Анатомия человека : Сапин М. Р., Билич Г. Л. учебник в 3 т. - изд. 3-е испр. 2007. - Т. 1, Т. 2, Т. 3.
|
|
|
|
В процессе изучения тела человека его структуры подразделяют на клетки, ткани, органы, системы и аппараты органов, которые и формируют организм.
КЛЕТКИ
Клетка является основной структурной единицей строения живого. В середине XIX в. Т. Шванн создал клеточную теорию. Ее основные положения свидетельствовали о том, что все ткани состоят из клеток, а клетки животных и растений принципиально сходны между собой; деятельность организмов - это итог жизнедеятельности отдельных клеток. Большое влияние на дальнейшее развитие клеточной теории оказал Р. Вирхов. Он не только свел воедино многочисленные разрозненные факты, но и убедительно показал, что клетки являются постоянной структурой и образуются только путем размножения, порождая себе подобных - «каждая клетка их клетки».
Клетка рассматривается как элементарная единица всего живого, потому что ей присущи все свойства живых организмов: высокоупорядоченное строение, получение энергии извне и ее использование для выполнения работы и поддержания упорядоченности (преодоление энтропии), обмен веществ, рост, размножение, передача биологической информации потомкам, активная реакция на раздражения, регенерация, адаптация к окружающей среде.
Клеточная теория в современной интерпретации включает следу- ющие главные положения:
- клетка является универсальной элементарной единицей живого;
- клетки всех организмов принципиально сходны по строению, функции и химическому составу;
- клетки хранят, перерабатывают и реализуют генетическую информацию;
- клетки размножаются только путем деления исходной клетки;
- многоклеточные организмы являются сложными клеточными ансамблями, образующими целостные системы;
- благодаря деятельности клеток в сложных организмах осуществляются рост, развитие, обмен веществ и энергии.
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
В состав веществ, участвующих в реакциях, связанных с жизнедеятельностью клетки, входят почти все известные химические элементы, причем на долю четырех из них приходится 98% массы клетки. Это кис- лород (65-75%), углерод (15-18%), водород (8-10%) и азот (1,5-3%). Остальные элементы подразделяются на макроэлементы (около 1,9%) и микроэлементы (около 0,1%). К макроэлементам относятся сера, фосфор, хлор, калий, натрий, магний, кальций и железо, к микроэлементам - цинк, медь, йод, фтор, марганец, селен, кобальт и др. Несмотря на очень малое содержание, микроэлементы играют важную роль. Они влияют на обмен веществ, без них невозможна нормальная жизнедеятельность каждой клетки в отдельности и организма как целого.
Клетка состоит из неорганических и органических веществ. Среди неорганических преобладает вода, ее относительное количество в клетке от 70 до 80%. Вода - универсальный растворитель, в ней происходят все биохимические реакции, при участии воды осуществляется тепло- регуляция клетки. Вещества, растворимые в воде (соли, основания, кислоты, белки, углеводы, спирты и др.), называются гидрофильными. Гидрофобные вещества (жиры и жироподобные вещества) не растворяются в воде. Есть органические вещества с вытянутыми молекулами, у которых один конец гидрофилен, другой гидрофобен; их называют амфипатическими. Примером амфипатических веществ могут служить фосфолипиды, участвующие в образовании биологических мембран.
Неорганические вещества (соли, кислоты, основания, положительные и отрицательные ионы) составляют от 1,0 до 1,5% массы клетки. Среди органических веществ преобладают белки (10-20%), жиры, или липиды (1-5%), углеводы (0,2-2,0%), нуклеиновые кислоты (1-2%). Содержание низкомолекулярных веществ в клетке не превышает 0,5%.
Молекула белка является полимером, который состоит из множества повторяющихся единиц (мономеров). Мономеры белка - аминокислоты
(их 20) соединены между собой пептидными связями, образующими полипептидную цепь (первичную структуру белка). Цепь закручивается в спираль, представляющую, в свою очередь, вторичную структуру белка.
Белки выполняют важнейшие функции. Ферменты - биологические катализаторы, увеличивающие скорость химических реакций в клетке в сотни тысяч - миллионы раз, являются белками. Белки, входя в состав всех клеточных структур, выполняют пластическую (строительную) функцию. Они образуют клеточный скелет. Движения клеток также осуществляют специальные белки (актин, миозин, динеин). Белки обеспечивают транспорт веществ в клетку, из клетки и внутри клетки. Антитела, которые наряду с регуляторными выполняют и защитные функции, также являются белками. Наконец, белки являются одним из источников энергии.
Углеводы подразделяются на моносахариды и полисахариды. Полиса- хариды, подобно белкам, построены из мономеров - моносахаридов. Среди моносахаридов в клетке наиболее важны глюкоза (содержит 6 атомов углерода) и пентоза (5 атомов углерода). Пентозы входят в состав нуклеиновых кислот. Моносахариды хорошо растворяются в воде, полисахариды - плохо. В животных клетках полисахариды представлены гликогеном. Углеводы являются источником энергии. Сложные углеводы, соединенные с белками (гликопротеины) и/или жирами (гликолипиды), участвуют в образовании клеточных поверхностей и взаимодействиях клеток.
К липидам относятся
жиры и жироподобные вещества. Молекулы жиров построены из глицерина и
жирных кислот. К жироподобным веществам относятся холестерин, некоторые
гормоны, лецитин. Липиды, являющиеся основным компонентом клеточных
мембран (они описаны ниже), выполняют тем самым строительную функцию.
Они являются важнейшим источником энергии. Так, если при полном
окислении
Нуклеиновые кислоты являются полимерными молекулами, образованными мономерами - нуклеотидами, каждый из которых состоит из пуринового или пиримидинового основания, сахара пентозы и остатка фосфорной кислоты. Во всех клетках существует два типа нуклеиновых кислот: дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК), которые различаются по составу оснований и сахаров (табл. 1).
Молекула ДНК состоит из двух разнонаправленных полинуклеотидньгх цепей, закрученных одна вокруг другой в виде двойной спирали. Основания расположены внутри двойной спирали, а сахарофосфатный скелет -
Таблица 1. Состав нуклеиновых кислот
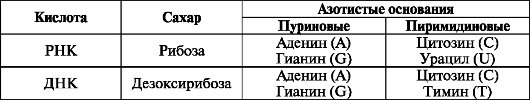
снаружи. Азотистые основания обеих цепей соединены между собой комплементарно водородными связями, при этом аденин соединяется только с тимином, а цитозин с гуанином (А=Т, G=C). Количество гуанина всегда равно количеству цитозина, количество тимина - количеству гуанина. На один виток спирали длиной 3,4 нм приходится 10 пар азотистых оснований. ДНК несет в себе генетическую информацию, закодированную последовательностью азотистых оснований. Она определяет специфичность синтезируемых клеткой белков, т. е. последовательность аминокислот в полипептидной цепи. Вместе с ДНК дочерним клеткам передается генетическая информация, определяющая (во взаимодействии с условиями среды) все свойства клетки. ДНК содержится в ядре и митохондриях. Молекула РНК образована одной полинуклеотидной цепью.
Организм человека состоит примерно из 6*1013
клеток (Дубинин Н.П., 1963). Размеры клеток человека варьируют от
нескольких микрометров (например, малые лимфоциты - около 7 мкм) до 200
мкм (яйцеклетка). Напомним, что 1 микрометр (мкм) = 10-6 м; 1 нанометр (нм) = 10-
Основными функциональными структурами клетки являются ее поверхностный комплекс, цитоплазма и ядро. Поверхностный комплекс включает в себя гликокаликс, клеточную мембрану (цитолемму)и кортикальный слой цитоплазмы. Нетрудно увидеть, что резкого отграничения поверхностного комплекса от цитоплазмы нет. В цитоплазме выделяют гиалоплазму (матрикс, цитозоль), органеллы и включения (табл. 2, рис. 1).
Структурными компонентами ядра являются кариолемма (кариотека), кариоплазма ихромосомы;петли некоторых хромосом могут переплетаться, и в этой области образуется ядрышко.
Цитолемма, кариолемма и часть органелл образованы биологическими мембранами.
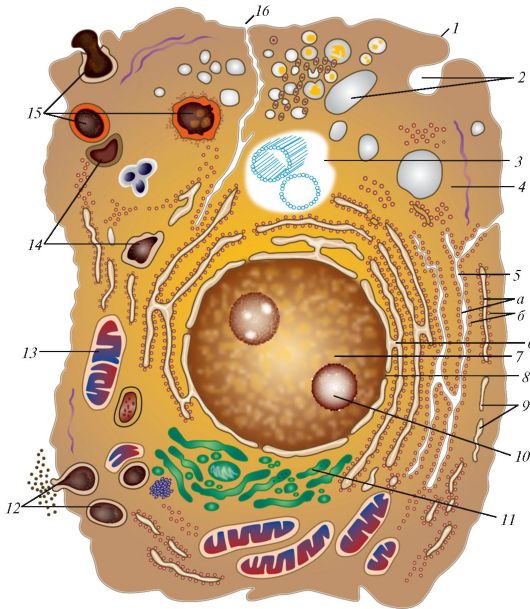
Рис. 1. Ультрамикроскопическое строение клетки: 1 - цитолемма (цитоплазматическая мембрана); 2 - пиноцитозные пузырьки; 3 - центросома (клеточный центр; цитоцентр); 4 - гиалоплазма; 5 - зернистая эндоплазматическая сеть: а - мембрана зернистой сети, б - рибосомы; 6 - связь перинуклеарного пространства с полостями эндоплазматической сети; 7 - ядро; 8 - ядерная пора; 9 - незернистая (гладкая) эндоплазматическая сеть; 10 - ядрышко; 11 - внутренний сетчатый аппарат (комплекс Гольджи); 12 - секреторные вакуоли; 13 - митохондрия; 14 - лизосомы; 15 - три последовательные стадии фагоцитоза; 16 - связь клеточной оболочки (цитолеммы) с мембранами
эндоплазматической сети
Таблица 2. Структурные компоненты клетки
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Мембрана состоит из двух слоев амфипатических молекул липидов (билипидный слой, или бислой). Каждая такая молекула имеет две части - головку и хвост. Хвосты гидрофобны и обращены друг к другу. Го- ловки, напротив, гидрофильны и направлены кнаружи и внутрь клетки. В билипидный слой погружены молекулы белка (рис. 2). Билипидный слой ведет себя как жидкость, обладающая значительным поверхностным натяжением. Вследствие этого он образует замкнутые полости, которые не спадаются.
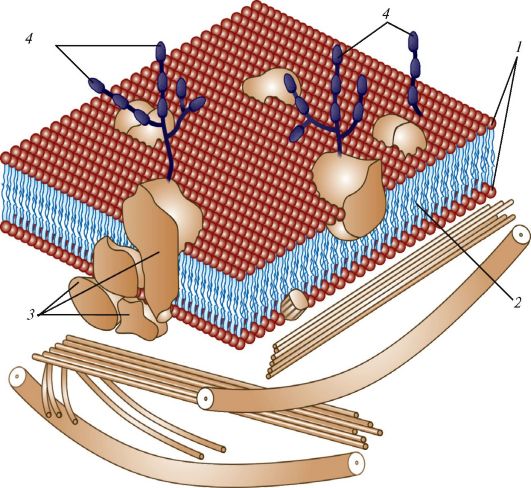
Рис. 2. Строение цитоплазматической мембраны: 1 - липиды; 2 - гидрофобная зона липидных молекул; 3 - белковые молекулы; 4 - полисахариды гликокаликса
Некоторые белки проходят через всю толщу мембраны, так что один конец молекулы обращен в пространство по одну сторону мембраны, другой - по другую. Их называют интегральными (трансмембранными). Другие белки расположены так, что в околомембранное пространство обращен лишь один конец молекулы, а второй конец лежит во внутреннем или в наружном монослое мембраны. Такие белки называют полуинтегральными. Некоторые белки (обычно переносимые через мембрану и временно находящиеся в ней) могут лежать между фосфолипидными слоями.
Концы белковых молекул, обращенные в околомембранное пространство, могут связываться с различными веществами, находящимися в этом пространстве, поэтому интегральные белки играют большую роль в организации трансмембранных процессов. С полуинтегральными белками всегда связаны молекулы, осуществляющие реакции по восприятию сигналов из среды (молекулярные рецепторы) или по передаче сигналов от мембраны в среду. Подобно липидам, белковые молекулы также являются амфипатическими - их гидрофобные участки окружены аналогичными «хвостами» липидов, а гидрофильные обращены наружу или внутрь клетки.
Белки осуществляют большую часть мембранных функций: одни являются рецепторами, другие - ферментами, третьи - переносчиками. Несколько белковых молекул могут образовать канал, через который проходят определенные ионы или молекулы.
Концентрация веществ, в частности ионов, по обе стороны мембраны не одинакова. Каждая сторона несет свой электрический заряд. Различия концентрации ионов создают соответственно и разность элект- рических потенциалов. Одной из важнейших функций биологических мембран, в том числе клеточной мембраны, является транспорт.
Биологической мембраной является цитолемма. Ее толщина около 10 нм, так что при световой микроскопии она неразличима. Цитолемма обеспечивает в первую очередь разграничительную функцию клетки с внешней средой. Она выполняет также транспортную и рецепторную функции (восприятие сигналов из внешней для клетки среды). Цитолемма, таким образом, обеспечивает поверхностные свойства клетки.
Наружный и внутренний электронноплотные слои цитолеммы имеют каждый толщину около 2-5 нм, средний электронно-прозрачный слой - около 3 нм. В билипидный слой цитолеммы погружены молекулы белка. Некоторые из них (интегральные, или трансмембранные) проходят через всю толщину мембраны, другие (периферические, или
внешние) лежат во внутреннем или наружном монослое мембраны. Некоторые интегральные белки связаны нековалентными связями с белками цитоплазмы.
Внешняя поверхность цитолеммы покрыта гликокаликсом. Его толщина различная и колеблется даже на разных участках поверхности одной клетки от 7,5 до 200 нм. Гликокаликс представляет собой совокупность молекул, связанных с белками мембраны. По составу эти молекулы могут представлять собой цепочки полисахаридов, гликолипидов и гликопротеинов.
Многие молекулы гликокаликса функционируют в качестве специфических молекулярных рецепторов. Чем больше конкретных спе- цифических рецепторов находится в гликокаликсе, тем активнее клетка реагирует на соответствующие сигнальные вещества. Если в гликокаликсе нет таких молекул-рецепторов, клетка на внешние вещества не реагирует.
МЕЖКЛЕТОЧНЫЕ СОЕДИНЕНИЯ
При контакте клеток друг с другом их цитолеммы вступают во взаимодействия. При этом образуются особые объединяющие структуры - межклеточные соединения (рис. 3). Они формируются при образовании многоклеточного организма во время эмбрионального развития и при образовании тканей. Межклеточные соединения (junctiones intercellulares) подразделяются на простые и сложные. В простых соединениях цитолеммы соседних клеток соприкасаются друг с другом или образуют изгибы наподобие зубцов, так что зубцы цитолеммы одной клетки внедряются между двумя зубцами другой (зубчатое соединение). Встречаются соединения в виде глубоко вдающихся впячиваний цитолеммы одной клетки в цитолемму другой (пальцевидное соединение). Между цитолеммами соседних клеток во многих соединениях сохраняется межклеточная щель шириной 15-20 нм.
Сложные (специальные) соединения, в свою очередь, подразделяются на адгезионные (пятно смыкания), запирающие зоны и зоны слипания. Пятно слипания, или десмосома (macula adherens), состоит из двух электронноплотных пластинок, принадлежащих цитолеммам соседних клеток, разделенных межклеточным пространством размером около 25 нм, заполненным тонкофибриллярным веществом гликопротеиновой природы.
