низших водных позвоночных сходно со строением примитивного глаза. У рыб, земноводных и пресмыкающихся это фоторецепторный орган, одновременно выделяющий гормон мелатонин, регулирующий суточную активность животных и контролирующий синтез и разрушение темного пигмента меланина. У млекопитающих фоторецепторная функция эпифиза исчезает, однако он сохраняет тесную функциональную связь с сетчаткой глаза и получает от нее информацию как об освещенности, так и о продолжительности дня и ночи. От этих факторов зависит се-
креция мелатонина эпифизом. Поэтому функция регуляции суточной и годичной ритмической активности этой железой у млекопитающих, в том числе человека, в полной мере сохраняется.
Щитовидная железа, гормон которой тироксин регулирует энергетический обмен, среди хордовых как компактный орган впервые появляется у рыб. Однако уже у ланцетника отдельные тироксин-синтезирующие клетки обнаруживаются в эндостиле - желобке на вентральной стороне глотки. Щитовидная железа рыб закладывается также в виде желобка на вентральной стороне глотки между 1-й и 2-й жаберными щелями в области зачатка основания языка. Позже этот клеточный материал погружается под слизистую оболочку и формирует фолликулы, характерные для щитовидной железы. У других позвоночных железа закладывается так же, как у рыб, но затем она перемещается в область подъязычной кости (у земноводных) или в шейную область (у пресмыкающихся и млекопитающих).
У человека в эмбриогенезе щитовидной железы происходит рекапитуляция предковых состояний. Гетеротопия ее осуществляется посредством миграции клеток из эпителия корня языка в область расположения щитовидного хряща гортани. При этом образуется клеточный тяж, полый внутри, называющийся щитоязычным протоком - canalis thyreoglossus. При нормальном развитии этот проток полностью редуцируется, рудиментом его является слепое отверстие foramen caecum в корне языка.
Свидетельство эмбрионального перемещения железы - и расположение верхней щитовидной артерии a. thyreoidea superior, которая, начинаясь от общей сонной артерии, резко поворачивает вниз и входит в железистую ткань. На ранних этапах эмбрионального развития эта артерия направляется вверх к зачатку щитовидной железы, а затем меняет свое направление вместе с его перемещением книзу.
Персистирование участков щитоязычного протока в постнатальном периоде сопровождается накоплением в них жидкости и образованием срединных кист шеи, которые могут располагаться в любом месте от корня языка до верхней границы щитовидного хряща. Иногда кисты загнаиваются и прорываются с образованием срединных свищей шеи. Не менее известны такие пороки развития, как эктопия щитовидной железы, которая может развиться в результате нарушения клеточной миграции (рис. 14.45). Встречается в качестве аномалии расположение ткани щитовидной железы в области корня языка, а также существенно выше места ее дефинитивного положения - в области подъязычной
Рис. 14.45. Аномалии щитовидной железы: 1 - нормальное расположение железы; 2 - место эмбриональной закладки железы; 3 - варианты аномального расположения железы; показано направление перемещения зачатка щитовидной железы в онтогенезе
кости или несколько ниже ее - так называемая переднешейная локализация.
Из эпителия глотки в области III-V жаберных карманов у позвоночных развиваются мелкие эндокринные образования, связанные по месту окончательного положения со щитовидной железой. Это паращитовид-ные железы и ультимобранхиальные тельца. Первые, выделяя гормон паратиреоидин, повышающий содержание ионов кальция в крови и уменьшающий их количество в костях, развиваются как самостоятельные железы только у наземных позвоночных, а у земноводных - лишь после метаморфоза. Клетки вторых выделяют кальцитонин, антагонист паратиреоидина. Таким образом, паращитовидные железы и ультимобранхиальные тельца регулируют кальциевый обмен. Значение их наиболее велико у наземных позвоночных. В филогенетическом ряду позвоночных они постепенно перемещаются из глоточной области в сторону щитовидной железы, а у млекопитающих даже срастаются с ней. Паращитовидные железы еще сохраняют самостоятельность, а клетки ультимобранхиальных телец мигрируют между фолликулами щитовидной железы и сохраняются под названием парафолликулярных клеток. Таким образом, на примере щитовидной, паращитовидных желез и ультимобранхиальных телец видна интеграция железистых структур в сложное надорганное образование, выполняющее целый комплекс взаимосвязанных функций.
