Биохимия тканей и жидкостей полостей рта: учебное пособие / Вавилова Т.П. - 2-е изд., испр. и доп. - 2008. - 208 с. : ил.
|
|
|
|
ГЛАВА 8 ПОВЕРХНОСТНЫЕ ОБРАЗОВАНИЯ НА ЗУБАХ
8.1. КУТИКУЛА. ПЕЛЛИКУЛА. ЗУБНОЙ НАЛЁТ
В физиологических условиях на поверхности эмали образуются кутикула и пелликула.
Кутикула
Ку т и к ула покрывает поверхность эмали зубов после их прорезыва- ния в виде тонкой оболочки, состоящей из двух слоёв и обозначаемых как первичная и вторичная кутикула. Первичная кутикула - внутренний тонкий (около 0,5-1,5 мкм) гомогенный слой гликопротеинов, являющимся последним секреторным продуктом энамелобластов. Вторичная кутикула образована наружным (около 10 мкм) слоем редуцированного эпителия эмалевого органа.
После прорезывания зубов кутикула стирается и частично может сохраняться на апроксимальных поверхностях.
В отличие от кутикулы на протяжении всей жизни на поверхности эмали могут формироваться пелликула зуба и зубной налёт.
Пелликула
Пелликула - приобретенная тонкая органическая пленка, которая образуется из гликопротеинов слюны на поверхности зуба. Толщина пелликулы 1-4 мкм. Является бесструктурным образованием, не содержащим бактерий, и плотно фиксирована на поверхности зуба. На формирование приобретённой пелликулы влияет рН слюны. Пелликула формируется в течение 20-30 мин и её образование начинается с адсорбции специфических белков слюны на гидроксиапатитах эмали. Между поверхностью эмали и осаждающимися белками образуются ионные связи и гидрофобные взаимодействия.
В образовании пелликулы участвуют кислые и гликозилированные белки, богатые пролином, лактоферрин, лактопероксидаза, гистатин 1, цистатин SAIII и в более низких концентрациях - лизоцим и гистатин-5, а также аминосахара и моносахариды. Пелликула обладает избирательной проницаемостью и обеспечивает процессы диффузии ионов в поверхностный слой эмали, а также защищает эмаль зубов от воздействия химичес-
ких агентов. Зубная пелликула представляет собой барьер, через который регулируются процессы минерализации и деминерализации эмали, а также осуществляется контроль за составом микробной флоры, участвующей в образовании зубного налёта. После механической очистки пелликула восстанавливается на поверхности эмали в течение нескольких часов.
Все другие поверхностные образования на зубах, за исключением кутикулы и пелликулы, играют определённую роль в развитии стоматологической патологии.
Зубной налёт
Зубной налёт (зубная бляшка) - структура, состоящая из скопления различных видов микроорганизмов, погружённых в матрикс, образованный продуктами их жизнедеятельности, компонентами слюны и неорганическими веществами. Зубной налёт образуется путем осаждения микроорганизмов на поверхности пелликулы и растет за счет постоянного наслаивания новых видов бактерий. Определенную роль в формировании зубного налёта играют не только белки слюны и микроорганизмы, но и клетки слущенного эпителия.
Состав зубного налёта непостоянен и по мере его старения меняется.
Это зависит от состава микрофлоры и метаболических реакций, протекающих с участием микроорганизмов. По мере роста налёта начинает преобладать анаэробная флора, для которой характерно высокая ферментативная активность и образование органических кислот.
Зубной налёт на 78-80% состоит из воды. В сухом веществе определяются минеральные вещества, белки, углеводы.