Полудесмосома, образованная лишь одной пластинкой с входящими в нее тонофиламентами, прикрепляет клетку к базальной мембране. Зона
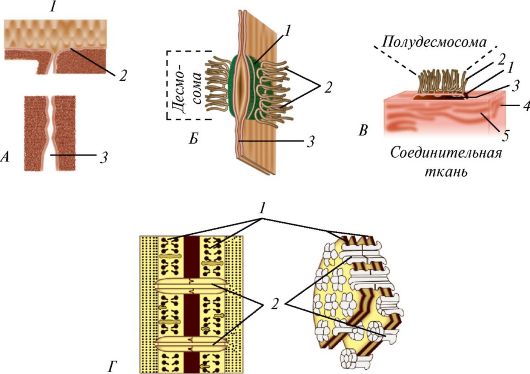
Рис. 3. Межклеточные соединения: А - плотный контакт: 1 - просвет контакта; 2 - плотный контакт; 3 - межклеточное пространство; Б - десмосома: 1 - электронноплотная пластинка; 2 - тонофиламенты; 3 - межклеточное пространство; В - полудесмосома: 1 - электронноплотная пластинка; 2 - тонофиламенты; 3 - межклеточное пространство; 4 - гликокаликс; 5 - базальная мембрана; Г - нексус (щелевидное соединение): 1 - цитолеммы смежных клеток; 2 - коннексоны, каждый из которых состоит из 6 субъединиц с цилиндрическим каналом
слипания (zonula adherens), или лентовидная десмосома, представляет собой «ленту», которая огибает всю поверхность клетки вблизи ее апикального отдела. Ширина межклеточного пространства, заполненного волокнистым веществом, не превышает 15-20 нм. Цитоплазматическая поверхность «ленты» уплотнена и укреплена сократительным пучком актиновых филаментов.
Запирающая зона (zonula occludens), или плотное соединение, имеет вид пояска шириной 0,5-0,6 мкм, поэтому через плотные контакты молекулы не проходят.
Нексус, или щелевидный контакт (nexus, s. macula communicans), или синапс, рассматривается как проводящее соединение. Через такие сое- динения из одной клетки в другую проходят водорастворимые малые
молекулы с молекулярной массой не более 1500 Д. Такими контактами соединены очень многие клетки человека. В нексусе между цитолеммами соседних клеток имеется пространство шириной 2-4 нм. Щелевые контакты играют важную роль в осуществлении функции клеток, обладающих выраженной электрической активностью (например, кардиомиоцитов). Синапсы играют важную роль в осуществлении функций нервной системы (см. «Нервная ткань»).
У многих клеток цитолемма образует различной величины вытягивания - микроворсинки, которые обеспечивают увеличение клеточной поверхности. Это, как правило, связано с осуществлением функции вса- сывания веществ из внешней для клетки среды. Микроворсинки представляют собой выпячивания цитолеммы длиной 1-2 мкм и диаметром до 0,1 мкм. В гиалоплазме ворсинок проходят продольные пучки актиновых микрофиламентов (30-40), поэтому длина микроворсинок может изменяться. У основания микроворсинки в поверхностном комплексе клетки происходит объединение ее микрофиламентов с элементами цитоскелета.
Поверхность микроворсинок также покрыта гликокаликсом. При особой активности всасывания микроворсинки близко располагаются друг возле друга. При этом микроворсинки образуют щеточную каемку. У щеточной каемки многие молекулы гликокаликса обладают фермен- тативной активностью.
Особо крупные микроворсинки длиной до 7 мкм называют стереоцилиями. Они имеются у некоторых специализированных клеток (например, у сенсорных клеток в органах равновесия и слуха).
ЦИТОПЛАЗМА
Основными структурами цитоплазмы являются, как упомянуто ранее, гиалоплазма (матрикс), органеллы и включения.
Гиалоплазма (от греч. hyalos - стекло), занимающая около 53-55% объема клетки, в физико-химическом отношении представляет собой коллоид, состоящий из воды, ионов и многих молекул органических веществ, комплексных соединений типа гликолипидов, гликопротеинов и липопротеинов. В гиалоплазме взвешены органеллы и включения. В гиалоплазме осуществляется наиболее распространенный процесс выделения энергии - гликолиз, представляющий собой последовательную цепь ферментативных реакций, в результате которых шестиуглеродная молекула глюкозы распадается на две трехуглеродные
молекулы пировиноградной кислоты. При гликолитическом расщеплении одной молекулы глюкозы образуются 4 молекулы АТР.
Органеллы - это структуры цитоплазмы, выполняющие конкретные функции, необходимые для поддержания жизнедеятельности клетки. Это обеспечение ее энергетического обмена, синтетических процессов, транспорта веществ и т. п. Органеллы, присущие всем клеткам, называют органеллами общего значения, присущие некоторым специализированным видам клеток - специальными. В зависимости от того, включает структура органеллы биологическую мембрану или нет, различают органеллы мембранные и немембранные.
Органеллы общего значения. Мембранные органеллы. Каждая мембранная органелла представляет собой структуру цитоплазмы, ограниченную биологической мембраной. К мембранным органеллам относятся митохондрии, эндоплазматическая сеть, внутренний сетчатый аппарат (комплекс Гольджи), лизосомы и пероксиомы.
Митохондрии участвуют в процессах клеточного дыхания и преобра- зуют энергию, которая при этом освобождается, в форму, доступную для использования другими структурами клетки. За ними закрепилось образное название «энергетические станции клетки».
Митохондрии, в отличие от других органелл, обладают собственной генетической системой, необходимой для самовоспроизведения и синтеза белков. Они имеют свои ДНК, РНК и рибосомы, отличающиеся от таковых в ядре и в других отделах цитоплазмы клетки. Митохондрии способны размножаться в клетке путем бинарного деления. Таким образом, они являются самовоспроизводящимися органеллами. Вместе с тем часть белков кодируется ядерными генами и поступает в митохондрии из гиалоплазмы, поэтому митохондрии в отношении самовоспроизведения называют полуавтономными структурами.
При световой микроскопии митохондрии выглядят как округлые, удлиненные или палочковидные структуры длиной 0,3-5 мкм и шири- ной 0,2-1 мкм. Каждая митохондрия образована двумя мембранами - внешней и внутренней (рис. 4). Между этими мембранами расположено межмембранное пространство шириной 10-20 нм. Внешняя мембрана ровная, внутренняя образует многочисленные складки - кристы. Благодаря кристам площадь внутренней мембраны существенно увеличивается. Так, поверхность одной митохондрии гепатоцита составляет около 13 мкм2, а площадь ее крист - около 16 мкм2 (Krstic R., 1976). На внутренней (обращенной к матриксу) поверхности крист лежит множество электронноплотных субмитохондриальных элементарных частиц
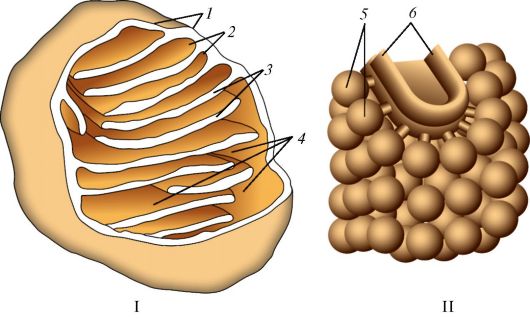
Рис. 4. Митохондрия: I - общая схема строения: 1 - наружная мембрана; 2 - внутренняя мембрана; 3 - кристы; 4 - матрикс; II - схема строения кристы: 5 - грибовидные тельца (АТР-синтетаза); 6 - складка внутренней мембраны (по Албертсу и соавт. и по
де Дюву, с изменениями)
(до 4000 на 1 мкм2 мембраны), имеющих форму гриба. Эти частицы содержат фермент АТР-синтетазу, осуществляющую синтез и гидролиз АТР.
Пространство, ограниченное внутренней мембраной, заполнено коллоидным митохондриальным матриксом. Он имеет мелкозернистую структуру и содержит множество различных ферментов. В матриксе также заключен собственный генетический аппарат митохондрий. Митохондрии обладают также РНК и рибосомами (размерами около 15 нм), которые также находятся в матриксе. Кристы могут иметь вид складок, гребней. Иногда кристы имеют вид трубочек диаметром 20-60 нм. Это наблюдается в клетках, которые синтезируют стероиды, например в клетках яичек. Встречаются кристы в виде призм, например в астроцитах головного мозга. У человека наиболее распространены митохондрии с кристами. Обычно кристы ориентированы поперек длинной оси митохондрий, но могут иметь и продольную ориентацию. Кристы, как правило, не достигают противоположной стороны митохондриальной мембраны. Благодаря указанным складкам площадь внутренней мембраны резко возрастает.
Число, размеры и расположение митохондрий зависят от функции клетки, в частности от ее потребности в энергии и от места, где энергия
расходуется. Так, в одной печеночной клетке число митохондрий достигает 2500. Множество крупных митохондрий содержится в кардиомиоцитах и миосимпластах мышечных волокон. В спермиях богатые кристами митохондрии окружают аксонему промежуточной части жгутика.
Эндоплазматическая сеть представляет собой единую непрерывную структуру, ограниченную мембраной толщиной около 10 нм, образующей инвагинации и складки. На электронно-микроскопических фотографиях эндоплазматическая сеть выглядит как множество трубочек, плоских или округлых цистерн, мембранных пузырьков, имеющих сообщение с перинуклеарным пространством клетки (см. рис. 1). На мембранах эндоплазматической сети совершается многообразный первичный синтез веществ, необходимых для жизнедеятельности клетки. Их можно условно назвать первичными, потому что молекулы этих веществ будут подвергаться дальнейшим химическим превращениям в других структурах.
Большинство веществ синтезируется на наружной поверхности мембран эндоплазматической сети. Затем эти вещества переносятся через мембрану внутрь этой сети и там транспортируются к местам дальнейших биохимических превращений. Различают два типа эндоплазматической сети: зернистую (гранулярную, шероховатую) и незернистую (гладкую, агранулярную).
Мембраны зернистой (гранулярной) эндоплазматической сети покрыты рибосомами, поэтому при световой микроскопии зернистая эндоплазматическая сеть выглядит в виде базофильного вещества, дающего положительную окраску на РНК. На структурах зернистой эндоплазма- тической сети синтезируются интегральные мембранные белковые молекулы, которые транспортируются в другие участки клетки.
Поверхность незернистой (гладкой) эндоплазматической сети не имеет рибосом. Сама сеть представляет собой множество мелких трубочек диаметром около 50 нм каждая. Между трубочками часто расположены гранулы гликогена. На мембранах незернистой эндоплазматической сети синтезируются углеводы и липиды, среди них - гликоген и холестерин, она участвует и в синтезе стероидных гормонов (в клетках Лейдига, в корковых эндокриноцитах надпочечника). Незернистая эндоплазматическая сеть участвует также в выделении ионов хлора в париетальных клетках эпителия желез желудка. Как депо ионов кальция гладкая эндоплазматическая сеть участвует в сокращении кардиомиоцитов и волокон скелетной мышечной ткани. Чрезвычайно важна ее роль в детоксикации гепатоцитами веществ, которые поступают от кишки по воротной вене в печеночные капилляры.
По просветам структур эндоплазматической сети синтезированные вещества транспортируются к комплексу Гольджи. Одной из важнейших функций эндоплазматической сети является синтез белков и липидов для всех клеточных органелл.
Внутренний сетчатый аппарат (комплекс Гольджи) представляет собой совокупность цистерн, пузырьков, пластинок, трубочек, мешочков которые при световой микроскопии имеют вид сеточки. Чаще всего в комплексе Гольджи выявляются 3 мембранных элемента: уплощенные мешочки (цистерны), которые связаны между собой каналами, пузырьки и вакуоли. Концы цистерн расширены. От них отщепляются пузырьки и вакуоли, окруженные мембраной и содержащие различные вещества. Мембранные пузырьки имеет диаметр 50-65 нм. Более крупные секреторные гранулы имеют диаметр от 66 до 100 нм. Часть вакуолей содержит гидролитические ферменты, это предшественники лизосом.
Наиболее широкие уплощенные цистерны обращены в сторону эндоплазматической сети. В цистернах продолжается синтез полисахаридов, образуются комплексы белков, углеводов и липидов. По мере мо- дификации вещества переходят из одних цистерн в другие. На боковых поверхностях цистерн образуются выросты, куда перемещаются вещества. Выросты отщепляются в виде пузырьков, которые удаляются от комплекса Гольджи в различных направлениях по гиалоплазме.
Пузырьки, отщепляющиеся от комплекса Гольджи, направляются к поверхности клетки и выводят синтезированные вещества в межклеточный матрикс.
Ту сторону комплекса Гольджи, куда поступают вещества от эндоплазматической сети, называют цис-полюсом, противоположную - трансполюсом. Таким образом, комплекс Гольджи структурно и биохимически поляризован. По направлению от цис-поверхности к транс-поверхности увеличивается толщина мембран (от 6 до 8 нм), а также содержание в них холестерина и углеводных компонентов в мембранных гликопротеинах.
В процессе упаковки веществ в пузырьки расходуется значительное количество материала мембран, поэтому сборка мембран - еще одна функция комплекса Гольджи. Эта сборка совершается из веществ, поступающих, как обычно, от эндоплазматической сети. Секреторный путь и обновление мембран представлены на рис. 5.
Положение комплекса Гольджи в клетке обусловлено ее функциональной специализацией. В секретирующих клетках он находится между ядром и поверхностью выведения структур из клетки. Так, в бокаловидных слизистых клетках ядро смещено к базальному концу, а комплекс
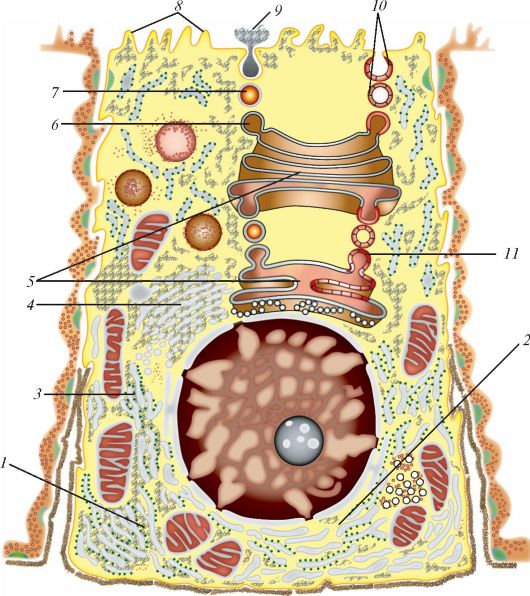
Рис. 5. Схема секреторного пути и обновления мембран: 1 - область, где происходит синтез белков, предназначенных для экспорта из клетки; 2 - область, где происходит синтез белков, предназначенных для обновления мембран; 3 - область, где происходит гликозилирование (1 + 2+3 - зернистая эндоплазматическая сеть); 4 - транспортные пузырьки, где происходит образование дисульфидных мостиков; 5 - комплекс Гольджи, где происходят добавление липидов, сульфатирование, удаление боковых цепей, терминальное гликозилирование; 6 - просекреторная гранула, где происходит протеолитическая доработка; 7 - секреторная гранула, где происходит концентрация секрета; 8 - цитолемма; 9 - экзоцитоз;10 - встраивание в мембрану; 11 - сборка элементов мембраны (по К. де Дюву, с изменениями)
Гольджи находится между ним и апикальной поверхностью. В клетках эндокринных желез, из которых секрет выводится в кровеносные капилляры, комплекс Гольджи представлен многими поверхностно лежащими структурами. В гепатоцитах (клетках паренхимы печени) структуры комплекса Гольджи располагаются группами: одни около желчных путей, другие - около сосудистых. В цитоплазме клеток (при световой микроскопии) комплекс занимает светлую зону около ядра, он окружен гранулярной эндоплазматической сетью и на ее базофильном фоне выглядит как «светлый дворик». Во всех случаях вблизи комплекса Гольджи концентрируются митохондрии. Это связано с происходящими в нем энергозависимыми реакциями.
Лизосомы, которые непосредственно отделяются от комплекса Гольд- жи, называют первичными. Каждая лизосома представляет собой мембранный пузырек диаметром 0,4-0,5 мкм, в котором содержится около 50 видов различных гидролитических ферментов в неактивированном состоянии (протеазы, липазы, фосфолипазы, нуклеазы, гликозидазы, фосфатазы, в том числе кислая фосфатаза). Молекулы этих ферментов, как всегда, синтезируются на рибосомах зернистой эндоплазматической сети, откуда переносятся транспортными пузырьками в комплекс Гольджи.