У человека известны аномалии расположения паращитовидных желез, связанные с нарушением их гетеротопий.
Надпочечники позвоночных имеют двойственное происхождение. У рыб и земноводных ткани, соответствующие мозговому и корковому веществам этих желез, расположены отдельно друг от друга. Зачатки мозгового вещества связаны по происхождению с симпатическими нервными узлами и расположены метамерно. Зачатки коркового вещества развиваются из утолщений эпителия брюшины. У наземных позвоночных мозговое и корковое вещества объединяются в компактные эндокринные железы, имеющие сложное гистологическое строение. У млекопитающих они прилежат к переднему концу почек. Мозговое вещество выделяет в основном адреналин - регулятор кровообращения и энергетического обмена, а корковое вещество - разнообразные стероидные гормоны, влияющие на минеральный и углеводный обмены, функции почек, половых желез и формирование ряда вторичных половых признаков.
14.7. ЗАЩИТА И ПОДДЕРЖАНИЕ ЦЕЛОСТНОСТИ ОРГАНИЗМА, ПОСТОЯНСТВА ЕГО ВНУТРЕННЕЙ СРЕДЫ, ОПТИМАЛЬНЫХ УСЛОВИЙ ДЛЯ ПРОЦЕССОВ ЖИЗНЕДЕЯТЕЛЬНОСТИ. ЭВОЛЮЦИЯ ИММУННОЙ
СИСТЕМЫ
Живые формы в среде обитания испытывают на себе влияние многих факторов абиотической и биотической природы, действие которых является вредоносным, снижающим показатели жизнеспособности вплоть до состояния несовместимости с жизнью. Некоторые факторы защиты, повышающие шансы на выживание, общеизвестны - панцирь черепахи, внешние покровы червей, паразитирующих в просвете тонкой кишки человека (см. выше). В среде жизни многоклеточных организмов присутствует множество одноклеточных форм, ведущих паразитическое существование, патогенных грибов, микроорганизмов и вирусов. Задача воспрепятствовать их проникновению и вмешательству в процессы жизнедеятельности настолько важна, что процесс исторического развития жизни на Земле не мог остаться в стороне от нахождения и закрепления в структурно-функциональной организации живых форм соответствующих механизмов.
Одна из «удач» процесса исторического развития на этапе многокле-точности - появление иммунной системы. Вместе с тем в филогенезе
системы иммунитета в ее наиболее эволюционно продвинутом варианте, характерном для подтипа Позвоночные, принято выделять доиммун-ный и иммунный периоды. При этом ряд неспецифических клеточных (фагоцитоз, натуральные клетки-киллеры), неклеточных гуморальных (система комплемента, белки острой фазы, интерфероны), связанных с местными локальными или общими реакциями организма (воспаление, подъем температуры или лихорадка) механизмов, появившихся в доиммунном периоде, функционирует также в организме позвоночных животных, в том числе млекопитающих.
Важнейшая отличительная черта иммунного периода в сравнении с доиммунным состоит, видимо, в появлении у представителей подтипа Позвоночные лимфоцитов (впервые в группе Бесчелюстные - кругло-ротые рыбы, миноги), которые подразделяются на Т- и В-лимфоциты (впервые в группе Челюстные - хрящевые рыбы, акулы). Субпопуляцией В-лимфоцитов обеспечивается гуморальная составляющая системы иммунитета позвоночных, основанная на образовании антител, что придает иммунному ответу свойство специфичности. Субпопуляция Т-лимфоцитов, играющая ключевую роль в клеточной составляющей системы иммунитета позвоночных, также функционирует на основе специфических взаимодействий «антитело-антиген».
