Медицинская и биологическая физика: учебник / А. Н. Ремизов. - 4-е изд., испр. и перераб. - 2012. - 648 с. : ил.
|
|
|
|
Глава 13. Физические процессы в биологических мембранах
Важной частью клетки являются биологические мембраны. Они ограничивают клетку от окружающей среды, защищают ее от вредных внешних воздействий, управляют обменом веществ между клеткой и ее окружением, способствуют генерации электрических потенциалов, участвуют в синтезе универсальных аккумуляторов энергии АТФ в митохондриях и т.д. По существу, мембраны формируют структуру клетки и осуществляют ее функции. Многие заболевания (атеросклероз, отравления и др.) связаны с нарушением структуры и функции мембран.
В главе рассматриваются физические свойства биологических мембран и основные физические процессы, которые в них происходят.
13.1. СТРОЕНИЕ И МОДЕЛИ МЕМБРАН
Мембраны окружают все клетки (плазматические или наружные клеточные мембраны). Без мембраны содержимое клетки просто бы растеклось, диффузия привела бы к термодинамическому равновесию, что означает отсутствие жизни. Можно сказать, что первая клетка появилась тогда, когда она смогла отгородиться от окружающей среды мембраной.
Внутриклеточные мембраны подразделяют клетку на ряд замкнутых отсеков (компартаментов), каждый из них выполняет определенную функцию.
Толщина мембран порядка нескольких нанометров, поэтому ее нельзя увидеть в оптический микроскоп (см. 26.8), но можно рассмотреть в электронный микроскоп (см. 28.2).
Основу структуры любой мембраны представляет двойной ли-пидный слой (в значительной степени фосфолипиды). Молекулы липи-дов, образующие мембрану, являются алимфатическими соединениями, т.е. состоят из двух функционально различных частей: полярной «головки» и неполярного гидрофобного «хвоста» (рис. 13.1). Двойной липид-ный слой образуется из двух монослоев липидов так, что гидрофобные «хвосты» обоих слоев направлены внутрь. При этом обеспечивается


электрического потенциала и т.д. Схематично жидкомозаичная структура мембраны показана на рис. 13.3. Здесь 1 - поверхностные белки, 2 - полупогруженные белки, 3 - полностью погруженные (интегральные) белки, 4 - белки, формирующие ионный канал 5.
Мембраны не являются неподвижными, «спокойными» структурами. Липиды и белки обмениваются местами и перемещаются как вдоль плоскости мембраны - латеральная диффузия, так и поперек нее - так называемый «флип-флоп». Латеральной диффузии соответствует высокая подвижность липидов, а «флип-флопу» - низкая, т.е. обмен местами липидов, находящихся на разных сторонах мембраны, является редким процессом.
Уточнение строения биомембраны и изучение ее свойств оказалось возможно при использовании физико-химических моделей мембраны (искусственные мембраны). Наибольшее распространение получили три такие модели.
Рассмотрим первую модель - монослои фосфолипидов на границе раздела вода-воздух или вода-масло.
На таких границах молекулы фосфолипидов расположены так, что гидрофильные головки находятся в воде, а гидрофобные «хвосты» - в воздухе или в масле. Если постепенно уменьшать площадь, занимаемую монослоем, то в конце концов удастся получить монослой, в котором молекулы расположены так же плотно, как и в одном из бислоев мембраны (рис. 13.4).
Второй широко распространенной моделью биомембраны являются липосомы.