Содержание макро- и микроэлементов в зубном налёте вариабельно. Известно, что на 1 мг сухой массы зубного налёта приходится около 3,4 мкг кальция, 8,4 мкг фосфора, 4,2 мкг калия и 1,3 мкг натрия. Кальций и фосфор в зубном налёте в основном поступает из слюны, хотя и не исключен выход этих элементов из эмали зубов. По мере созревания зубного налёта количество кальция и фосфора продолжает расти. Помимо макроэлементов в зубном налёте присутствуют микроэлементы, которые представлены ионами Ca2+, Sr2+, Fe3+, Mg3+, Mn3+, F- и др. Содержание фтора в зубном налёте может быть в десятки и даже в сотни раз больше, чем в слюне (6-180 мкг/г). Эта концентрация в значительной степени зависит от уровня фтора в воде. Включение фтора в зубной налёт происходит через образование фторапатита и СаF2 и связывание фтора с белками матрикса налёта. Фтор способен проникать внутрь бактерий.
Количество липидов в зубном налёте невелико и в раннем налёте определяются триацилглицеролы, холестерин, глицерофосфолипи- ды. По мере созревания налёта количество липидов не уменьшается, но они образуют комплексы с углеводами.
Белки составляют около 8% от сухой массы зубного налёта. Аминокислотный состав белков зубного налёта сходен, но не идентичен аминокислотному составу белков слюны и эпителиальных клеток. В зубном налёте также присутствуют свободные аминокислоты. По мере созревания зубного налёта аминокислотный состав меняется, исчезают глицин, аргинин, лизин и растет количество глутаминовой кислоты. Эти изменения обусловлены тем, что микроорганизмы способны расщеплять кислыми и слабощелочными протеиназами белки с освобождением пептидов и аминокислот, которые, в свою очередь, дезаминируются и декарбоксилируются. Увеличение количества глутаминовой кислоты связано с тем, что аминокислоты подвергаются трансаминированию.
Из свободных аминокислот в анаэробных условиях образуются продукты гниения: Н2S, аммиак, крезол, фенол, метилмеркаптан, индол, скатол, а также органические кислоты и низкомолекулярные летучие альдегиды, кетоны, придающие неприятный запах дыханию (галитоз).
В зубном налёте определяются свободная фруктоза, глюкоза, гексозамины, сиаловые кислоты, глюкозаминогликаны и гомополисахариды. Всего углеводов содержится 7-14 % от сухой массы.
В зубном налёте очень высока активность гликозидаз (а- и (3-глюкози- дазы, (3-галактозидаза, (3-глюкуронидаза, гиалуронидаза и β-N-ацетилгек- созаминидаза): она на порядок выше, чем в смешанной слюне. Осаждённые гликопротеины слюны под действием гликозидаз повергаются дегликозилированию, и освободившиеся свободные углеводы используются микроорганизмами для обеспечения энергетическх затрат. Поступающая с пищей сахароза гидролизуется бактериальной сахаразой до α-D-глюкозы и β-D-фруктозы. У бактерий глюкоза превращается в пировиноградную кислоту, которая у аэробных бактерий распадается до молекул СО2 и H2O, а у анаэробных микроорганизмов восстанавливается до молочной кислоты. Возможно также образование из пирувата уксусной и муравьиной кислот.
Лактатдегидрогеназа бактерий активируется высокими концентрациями фруктозо-1-6-бисфосфата, в то время как образование уксусной и муравьиной ингибируется большими концентрациями глицераль- дегид-3-фосфата. Этим объясняется активное образование молочной кислоты при избыточном поступлении углеводов, а при их недостатке пируват превращается в ацетат и муравьиную кислоту, но не в лактат.
Некоторые бактерии полости рта при избытке углеводов способны синтезировать гликоген. Механизм синтеза и распада гликогена бактериями подобен таковым у млекопитающих, но для синтеза гликогена используются молекулы не УДФ-глюкозы, а АДФ-глюкозы. Наиболее активны в этом отношении стрептококки. Так Str. mitis может синтезировать гликоген в количестве до 37% от своей сухой массы, который используется этими бактериями для жизнеобеспечения при отсутствии углеводов. Бактерии зубного налёта могут использовать сахарозу и для синтеза внеклеточных полисахаридов.