У позвоночных животных выделяют три «эволюционно проработанных» линии защиты от неблагоприятных последствий взаимодействия с микроорганизмами. На масштаб таких последствий указывает тот факт, что за 18 мес 1918-1919 гг. в связи с пандемией гриппа («испанка»), вызванной соответствующим вирусом, на планете погибло порядка 22 млн человек. Как отмечалось выше, некоторые из механизмов, в частности, второй линии защиты функционируют как у представителей подтипа Позвоночные, так и у других многоклеточных животных. Этот факт можно рассматривать как проявление феномена преадаптации в эволюции иммунной системы высших животных.
Первая линия защиты, сложившаяся на заре возникновения живых существ как дискретных обособленных от внешнего мира единиц (организмов, особей), связана с механо-химическими, барьерными в своей основе свойствами пограничных структур, отграничивающих организм от окружающей среды - клеточные оболочки прокариот и одноклеточных эукариот, кожные и другие покровные образования, слизистые оболочки многоклеточных. На этом этапе, видимо, произошла дивергенция эволюционной стратегии: в прокариотические клетки чужеродная генетическая (био)информация обычно проникает в виде информационной
макромолекулы или ее фрагментов фактически без белков, что определило развитие семейства ферментов, разрушающих чужеродную ДНК; в эукариотические клетки, особенно многоклеточных живых форм чужеродная генетическая (био)информация проникает в виде ДНК с сопровождающими ее продуктами экспрессии биоинформации, то есть белками, на которые (как на чужеродные) эукариотический организм реагирует в первую очередь.
Вторая линия защиты включается в случаях, когда микроорганизм преодолевает внешние барьеры и проникает внутрь организма. Действующие агенты этой линии защиты - совокупность клеток определенного типа и набор определенных химических (преимущественно белковых) соединений, способных к обнаружению чужеродного агента и его разрушению. Клеточные и гуморальные защитные факторы второй линии защиты неспецифичны относительно агента, против которого направлено их действие. В своей совокупности с наличием этих факторов связывают неспецифический иммунный ответ организма на проникновение в него чужеродного, в частности инфекционного, агента (инфекционная иммунология).
Указанный ответ обеспечивается, во-первых, наличием клеток-фагоцитов - у позвоночных макрофагов и нейтрофилов, непосредственно разрушающих микроорганизмы и другие патогены, а также наличием натуральных (естественных, природных) клеток-киллеров (natural killer cells), которые разрушают не инфекционный агент непосредственно, а собственные клетки организма, пораженные названным агентом.
Возникшие в эволюции наиболее рано (фактически с появлением многоклеточных живых форм) механизмы защиты и сохранения постоянства внутренней среды были связаны с клетками, фагоцитирующими чужеродные агенты биологического происхождения. Действительно, уже у губок наблюдается отторжение реципиентом клеточного материала донора при межвидовых пересадках. Для того чтобы указанный механизм работал, необходимо, чтобы многоклеточный организм мог отличать «свое» и «чужое», причем эволюционно возможность такого разграничения должна была оформиться достаточно рано. Так оно, видимо, и случилось.
У современных беспозвоночных животных, например, указанное разграничение, исключающее разрушение фагоцитами собственных структур, происходит благодаря особой белковой метке в оболочке «своих» клеток. Таким образом, в мире беспозвоночных животных «чужой» идентифицируется как «не свой» (отрицательный принцип
тестирования). Механизмы идентификации «чужого» как действительно «чужого» на основе отличий в белковом (антигенном) составе (положительный принцип тестирования), что отражает генетически-биоинформационные различия между инфицируемым объектом и ин-фектом или между донором и реципиентом биологического материала, появляются в филогенезе позже (подтип Позвоночные) и связаны с оформлением системы специфического иммунитета, в основе которой лежит жизнедеятельность особых клеток Т- и В-лимфоцитов, продукция организмом специальных белков-антител и появление особых антиген-представляющих (антигенпрезентирующих) клеток. С возникновением системы специфического иммунитета стал реальностью важный биологический феномен «иммунологической памяти», благодаря которому оказалась возможной вакцинопрофилактика ряда заболеваний, прежде всего инфекционных (бактериальных, вирусных). В отличие от макрофагов и нейтрофилов, механизм действия натуральных клеток-киллеров - не фагоцитоз. Выделяя белки перфорины, клетки-киллеры проделывают в оболочке агента-мишени отверстия (поры), через которые другие белки (гранзимы), проникая внутрь агента, вызывают его разрушение (например, запуская процесс апоптоза). Объектом действия натуральных киллеров могут служить собственные клетки организма, в частности те, которые, выйдя из-под контроля общеорганизменных регуляторных механизмов, встают на путь онкотрансформации, то есть превращаются в опухолевые (раковые). Такое уничтожение онкотранс-формированных клеток происходит нередко задолго до того, как образуется опухоль, которую удается диагностировать - эволюционный принцип расширения и смены функций.