Все лизосомы клетки формируют лизосомное пространство, в котором с помощью протонного насоса постоянно поддерживается кислая среда - рН колеблется в пределах от 3,5 до 5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия. Это связано с особым расположением молекул лизосомной мембраны. Повреждение или нарушение проницаемости лизосомной мембраны приводит к активации ферментов и тяжелым повреждениям клетки вплоть до ее гибели.
Функция лизосом - внутриклеточный лизис («переваривание») высокомолекулярных соединений и частиц. Это могут быть собственные органеллы и включения или частицы, поступившие в клетку извне в ходе эндоцитоза. Захваченные частицы обычно окружены мембраной. Такие лизосомы называют фагосомами.
Процесс внутриклеточного лизиса (переваривания) осуществляется в несколько этапов. Сначала первичная лизосома сливается с фагосомой. Их комплекс называют вторичной лизосомой (фаголизосомой). Во вторичной лизосоме ферменты активируются и расщепляют поступившие в клетку полимеры до мономеров. Продукты расщепления транспортируются через лизосомную мембрану в цитозоль. Непереваренные вещества остаются в лизосоме и могут сохраняться в клетке, окруженные мембраной, очень долго в виде остаточного тельца. Остаточные тельца относят
уже не к органеллам, а к включениям. Возможен и другой путь превращений: вещества в фагосоме расщепляются полностью, после чего мембрана фагосомы распадается.
В процессе жизнедеятельности клетки на разных иерархических уровнях ее организации, начиная от молекул и кончая органеллами, постоянно происходит перестройка структур. Вблизи поврежденных или требующих замены участков цитоплазмы, обычно по соседству с комплексом Гольджи, образуется полулунная двойная мембрана, которая растет, окружая со всех сторон поврежденные зоны. Затем эта структура сливается с лизосомами. В такой аутофагосоме (аутосоме) совершается лизис структур органеллы.
Вторичные лизосомы могут сливаться между собой, а также с другими первичными лизосомами. При этом иногда образуются своеобразные вторичные лизосомы - мультивезикулярные тельца.
В некоторых случаях непереваренные остатки накапливаются в лизосомах, что приводит к их перегрузке («хронический запор»). Выделение непереваренных остатков путем экзоцитоза и их накопление во внеклеточной среде могут вызвать повреждение внеклеточных структур.
Пероксисомы представляют собой мембранные пузырьки диаметром от 0,2 до 0,5 мкм. Как и лизосомы, они отщепляются от цистерн трансполюса комплекса Гольджи. Различают две формы пероксисом. Мелкие пероксисомы (диаметром 0,15-0,25 мкм) имеются практически во всех клетках, они содержат мелкозернистый осмиофильный материал и морфологически мало отличаются от первичных лизосом. Крупные пероксисомы (диаметром более 0,25 мкм) присутствуют лишь в некоторых тканях (печень, почки). В них имеется кристалловидная сердцевина, в которой находятся ферменты в концентрированном виде. Наряду с пероксисомами встречаются и другие мембранные микротельца диаметром от 0,5 до 10 мкм, содержащие различные ферменты.
Пероксисомы содержат ферменты (пероксидазу, каталазу и оксидазу D-аминокислот). Пероксидаза участвует в обмене перекисных соединений, в частности перекиси водорода, которая токсична для клетки. Для биохимических реакций в пероксисомах используется молекулярный кислород. Пероксисомы принимают также участие в нейтрализации многих других токсичных соединений, например этанола. Пероксисомы участвуют также в обмене липидов, холестерина и пуринов.
Немембранные органеллы. К немембранным органеллам относятся цитоскелет, клеточный центр и рибосомы.
Цитоскелет (клеточный скелет) включает микротрубочки, микрофиламенты и промежуточные филаменты. Микротрубочки (см. рис. 1) расположены
в цитоплазме клетки. Они представляют собой полый цилиндр диаметром 20-30 нм. Стенка микротрубочки имеет толщину 6-8 нм. Многие микротрубочки расположены радиально по отношению к центриолям. Микро- трубочки прочны и образуют опорные структуры цитоскелета. Часть микротрубочек располагается в соответствии с силами сжатия и натяжения, которые испытывает клетка. Особенно хорошо это заметно в клетках эпителиальных тканей, которые разграничивают разные среды организма. Микротрубочки участвуют в транспорте веществ внутри клетки. Структура стенок микротрубочек может меняться при различных воздействиях на клетки. В подобных случаях может нарушаться внутриклеточный транспорт.
Микрофиламенты - это белковые нити толщиной около 4 нм. Большинство микрофиламентов образовано молекулами актинов, которых около 10 видов. Кроме того, актиновые филаменты могут группироваться в пучки, образующие собственно опорные структуры цитоскелета. Филаменты актина способны образовывать комплексы с полимерными молекулами белка миозина. Микрофиламенты, будучи соединенными с цитолеммой, способны менять ее конфигурацию. Это важно для поступления веществ в клетку посредством пиноцитоза и фагоцитоза. Проме- жуточные филаменты образованы длинными белковыми молекулами. Толщина таких филаментов 8-10 нм. Они тоньше микротрубочек, но толще микрофиламентов, за что и получили свое название.
Клеточный центр (рис. 6) образован двумя центриолями (диплосома) и центросферой. Обе центриоли диплосомы расположены под углом друг к другу. Основная функция клеточного центра - сборка микротрубочек. Каждая центриоль представляет собой цилиндр, стенки которого, в свою очередь, состоят из 9 комплексов микротрубочек длиной около 0,5 мкм и диаметром около 0,25 мкм. Каждый комплекс состоит из трех микротрубочек и поэтому называется триплетом. Триплеты, расположенные по отношению друг к другу под углом около 50?, состоят из трех микротрубочек (изнутри кнаружи): полной А и неполных В и С диаметром около 20 нм каждая. От трубочки А отходят две ручки. Одна из них направлена к трубочке С соседнего триплета, другая - к центру цилиндра, где внутренние ручки образуют фигуру звезды или спиц колеса. Центриоли являются саморегулирующимися структурами, которые при делении клетки удваиваются и расходятся к противоположным полюсам клетки. Центриоли участвуют в образовании базальных телец, ресничек, жгутиков и митотического веретена.
Рибосомы - тельца размерами 20-30 нм, состоят из двух субъединиц - большой и малой. Каждая субъединица представляет собой комплекс рибосомной РНК (рРНК) с белками. Основная функция рибосом - сборка
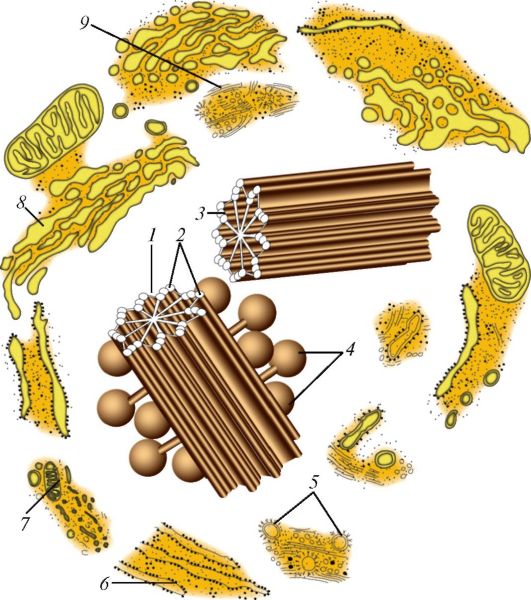
Рис. 6. Клеточный центр и другие структуры цитоплазмы: 1 - центросфера; 2 - центриоль на поперечном срезе (триплеты микротрубочек, радиальные спицы, центральная структура «колеса телеги»); 3 - центриоль (продольный разрез); 4 - сателлиты; 5 - окаймленные пузырьки; 6 - зернистая эндоплазматическая сеть; 7 - митохондрия; 8 - внутренний сетчатый аппарат (комплекс Гольджи); 9 - микротрубочки (по Р. Крстичу, с изменениями)
белковых молекул из аминокислот, доставляемых к ним транспортными РНК (тРНК). Между субъединицами рибосомы имеется щель, в которой проходит молекула информационной РНК (иРНК), а на большой субъединице - бороздка, в которой располагается и по которой сползает формирующаяся белковая цепь. Сборка аминокислот производится в соответствии с чередованием нуклеотидов в цепи иРНК. Таким способом осуществляется трансляция генетической информации.
Рибосомы могут находиться в гиалоплазме по одиночке либо группами в виде розеток, спиралей, завитков. Такие группы называют поли- рибосомами (полисомами). Значительная часть рибосом прикреплена к мембранам: к поверхности эндоплазматической сети и к наружной мембране кариотеки. Свободные рибосомы синтезируют белок, необходимый для жизнедеятельности самой клетки, прикрепленные - белок, подлежащий выведению из клетки. Число рибосом в клетке может достигать десятков миллионов.
Среди специальных органелл следует рассматривать также мерцательные реснички и жгутики как наиболее распространенные. Описание остальных специальных органелл дано в соответствующих в разделах курсов эмбриологии, общей и частной гистологии.
Ресничка представляет собой вырост клетки, окруженный цитолем- мой (рис. 7). У основания реснички на уровне кортикального слоя цитоплазмы находится базальное тельце (кинетосома), которое образовано 9 периферическими триплетами коротких микротрубочек, окружающих один центральный триплет. Над базальным тельцем в названный цитоплазматический вырост направляется осевая нить (осевой филамент, аксонема), тоже образованная микротрубочками. Сразу над базальным тельцем микротрубочки аксонемы тоже образуют 9 периферических триплетов. На поперечном сечении реснички ее возвышающаяся часть напоминает колесо с 9 спицами, в центре которого лежит нечетко контурирующаяся центральная капсула, окружающая две центральные одиночные микротрубочки, с которыми она связана белковыми отростками.
Все реснички клетки совершают координированные колебательные движения. Число ресничек достигает нескольких сотен. Так, до 250 ресничек длиной 5-15 мкм и диаметром 0,15-0,25 мкм покрывают апикальную поверхность реснитчатых эпителиоцитов верхних дыхательных путей, маточных труб, семенных канальцев.
Жгутики также выполняют функцию движения. Это достигается посредством скольжения дуплетов микротрубочек относительно друг друга. Оно обусловлено изменениями конфигурации молекул белка динеина.
Включения представляют собой скопления веществ в клетке, обра- зующиеся как продукты ее метаболизма или попавшие в клетку извне. Среди включений довольно условно различают трофические, пигментные и секреторные. К трофическим включениям относят капли жира, гранулы гликогена, белковые гранулы. Эти вещества накапливаются в клетке, а затем расходуются ею при соответствующих функциональных потребностях. Пигментные включения могут лежать свободно, но могут быть окружены мембраной.
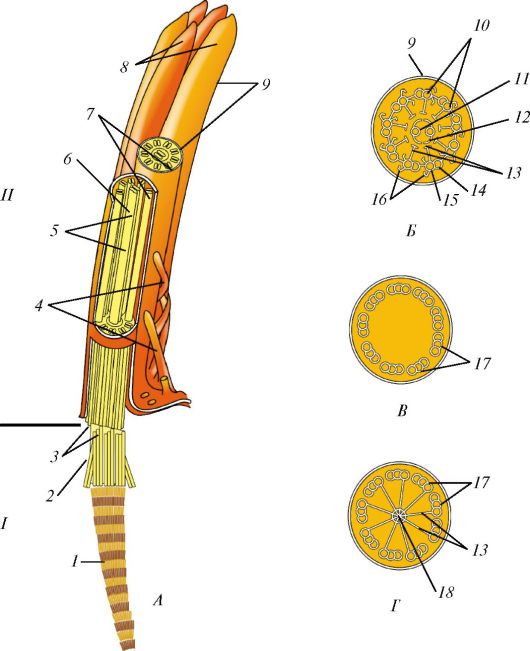
Рис. 7. Ресничка:
А - ресничка на продольно-поперечном разрезе: I - внутриклеточная часть; II - внеклеточная часть; 1 - базальный корешок; 2 - базальное тельце; 3 - наружные микротрубочки; 4 - микроворсинки; 5 - центральная капсула; 6 - центральный дуплет; 7 - периферические дуплеты; 8 - реснички; 9 - цитолемма; Б - поперечный срез через внеклеточную часть: 10 - периферические дуплеты; 11 - центральный дуплет; 12 - центральная капсула; 13 - спицы; 14 - субфибрилла а; 15 - субфибрилла б; 16 - динеин; В - поперечный срез через промежуточную область, соединяющую аксонему с базальным тельцем: 17 - триплеты микротрубочек; Г - срез через базальное тельце: 17 - триплеты; 18 - центральный цилиндр
ТРАНСПОРТ ВЕЩЕСТВ И МЕМБРАН В КЛЕТКЕ
Вещества перемещаются в клетке, будучи упакованными в мембраны («передвижение содержимого клетки в контейнерах»). Сортировка веществ и их передвижение связаны со специальными белками-рецеп- торами в мембранах комплекса Гольджи. Транспорт через мембраны, в том числе и через цитоплазматическую мембрану (цитолемму), является одной из важнейших функций живых клеток. Различают два типа транспорта: пассивный и активный. Пассивный транспорт не требует затрат энергии, активный транспорт - энергозависимый. Пассивный транспорт не требует затрат энергии, активный транспорт - энергозависимый. Пассивный транспорт незаряженных молекул осуществляется по градиенту концентрации путем диффузии. Транспорт заряженных веществ зависит от разности потенциалов на поверхности цитолеммы. Как правило, внутренняя поверхность цитоплазматической мембраны (цитолеммы) несет отрицательный заряд, что облегчает проникновение в клетку положительно заряженных ионов.
Переход ионов или молекул из зоны, где эти вещества находятся в более высокой концентрации, в зону с более низкой концентрацией получил название диффузии. Специфические транспортные белки, встроенные в мембрану, переносят через нее небольшие молекулы. Каждый транспортный белок осуществляет транспорт молекул одного класса или только одного соединения. Трансмембранные белки либо являются переносчиками, либо образуют каналы. Диффузия может быть нейтральной, когда незаряженные вещества проходят между липидными молекулами или через белки цитолеммы, формирующие каналы. Облегченная диффузия происходит при участии специфических белков-переносчиков, связывающих вещество и переносящих его через мембрану. Облегченная диффузия протекает быстрее, чем нейтральная.
Активный транспорт осуществляют белки-переносчики. Для этого расходуется энергия, получаемая вследствие гидролиза АТР, а также при разности потенциалов (зарядов) на различных поверхностях клеточной мембраны. Активный транспорт происходит против градиента концентрации. На цитолемме с помощью натриево-калиевого насоса поддерживается мембранный потенциал. Этот насос, накачивающий ионы калия в клетку против градиентов концентрации, а ионы натрия - во внеклеточное пространство, является ферментом АТР-азой. Благодаря АТР-азе натрий переносится через мембрану и выводится во внеклеточную среду, а калий переносится внутрь клетки. АТР-аза осуществляет также активный транспорт аминокислот и сахаров.
КЛЕТОЧНОЕ ЯДРО
В большинстве клеток ядро шаровидное или овоидное. Однако встречаются ядра и другой формы (кольцевидные, палочковидные, веретенообразные, бобовидные, сегментированные и др.). Размеры ядра колеблются в широких пределах - от 3 до 25 мкм. Наиболее крупным ядром обладает яйцеклетка. Большинство клеток человека имеет одно ядро, но имеются двухъядерные (например, некоторые нейроны, клетки печени, кардиомиоциты). Иногда многоядерными клетками называют структуры, которые образовались не вследствие полиплоидизации исходной клетки, а в результате слияния нескольких одноядерных клеток. Такие структуры имеют специальное название «симпласты»; они встречаются, в частности, в составе скелетных поперечно-полосатых мышечных волокон.