При участии бактериальных гликозилтрансфераз образуются липкие полисахариды (гликаны), которые адсорбируются на поверхности зуба и через гликан-связывающий белок связывают микроорганизмы. Гликаны в зубном налёте представлены леванами и декстранами.
Леван - полисахарид, состоящий из остатков фруктозы, связанных β (2-»6) - гликозидными связями и соединённый с молекулой сахарозы. Молекула декстрана - разветвлённый полисахарид, образованный остатками α-D-глюкозы, соединённых α (1-»6) и α (1-»3) - гликозидными связями (рис. 8.1).
Рис. 8.1. Структурная формула молекул: А - Левана; Б - Декстрана
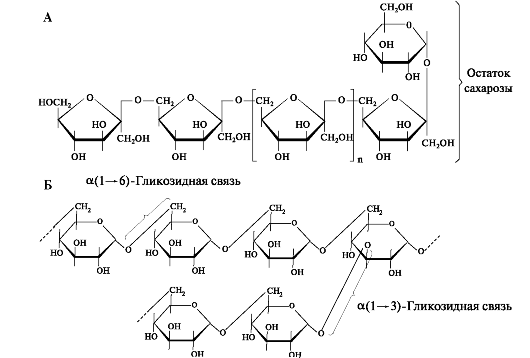
В синтезе леванов участвуют бактериальные фруктозилтрансферазы, а декстрана - глюкозилтрансферазы путём переноса остатков глюкозы от сахарозы. Молекулы декстрана достаточно долго сохраняются в зубном налёте, а молекулы левана легко растворимы и быстро гидролизуются леваназой некоторых стрептококков (рис. 8.2).
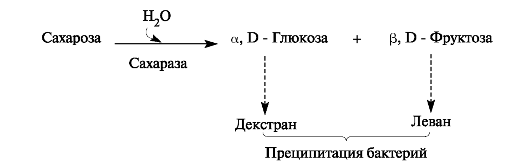
Рис. 8.2. Образование липких полисахаридов в зубном налёте.
Липкие полисахариды помогают бактериям занять определенное место в зубном налёте и обеспечивают их адгезию к эмали. Связь поверхности апатитов эмали с полисахаридами бактерий обеспечивают водородные связи, ионов Ca2+ и белки адгезины. К белкам адгезинам относится гликопротеин с мол. массой 200 кДа, который выделяется стрептококками.
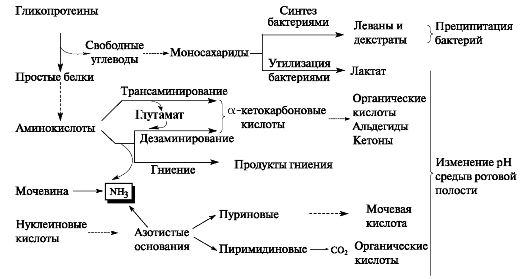
Рис. 8.3. Метаболические процессы в зубном налёте.
Зубной налёт отличается высокой метаболической активностью. В нём определяется активность свыше 50 различных ферментов, в основном бактериального происхождения. Помимо вышеуказанных гидролаз, трансфераз в зубном налёте присутствуют кислая и щелочная фосфатазы, РНК-азы и ДНК-азы, ферменты гликолиза, цикла трикарбоновых кислот, пероксидазы и другие энзимы. Активность всех ферментов возрастает при множественном кариесе и воспалении тканей пародонта (рис. 8.3).
