Биохимия тканей и жидкостей полостей рта: учебное пособие / Вавилова Т.П. - 2-е изд., испр. и доп. - 2008. - 208 с. : ил.
|
|
|
|
ГЛАВА 7 СЛИЗИСТАЯ ОБОЛОЧКА ПОЛОСТИ РТА
Слизистая оболочка полости рта покрывает внутреннюю поверхность щёк, губ, альвеолярные отростки, твёрдое и мягкое нёбо, язык, дно полости рта и постоянно увлажняется секретом слюнных желёз. Она обладает рядом характерных особенностей и имеет неоднородное строение. Это разнообразие позволяет слизистой оболочке осуществлять множество функций.
7.1. СТРОЕНИЕ И ФУНКЦИИ СЛИЗИСТОЙ ОБОЛОЧКИ ПОЛОСТИ РТА
Слизистая оболочка полости рта представлена двумя слоями: эпителием и собственной пластинкой, образованной соединительной тканью, между которыми располагается базальная мембрана.
В собственной пластинке слизистой оболочки полости рта имеются три основных типа волокон: коллагеновые, ретикулярные и эластические. Коллагеновые волокна представлены в основном коллагенами III, IV, V и VI типов. С фибриллобразующим коллагеном V типа связаны коллаген VI типа, протеогликаны, гликозаминогликаны и гликопротеины. Коллаген IV типа участвует в формировании базальной мембраны слизистой оболочки. Коллаген III типа характерен для тканей, содержащих большое количество кровеносных сосудов. Ретикулярные волокна слизистой оболочки представлены коллагеном III типа, которые обеспечивают упругость слизистой. В межкле- точном веществе тканей периодонта, корня языка, в подслизистом слое губ и щёк в больших количествах присутствует эластиновые волокна, придающие устойчивость к сжатию и растяжению.
Базальная мембрана. На ультраструктурном уровне в базальной мембране выявляется светлый мелкозернистый слой, прилежащий к внешней клеточной мембране эпителиоцитов базального слоя (светлая пластинка), а также более глубоко лежащий слой, образованный мелкозернистым или фибриллярным материалом (плотная пластинка) (рис. 7.1).
Светлая пластинка образована гликопротеинами, в том числе ламинином и протеогликанами, содержащими гепарансульфаты.
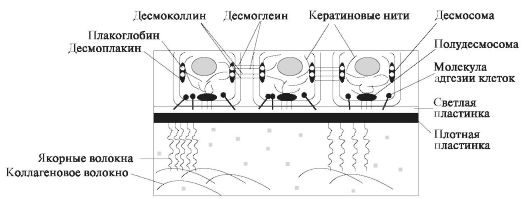
Рис. 7.1. Ультраструктурная организация слизистой оболочки полости рта.
• Плотная пластинка содержит коллаген IV типа и белок энтактин, через который осуществляется связь с белком ламинином. Якорные фибриллы построены из коллагена VII типа, а связанные с ними фибриллы - из коллагенов I и III типов. В составе базальной мембраны определяется адгезивный гликопротеин фибронектин. Базальная мембрана слизистой оболочки полости рта: обеспечивает прочную связь эпителия с подлежащей соединительной тканью: к ней с помощью полудесмосом с одной стороны прикрепляются базальные клетки эпителия, а с другой стороны посредством якорных фибрилл - коллагеновые волокна; •способствует дифференцировке эпителия и сохраняет его архитектонику. •осуществляет избирательное поступление веществ в эпителий. Базальная мембрана может задерживать ряд молекул с высокой мол. массой, например: комплексы антиген-антитело. Помимо описанных функций для клеток слизистой оболочки полости рта характерна высокая способность к регенерация и тесное взаимодействие эпителиальных клеток как между собой, так и с внеклеточным матриксом.
В межклеточном веществе определяются сложные надмолекулярные комплексы, состоящие из протеогликанов, гликозаминогликанов и гликопротеинов. Протеогликаны слизистой оболочки, представленные декорином, бигликаном, версиканом и синдеканом-1, связаны с дерматансульфатом (60%), гепарансульфатом и хондроитин-4-суль- фатом (10%). Гиалуроновая кислота, которая содержится в достаточно большом количестве (30%), связывает воду, ионы и обеспечивает тургор слизистой, а также выполняет защитную функцию. В формиро-
вании надмолекулярных комплексов и связывании клеток участвуют фибронектин и другие гликопротеины основного вещества.
Эпителий слизистой оболочки полости рта представлен несколькими слоями (базальный, шиповатый, зернистый и роговой), и к базальной мембране присоединяются клетки базального слоя.