Ядро отделено от цитоплазмы ядерной оболочкой - кариотекой, или кариолеммой, которая образована двумя мембранами - внутренней и наружной (рис. 8). Пространство между мембранами называют перинуклеарным пространством. Оно имеет ширину 20-50 нм и сохраняет сообщение с полостями эндоплазматической сети. Со стороны цитоплазмы наружная мембрана нередко покрыта рибосомами.
Местами внутренняя и наружная мембраны кариотеки (ядерной оболочки) сливаются, а в месте слияния образуется закрытое гранулами отверстие - ядерная пора. Ядерная пора не зияет: в ней упорядоченно располагаются белковые молекулы, которые формируют поровый комплекс. Комплекс поры представляет собой сложную структуру, состоящую из двух рядов связанных между собой белковых гранул. Отверстие поры закрыто тонкой диафрагмой.
Через поровые комплексы осуществляется избирательный транспорт молекул и частиц из ядра в цитоплазму и обратно. Поры могут занимать до 25% поверхности ядра. Число пор у одного ядра достигает 3000-4000, а их плотность составляет около 11 на 1 мкм2 ядерной оболочки. Из ядра в цитоплазму транспортируются в основном разные виды РНК. Из цитоплазмы в ядро поступают все ферменты, необходимые для синтеза РНК. Внутренняя поверхность кариотеки связана с многочисленными промежуточными филаментами.
Под ядерной оболочкой находятся нуклеоплазма (кариоплазма) (karioplasma, s. nucleoplasma), имеющая гомогенное строение, и ядрышко. В нуклеоплазме неделящегося ядра, в его ядерном белковом матриксе находятся осмиофильные гранулы (глыбки) хорошо окра- шивающегося гетерохроматина. Участки более разрыхленного хроматина, расположенные между гранулами, называют эухроматином.
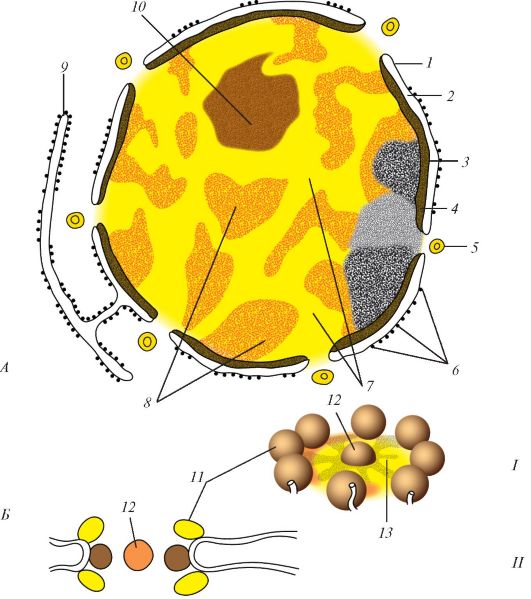
Рис. 8. Ядро клетки: А - ядро: 1 - наружная мембрана кариотеки (наружная ядерная мембрана); 2 - перинуклеарное пространство; 3 - внутренняя мембрана кариотеки (внутренняя ядерная мембрана); 4 - ядерная пластинка; 5 - поровый комплекс; 6 - рибосомы; 7 - нуклеоплазма (ядерный сок); 8 - хроматин; 9 - цистерна зернистой эндоплазматической сети; 10 - ядрышко; Б - поровый комплекс: I - пространственная реконструкция; II - схема основных структур; 11 - пери- ферические гранулы; 12 - центральная гранула; 13 - дифрагма поры (по Б. Албертсу и др., с изменениями)
Разрыхленный хроматин называют деконденсированным хроматином, в нем наиболее интенсивно протекают синтетические процессы. Во время деления клетки хроматин уплотняется, конденсируется, образует хромосомы.
Хроматин неделящегося ядра и хромосомы делящегося образованы молекулами дезоксирибонуклеиновой кислоты (ДНК), связанной с ри- бонуклеиновой кислотой (РНК) и белками (гистонами и негистонами). Следует подчеркнуть химическое тождество хроматина и хромосом.
Каждая молекула ДНК состоит из двух длинных правозакрученных полинуклеотидных цепей (двойных спиралей). Каждый нуклеотид состоит из азотистого основания, сахара и остатка фосфорной кислоты. Основание расположено внутри двойной спирали, а сахарофосфатный скелет - снаружи.
Наследственная информация в молекулах ДНК записана в линейной последовательности ее нуклеотидов. Элементарной частицей наследственности является ген. Ген - это участок ДНК, имеющий определенную последовательность расположения нуклеотидов, ответственных за синтез одного определенного специфического белка.
Молекула ДНК в ядре
упакована компактно. Так, одна молекула ДНК, содержащая 1 млн
нуклеотидов, при их линейном расположении заняла бы отрезок длиной
всего
Молекулы ДНК, связанные с белками-гистонами, образуют нуклеосомы, являющиеся структурными единицами хроматина. Нуклеосомы имеют вид бусинок диаметром 10 нм. Каждая нуклеосома состоит из гистонов, вокруг которых закручен участок ДНК, включающий 146 пар нуклеотидов. Между нуклеосомами располагаются линейные участки ДНК, состоящие из 60 пар нуклеотидов.
Хроматин представлен фибриллами, которые образуют петли длиной около 0,4 мкм, содержащие от 20 000 до 30 000 пар нуклеотидов.
В результате уплотнения (конденсации) и закручивания (суперспирализации) дезоксирибонуклеопротеидов (ДНП) в делящемся ядре они становятся видимыми. Эти структуры (хромосомы) представляют собой удлиненные палочковидные образования, имеющие два плеча, разделенные так называемой перетяжкой - центромерой. В зависимости от расположения центромеры и взаимного расположения и длины плеч (ножек) выделяют 3 типа хромосом: метацентрические, имеющие примерно одинаковые плечи, субметацентрические, у которых длина плеч различная, а также акроцентрические хромосомы, у которых одно плечо длинное, а другое очень короткое, еле заметное. В хромосоме имеются эу- и гетерохроматиновые
участки. Последние в неделящемся ядре и в ранней профазе митоза остаются компактными. Чередование эу- и гетерохроматиновых участков используют для идентификации хромосом. Поверхность хромосом покрыта различными молекулами, главным образом рибонуклеопротеинами.
В соматических клетках имеются по две копии каждой хромосомы, их называют гомологичными. Они одинаковы по длине, форме, строению, несут одни и те же гены, которые расположены одинаково. Особенности строения, число и размеры хромосом называют кариотипом. Нормальный кариотип человека включает 22 пары аутосом и одну пару половых хромосом (XX у женщин или XY у мужчин). Соматические клетки человека (диплоидные) имеют удвоенное число хромосом - 46. Половые клетки содержат гаплоидный (одинарный) набор - 23 хромосомы, в половых клетках ДНК в 2 раза меньше, чем в диплоидных соматических клетках. Разные участки хромосом обеспечивают синтез различных РНК. Особенно выделяются участки, синтезирующие рибосомные РНК (рРНК); ими обладают не все хромосомы. Эти участки называют ядрышковыми организаторами.
Ядрышко (nucleoolus) (одно или несколько) выявляется во всех неделящихся клетках. Оно имеет вид интенсивно окрашивающегося округлого тельца, величина которого пропорциональна интенсивности белкового синтеза. Ядрышко состоит из электронноплотной нуклеолонемы (от греч. nema - нить), в которой различают нитчатую (фибриллярную) часть, состоящую из множества переплетающихся нитей РНК толщиной около 5 нм, и гранулярную часть. Гранулярная (зернистая) часть образована зернами диаметром около 15 нм, представляющими собой частицы рибонуклеопротеидов (РНП) - предшественников рибосомных субъединиц. Околоядрышковый хроматин внедряется в углубления нуклеолонемы. В ядрышке образуются предшественники рибосом, которые перемещаются к поровым комплексам, проходят через них в цитоплазму клетки и превращаются в рибосомы.
ФУНКЦИИ КЛЕТОК
В клетках постоянно осуществляется обмен веществ, или метаболизм (от греч. metabole - перемена, превращение), который представляет собой совокупность процессов ассимиляции (реакций биосинтеза сложных биологических молекул из более простых) и диссимиляции (реакций расщепления). В результате диссимиляции освобождается энергия, заключенная в химических связях пищевых веществ. Эта энергия используется клеткой для осуществления различной работы, в том числе и ассимиляции.
Все биохимические реакции в клетке строго структурированы и осуществляются при участии высокоспецифических биокатализаторов - ферментов, или энзимов (от греч. en - в, zyme - брожение, закваска). Ферменты ускоряют реакцию на 10 порядков (в 1010 раз). По общему типу воздействия ферменты подразделяются на 6 классов. Оксиредуктазы катализируют окислительно-восстановительные реакции, трансферазы уча- ствуют в переносе функциональных групп, гидролазы обеспечивают реакции гидролиза, лиазы - присоединение групп по двойным связям, изомеразы осуществляют перевод соединений в другую изомерную форму, а лигазы (не путать с лиазами!) связывают молекулярные группировки в цепи.
Все процессы в клетках происходят при участии поступающих в клетку веществ (эндоцитоз) и выведения веществ из клетки (экзоцитоз). Эти процессы, а также внутриклеточный перенос веществ в мембранных пузырьках связаны со слипанием и объединением бислоев мембран (рис. 9). При эндоцитозе слипаются два наружных монослоя плазматической мембраны, при экзоцитозе - два внутренних, обращенных к цитоплазме слоя.
Эндоцитоз. Существует несколько способов эндоцитоза (от греч. endon - внутри, kytos - клетка). При эндоцитозе этот транспорт осуществляется с участием цитолеммы. Кроме того, существуют более сложные способы - пиноцитоз (от греч. pino - пью) и фагоцитоз (от греч. phagos - пожирающий). Обычно под пиноцитозом понимают захват клеткой жидких коллоидных частиц, под фагоцитозом - захват корпускул (более плотных и крупных частиц, а также других клеток). Механизм пино- и фагоцитоза различен.
Пиноцитоз. Для того, чтобы внешние молекулы поступили в клетку, они должны быть сначала связаны рецепторами гликокаликса. Цитолемма вместе с присоединенными извне молекулами впячивается, затем ее края сближаются и смыкаются. В результате от цитолеммы отщепляется пузырек, несущий в себе захваченные молекулы. Такие пузырьки получили название окаймленных. Окаймленные пузырьки беспрепятственно транспортируются в клетке именно к тем участкам ци- топлазмы, где должно использоваться их содержимое. Если вещества переносятся через клетку из одной среды в другую без изменений, то этот процесс называют трансцитозом. Путем трансцитоза могут переноситься и белковые молекулы, в частности иммуноглобулины.
Фагоцитоз. Крупные частицы также могут распознаваться рецепто- рами клетки и захватываться клетками. Выросты клетки охватывают частицу и объединяются над ней. Наружные листки выростов сливаются, замыкая поверхность клетки. Глубокие листки выростов образуют мембрану вокруг поглощенной частицы - формируется фагосома. Фагосома
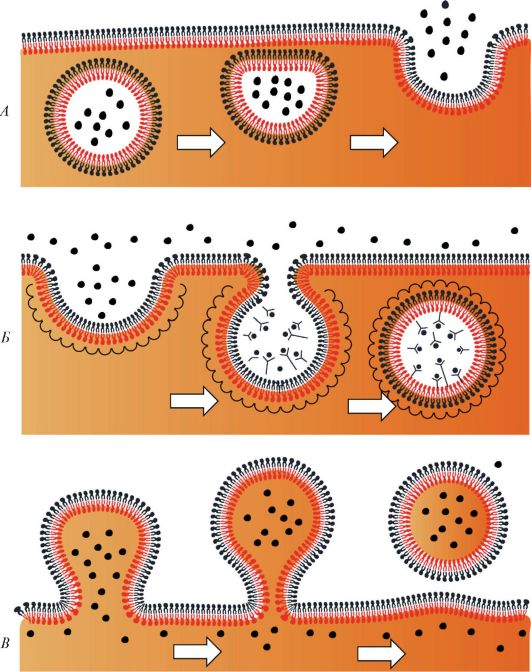
Рис. 9. Слипание и объединение бислоев (мембран) в трех процессах: в экзоцитозе (А), в эндоцитозе (Б) и в отпочковывании (В). Стрелками показана последовательность этих процессов (по Б. Албертсу и др., с изменениями)
сливается с лизосомами, в результате чего возникает их комплекс - гетеролизосома (гетеросома).
Способностью к фагоцитозу потенциально обладают все клетки, но в организме лишь некоторые специализируются в этом направлении. Таковы нейтрофильные лейкоциты и макрофаги.
Экзоцитоз. Выведение веществ из клетки осуществляется благодаря нескольким механизмам. Один из них - пассивный транспорт вследствие разности концентраций веществ внутри и вне цитолеммы, другой - активный транспорт. Таким путем выводятся из клетки ионы и мелкие молекулы. Третий механизм обеспечивает выведение крупномолекулярных соединений из клетки.
Крупномолекулярные соединения в виде транспортных пузырьков с участием микротрубочек направляются к клеточной поверхности. Мембрана пузырька сливается с цитолеммой, и содержимое пузырька оказывается за пределами клетки.
Слияние пузырька с цитолеммой может совершаться без каких-либо дополнительных сигналов. Такой экзоцитоз называют конститутивным. Так выводится из клетки большинство продуктов ее собственного метаболизма. Ряд клеток предназначен для синтеза специальных соединений - секретов, которые используются в других частях организма. Для того чтобы транспортный пузырек с секретом слился с цитолеммой, необходимы сигналы извне. Такой экзоцитоз называют регулируемым. Сигнальные молекулы, способствующие выведению секретов, называют либеринами (рилизинг- факторами), а препятствующие выведению - статинами. Мембрана транспортного пузырька встраивается в цитолемму и становится ее частью. Экзоцитоз и возврат мембран эндосом в нормально функционирующей клетке уравновешены с поглощением мембран в ходе пино- и фагоцитоза.
Внутриклеточный синтез. Управление внутриклеточным синтезом осуществляется из ядра клетки. На активных участках хромосом синтезируются молекулы РНК. Они транспортируются к поровым комплексам и поступают в цитоплазму. На рибосомах из аминокислот происходит сборка белков, которые в соответствии с их назначением можно отнести к трем группам. Одна группа - это структурные белки, которые используются клеткой для построения собственных органелл, другая - белки, выделяемые клеткой вовне, это ее секреты; третья группа - ферменты, которые обеспечивают все внутриклеточные биохимические превращения как катализаторы. Часть ферментов остается в цитоплазме. Одни из них функционируют в гиалоплазме, другие встраиваются в органеллы. Третья часть ферментов направляется в ядро и там регулирует считывание генетической информации с ДНК и матричный синтез РНК.
В ядро возвращаются те белки, которые участвуют в построении самих хромосом.
Вещества, которые синтезируются на мембранах эндоплазматической сети, поступают в транспортные пузырьки и доставляются к комплексу Гольджи. В нем протекают заключительные этапы синтеза.
Синтез белка (трансляция) связан с процессом транскрипции - пере- писывания информации, хранящейся в ДНК. Благодаря ядерной оболочке в клетках процессы транскрипции и трансляции проходят в разных структурах и разделены во времени.
Транскрипция осуществляется в ядре. Информация о структуре белка, заключенная в ДНК, «переписывается» на информационную РНК (иРНК). При этом с одного гена может «переписываться» множество молекул иРНК, которые транспортируются из ядра в цитоплазму, где и выполняют свои функции.
В клетках существует 3 типа РНК. Среди них иРНК переносит информацию о нуклеотидной последовательности ДНК к рибосомам. В образовании рибосом участвует рибосомная РНК (рРНК). Небольшие транспортные РНК (тРНК) выполняют двойную функцию: они присоединяют молекулу аминокислоты, транспортируют ее к рибосоме и узнают триплет, соответствующий этой аминокислоте в молекуле иРНК. Рибосомная и транспортная РНК (рРНК и тРНК) синтезируются на идентичных генах, которые (в отличие от генов иРНК) в каждой клетке имеются в виде множества копий. Реакции синтеза белка осуществляют рибосомы, которые считывают информацию.