8.2. ЗАМЕНИТЕЛИ САХАРОВ
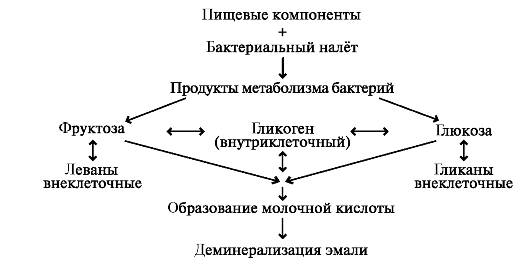
Накопление органических кислот, образующихся при распаде сахарозы, сопровождается локальным понижением рН. Накапливающиеся протоны H+ начинают замещать ионы Ca2+ в кристаллах гидроксиапатита эмали, что приводит к разрушению её минерального компонента. Результаты экспериментальных исследований показали, что протеолитические ферменты зубного налёта также способны разрушать органическую фракцию эмали с последующим освобождением фосфатов. Все эти реакции инициируют развитие кариозного процесса (рис. 8.4).
Рис. 8.4. Участие органических кислот в деминерализации эмали зуба.
Вместе с тем полагают, что роль протеиназ зубного налёта в развитии кариеса не столь велика, но более существенна в патогенезе гингивита и пародонтита, когда активность кислых и слабощелочных протеиназ в зубном налёте и десне возрастает в 4-5 раз, что
сопровождается деградацией гликопротеинов и других белков тканей пародонта. В расщеплении кислых гликозаминогликанов и гли- копротеинов межклеточного вещества и мембран клеток пародонта участвуют ферменты (3-глюкуронидаза, гиалуронидаза и β-N-ацети- лгексозаминидаза.
Полоскание полости рта 5% раствором глюкозы вызывает падение рН в течение 10-12 мин после процедуры, создавая локальные условия для деминерализации. На восстановление рН требуется более 1 ч. Динамика изменения рН у разных людей различна, что позволяет использовать этот показатель в качестве своеобразного теста для разделения людей на группы кариесрезистентных и кариесвосприимчивых.
В настоящее время выпускают большое количество разных веществ со сладким вкусом, что позволяет исключить из продуктов питания сахарозу (рис. 8.5).

Рис. 8.5. Распределение заменителей сахарозы по степени сладости.
Сорбитол - шестиатомный сахароспирт, который содержится в малых количествах во фруктах и синтезируется в печени. В слюне отсутствует сорбитолдегидрогеназа и сорбитол не метаболизируется в полости рта, поэтому его приём не вызывает значительного снижения рН в зубном налёте. В кишечнике сорбитол всасывается пассивно и не полностью. Невсосавшийся сорбитол (30%) оказывает осмотический послабляющий эффект. Максимально рекомендуемый прием сорбита составляет 150 мг/кг массы тела в день. Он используется как добавка в шоколад для больных сахарным диабетом и в жевательной резинке для профилактики кариеса зубов.
Ксилитол - пятиатомный сахароспирт. Входит в состав овощей и фруктов, а также его получают из коры деревьев. По сладости близок к сахарозе. Ксилитол подвергается незначительному превращению под действием бактерий зубного налёта, поэтому промывание полос-
ти рта ксилитом, в отличие от глюкозы, не вызывает существенного падения рН, а у людей, потребляющих продукты с добавкой ксилита, отмечена меньшая заболеваемость кариесом. Ксилитол приводит к лизису кариесогенных микроорганизмов. Попадая в микробную клетку, ксилитол фосфорилируется и ингибирует транспорт глюкозы и фруктозы. Нерегулируемое накопление ксилитол-5-фосфата в бактериальной клетке сопровождается осмотическим поступлением воды с последующим лизисом микроорганизмов. Подобно сорбиту ксилит вызывает осмотическую диарею.
В последнее время из плодов дикорастущих африканских растений выделены чрезвычайно сладкие на вкус белки - миракулин, монелин, тауматин. Они используются в жевательных резинках и зубных пастах.
Кроме белков в качестве заменителя сахарозы используют дипептиды - аспартам (метиловый эфир L-аспартил- L-фенилаланин), аспартил - аланинамиды. Они также не влияют на рН зубного налёта. При катаболизме аспартама отщепляется метиловый спирт и освобождается аспарагиновая кислота и фенилаланин.