Помимо эпителиальных в слизистой оболочке присутствуют и неэпителиальные клетки, которые постоянно взаимодействуют друг с другом и образуют единую систему связанных элементов. Деятельность клеток каждого типа регулируется различными факторами. Так эпителиоциты синтезируют ИЛ-1 и ИЛ-6, фактор некроза опухолей, колониестимулирующий фактор роста, фактор роста эпителия, ТФР-(3. ИЛ-1, синтезируемый эпителиоцитами, в свою очередь, активирует синтез Т-лимфоцитов, те, в свою очередь, секретируют ИЛ-2. Взаимодействие клеток через цитокины обеспечивает иммунную реакцию в ответ на повреждение целостности слизистой оболочки. Цитокины эпителиального происхождения также воздействуют на рост и дифференцировку фибробластоподобных клеток, участвующих в регенерации эпителия.
В эпителии слизистой оболочки полости рта клетки разных слоёв постоянно взаимодействуют. Клетки базального и шиповатого слоёв связаны с базальной мембраной с помощью полудесмосом, а между собой десмосомами. Десмосомы соединяют клеточные мембраны с промежуточными филаментами цитоскелета и формируют непрерывную сеть, которая пронизывает всю ткань и обеспечивает значительную устойчивость тканей к растяжению, а полудесмосомы облегчают взаимодействие клетки с внеклеточным матриксом.
Десмосомы - сложноорганизованная специализированная структура клеточной адгезии, которая реализуется через специальные адгезивные молекулы - гликопротеины. Десмосома представлена в виде двух форм соединений. Одна из них - цитоплазматическая пластинка - осуществляет связь промежуточных филаментов клетки с плазматической мембраной. Вторая форма связывает плазматическую мембрану с внеклеточными межмембранными молекулами. Функцию десмосом обеспечивают кальций-связывающие белки - плакоглобины, десмоплакины, десмоколлины, десмоглеины, которые относятся к семейству кадгеринов. Эти клетки оказывают специфическое воздействие друг на друга (рис. 7.2).
Кроме того, между эпителиоцитами имеются щелевые контакты, а также плотные соединения.
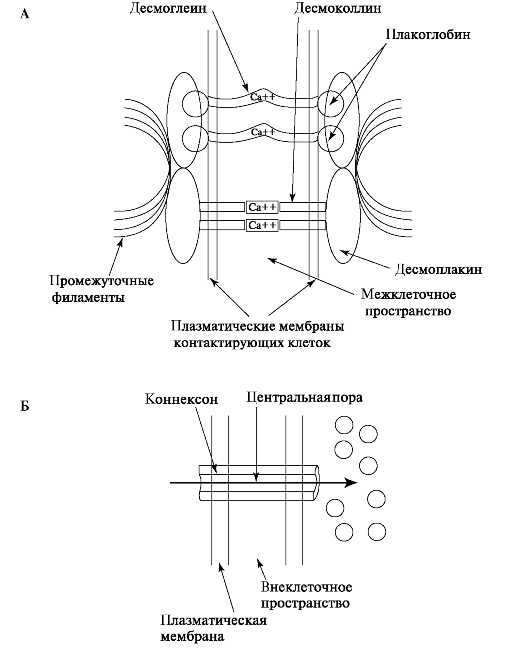
Рис. 7.2. Виды контактов в эпителии слизистой оболочки полости рта:
А - Строение и организация десмосом; Б - Схематичное изображение
щелевого контакта, связывающих клетки в эпителиальном слое слизистой
Щелевые контакты - скопление мембранных каналов (коннексоны), соединяющих содержимое соседних клеток в тканях (то есть эти каналы соединяют две соседние плазматические мембраны). Каналы сформированы обширной группой родственных белков и обычно находятся в открытом состоянии. Они закрываются, когда снижается скорость метаболизма. Сигналом для закрытия канала является повышение концентрации ионов Ca2+, изменение трансмембранного потенциала, закисление среды, а также фосфорилирование белков. Каждый канал представляет собой гексамерную структуру с центральной порой и состоит из 12 субъединиц, по 6 от каждой клетки. Каждая субъединица имеет полый стержень, пронизывающий бислой (рис. 7.3). В присутствии ионов Ca2+ субъединицы располагаются параллельно центральной оси канала, а в отсутствии этих ионов они несколько наклонены и переходят в открытое состояние. По щелевым соединениям из одной клетки в другую могут поступать неорганические ионы и большинство метаболитов - моносахариды, аминокислоты, нуклеотиды.
Вместе с тем белки, нуклеиновые кислоты и полисахариды больших размеров не проходят по этим каналам. Таким образом, через щелевые соединения и десмосомы клетки слизистой оболочки объединены в единую систему и таким путём достигается быстрый и синхронный ответ на стимуляцию. Через щелевые контакты происходит также питание клеток, удалённых от кровеносных сосудов.