Учитывая сказанное, наличие клеток-киллеров можно рассматривать как свидетельство в пользу функционирования в многоклеточном организме механизма активного контроля постоянства внутренней среды (в частности, клеточного состава) организма - «иммунологический надзор» (неинфекционная иммунология). У позвоночных с высокоразвитой иммунной системой функцию «иммунологического надзора» выполняет субпопуляция цитотоксических Т-лимфоцитов. Вопрос о том, является ли смена натуральных клеток-киллеров низших многоклеточных на цитотоксические Т-лимфоциты позвоночных животных примером тканевой субституции, остается открытым.
Эволюционно более ранние гуморальные факторы второй линии защиты от проникновения в многоклеточный организм чужеродного биологического объекта также неспецифичны в отношении объекта-
мишени и включают систему комплемента, интерфероны, белки острой фазы и некоторые другие.
Система комплемента позвоночных (млекопитающих, в том числе человека) представлена белками (порядка 30), циркулирующими в составе плазмы крови. При контакте со стенкой микробной клетки эти белки агрегируют друг с другом. Такой агрегат, внедряясь в клеточную стенку чужеродного объекта, образует пору. Через пору внутрь объекта поступает жидкость, в результате чего он набухает и, лопаясь («взрываясь»), погибает. Белки системы комплемента выполняют ряд других функций: являются аттрактантами для макрофагов и нейтрофилов, благодаря чему последние попадают именно в неблагополучный район организма, например в воспалительный очаг; покрывая поверхность чужеродной клетки и делая ее более грубой, они способствуют более быстрому фагоцитозу.
Интерфероны-α и β образуются в соматических клетках, особенно инфицированных вирусом, и препятствуют инфицированию расположенных в непосредственной близости непораженных клеток, а также блокируют репликацию вирусной ДНК и сборку вирусных белков. Лимфоциты определенной субпопуляции и натуральные клетки-киллеры образуют γ-интерферон. Конкретные точки приложения действия γ-интерферона не известны. Тем не менее общепринято мнение, что γ-интерферон играет ключевую антиинфекционную и антираковую роль - эволюционный принцип расширения функций.
Белки острой фазы сыворотки крови, в частности С-реактивный белок, представляющие собой растворимые рецепторы для патогенов, прежде всего, инфекционной природы. С этими белками связывают феномен опсонизации, заключающийся в том, что такие белки (опсонины) благодаря способности связываться одновременно с микроорганизмом (патогеном) и клеткой-фагоцитом способствуют более быстрому разрушению инфекта путем фагоцитоза.
К доиммунным, то есть эволюционно возникшим достаточно рано, механизмам защиты и поддержания целостности организма, постоянства его внутренней среды и оптимальных условий для процессов жизнедеятельности, направленным на уничтожение или локализацию (блок распространения патогена по организму) проникших в многоклеточный организм микроорганизмов, могут быть отнесены также общий подъем температуры тела и воспаление.
В эволюционном плане такая реакция на появление в организме чужеродного агента (бактериального, вирусного, паразитарного), как
подъем температуры тела (лихорадка), если отталкиваться от известных науке фактов, решалась на разных уровнях эволюции, особенно прогрессивной, по-разному. У высших позвоночных, в частности у млекопитающих (гомойотермные животные), макрофагами выделяется вещество (интерлейкин-I) с пирогенной регуляторной функцией, которое проникает в головной мозг и активирует определенные нервные клетки гипоталамической области, что, собственно, и приводит к повышению температуры тела. В условиях повышенной температуры активируется фагоцитоз, а печень и селезенка задерживают железо, необходимое для размножения микроорганизмов. Инфицированные ящерицы (пой-килотермные животные) перемещаются из прохладных участков места обитания в более теплые. Если такое перемещение невозможно, процесс выздоровления замедляется.