Это мельчайшие пузырьки (везикулы), состоящие из билипидной мембраны и полученные обработкой смеси воды и фосфолипидов ультразвуком. Липосомы представляют собой как бы биологическую мембрану, полностью лишенную белковых молекул. На липосомах часто проводятся эксперименты по изучению влияния различных факторов, например состава фосфолипидов, на свойства мембраны или,
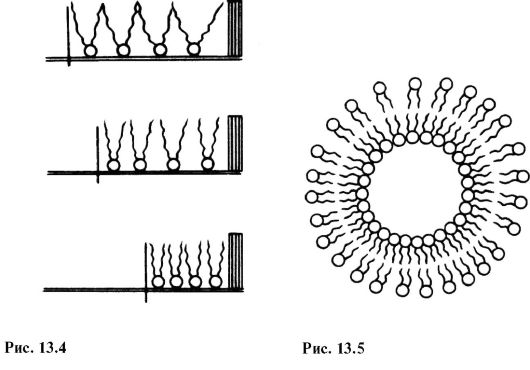
наоборот, влияния мембранного окружения на свойства встраиваемых белков. Схематически липосомы изображены на рис. 13.5. Третьей моделью, позволившей изучать некоторые свойства биомебран прямыми методами, была билипидная (бислойная липидная) мембрана (БЛМ).
Впервые такая модельная мембрана была создана в 1962 г. П. Мюллером с сотрудниками. Они заполнили отверстие в тефлоновой перегородке, разделяющей два водных раствора, фосфолипидом, растворенным в гептане. После того как растворитель и излишки липида растекаются по тефлону, в отверстии образуется бислой толщиной несколько нанометров и диаметром около 1 мм. Расположив по обе стороны мембраны два электрода, можно измерить сопротивление мембраны или генерируемый на ней потенциал. Если исходно по разные стороны перегородки поместить различные по химическому составу растворы, можно изучать проницаемость мембраны для различных агентов.
Мембраны выполняют две важные функции: матричную, т.е. являются матрицей, основой для удержания белков, выполняющих разные функции, и барьерную - защищают клетку и отдельные компартамен-ты от проникновения нежелательных частиц. Если эти функции мембран нарушаются, то происходят изменение нормального функционирования клеток и, как следствие, заболевание организма.
13.2. НЕКОТОРЫЕ ФИЗИЧЕСКИЕ СВОЙСТВА И ПАРАМЕТРЫ МЕМБРАН
Измерение подвижности молекул мембраны и диффузии частиц через мембрану свидетельствует о том, что билипидный слой ведет себя подобно жидкости. В то же время мембрана - упорядоченная структура. Эти два фактора заставляют думать, что фосфолипиды в мембране при ее естественном функционировании находятся в жидкокристаллическом состоянии (см. 10.2). Жидкостные свойства мембраны подтверждаются методами ЭПР (см. 30.3) и ЯМР (см. 30.4).
Вязкость липидного слоя мембран приблизительно на два порядка выше вязкости воды, она равна 30-100 мПас, что соответствует примерно вязкости растительного масла. Поверхностное натяжение на 2-3 порядка ниже (0,03-1 мН/м), чем у воды.
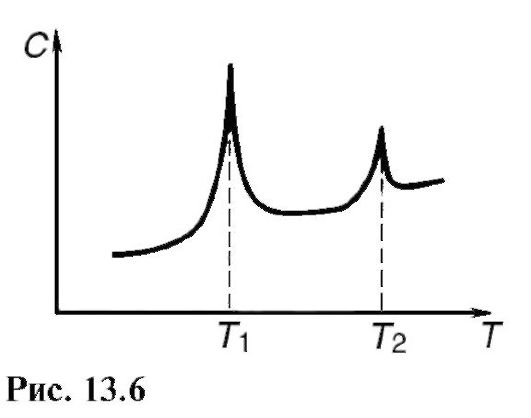
При изменении температуры в мембране можно наблюдать фазовые переходы: плавление липидов при нагревании и кристаллизацию при охлаждении. Фазовые переходы связаны с изменением энергии и поэтому могут быть обнаружены, в частно -сти, по увеличению теплоемкости с при изменении температуры (рис. 13.6; при температурах T1 и Т2 - фазовые переходы). Жидкокристаллическое состояние бислоя имеет меньшую вязкость и большую растворимость различных веществ, чем твердое состояние. Толщина жидкокристаллического биослоя меньше, чем твердого.
Конформация (структура) молекул в жидком и твердом состояниях различна, в чем можно убедиться при рентгеноструктурном анализе (см. 24.7). В жидкой фазе молекулы фосфолипидов могут образовывать полоски (кинки), в которые способны внедряться молекулы диффундирующего вещества.