Перемещаясь по цепи иРНК, рибосома присоединяет аминокислоты, которые связываются между собой, а молекулы тРНК отделяются, чтобы вскоре присоединить новую аминокислоту.
ЖИЗНЕННЫЙ ПУТЬ КЛЕТОК
Как известно, клетки образуются в результате деления. Во вновь возникшей клетке не всегда сразу существуют все системы, обеспечиваю- щие ее специфическую функцию. Должно пройти некоторое время (время созревания), чтобы сформировались все органеллы и были бы синтезированы все необходимые ферменты.
Зрелая клетка может функционировать различное время. Некоторые клетки сохраняются в течение всей жизни особи (например, нейроны). Таких видов клеток немного. Большинство клеток по истечении опре- деленного времени гибнет и по мере убыли замещается новыми. Скорость замещения у разных клеток неодинакова. Конечно, клетка может
погибнуть в результате многих внешних случайных причин, например, травмы, химического или радиационного поражения. В таком случае разрушение клетки происходит хаотично, а продукты ее распада сами оказывают раздражающее действие на окружение. Развивается воспалительная реакция. Подобная случайная гибель клеток называется некрозом и служит предметом изучения патологической анатомии. Большинство клеток, однако, погибает тогда, когда проявляются особые естественные генетические механизмы. Генетически запрограммированную клеточную гибель называют апоптозом. Механизм возникновения апоптоза весьма сложен.
Для поддержания жизненного равновесия клетка должна получать сигналы от других клеток, нередко от клеток другого вида. Обычно в качестве сигнальных выступают специфические молекулы олигопептидов. Поскольку они поддерживают жизнь клеток, их назвали цитокинами. Известно несколько десятков цитокинов, их действие разнообразно. На клетки одних видов цитокины оказывают более сильное действие, на клетки других видов - слабое.
В жизненном пути клеток многих видов наступает момент, когда их функциональные возможности исчерпываются. У таких клеток нарушается чувствительность к цитокинам и изменяется соотношение активности генов, обеспечивающих внутреннее равновесие. Гены, обеспечиваю- щие размножение клетки, блокируются. Напротив, гены, обеспечивающие синтез литических ферментов, стимулируются. Ферменты поступают в ядро и лизируют хроматин.
Хромосомы распадаются, синтез в клетке прекращается. Внешние проявления такой гибели клеток разнообразны. Их называли пикнозом (сморщивание ядра), хроматолизисом (снижение окрашиваемости ядра), кариорексисом (распад ядра на части).
Вслед за гибелью ядра разрушается цитоплазма. Остатки клетки фагоцитируются макрофагами. Материал погибших клеток перераба- тывается макрофагами и может выводиться ими на поверхность клетки. В таком случае этот материал может опять использоваться другими клетками. Вокруг клеток, подвергшихся апоптозу, воспалительный процесс не возникает. Жизнедеятельность ткани, часть которой составляли погибшие клетки, продолжается без нарушений.
Деление клеток. Клеточный цикл. Рост организма, увеличение числа клеток, их размножение происходят путем деления. Основными способами деления клеток в человеческом организме являются митоз и мейоз. Процессы, происходящие при этих способах деления клеток, протекают одинаково, но приводят к разным результатам.
Митотическое деление клеток (митоз) приводит к увеличению числа клеток, к росту организма. Таким способом обеспечивается обновление
клеток при их износе, гибели. Известно, что клетки эпидермиса живут 10-30 дней, эритроциты - до 4-5 мес. Нервные и мышечные клетки (волокна) живут в течение жизни человека.
У всех клеток при размножении (делении) наблюдаются изменения, укладывающиеся в рамки клеточного цикла. Клеточным циклом называют процессы, которые происходят в клетке от деления до деления. В клеточном цикле выделяют подготовку клетки к делению (интерфазу) и митоз (процесс деления клетки).
В интерфазе, которая длится примерно 20-30 ч, скорость биосинтетических процессов возрастает, увеличивается количество органелл. В интерфазе происходит матричный синтез ДНК и удвоение хромосом. В интерфазе удваивается масса клетки и всех ее структурных компонентов, в том числе центриолей. Происходит репликация (повторение, удвоение) молекул ДНК. Это процесс передачи генетической информации, хранящейся в родительской ДНК, путем точного ее воспроизведения в дочерних клетках. При этом родительская цепь ДНК служит матрицей для синтеза дочерних дезоксири- бонуклеиновых кислот. В итоге репликации каждая из двух дочерних молекул ДНК состоит из одной старой и одной новой цепей. В период подготовки к митозу в клетке синтезируются белки, необходимые для деления клетки (митоза). К концу интерфазы хроматин в ядре конденсирован.
Митоз (от греч. mitos - нить) представляет собой период, когда ма- теринская клетка разделяется на две дочерние (рис. 10). Митотическое деление клеток обеспечивает равномерное распределение структур клетки, ее ядерного вещества - хроматина - между двумя дочерними клетками. Длительность митоза от 30 мин до 3 ч. Митоз подразделяют на профазу, метафазу, анафазу, телофазу.
К началу профазы хроматин конденсируется, в результате чего в ядре образуется плотный клубок, в ядре становятся видимыми многочисленные плотные базофильные скопления, затем начинается спирализация хромосом. Вследствие этого хромосомы становятся индивидуально различимыми. В профазе постепенно распадается ядрышко, центриоли расходятся к полюсам клетки. Микротрубочки центриолей направлены к экватору, и в области экватора они перекрывают друг друга.
В метафазе разрушается ядерная оболочка, образуется биполярное митотическое веретено, состоящее из микротрубочек, хромосомные нити направляются к полюсам, сохраняя связь с экваториальной областью клетки. Структуры эндоплазматической сети и комплекса Гольджи распадаются на мелкие пузырьки (везикулы), которые вместе с митохонд- риями распределяются в обе половины делящейся клетки. В ходе метафазы хромосомы перемещаются и располагаются в одной плоскости перпендикулярно оси между полюсами. Образуется фигура, называемая
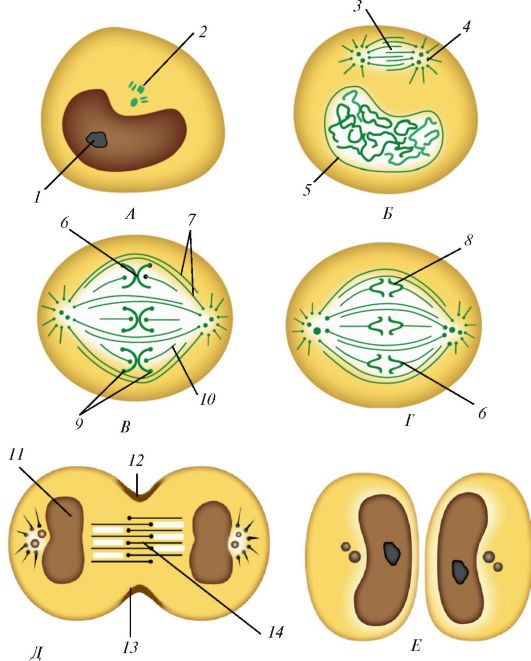
Рис 10. Стадии митоза. Показаны конденсация хроматина с образованием хромосом, образование веретена деления и равномерное распределение хромосом и центриолей по двум дочерним клеткам: А - интерфаза; Б - профаза; В - метафаза; Г - анафаза; Д - телофаза; Е - поздняя телофаза; 1 - ядрышко; 2 - центриоли; 3 - веретено деления; 4 - звезда; 5 - ядерная оболочка; 6 - кинетохор; 7 - непрерывные микротрубочки; 8, 9 - хромосомы; 10 - хромосомные микротрубочки; 11 - формирование ядра; 12 - борозда дробления; 13 - пучок актиновых нитей; 14 - остаточное (срединное) тельце (по А. Хэму и Д. Кормаку, с изменениями)
материнской звездой. При этом все хромосомы располагаются так, что их центромеры находятся в экваториальной плоскости, пересекающей продольную ось веретена под прямым углом (метафазная пластинка). В конце метафазы каждая хромосома начинает расщепляться продольной щелью на две новые дочерние хромосомы.
В анафазе хромосомы отделяются друг от друга и расходятся к полюсам клетки со скоростью до 0,5 мкм/мин. В конце анафазы цитоплазматическая мембрана впячивается по экватору клетки перпендикулярно ее продольной оси, образуя борозду деления.
В телофазе хромосомы, разошедшиеся к полюсам клетки, деконденсируются и начинается транскрипция (продукция) РНК. Образуются ядерная оболочка, ядрышко, быстро формируются мембранные структуры будущих дочерних клеток. На поверхности клетки, по ее экватору, перетяжка углубляется, клетка разделяется на две дочерние клетки.
Благодаря митотическому делению дочерние клетки получают набор хромосом, идентичный материнскому. Митоз обеспечивает генетическую стабильность, увеличение числа клеток и, следовательно, рост организма, а также процессы регенерации.
Мейоз (от греч. meiosis - уменьшение) наблюдается у клеток-пред- шественниц половых клеток. В результате деления этих клеток образуются новые клетки с одинарным (гаплоидным) набором хромосом, что важно для передачи генетической информации. При слиянии одной половой клетки с клеткой противоположного пола во время оплодотворения набор хромосом удваивается и становится полным, двойным (диплоидным). В образовавшейся после слияния половых клеток диплоидной (двуядерной) зиготе оказывается два набора одинаковых (гомологичных) хромосом. Одна пара гомологичных хромосом клетки происходит из ядра яйцеклетки, другая - из ядра сперматозоида.
В результате мейоза половых клеток в зрелом организме в каждую дочернюю клетку попадает лишь по одной из всех пар гомологичных хромосом исходных клеток. Это становится возможным, поскольку при мейозе происходит лишь репликация ДНК и два последовательных деления ядер. В результате из одной диплоидной клетки образуются две гаплоидные клетки. В каждой из таких дочерних клеток содержится вдвое меньше хромосом (23), чем в ядре материнской клетки (46). В результате мейоза гаплоидные половые клетки имеют не только уменьшенное вдвое число хромосом, но и иное расположение женских и мужских (отцовских и материнских) генов в хромосомах. В связи с этим новый организм не просто несет те или иные признаки своих родителей, но и имеет собственные индивидуальные черты.
Сравнительная характеристика митоза и мейоза представлены в табл. 3.
Таблица 3. Сравнительная характеристика митоза и мейоза
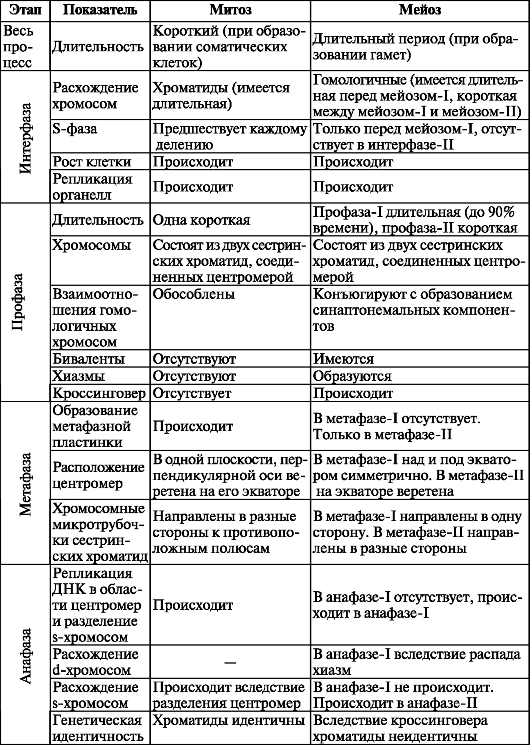
Окончание таблицы 3

ТКАНИ
Клетки и их производные объединяются в ткани. Ткань - это сложившаяся общность клеток и межклеточного вещества, объединенных единством происхождения, строения и функции. В организме человека выделяют 4 типа тканей: эпителиальную, соединительную, мышечную и нервную. Каждая ткань развивается из определенного зародышевого листка. Эпителиальная ткань происходит из энто-, экто- и мезодермы. Соединительные и мышечные ткани образуются из мезодермы (кроме мышц радужки и миоэпителиоцитов, происходящих из эктодермы). Нервная ткань развивается из эктодермы.
ЭПИТЕЛИАЛЬНАЯ ТКАНЬ
Эпителиальная ткань (tdxtus epithelialis) покрывает поверхности тела, выстилает слизистые оболочки, отделяя организм от внешней среды (покровный эпителий), а также образует железы (железистый эпителий). Кроме того, выделяют сенсорный эпителий, клетки которого воспринимают специфические раздражения в органах слуха, равновесия и вкуса. Некоторые авторы называют нейросенсорным эпителием измененные нервные клетки, воспринимающие световые и обонятельные раздражения.
Классификация эпителиев. В зависимости от положения относительно базальной мембраны покровный эпителий подразделяют на простой (однослойный) и многослойный (рис. 11, табл. 4). Все клетки простого (однослойного) эпителия лежат на базальной мембране и образуют один клеточный слой. У многослойного эпителия клетки образуют несколько слоев и лишь клетки нижнего (глубокого) слоя лежат на базальной мембране. Простой (однослойный) эпителий, в свою очередь, подразделяется на однорядный,
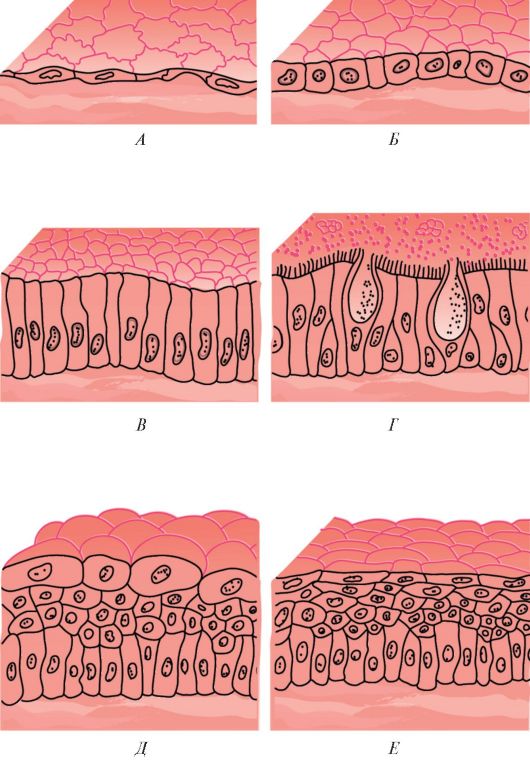
Рис. 11. Строение покровного эпителия: А - простой сквамозный (плоский ) эпителий (мезотелий); Б - простой кубический эпителий; В - простой столбчатый эпителий; Г - реснитчатый эпителий; Д - переходный эпителий; Е - неороговевающий многослойный (плоский) сквамозный эпителий
Таблица 4. Характеристика типов эпителия
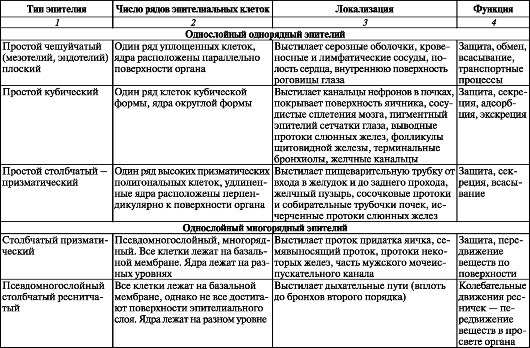
Окончание таблицы 4
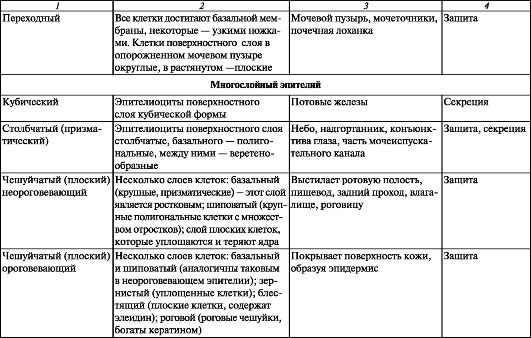
Таблица 5.