Исследования обменных процессов в клетках слизистой оболочки полости рта показали высокую активность окислительно-восстановительных ферментов, а источником АТФ является окислительное фосфорилирование. В клетках определяется низкая активность кис-
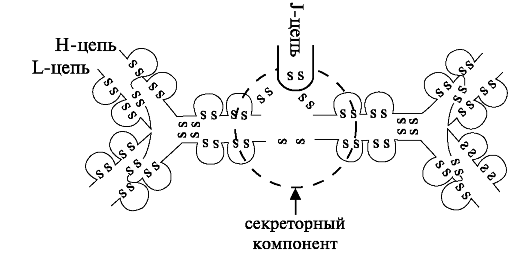
Рис. 7.3. Строение молекулы секреторного иммуноглобулина (SIgA).
лых и слабощелочных протеиназ и гликозидаз, что, по-видимому, способствует сохранению целостности слизистой оболочки.
Для неороговевающего эпителия ротовой полости человека характерна способность к синтезу и накоплению большого количества гликогена, преимущественно в цитоплазме клеток шиповатого слоя. Базальный слой эпителия не содержит гликогена. В эпителии твёрдо- го нёба и дёсен гликоген в норме отсутствует и появляется при патологических процессах.
Базальный слой. Среди клеток базального слоя имеются стволовые клетки - родоначальницы пролиферативных единиц эпидермиса. Митотическая активность базальных клеток зависит от толщины эпителиального пласта и контролируется гормонами и факторами роста. Деление кератиноцитов стимулируют эпидермальный фактор роста и фактор роста кератиноцитов. Аналогичным действием обладает и вещество P, выделяющееся из терминалей чувствительных нервных волокон. В этом слое клеток происходит синтез фибриллярных белков промежуточных филаментов цитоскелета - кератинов. Особенностью кератиновых белков является присутствие в их составе 10-12% остатков цистеина, защищающих клетки слизистой оболочки от воздействия АФК. Кератиновые белки формируют кератиновые филаменты, которые обеспечивают связывание клеток с базальной мембраной с помощью полудесмосом. В базальном слое эпителия располагаются клетки Меркеля, связанные с афферентным нервным волокном, и осуществляющие рецепторную функцию. Особенностью клеток Меркеля является наличие гранул, содержащих нейромедиаторы, которые при деформации отростков выделяются в синаптическую щель. В качестве медиаторов выступают ВИП, пептид гистидин-изолейцин, вещество P, пептид связанный с кальцитониновым геном и др.
Клетки следующего слоя - шиповатые - содержат многочисленные отростки, которые соединяются между собой десмосомами. В отростках содержатся пучки тонофиламентов, которые представля- ют собой скопление белков, формирующих фибриллярные структуры, связанные с десмосомами. Для клеток этого слоя характерна высокая метаболическая и митотическая активность. В шиповатых клетках, расположеных ближе к поверхности эпидермиса, имеются гранулы, содержащие липиды и белки. Содержимое этих гранул освобождается в межклеточное пространство и скрепляет клетки между собой.
Зернистые клетки сохраняют способность к синтезу белков, который ингибируется с переходом клеток в роговой слой. Они содержат
пучки промежуточных филаментов и два вида гранул: в гранулах I типа (кератиносомы) определяются гидролитические ферменты и липиды, которые выделяются в межклеточное вещество, где они образуют водонепроницаемый барьер, а гранулы II типа сформированы кератогиалином. Гранулы II типа не окружены мембраной и представляют собой скопления кератиновых белков (цитокератинов), соединённых с белками, богатыми гистидином и цистеином. В состав гранул кератогиалина входят два специфических белка - лорикрин и инволюкрин, для которых характерно малое количество пролина. Эти белки при участии фермента трансглутаминазы связывают эпителиальные клетки с базальной мембраной. В слое клеток также синтезируются белки-маркёры эпителия филлагрин и трихогиалин. Совместно с кальцием и ретиноевой кислотой путём регуляции активности различных протеиназ и протеинкиназ они участвуют в процессах дифференцировки клеток. Таким способом обеспечивается целостность слизистой оболочки.
Определение характера синтезируемых цитокератинов имеет опре- делённое диагностическое значение для прогнозирования развития опухолевых процессов.