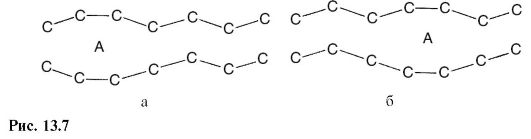
Перемещение «кинка» в этом случае будет приводить к диффузии молекулы поперек мембраны (рис. 13.7).
Двойной фосфолипидный слой уподобляет мембрану конденсатору, электроемкость 1 мм2 мембраны составляет 5-13 нФ.
13.3. ПЕРЕНОС МОЛЕКУЛ (АТОМОВ) ЧЕРЕЗ МЕМБРАНЫ
Важным элементом функционирования мембран является их способность пропускать или не пропускать молекулы (атомы) и ионы. Существенно, что вероятность такого проникновения частиц зависит как от направления их перемещения, например в клетку или из клетки, так и от разновидности молекул и ионов.
Эти вопросы рассматриваются в разделе физики, относящемся к явлениям переноса. Таким термином называют необратимые процессы, в результате которых в физической системе происходит пространственное перемещение (перенос) массы, импульса, энергии, заряда или какой-либо другой физической величины.
К явлениям переноса относят диффузию (перенос массы вещества), вязкость (перенос импульса), теплопроводность (перенос энергии), электропроводность (перенос электрического заряда). Здесь и в следующих параграфах рассматриваются наиболее существенные для биологических мембран явления: перенос вещества и перенос заряда. Как синоним переноса частиц в биофизике широкое распространение получил термин «транспорт частиц».
Выведем основное уравнение диффузии (уравнение Фика), рассматривая процесс переноса в жидкостях.
Пусть через некоторую площадку площадью S (рис. 13.8) во всех направлениях перемещаются молекулы жидкости. Учитывая теорию молекулярного строения жидкости (см. 9.6), можно сказать, что молекулы пересекают площадку, перескакивая из одного положения равновесия в другое.
На расстояниях, равных среднему перемещению δ молекул (среднее расстояние между молекулами жидкости), вправо и влево от площадки построим прямо-
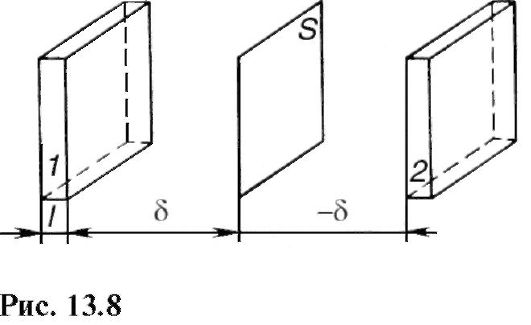
угольные параллелепипеды небольшой толщины l (l < δ). Объем каждого параллелепипеда равен Sl. Если п - концентрация молекул, то внутри выделенных параллелепипедов имеется Sln молекул. Предположим, что концентрация молекул изменяется в пространстве, в левом (1) выделенном параллелепипеде концентрация равна n1, а в правом (2) - п2. Следовательно, в одном параллелепипеде Sln1 молекул, а в другом - Sln2 молекул.
Все молекулы вследствие хаотичного их движения можно условно представить шестью группами, каждая из которых перемещается вдоль или против направления одной из осей координат. Отсюда следует, что в направлении, перпендикулярном площадке S, т.е. вдоль оси ОХ, от первого параллелепипеда перескакивает 1/6 Sln1 молекул, а противоположно оси ОХ от второго параллелепипеда перескакивает 1/6 Sln2 молекул.