или изоморфный (плоский, кубический, столбчатый), и псевдомногослойный (многорядный). У однорядного эпителия ядра всех клеток эпителиального пласта расположены на одном уровне и все клетки имеют одинаковую высоту. У многорядного эпителия ядра клеток расположены на разных уровнях. В зависимости от формы клеток и их способности к ороговению различают неороговевающий многослойный (плоский) чешуйчатый эпителий и ороговевающий многослойный (плоский) чешуйчатый эпителий.
Эпителиоциты отличаются большим разнообразием формы и размеров. В зависимости от формы клеток различают следующие виды эпителиоцитов: чешуйчатые (плоские), кубические, столбчатые (призматические), реснитчатые, жгутиковые, микроворсинчатые. Кроме того, имеются пигментные и секреторные (железистые) эпителиоциты.
Строение клеток различных видов эпителия неодинаково. Однако все они имеют общие структурные особенности. Эпителиоциты полярны - их апикальная часть отличается от базальной. За редким исключением (атипичный эпителий) они образуют пласт, который располагается на базальной мембране и лишен кровеносных сосудов. В эпителиоцитах имеются все описанные выше органеллы общего назначения, их развитие зависит от выполняемой клеткой функции. Так, клетки, секретирующие белок, богаты элементами зернистой эндоплазматической сети, клетки, продуцирующие стероиды, - элементами незернистой эндоплазматичес- кой сети. И в тех и в других хорошо развит комплекс Гольджи. Всасывающие клетки имеют множество микроворсинок, а эпителиоциты, покрывающие слизистую оболочку дыхательных путей, - реснички.
Эти особенности приводятся ниже при описании различных эпителиев.
Покровный эпителий выполняет многочисленные функции. Это в первую очередь барьерная и защитная функции, которые осуществляют все виды эпителия, а также внешний обмен, всасывание (однослойный эпителий тонкой кишки, эпителий - мезотелий брюшины, плевры, эпителий канальцев нефрона и др.), секреция (клетки амниотического эпителия, эпителия сосудистой полоски улиткового лабиринта, большие (гранулярные) альвеолоциты, экскреция (эпителий канальцев нефрона), газообмен (дыхательные альвеолоциты), моторика (осуществляют рес- нички и жгутики).
Некоторые виды эпителия у человека утратили свойство пограничности, например, эпителий желез внутренней секреции.
Подробная морфофункциональная характеристика покровного и железистого эпителиев приводится ниже.
Однослойные эпителии. Простой чешуйчатый (плоский) эпителий
представляет собой пласт тонких, плоских клеток, лежащих на базальной мембране. Лишь в зоне залегания ядер имеются выпячивания свободной поверхности клетки. Эпителиоциты имеют полигональную форму, границы между ними видны при импрегнации солями серебра при световой микроскопии. Плоские эпителиоциты покрывают поверхности серозных оболочек (мезотелий), образуют наружную стенку капсулы почечных клубочков, задний эпителий роговицы. Такие клетки выстилают просветы всех кровеносных и лимфатических сосудов и полостей сердца (эндотелий), просвет альвеол (респираторные эпителиоциты). У некоторых органов простой чешуйчатый (плоский) эпителий лишен ресничек, но имеет большее или меньшее количество микроворсинок. Например, задний эпителий роговицы глаза имеет лишь единичные микроворсинки, располагающиеся над ядром.
Мезотелиоциты, покрывающие серозные оболочки (брюшину, плевру, перикард), имеют полигональную форму, очень тонкую цитоплазму. Их свободная поверхность покрыта множеством микроворсинок, у некоторых клеток имеется по 2-3 ядра. В цитоплазме содержатся единичные митохондрии, небольшое количество элементов зернистой эндоплазматической сети и комплекса Гольджи. Мезотелиоциты облегчают взаимное скольжение внутренних органов и предотвращают образование спаек между ними.
Эндотелиоциты - это уплощенные, удлиненные, иногда веретено- образные клетки с очень тонким слоем цитоплазмы. Ядросодержащая часть клетки утолщена, в результате тело клетки слегка выбухает в просвет сосуда. Клетки соединены между собой простыми (зубчатыми) и сложными межклеточными соединениями (запирающими зонами). Микроворсинки расположены, в основном, над ядром. Цитоплазма содержит микропиноцитозные пузырьки, единичные митохондрии, элементы зернистой эндоплазматической сети и комплекса Гольджи.
Респираторные (дыхательные) эпителиоциты имеют большие размеры (50-100 мкм), их цитоплазма богата микропиноцитозными пузырьками и рибосомами. Прочие органеллы представлены слабо.
Простой кубический эпителий образован одним слоем клеток гексагональной формы, имеющих на срезах, перпендикулярных поверхности, форму, близкую к квадрату. В центре клетки расположено округлое ядро. Апикальная поверхность клетки покрыта микроворсинками. Особенно много микроворсинок на апикальной стороне эпителиоцитов сосудистого сплетения. Различают безресничные кубические эпителиоциты
(у некоторых собирательных трубочек почки, дистальных прямых канальцев нефронов, желчных проточков, сосудистых сплетений мозга, пигментного эпителия сетчатки глаза и др.) и реснитчатые (у терминальных и респираторных бронхиол, эпендимоцитов, выстилающих полости желу- дочков мозга). Передний эпителий хрусталика также представляет собой простой кубический эпителий. Поверхность этих клеток гладкая.
Пигментные эпителиоциты несут на апикальной стороне крупные выросты, содержащие веретенообразные гранулы меланина.
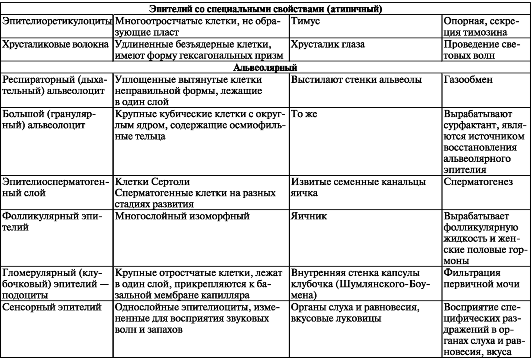
Простой столбчатый (призматический) эпителий широко распространен в организме человека. Он покрывает слизистую оболочку желудочно-кишечного тракта от входа в желудок до заднего прохода.
Столбчатые эпителиоциты - высокие, узкие, призматические, многоугольные или округлые клетки, плотно связанные друг с другом комплексом межклеточных соединений, которые расположены вблизи поверхности. Округлое или эллипсоидное ядро обычно располагается в нижней трети клетки. Столбчатые эпителиоциты зачастую имеют множество микроворсинок, стереоцилий или ресничек (рис. 12). В цитоплазме находятся многочисленные митохондрии, хорошо развитый аппарат Гольджи, элементы незернистой и зернистой эндоплазматической сети. Микроворсинчатые клетки превалируют в эпителии слизистой
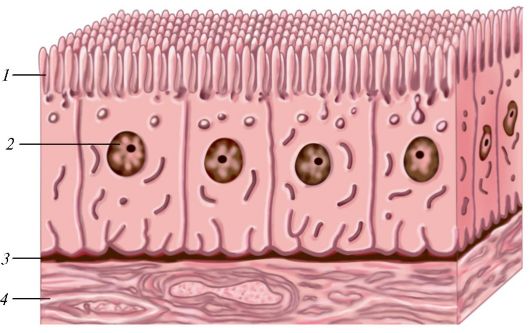
Рис. 12. Строение столбчатых эпителиальных клеток: 1 - микроворсинки; 2 - ядро эпителиоцита; 3 - базальная мембрана; 4 - соеди- нительная ткань (по В.Г. Елисееву и др.).
оболочки кишечника и желчного пузыря. В слизистой оболочке этих органов, помимо микроворсинчатых клеток, имеется множество бокаловидных экзокриноцитов, вырабатывающих слизь. Стенки сосочковых протоков и собирательных трубочек почек и исчерченных протоков слюнных желез также образованы столбчатыми эпителиоцитами, микроворсинок у которых немного. Реснитчатые эпителиоциты в большом количестве встречаются у слизистой оболочки бронхов третьего порядка, бронхиол, матки и маточных труб.
Псевдомногослойный (многорядный) эпителий образован преимущественно высокими клетками с овальными ядрами, которые располагаются на различных уровнях. Все клетки лежат на базальной мембране, но не все они достигают просвета органа. У эпителия этого типа различают 4 вида клеток:
- высокодифференцированные поверхностные эпителиоциты - удлиненные клетки, достигающие просвета органа. Эти клетки имеют округлое ядро и хорошо развитые органеллы, особенно комплекс Гольджи и эндоплазматическую сеть. Их апикальная цитолемма образует микроворсинки, стереоцилии или реснички. Реснитчатые клетки покрывают слизистую оболочку носа, трахеи, бронхов. Безресничные клетки покрывают слизистую оболочку части мужской уретры, выводных протоков многих желез, протоки придатка яичка и семявыносящий проток;
- вставочные эпителиоциты, удлиненные, малодифференцированные, лишенные ресничек и микроворсинок и не достигающие просвета. Эти клетки расположены между поверхностными клетками и соединены с ними межклеточными соединениями;
- базальные эпителиоциты, образующие самый глубокий ряд клеток. Они являются источником обновления эпителия (ежедневно до 2% клеток популяции);
- бокаловидные экзокриноциты, богатые гранулами слизи, залегающие между реснитчатыми клетками.
В эпителии протоков придатка яичка и семявыносящего протока имеются клетки лишь двух видов: поверхностные (со стереоцилиями) и базальные (лишенные ресничек и микроворсинок).
Многослойные эпителии. Неороговевающий многослойный (плоский) эпителий (рис. 13) состоит из трех слоев клеток, среди которых различают базальный, промежуточный чешуйчатый (шиповатый) и поверхностный:
- базальный слой образован относительно крупными призматическими или полиэдрическими клетками, которые прикреплены к базальной мембране с помощью многочисленных полидесмосом;
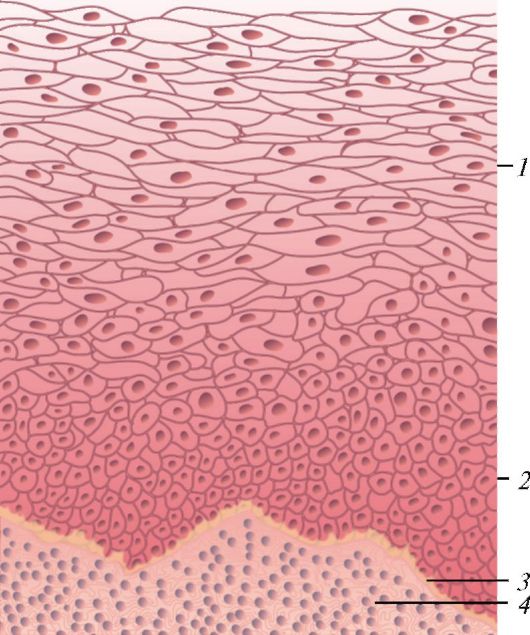
Рис. 13. Многослойный неороговевающий плоский (сквамозный) эпителий: 1 - поверхностный слой; 2 - шиповатый слой; 3 - базальный слой; 4 - подлежащая соединительная ткань (по В.Г. Елисееву и др.).
- шиповатый (промежуточный) слой образован крупными отростчатыми полигональной формы клетками, отростки которых связаны между собой многочисленными десмосомами, а цитоплазма богата тонофи- ламентами;
- поверхностный слой образован плоскими клетками, многие из которых лишены ядра. Однако эти клетки остаются связанными между собой с помощью десмосом.
Оба первых слоя образуют ростковый (герминативный) слой. Эпителиоциты делятся митотически и, продвигаясь наверх, уплощаются и заменяют слущивающиеся клетки поверхностного слоя. Самые поверхностные клетки превращаются в тонкие чешуйки, которые теряют связи друг с другом и отпадают. Свободная поверхность многих клеток покрыта короткими микроворсинками и мелкими складками. Эпителий этого типа покрывает слизистую оболочку ротовой полости, пищевода, влагалища, голосовых складок, переходной зоны анального канала, женской уретры,
Рис. 14. Строение многослойного чешуйчатого ороговевающего эпителия: 1 - роговые чешуйки; 2 - роговой слой; 3 - блестящий слой; 4 - зернистый слой; 5 - шиповатый слой; 6 - базальный слой; 7 - меланоцит; 8 - межклеточные щели; 9 - базальная мембрана (по Р. Крстичу, с изменениями)
а также образует передний эпителий роговицы. Иными словами, неороговевающий многослойный плоский эпителий покрывает поверхности, постоянно увлажняемые секретом желез, расположенных в субэпители- альной рыхлой неоформленной соединительной ткани.
Ороговевающий многослойный (плоский) чешуйчатый эпителий покрывает всю поверхность кожи, образуя ее эпидермис (рис. 14). У эпидермиса кожи выделяют пять слоев: базальный, шиповатый, зернистый, блестящий, роговой:
- в базальном слое располагаются клетки призматической формы, которые имеют множество мелких отростков, окруженных базальной мембраной. В цитоплазме, расположенной над ядром, находятся гранулы меланина. Между базальными эпителиоцитами залегают пигментсодержащие клетки - меланоциты;
- шиповатый слой образован несколькими слоями крупных полигональных шиповатых эпителиоцитов, соединенных между собой множеством десмосом, располагающихся на отростках. Цитоплазма богата тонофибриллами и тонофиламентами. Оба описанных слоя формируют ростковый слой, клетки которого делятся митотически и продвигаются наверх;
- зернистый слой состоит из чешуйчатых (плоских) эпителиоцитов, богатых гранулами кератогиалина. По мере увеличения его количества клетки медленно дегенерируют;
- блестящий слой обладает сильной светопреломляющей способностью из-за чешуйчатых (плоских) эпителиоцитов, содержащих элеидин;
- роговой слой образован слущивающимися роговыми чешуйками.
Переходный эпителий изменяет свою форму в зависимости от функционального состояния органа. Переходный эпителий, покрывающий слизистую оболочку почечных лоханок, мочеточников, мочевого пузыря, начала мочеиспускательного канала, меняет свою форму в зависимости от состояния органа. При растяжении стенок органов эти эпителиоциты становятся плоскими, а их цитоплазматическая мембрана растягивается. При расслаблении стенок органов клетки становятся вы- сокими. Поверхностные клетки полиплоидные, они имеют одно крупное или два мелких ядра. В апикальной части этих клеток расположены комплекс Гольджи, многочисленные, окруженные мембраной веретенообразные везикулы и микрофиламенты. Веретенообразные пузырьки, по-видимому, происходят из комплекса Гольджи. Они подходят к цитолемме, как бы сливаются с ней. У растянутого (наполненного) мочевого пузыря эпителиальный покров не прерывается. Эпителий остается непроницаемым для мочи и надежно предохраняет мочевой пузырь от ее
всасывания. Это обеспечивается, с одной стороны, плотными контактами между клетками (десмосомы) и многочисленными интердигитациями цитолемм соседних клеток, а с другой - множеством утолщений на наружной поверхности цитоплазматической мембраны за счет плотного вещества неизвестной природы - «бляшек», к которым изнутри клетки подходит множество нитей наподобие якорей. При расслаблении стенки пузыря цитоплазматическая мембрана поверхностных клеток становится складчатой, сгибаясь в участках между бляшками. В клетках имеются митохондрии, свободные рибосомы и включения гликогена. Под поверхностным слоем лежат клетки, имеющие форму теннисных ракеток с узкими ножками, соприкасающимися с базальной мембраной. Эти клетки имеют крупное ядро неправильной формы, в цитоплазме расположены митохондрии, умеренное количество элементов эндоплазматической сети и комплекса Гольджи. Непосредственно на базальной мембране лежат мелкие клетки с ядрами неправильной формы и малым количеством органелл. У пустого мочевого пузыря клетки высокие, на препарате видно до 8-10 рядов ядер; у наполненного (растянутого) клетки уплощены, число рядов ядер не превышает 2-3, цитолемма поверхностных клеток гладкая.