В настоящее время достаточно часто в качестве подсластителя применяют сахарин - амид ортосульфобензойная кислота. Он достаточно быстро всасывается в пищеварительном тракте и 98% его выделяется с мочой. Он является слабым канцерогеном и вызывает рак мочевого пузыря у крыс.
Цикламаты - производные амино-N-сульфоновой кислоты. Часть цикламата (до 1%) в организме превращается в токсичное соединение циклогиксиламин или канцерогенный дициклогиксиламин, который вызывает развитие рака мочевого пузыря.
На устойчивость зубов к кариесу оказывают влияние микроэлементы. Так, фтор участвует в образовании кислотоустойчивых фторапатитов и ингибирует рост бактерий. Поступающие молибден и ванадий повышают кариесрезистентность, в то время, как селен, напротив её снижает.
Отложение минералов в зубном налёте приводит к формированию зубного камня.
8.3. ЗУБНОЙ КАМЕНЬ
Кристаллы фосфата кальция откладываются внутри зубного налё- та и тесно связываются с поверхностью эмали. Процесс отложения
неорганических веществ в зубном налёте занимает около 12 сут и после минерализации камень уже не так легко удаляется механическим воздействием или током слюны. Бактерии продолжают накапливаться на поверхности образующегося зубного камня, способствуя его росту. В зависимости от расположения на поверхности зуба различают над- и поддесневой зубной камень. По своему составу над- и поддесневой зубные камни сходны, однако они различаются по источникам поступления фосфорно-кальциевых соединений. В наддесневой зубной камень они поступают из слюны, а в поддесневой зубной камень - из десневой жидкости.
Химический состав зубного камня. Большая часть зубного камня (70-90% сухого остатка) представлена неорганическими веществами. В зубном камне определяется 29-57% кальция и 16-29% неорганического фосфата и около 0,5% магния. В следовых количествах присутствует свинец, молибден, кремний, алюминий, стронций, кадмий, фтор и других химические элементы.
Кальций и фосфор осаждаются на органической матрице в виде солей. На начальных этапах в основном образуется брушит [СаНРО4?2Н2О], который составляет до 50% от всех видов кристаллов. Кристаллы брушита имеют клиновидную форму. Их накопление приводит к формированию слабоминерализованного, легко удаляемого зубного камня. Помимо брушита образуются другие виды кристаллов. Это витлокит, монетит, октокальций фосфат, гидроксиапатит и другие апатиты. Витлокит представляет собой кристаллы, имеющие форму ромба. В их структуре определяются безводный фосфат кальция [Ca3 PO4]2 и ионы Mg2+, Mn2+, Fe3+. Монетит - вторичная соль фосфорной кислоты [CaHPO4], которая кристаллизуется в виде треугольных пластинок. Растворимость кристаллов монетита быстро увеличивается при кислых значениях рН-среды. Промежуточным связующим звеном между кислыми солями - монетитом и брушитом и основной солью - гидроксиапатитом является октокальций фосфат - [Ca8(HPO4)2(PO4)4?5H2O]. Он напоминает кристаллы гидроксиапатита, но имеет слоистую структуру с чередованием слоёв соли толщиной 1,1 нм и слоёв воды толщиной 0,8 нм. Кристаллы октакальция фосфата растут в форме тонких пластинок длиной до 250 мкм. Они содержат кислый фосфатный ион и не имеют гидроксильных групп. Октакальций фосфат играет важную роль в нуклеации апатитных солей и подобно монетиту и брушиту при щелочных значениях рНсреды превращается в гидроксиапатит.
В составе зубного камня фтор присутствует в виде фторапатита, фторида кальция [СаF2] и комплекса с органическими соединениями в составе бактерий. Низкие концентрации фтора (20-100 мкг/л) в зубном камне ускоряют скорость образования апатитов. Применение фторсодержащих паст для чистки зубов уже в первые 10 сут приводит к накоплению фтора в составе зубного камня.