У людей подъем температуры свыше 39,4 °С рассматривается как опасный для жизни: при названной температуре многие жизненно необходимые (критические) белки-ферменты денатурируют с утратой каталитической активности. Подъем температуры тела человека до 40,6 °С нередко оказывается фатальным.
Воспалительный процесс можно рассматривать как эволюционно обусловленную местную (локальную) достаточно сложную (в частности, по составу участников - веществ и клеток) реакцию многоклеточного организма на проникновение в него инфекционного агента с целью предотвратить распространение (локализовать) и уничтожить названный агент. В очаге воспаления создаются условия (интенсивный кровоток, отек, местный подъем температуры, скопление макрофагов и нейтрофи-лов), способствующие достижению указанной цели. Поскольку воспалительная реакция может быть следствием травмы или проникновения в организм чужеродного тела (заноза), она полностью лишена свойства специфичности и не может рассматриваться исключительно как фактор антимикробной направленности.
Как уже отмечалось, третья, эволюционно наиболее поздняя линия защиты целостности многоклеточного организма, постоянства его внутренней среды (включая клеточный состав) связана с появлением иммунной системы, основанной на появлении специализированных клеток (Т- и В-лимфоциты, антигенпредставляющие клетки и ряд других клеточных типов), на возникновении специфических органов, формирующих иммунную систему высших животных (лимфатические узлы, селезенка, тимус млекопитающих, возможно, фабрициева сумка птиц, скопления лимфоидной ткани - миндалины, пейеровы бляшки). В по-
строении и функционировании названной системы, как это свойственно процессу исторического развития в принципе (явление преадаптации), эффективно используются другие уже состоявшиеся эволюционные «наработки» - системы циркуляции: незамкнутая (лимфообращение) и замкнутая (кровообращение). Принципиальный момент, отличающий механизмы третьей линии защиты (иммунная система) у высших многоклеточных организмов состоит в том, что в основе функции иммунной системы в ее эволюционно наиболее продвинутом варианте (теплокровные, в частности, млекопитающие) лежит принцип «белок-белковых» взаимодействий (идентификация антигенов и образование к ним антител). Это делает иммунитет специфичным, расширяет функции иммунной системы (инфекционная и неинфекционная иммунология), создает как благоприятные для практического здравоохранения (иммунологическая память), так и проблемные (аутоиммунные патологические состояния, иммунология онкотрансформированных клеток) ситуации.
Вопросы для самоконтроля
1. Каковы общие закономерности эволюции систем органов хордовых?
2. Приведите примеры рекапитуляций и филэмбриогенезов в эволюции конкретных систем органов позвоночных.
3. Приведите примеры врожденных пороков развития разных систем органов человека и объясните возможные механизмы их формирования.
4. Приведите примеры пороков развития человека, в формировании которых ведущим механизмом является нарушение диффе-ренцировки.
5. Приведите примеры пороков развития, возникающих в результате нарушения гетеротопии.
6. Как можно объяснить механизмы формирования в зародышевом развитии следующих аномалий: двурогая матка, полидактилия, гипертрихоз, расщелина твердого нёба, язычная локализация щитовидной железы, крипторхизм, полидентия, шейные свищи, кистозная гипоплазия легких, персистирование обеих дуг аорты, тазовое расположение почки, платиневрия.
7. Какие пороки развития человека могут объясняться нарушением тканевой субституции, редукции и сращения парных зачатков?
8. Приведите примеры пороков развития, в формировании которых ведущим механизмом является нарушение онтогенетических корреляций.
9. Объясните, почему пороки развития, возникающие на более ранних стадиях онтогенеза, обычно характеризуются множественностью и высокой степенью тяжести?
10. Объясните, почему врожденная гипоплазия нижней челюсти часто сопровождается аномалиями слухового анализатора, щитовидной железы, глотки и гортани.
11. Какие пороки развития человека могут быть объяснены нарушениями эргонтических корреляций?
12. Приведите примеры геномных корреляций и их нарушений у человека.