Многослойный кубический эпителий образован несколькими (от 3 до 10) слоями клеток. Поверхностный слой представлен клетками кубической формы. Клетки имеют микроворсинки и богаты гранулами гликогена. Под ними расположено несколько слоев удлиненных клеток веретенообразной формы. Непосредственно на базальной мембране лежат полигональные или кубические клетки. Все клетки соединены между собой зубчатыми и пальцевидными межклеточными соединениями, а клетки поверхностного слоя - сложными соединениями. Эпителий этого типа встречается редко. Он расположен небольшими участками на коротком протяжении между многорядным призматическим и многослойным плоским неороговевающим эпителиями (слизистая оболочка задней части преддверия полости носа, надгортанник, часть мужского мочеиспускательного канала, выводные протоки потовых желез).
Многослойный столбчатый эпителий также состоит из нескольких слоев клеток (3-10). Поверхностные эпителиоциты имеют призматическую форму и зачастую несут на своей поверхности реснички. Глубже лежащие эпителиоциты полиэдрические и кубические. Эпителий этого типа встречается в некоторых участках выводных протоков слюнных и молочных желез, слизистой оболочки глотки, гортани и мужского мочеиспускательного канала.
Железистый эпителий. Клетки железистого эпителия (гландулоциты) образуют паренхиму многоклеточных желез и одноклеточные железы. Железы подразделяются на экзокринные, имеющие выводные протоки, и эндокринные, не имеющие выводных протоков и выделяющие синтезируемые ими продукты непосредственно в межклеточные пространства, откуда они поступают в кровь и лимфу; смешанные железы состоят из экзо- и эндокринных отделов (например, поджелудочная железа). Экзокриноциты выделяют синтезируемые ими продукты на поверхность органов (пищевода, кишки, желудка и др.), кожных покровов тела.
Во время эмбрионального развития в определенных участках покровного эпителия дифференцируются клетки, которые впоследствии специализируются на синтезе веществ, подлежащих секреции. Некоторые из этих клеток остаются в пределах эпителиального пласта, образуя эн-
доэпителиальные железы, другие усиленно делятся митотически и врастают в подлежащую ткань, формируя экзоэпителиальные железы. Одни железы сохраняют связь с поверхностью благо- даря протоку - это экзокринные железы, другие в процессе развития теряют эту связь и становятся эндокринными железами.
Экзокринные железы подразделяются на одноклеточные и многоклеточные (табл. 5).
Одноклеточные (экзокринные) железы. В организме человека имеется множество одноклеточных бокаловидных экзокриноцитов, лежащих среди других эпителиальных клеток, покрывающих слизистую оболочку полых органов пищеварительной, дыхательной и
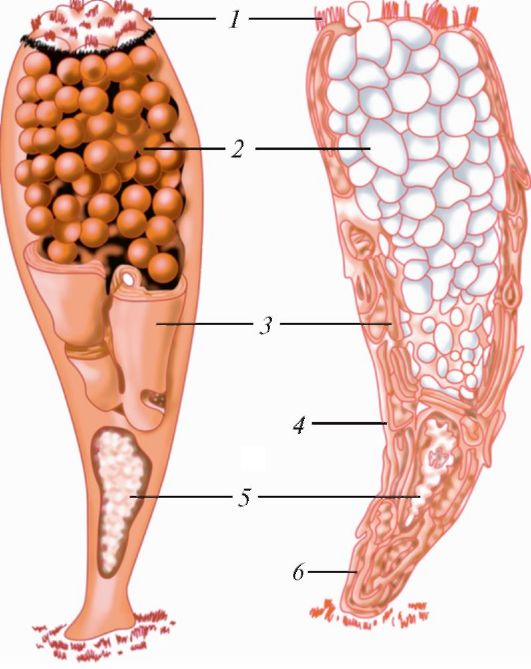
Рис. 15. Строение железистой клетки - бокаловидного экзокриноцита: 1 - клеточные микроворсинки; 2 - гранулы слизистого секрета; 3 - внутренний сетчатый аппарат; 4 - митохондрия; 5 - ядро; 6 - зернистая эндоплазматическая сеть
Таблица 5. Классификация экзокринных желез
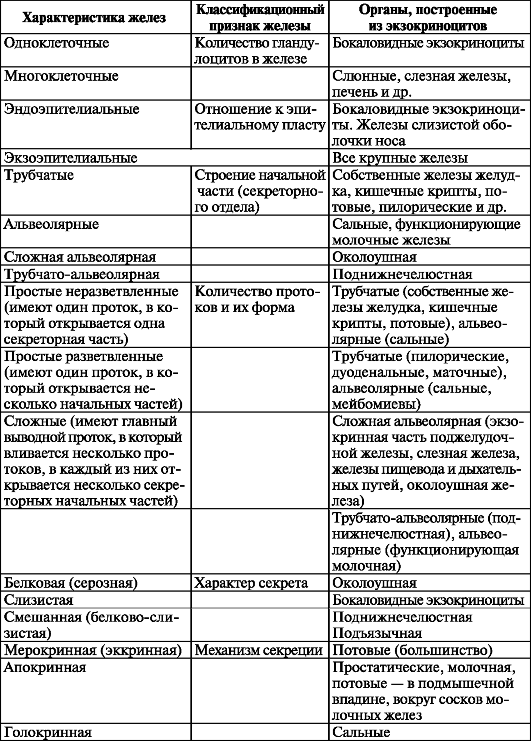
половой систем (рис. 15). Железы вырабатывают слизь, которая состоит из гликопротеидов. Структура бокаловидных клеток зависит от фазы секреторного цикла. Функционально активные клетки по форме напоми- нают бокал. Узкое, богатое хроматином ядро прилежит к базальной части клетки (ножке). Над ядром расположен хорошо развитый комплекс Гольджи, над которым в расширенной части клетки находятся конденсирующие вакуоли или просекреторные гранулы, а также множество секреторных гранул, выделяющихся из клетки по мерокриновому типу. После выделения секреторных гранул клетка становится узкой, на ее апикальной поверхности видны микроворсинки.
В процессе синтеза и образования слизи участвуют рибосомы, эндоплазматическая сеть, комплекс Гольджи. Белковый компонент синтезируется полирибосомами зернистой эндоплазматической сети, которая в большом количестве находится в базальной части клетки, и с помощью транспортных пузырьков переносится в комплекс Гольджи. Углеводный компонент синтезируется комплексом Гольджи, здесь же происходит и связывание белков с углеводами. В комплексе Гольджи формируются пресекреторные гранулы, которые отделяются и превращаются в секреторные. Количество гранул увеличивается в направлении к апикальной поверхности клетки. Секреция гранул слизи из клетки на поверхность слизистой оболочки, как правило, осуществляется путем экзоцитоза.
Многоклеточные железы. Экзокриноциты образуют начальные секреторные отделы экзокринных многоклеточных желез, которые вырабатывают различные секреты, и их трубчатых протоков, через которые секрет выделяется. Морфология экзокриноцитов зависит от характера секреторного продукта и фазы секреции. Железистые клетки структурно и функционально поляризованы. Их секреторные капли или гранулы сосредоточены в апикальной (надъядерной) зоне и выделяются в просвет через покрытую микроворсинками апикальную цитолемму. Клетки богаты митохондриями, элементами комплекса Гольджи и эндоплазматической сети. Зернистая сеть преобладает в белоксинтезирующих клетках (например, экзокринные панкреатоциты, гландулоциты околоушной железы), незернистая сеть - в клетках, синтезирующих липиды или углеводы (гепатоциты, корковые эндокриноциты надпочечников). Клетки в области их верхушек соединены между собой сложными межклеточными соединениями, между боковыми поверхностями базальных частей имеются широкие межклеточные щели. Базальная цитолемма нередко складчатая.
Синтез белка и выведение секреторного продукта представляют сложный процесс, в котором участвуют различные клеточные структуры: полирибосомы и эндоплазматическая (зернистая) сеть, комплекс Гольджи, секреторные гранулы, цитоплазматическая мембрана. Секреторный процесс происходит циклично, в нем выделяют четыре фазы (Pallade G., 1975). В первой фазе в клетку поступают необходимые для синтеза вещества. В базальной части белоксинтезирующих клеток хорошо видны многочисленные микропиноцитозные пузырьки. Во второй фазе происходит синтез веществ, которые с помощью транспортных пузырьков перемещаются к формирующейся поверхности комплекса Гольджи и сливаются с ней. В комплексе Гольджи вещества, подлежащие секреции (например, белки), сначала накапливаются в конденсирующих вакуолях умеренной электронной плотности, в которых белки концентрируются. В результате конденсирующие вакуоли превращаются в отделенные от комплекса Гольджи электронноплотные секреторные гранулы, расположенные между хорошо выраженными цистернами зернистой эндоплазматической сети. Секреторные гранулы передвигаются в апикальном направлении. В третьей фазе секреторные гранулы выделяются из клетки. В четвертой фазе секреции экзокриноцит восстанавливается.
Возможны три способа выделения секрета. При мерокриновом (эккриновом) секреторные продукты выделяются путем экзоцитоза. Этот способ наблюдается у серозных (белковых) желез. При этом структура клеток не нарушается. Апокриновый способ (например, лактоциты) сопровождается разрушением апикальной части клетки (макроапокриновый тип) либо верхушек микроворсинок (микроапокриновый тип). При голокриновом способе секреции гландулоциты полностью разрушаются и их цитоплазма входит в состав секрета (например, сальные железы).
В зависимости от строения начального (секреторного) отдела различают трубчатые (напоминают трубку), ацинозные (напоминают грушу) и альвеолярные (напоминают шарик), а также трубчато-ацинозные и трубчато-альвеолярные железы, начальные отделы которых имеют и ту, и другую форму (рис. 16).
В зависимости от строения протоков железы подразделяются на простые, имеющие простую, неразветвленную или малоразветвленную форму, и сложные, имеющие несколько начальных (секреторных) отделов. Простые железы подразделяются на простые неразветвленные, имеющие вид трубочки, груши или шарика, и простые разветвленные, имеющие
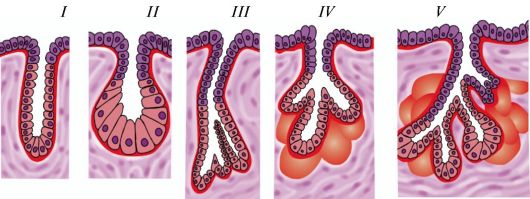
Рис. 16. Типы экзокринных желез: I - простая трубчатая железа с неразветвленным начальным отделом; II - простая альвеолярная железа с неразветвленным начальным отделом; III - простая трубчатая железа с разветвленным начальным отделом; IV - простая альвеолярная железа с разветвленным начальным отделом; V - сложная альвеолярно-трубчатая железа с разветвленными начальными отделами (по И.В. Алмазову и Л.С. Сутулову)
вид раздвоенной или утроенной трубочки, или ацинуса, или альвеолы. К простым трубчатым неразветвленным железам относятся собственные железы желудка, кишечные крипты, потовые железы, к простым альве- олярным неразветвленным - сальные. Простые трубчатые разветвленные - это пилорические, дуоденальные и маточные железы, простые альвеолярные разветвленные - мейбомиевы железы.
Сложные железы подразделяются на трубчатые (железы полости рта), трубчато-ацинарные (экзокринная часть поджелудочной железы, слез- ные, околоушные, крупные железы пищевода и дыхательных путей); трубчато-альвеолярные (поднижнечелюстная) и альвеолярные (функционирующая молочная железа). Железы вырабатывают белковый секрет (серозные железы), слизь (слизистые) или смешанный секрет.
Секреция липидов сальными железами включает синтез, накопление и выделение жирных кислот, триглицеридов, холестерола и его эфиров. В этом процессе участвуют незернистая эндоплазматическая сеть, комплекс Гольджи, митохондрии. В клетках сальных желез вместо типичных секреторных гранул имеются липидные капли. Первичные липидные вещества появляются в пределах везикул комплекса Гольджи, количество везикул увеличивается. Они образуют липидные капли, некоторые из них ограничены тонкой мембраной. Капли окружены элементами незернистой цитоплазматической сети.
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
Соединительные ткани (textus connectivus) представляют обширную группу, включающую собственно соединительные ткани (рыхлую волокнистую и плотную волокнистую, неоформленную и оформленную), ткани со специальными свойствами (ретикулярную, пигментную, жировую), твердые, скелетные (костные и хрящевые) и жидкие (кровь) (табл. 6). Соединительные ткани выполняют опорную, защитную (механическую) функции (плотная волокнистая соединительная ткань, хрящ, кость), а также трофическую (питательную), защитную (фагоцитоз) функции (рыхлая волокнистая и ретикулярная соединительные ткани, кровь). Соединительные ткани сформированы из многочисленных клеток и межклеточного вещества, а также различных волокон (коллагеновых, эластических, ретикулярных). Межклеточное вещество кости твердое, крови - жидкое.
Все названные виды соединительной ткани являются производными мезенхимы (от греч. mesos - средний, enchyma - налитое), которая, в свою очередь, образуется из мезодермы. Клетки мезенхимы образуют тонкие отростки, связывающие их между собой, формируется трехмерная сеть, в которой образуются кровеносные капилляры. Мезенхимные клетки имеют звездчатую форму, они слабо дифференцированы. Крупное, бедное хроматином ядро содержит большое ядрышко, обычно лежащее вблизи нуклеолеммы. Число органелл невелико. Это митохондрии, элементы зернистой эндоплазматической сети, умеренно развитый комплекс Гольджи, небольшое количество свободных рибосом, единичные лизосомы и липидные гранулы. В жидком аморфном межклеточном веществе умеренной электронной плотности находятся единичные микрофибриллы. Клетки активно митотически делятся. Во время деления они округляются, но их связь с окружающими клетками сохраняется благодаря тонким отросткам. Дочерние клетки принимают исходную звездчатую форму.
Делящиеся клетки образуют отдельные скопления, из которых дифференцируются стволовые клетки крови, фибробласты (превращаются в фиброциты), остеобласты (превращаются в остеоциты), одонтоблас- ты, хондробласты (превращаются в хондроциты), тучные клетки, адипоциты, ретикулярные клетки, макрофаги оседлые и блуждающие, эндотелиоциты, гладкие миоциты, миосимпласты. Таким образом, мезенхимные клетки являются полипотентными.
Слизистая соединительная ткань (textus mucosus) имеется только у зародыша, поэтому ее относят к эмбриональным. Она входит в состав пупочного канатика и пластинки хориона, окружает кровеносные сосуды.
Таблица 6. Классификация и характеристика видов соединительной ткани

Продолжение таблицы 6.
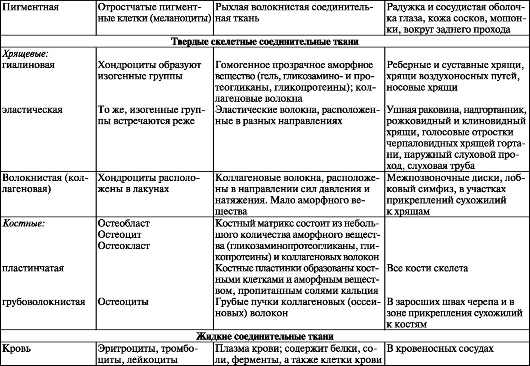
Слизистая ткань пупочного канатика (вартонов студень) образована слизистыми клетками (называемыми мукоцитами), которые имеют отростчатую форму и напоминают мезенхимные, и межклеточным веществом, окрашиваемым толуидиновым синим метахроматично в розовый цвет из-за большого количества мукополисахаридов. В петлях, образуемых клетками слизистой ткани, проходят тонкие коллагеновые волокна. Многоотростчатые клетки формируют трехмерную сеть. Переплетающиеся пучки коллагеновых микрофибрилл обусловливают прочность пупочного канатика, а способность гиалуроновой кислоты связывать воду обеспечивает тургор и препятствует сдавлению сосудов при перекручивании пупочного канатика. С увеличением возраста плода увеличивает- ся количество коллагеновых волокон в слизистой ткани.