Неорганические вещества в зубном камне связываются с белками, количество которых составляет 0,1-2,5% и зависит от вида зубного камня. Наибольшее количество белка (2,5%) присутствует в светлом наддесневом зубном камне. В тёмном наддесневом камне содержание белка снижается до 0,5%, а в поддесневом зубном камне оно составляет всего 0,1-0,3%. Помимо белков в зубном камне обнаруживают различные аминокислоты. В наибольшем количестве выявляются остатки аланина, лейцина, глицина, глутаминовой и аспарагиновой кислот, а также пролина, лизина, серина и треонина. Глутамат и аспартат способны связывать ионы Ca2+, а остатки серина, треонина и лизина - PO43-, что очень важно для инициации минерализации зубного налёта и дальнейшего образования зубного камня.
В зубном камне также присутствуют до 10% углеводов, в том числе гликозаминогликаны, галактоза, фруктоза, манноза и аминосахара. Гликозаминогликаны в основном встречаются в поддесневом зубном камне, и их источником, по всей вероятности, является деградированные протеогликаны межклеточного вещества тканей пародонта.
Содержание липидов в зубном камне невелико. Присутствующие глицерофосфолипиды, освобождаемые при распаде клеточных мемб- ран микроорганизмов, связывают ионы Ca2+ и инициируют образование фосфорно-кальциевых солей, а в дальнейшем гидроксиапатитов.
В образовании зубного камня участвуют молекулы АТФ, которые, с одной стороны, необходимы для фосфорилирования органических соединений, а с другой - при гидролизе АТФ освобождается ортофос- форная кислота, являющаяся компонентом различных минеральных кристаллов.
Механизм образования зубного камня. Для формирования зубного камня необходимы:
• образование органической матрицы (зубного налёта) микроорганизмами ротовой полости;
• отложение минеральных соединений на органической матрице с образованием центров кристаллизации;
• рост кристаллов в центрах минерализации.
Первичный слой бактерий, фиксированных на поверхности зуба, представлен в основном Str. sanguis, а на поверхности эпителиальных клеток слизистой оболочки десны Str. salivarium. На них оседают другие бактерии, образуется связь между множеством клеток, то есть возникают микробные ассоциации. По мере увеличения анаэробных бактерий освобождается большое количество молочной кислоты, которая поглощается и метаболизируется вейлонеллами. В этих участках происходит повышение значения рН-среды. Увеличение рН также способствует уменьшение количества молекул СО2 и накопление в зубном налёте аммиака. Аммиак освобождается из мочевины при участии уреолитических бактерий, обладающих уреазной активностью. Кроме того, микоплазмы гидролизуют аргинин, что также увеличивает количество аммиака. В реакциях трансаминирования α-кетокислоты превращаются в аспартат и глутамат. Выделившийся аммиак и образующиеся дикарбоновые кислоты активно соединяются с ионами PO43- , Mg2+, Ca2+ и формируются центры кристаллизации. Отложению фосфата способствует и изменение мицеллярной структуры слюны, когда фосфат кальция выпадает в осадок. Это возможно при изменении количества специфических белков слюны, а также количества пирофосфата. Статерины и пирофосфат являются ингибиторами образования зубного камня.
Таким образом, на поверхности и внутри бактерий образуется первичный преципитат, состоящий из брушита, невберита и струвита. Первичный преципитат затем трансформируется в пластинчатый гидроксиапатит, а в дальнейшем в гексагональный гидроксиапатит. Накопление фосфорно-кальциевых соединений и их трансформация протекает при участии щелочной фосфатазы и использовании молекул АТФ. Кристаллы зубного камня продолжают расти при послойном отложении минеральных солей и в тёмном зубном камне образуются сферолиты.