Время At «пролета» этими молекулами площадки S может быть найдено следующим образом. Предположим, что все молекулы из выделенных объемов движутся с одинаковыми средними скоростями 5. Тогда молекулы в объеме 1 или 2, дошедшие до площадки S, пересекают ее в течение промежутка времени:

т.е количество вещества (масса), которое за 1 с переносится через площадку. Изменение концентрации п1-п2 молекул можно представить как произведение dn/dx на расстояние 2δ между выделенными объемами:

противоположную градиенту концентрации), D - коэффициент диффузии, применительно к рассмотренному примеру диффузии в жидкости он равен



13.4. УРАВНЕНИЕ НЕРНСТА-ПЛАНКА. ПЕРЕНОС ИОНОВ ЧЕРЕЗ МЕМБРАНЫ
Как будет показано в 13.6, на мембране существует разность потенциалов, следовательно, в мембране имеется электрическое поле. Оно оказывает влияние на диффузию заряженных частиц (ионов и электронов). Между напряженностью поля и градиентом потенциала существует известное соотношение (см. 14.1):




Это соответствует электропроводимости в электролите (см. 15.3). Для нейтральных частиц (Z = 0 и ψ = 0) J = 0;
в) если мембрана непроницаема для частиц (Р = 0), то, естественно, плотность потока равна нулю.
13.5. АКТИВНЫЙ ТРАНСПОРТ
Явления переноса (см. 13.3 и 13.4) относятся к пассивному транспорту, диффузия молекул и ионов в направлении меньшей их концентрации, перемещение ионов в соответствии с направлением силы, действующей на них со стороны электрического поля.
Пассивный транспорт не связан с затратой химической энергии, он осуществляется в результате перемещения частиц в сторону меньшего электрохимического потенциала (см 12.5). Наряду с пассивным транспортом в мембранах клетки происходит перенос молекул и ионов в сторону большего электрохимического потенциала (молекулы переносятся в область большей их концентрации, ионы - против силы, действующей на них со стороны электрического поля).
Этот перенос осуществляется за счет энергии и не является диффузией - активный транспорт. Системы мембран, способствующие созданию градиентов ионов К+ и Na+, получили название натрий-калиевых насосов или, проще, натриевых насосов.
Натрий-калиевые насосы входят в состав цитоплазматических мембран, они работают за счет энергии гидролиза молекул АТФ с образованием молекул АДФ и неорганического фосфата (Фн):

Так, если принять концентрацию ионов К+ внутри клетки в 50 раз большей, чем вне клетки, то, согласно (13.38), при температуре 36 °С получим
А = 8,31 Дж/(моль-К) ? 309Kln50 = 10 кДж/моль.
Механизм работы натрий-калиевого насоса недостаточно ясен, однако существенно, что он работает при условии сопряжения калия и натрия. Это означает, что активного переноса ионов Na+ из клетки нет, если во внешней среде нет ионов К+, а ионов К+ - в клетку, если в клетке отсутствуют ионы Na+. Иначе говоря, ионы натрия активируют натрий-калиевый насос на внутренней поверхности клеточной мембраны, а ионы калия - на внешней.
Натрий-калиевый насос переносит из клетки во внешнюю среду три иона натрия в обмен на перенос двух ионов калия внутрь клетки. При этом создается и поддерживается разность потенциалов на мембране, внутренняя часть клетки имеет отрицательный заряд (см. 13.7).
13.6. РАЗНОВИДНОСТИ ПАССИВНОГО ПЕРЕНОСА МОЛЕКУЛ И ИОНОВ ЧЕРЕЗ БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Простая диффузия через липидный слой схематически показана на рис. 13.11, она подчиняется уравнению Фика для молекул (13.9) или в более общем случае для нейтральных и заряженных частиц - уравнению Нернста-Планка (13.31). В живой клетке такая диффузия обеспечивает прохождение кислорода и углекислого газа. Ряд лекарственных веществ и ядов также проникает через липидный слой по схеме, изображенной на рисунке. Как было отмечено в 13.2, определенная конфор-мация фосфолипидов способствует диффузии поперек мембраны благодаря перемещению кинков.
Однако подобная простая диффузия протекает достаточно медленно и не может снабдить клетку в нужном количестве питательными веществами. Поэтому есть и иные механизмы пассивного переноса вещества в мембране, к ним относятся диффузия через канал (пору) (рис. 13.12) и облегченная диффузия (в комплексе с переносчиком).