Роговой слой представлен плотно упакованными роговыми чешуйками, имеющими форму 14-гранной фигуры. Они постоянно слущиваются в наружных отделах слоя вследствие разрушения десмосом и не участвуют в обменных процессах. Этот слой хорошо выражен на поверхности десны, щеки по линии смыкания зубов и твёрдого нёба. Ороговение эпителия на этих поверхностях следует рассматривать как защитную реакцию на механическое раздражение, которое испытывает слизистая в процессе жевания. Для слущиваемых клеток характерно высокое содержание белка, в том числе катионных белков, инволюкрина и кератолеина, участвующих в реакциях защиты, а также активность эстераз и кислой фосфатазы. Эти белки защищают от разрушения плазмолемму гидролитическими ферментами, освобождаемых из кератиносом и лизосом.
Слизистая оболочка выполняет множество функций
• Защитная функция: защищает подлежащие ткани от повреждающего действия механических сил и стирания, которые возникают при откусывании и пережевывании пищи. Эпителий слизистой оболочки защищает также от химических воздействий и препятствует внедрению микроорганизмов. Устойчивость к неблагоприятным факторам, в частности, связана с десквамацией (слущиванием) эпителия, которая компенсируется активной его регенерацией.
Важным фактором, способствующим поддержанию барьерных свойств эпителия, служит постоянное смачивание его слюной, которая содержит ряд биологически активных веществ, влияющих на скорость дифференцировки и пролиферации эпителиальных клеток.
Во всех отделах слизистой оболочки полости рта, за исключением десны и передней части твёрдого нёба, рассеяны мелкие слюнные железы, в строме которых выявляют лимфоциты, тучные клетки, макрофа- ги и плазматические клетки, секретирующие преимущественно IgА. Последний синтезируется в плазматических клетках, находящихся в анатомической связи с эпителием слизистых оболочек и клетками слюнных желёз. Эпителиальные клетки концевых отделов слюнных желёз и выводных протоков синтезируют секреторный компонент - гликопротеин, который обеспечивает захват и трансэпителиальный перенос иммуноглобулинов в слюну. Этот иммуноглобулин называется секреторным IgА (SIgA). Структура его представлена на рис. 7.3.
SIgA отличается от сывороточного наличием секреторного компонента, связанного с двумя или тремя мономерами IgА. Синтезированный в плазматических клетках димер IgA, состоящий из 4Н, 4L, связанных J-цепью, покидает плазматические клетки и на плазматической мембране ацинарных клеток или клеток выводных протоков связывается с рецептором - гликопротеином с мол. массой ~71 кДа и поглощается клеткой. Далее димер IgA соединяется с другим гликопротеином (секреторным компонентом) с мол. массой ~80 кДа. Образовавшийся комплекс димер - SIgA перемещается к апикальной части клеток слюнных желёз и путем пиноцитоза поступает в слюнные протоки (рис. 7.4).
Секреторный компонент SIgA защищает молекулу антитела от разрушения ферментами различных клеток, а также повышает её устойчивость к воздействию денатурирующих факторов. По своей активности SIgA превосходит все другие иммуноглобулины. Свое антибактериальное действие он оказывает связываясь с лизоцимом. SIgA препятствует адгезии микроорганизмов на слизистых оболочках полости рта, а также адсорбции и репродукции вирусов в эпителиальных клетках.
Эпителиоциты также синтезируют белок кальпротектин, который обладает мощным антимикробным действием. Все эти вышеперечисленные белки принимают также активное участие в процессах регене- рации слизистой оболочки полости рта при повреждении.
• Сенсорная функция. На слизистой оболочке полости рта располагаются температурные, тактильные и болевые рецепторы.
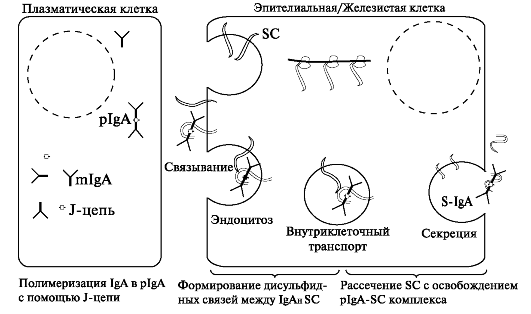
Рис. 7.4. Модель селективного транспорта IgA через эпителиальную/железис- тую клетку.
Восприятие вкуса осуществляется благодаря специализированным вкусовым рецепторам.
• Секреторная функция. В толще слизистой оболочки располагаются малые слюнные железы, а в некоторых участках - сальные железы.
• Иммунная функция участвует в обеспечении местного иммунитета. Это связано с тем, что она содержит клетки Лангерганса, макрофаги, лимфоциты, плазматические клетки, которые участвуют в различных звеньях иммунных реакций.
• Всасывательная функция. Слизистая оболочка полости рта на всей своей поверхности обладает большей проницаемостью, чем эпителий кожи. В области дна ротовой полости слизистая оболочка проницаема для ряда веществ, в частности, ионов I-, K+, Na+, некоторых аминокислот, лекарственных препаратов.