Порой или каналом называют участок мембраны, включающей белковые молекулы и липиды, который образует в мембране проход. Этот проход допускает проникновение через мембрану не только малых
молекул, например молекул воды, но и более крупных ионов. Диффузия через поры также описывается диффузионными уравнениями, однако наличие пор увеличивает проницаемость Р. Каналы могут проявлять селективность (избирательность) по отношению к разным ионам, это проявится и в различии проницаемости для разных ионов.
Еще одно облегчение диффузии - это перенос ионов специальными молекулами-переносчиками (рис. 13.13). Наиболее известна способность молекулы валиномицина (антибиотик) переносить через модельные бислойные мембраны ион калия. Эта молекула захватывает ион К+, образует растворимый в липидах комплекс и проходит через мембрану. За способность переносить ионы через мембраны валино-мицин и другие родственные ему соединения получили название ионо-форов.
Транспорт с помощью переносчиков может осуществляться и в варианте эстафетной передачи. В этом случае молекулы-переносчики образуют временную цепочку поперек мембраны и передают друг другу диффундирующую молекулу.
Диффузия комплекса молекулы валиномицина и иона калия также описывается общим уравнением диффузии.
Перенос ионов через мембраны исследовал В.Ф. Антонов.
13.7. ПОТЕНЦИАЛ ПОКОЯ
Поверхностная мембрана клетки не одинаково проницаема для разных ионов. Кроме того, концентрация каких-либо определенных ионов различна по разные стороны мембраны, внутри клетки поддерживается наиболее благоприятный состав ионов. Эти факторы приводят к появлению в нормально функционирующей клетке разности потенциалов между цитоплазмой и окружающей средой (потенциал покоя).

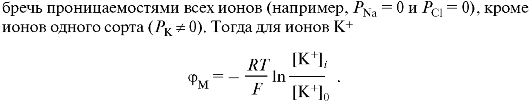
13.8. ПОТЕНЦИАЛ ДЕЙСТВИЯ И ЕГО РАСПРОСТРАНЕНИЕ
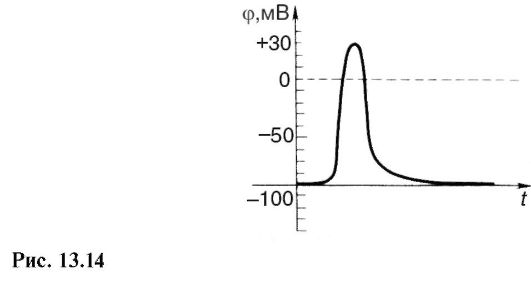
При возбуждении разность потенциалов между клеткой и окружающей средой изменяется, возникает потенциал действия (рис. 13.14). Он напоминает апериодические процессы, происходящие при зарядке и разрядке конденсатора (см. 18.1). В нервных волокнах происходит распространение потенциала действия.
Измерение сопротивления мембраны во время возникновения потенциала действия показало, что оно изменяется, повторяя по формуле временную зависимость потенциала действия. Чтобы решить вопрос, для каких ионов изменяется проницаемость мембраны, следует обратить внимание, что потенциал действия приводит к кратковременному возрастанию потенциала внутри клетки (рис. 13.14). Отрицательный относительно внешней среды потенциал становится положительным. Если по уравнению Нернста (12.46) вычислить равновесные потенциалы на мембране аксона кальмара, то получим соответственно для ионов К+, Na+ и С1- -90, +46, и -29 мВ. Так как при изменении проницаемости мембраны для какого-либо иона этот ион будет проникать через нее, стремясь создать равновесное состояние, то числовые данные показывают, что внутрь клетки проникают ионы Na+, создавая там положи-
тельный потенциал. Следовательно, при возбуждении клетки в начальный период увеличивается проницаемость мембран именно для ионов натрия.
Измерить проницаемость мембран для какого-либо иона (иначе говоря, электропроводимость или сопротивление мембраны для этого иона) можно, если на основании закона Ома найти отношение тока к напряжению, или наоборот. Практическая реализация такой задачи осложняется тем, что проницаемость (электрическое сопротивление) мембраны при возбуждении изменяется со временем. Это приводит к перераспределению электрического напряжения в цепи, и разность потенциалов на мембране изменяется. Ходжкин, Хаксли и Катц смогли создать опыт с фиксацией определенного значения разности потенциала на мембране. Это позволило им провести измерение ионных токов и, следовательно, проницаемости (сопротивления) мембран для ионов.
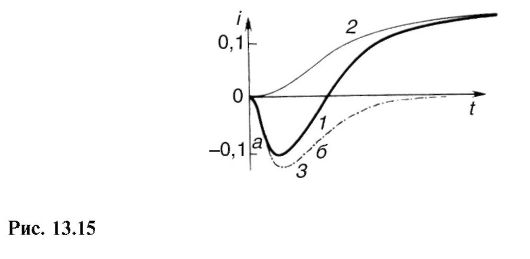
Результаты этой интересной работы приведены на рис. 13.15. Кривая 1 соответствует временной зависимости ионного тока через мембрану гигантского аксона кальмара, полученного при изменении потенциала на мембране до +56 мВ (потенциал покоя равен -60 мВ). Направление тока отрицательно, что означает прохождение положительных ионов через мембрану внутрь клетки. Этот ток в начальный период возбуждения, как уже было установлено, обусловлен прохождением ионов Na+ внутрь клетки, где концентрация их значительно меньше, чем снаружи. Естественно, что при таком нарушении равновесия ионы К+ начнут перемещаться наружу, где их концентрация существенно меньше. Для того чтобы выяснить, какая часть тока «натриевая», а какая «калиевая», можно провести то же возбуждение, но в искусственных условиях, когда в среде, окружающей аксон, нет Na+. В этом случае (см. кривую 2) ток обусловливается только выходом ионов К+ наружу из клетки. Раз-

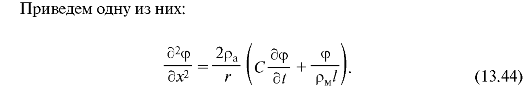
Здесь Ψ - внутриклеточный потенциал, измеряемый относительно потенциала покоя. Он зависит от расстояния х вдоль нервного волокна и времени t; pa и рм - удельные сопротивления соответственно аксо-плазмы и мембраны; r - радиус нервного волокна; См - электроемкость единицы площади мембраны.
Уравнение (13.44), как уравнение в частных производных, выходит за пределы этого курса.
Наряду с некоторой формальной аналогией между волной возбуждения (распространение потенциала действия по нервному волокну) и электромагнитной волной в двухпроводной линии между этими волнами имеется существенное принципиальное различие.
Электромагнитная волна, распространяясь в среде, ослабевает, так как растрачивает свою энергию. Имеет место диссипация энергии колебаний, т.е. превращение энергии колебаний (волн) в энергию молекулярно-теплового движения.
Источником энергии электромагнитной волны является источник этой волны: генератор, искра и т.п.
Волна возбуждения не затухает, так как получает энергию из самой среды, в которой она распространяется (в рассматриваемом примере - энергия заряженной мембраны).
Волны, получающие энергию из среды в процессе распространения, названы автоволнами, а среда - активной.
Таким образом, распространение потенциала действия по нервному волокну происходит в форме автоволны. Активной средой являются возбудимые клетки.
Расчеты показывают, что скорость распространения возбуждения по гладким немиелинизированным нервным волокнам примерно пропорциональна квадратному корню из их радиуса (υ и л/Т). Сравнительно высокие скорости распространения потенциала действия у некоторых беспозвоночных (20-30 м/с) обеспечиваются большим диаметром их волокон (до 1 мм).
У позвоночных животных повышение скорости распространения возбуждения достигается миелинизацией волокон. Удельное сопротивление миелина больше, чем у других биологических мембран, толщина
миелиновой оболочки в сотни раз превышает толщину обычной клеточной мембраны. Скорость распространения пропорциональна и толщине, и удельному сопротивлению мембраны, поэтому она достаточно высока и у позвоночных животных.
Нарушение миелиновой оболочки приводит к нарушению распространения потенциала действия по нервному волокну и, как следствие, к тяжелым нервным заболеваниям.