СОБСТВЕННО СОЕДИНИТЕЛЬНЫЕ ТКАНИ
Рыхлая волокнистая соединительная ткань (textus connectivus fibrdsus laxus) располагается преимущественно по ходу кровеносных и лимфатических сосудов, нервов, образует строму большинства внутренних орга- нов, собственную пластинку слизистой оболочки, подслизистую и подсерозную основы, адвентициальную оболочку. Рыхлая волокнистая соединительная ткань содержит многочисленные собственные и пришлые клетки: фибробласты, фиброциты, ретикулярные, перициты, макрофагоциты, тучные клетки (тканевые базофилы), плазмоциты, адипоциты, пигментные клетки, лимфоциты, лейкоциты, которые располагаются в межклеточном веществе (рис. 17). Межклеточное вещество продуцируется в основном фибробластами и представлено коллагеновыми, эласти- ческими, ретикулярными волокнами и основным (аморфным) веществом.
Тканевые базофилы (тучные клетки) лежат вблизи капилляров, перициты окружают капилляры, располагаясь в их базальной мембране. Тонкие микрофибриллы переходят из одного коллагенового волокна в другое. Эластические волокна, придающие рыхлой волокнистой соединительной ткани эластические свойства, широко разветвляются и анастомозируют между собой. Ретикулярные волокна входят в состав стромы лимфоидных (иммунных) органов. Пространства между всеми описанными структурами заняты аморфным веществом.
Фибробласты (fibroblastus, от греч. fibra - волокно, blastos - зародыш) - основные специализированные фиксированные клетки соединительной ткани, которые являются главными продуцентами межклеточного вещества (рис. 18). Количество фибробластов варьирует в соединительной
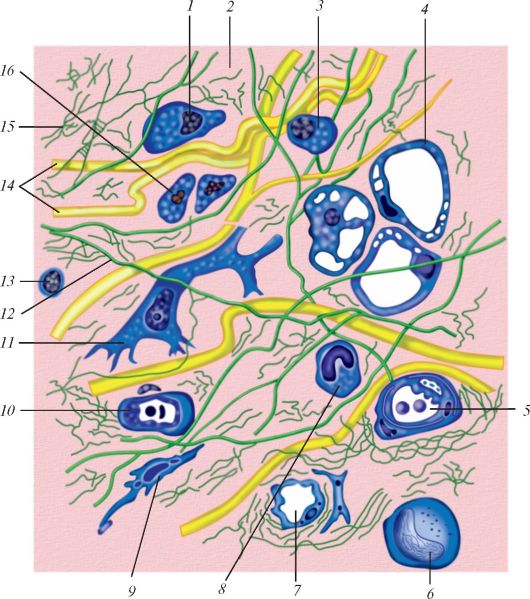
Рис. 17. Схема строения рыхлой волокнистой соединительной ткани: 1 - макрофагоцит; 2 - аморфное межклеточное (основное) вещество; 3 - плазмоцит (плазматическая клетка); 4 - липоцит (жировая клетка); 5 - кровеносный сосуд; 6 - моноцит; 7 - лимфатический капиляр; 8 - эозинофильный гранулоцит; 9 - фиброцит; 10 - кровеносный капилляр; 11 - фибробласт; 12 - эластическое волокно; 13 - лимфоцит; 14 - коллагеновые волокна ; 15 - ретикулярные волокна; 16 - тканевый базофил
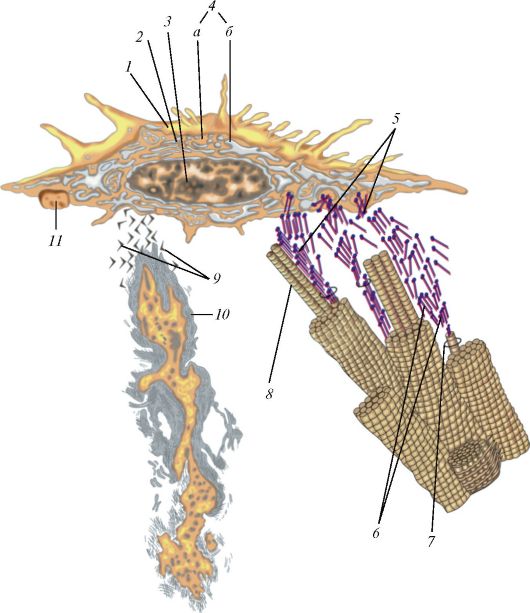
Рис. 18. Схема строения фибробласта и образования межклеточного вещества: 1 - фибробласт; 2 - сетчатый аппарат; 3 - ядро; 4 - эндоплазматическая сеть: а - незернистая, б - зернистая; 5 - молекулы тропоколлагена, выделяющиеся из клетки; 6 - полимеризация молекул тропоколлагена в профибриллы; 7 - микрофибриллы; 8 - фибриллы; 9 - молекулы эластина; 10 - микрофибриллярный структурный гликопротеин; 11 - эластичное волокно, погруженное в мембрану фибробласта (по Р. Крстичу, с изменениями)
ткани различных типов. Фибробластов особенно много в рыхлой волокнистой соединительной ткани. Фибробласты имеют овальное ядро, заполненное мелкими глыбками хроматина, четко различимым ядрышком и базофильной цитоплазмой, с множеством свободных и прикрепленных рибосом. Базофилия более выражена в молодых клетках, которые способны делиться. От поверхности клетки отходят мелкие цитоплазматические отростки и короткие микроворсинки. Под нуклеолеммой располагается тонкая зона гетерохроматина, имеются крупные ядрышки (1-2). Очень хорошо развита зернистая эндоплазматическая сеть. Многие ее цистерны расширены и заполнены гомогенным веществом умеренной электронной плотности. В цитоплазме много свободных рибосом. Сетчатый аппарат развит хорошо, видно множество вакуолей, пузырьков, расширенных мешочков. Обнаруживаются удлиненные мембранные гранулы, содержащие фибриллярный материал. В цитолемму фибробласта встроены многочисленные рецепторы. К наружной поверхности цитолеммы фиксированы коллаген I типа, а также гликопротеин фибронектин. Коллаген и фибронектин образуют на клеточной поверхности ячеистую сеть, которая служит своеобразным цитоскелетом и играет важную роль в процессах фибриллогенеза. По всей цитоплазме располагаются филаменты толщиной 5-7 нм. На внутренней поверхности цитолеммы располагаются микропиноцитозные пузырьки, окаймленные ямки, свидетельствующие об интенсивном эндоцитозе. Цитоплазму фибробласта заполняет трехмерная микротрабекулярная сеть, образованная тонкими белковыми филаментами (5-7 нм), которые соединяют между собой актиновые, миозиновые и промежуточные филаменты. Движения фибробластов связаны с актиновыми филаментами.
Межклеточное вещество рыхлой волокнистой соединительной ткани, которое синтезируют и секретируют фибробласты, образовано волокнами и бесструктурным основным веществом (аморфный компонент межклеточного вещества). Основное вещество является вязким гелем, состоящим главным образом из макромолекул полисахаридов и большого количества тканевой жидкости, связанной с ними. Полисахариды представлены гликозаминогликанами - несульфатированными (гиалуроновая кислота) и сульфатированными. Гликозоаминогликаны (старое название мукополисахариды) - это длинные неразветвленные цепи полисахаридов, образованных многократно повторяющимися дисахаридами. Все сульфатированные гликозоаминогликаны связаны с белками и образуют протеогликаны (старое название мукопротеины). Гидратированные цепи
гликозоаминогликанов образуют гели, в которых легко совершается диффузия различных веществ, передвигаются клетки и их отростки. В гель, образованный гликозоаминогликанами, погружены различные фибриллярные структуры, основной из них является коллаген, который подразделяют на несколько типов.
Коллаген I типа составляет до 90% всего коллагена организма человека. Он образует толстые фибриллы, в нем мало гидроксилина и углеводов. Коллаген I типа находится в толстых волокнах собственно кожи (дермы), сухожилий, связок, фасций, матрикса кости, в синовиальной оболочке суставов, в стенках аорты, капсуле и строме печени; в склере, роговице, радужке; в дентине, пульпе зуба, периодонте; в волокнах, оплетающих легочные альвеолы; в соединительной ткани мышц кишечника.
Коллаген II типа образует тонкие фибриллы, в нем много углеводов и гидроксилина. Этот коллаген образует матрикс хряща. Он содержится в пульпозном ядре и фиброзном кольце межпозвоночных дисков; в стек- ловидном теле, десцеметовой оболочке, передней части роговицы; в клапанах сердца; в стенках бронхов, трахеи.
Коллаген III типа содержит мало гидроксилизина и углеводов, много гидроксиполина. Он находится в тонких ретикулярных волокнах сосочкового слоя кожи; в ретикулярных волокнах костного мозга; в тонких волокнах надхрящницы, фиброзных колец межпозвоночных дисков, в стенках аорты, ретикулярных волокнах, печени; в дентине, пульпе зубов, периодонте; в соединительной ткани гладких мышц кишечника.
Коллаген IV типа присутствует в базальных мембранах и хрусталике глаза.
Коллаген V типа в незначительных количествах имеется во многих тканях организма.
Во время секреции проколлагена из клетки на поверхности цитоплазматической мембраны фибробластов ферменты проколлагенпептидазы отщепляют концевые пептиды, в результате чего проколлаген превращается в тропоколлаген. Вблизи поверхности фибробласта молекулы тропоколлагена объединяются между собой во внеклеточном пространстве путем самосборки, образуя протофибриллы. 5-6 протофибрилл, соединяясь между собой с помощью боковых связей, образуют микрофибриллы толщиной около 10 нм. Микрофибриллы, в свою очередь, соединяются, образуя длинные поперечно исчерченные фибриллы толщиной до 300 нм, которые формируют коллагеновые волокна толщиной 1-20 мкм. Наконец, множество волокон, собираясь вместе, формируют коллагеновые пучки толщиной до 150 мкм. При этом на всех уровнях
организации коллаген имеет спиральное строение, что обеспечивает создание весьма прочных структур (Серов В.В., Шехтер А.Б., 1981).
Коллагеновые волокна при световой микроскопии выглядят как изогнутые тяжи, состоящие из фибрилл. При поляризационной микроскопии определяется их сильное двойное лучепреломление, которое при растяжении усиливается, а при высыхании и нагревании уменьшается. При электронной микроскопии определяются продольная периодичность, чередование темных и светлых полос.
Молекулы коллагена расположены в фибрилле параллельными рядами, ступенчато, причем соседние сдвинуты одна относительно другой на 1/4 длины (64 нм), а промежуток между концом одной и началом другой молекулы составляет 35 нм. На поперечном сечении видно, что каждая I и VI молекулы коллагена начинаются и заканчиваются на одном и том же уровне. Такое расположение молекул создает поперечную исчерченность, которая четко видна на негативно окрашенных препаратах: темные и светлые сегменты правильно чередуются, образуя периоды длиной 64 нм каждый. Благодаря одинаковому расположению заряженных и незаряженных остатков аминокислот (кроме пролина, гидроксипролина и лизина) с периодичностью 64 нм правильная поперечная ис- черченность сохраняется в фибрилле любой толщины.
Процессы фибриллогенеза регулируются гликозоаминогликанами, протеогликанами и гликопротеинами. Важная роль принадлежит самому фибробласту, который не только секретирует компоненты межклеточного вещества, но и создает его архитектонику как тканевой систе- мы. Направление волокон соединительной ткани соответствует длинной оси фибробластов, которые регулируют сборку и трехмерное расположение волокон и их пучков в межклеточном веществе (Серов В.В., Шех-
тер А.Б., 1981).
Эластические волокна, в отличие от коллагеновых, способны растягиваться в 1,5 раза, после чего возвращаются в исходное состояние. Эластические волокна анастомозируют и переплетаются между собой, образуя сети, фенестрированные пластины и мембраны. Эластические волокна образованы эластином, в котором отсутствует гидроксилизин, содержится мало гидроксипролина и очень много пролина и глицина. Молекулы проэластина синтезируются фибробластами и гладкими миоцитами на рибосомах зернистой эндоплазматической сети и секретируются во внеклеточное вещество. Микрофибриллы толщиной около 13 нм вблизи клеточной поверхности во внеклеточном пространстве образуют мелкопетлистую сеть, являющуюся каркасом, который заполняется
эластином. Зрелое эластическое волокно снаружи покрыто микрофибриллярным гликопротеином. Микрофибриллы имеются и в толще аморфного эластина. При световой микроскопии эластические волокна пред- ставляют собой лентовидные гомогенные тяжи, которые формируют правильную широкоплетистую сеть или окончатые мембраны. При поляризационной микроскопии видно их слабое двойное лучепреломление, которое при растяжении волокна уменьшается. При электронной микроскопии периодичность и поперечная исчерченность отсутствуют.
Ретикулярные волокна тонкие (от 100 нм до 1,5 мкм), разветвленные, анастомозирующие друг с другом, образуют мелкопетлистые сети, в ячейках которых расположены клетки. Ретикулярные волокна вместе с ретикулярными клетками образуют каркас (строму) органов кроветворения и иммунной системы, связывают базальные мембраны эпителия и миосимпластов с коллагеновыми волокнами подлежащей соединительной ткани, участвуют в образовании стромы печени, поджелудочной железы и других паренхиматозных органов, окружают капилляры, кро- веносные и лимфатические сосуды. При поляризационной микроскопии выявляется их уменьшенное двойное лучепреломление, при электронной микроскопии определяется продольная исчерченность с периодом 64 нм. Ретикулярные волокна располагаются в углублениях, образованных цитоплазмой ретикулярных клеток, ограниченных цитоплазматической мембраной. Ретикулярные волокна являются производными не только фибробластов, но и ретикулярных клеток. Каждое ре- тикулярное волокно содержит множество фибрилл диаметром 30 нм с поперечной исчерченностью, такой как у коллагеновых фибрилл. Химический состав ретикулярных волокон не отличается от коллагеновых, однако они покрыты гликопротеидами и протеогликанами, что обусловливает их положительную окраску с помощью ШИК-реакции. Ретикулярные волокна окрашиваются в черный цвет при импрегнации серебром. Сравнительная характеристика волокон соединительной ткани приведена в табл. 7.
Фибробласты по мере старения превращаются в фиброциты, которые весьма слабо синтезируют компоненты межклеточного вещества рыхлой волокнистой соединительной ткани.
Фиброцит представляет собой многоотростчатую клетку веретенооб- разной формы с крупным эллипсоидным ядром, мелким ядрышком и небольшим количеством бедной органеллами цитоплазмы. Зернистая эндоплазматическая сеть и сетчатый аппарат развиты слабо. Цитоплазма вакуолизирована, в ней имеются лизосомы, аутофагосомы.
Таблица 7. Характеристика волокон соединительной ткани
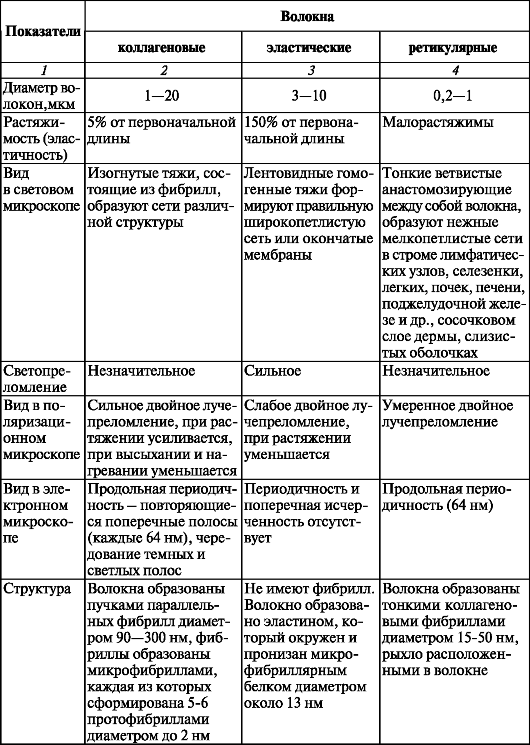
Окончание таблицы 7
В рыхлой волокнистой соединительной ткани присутствуют клетки, разрушающие компоненты межклеточного вещества, - фибробласты. Эти клетки по структуре весьма напоминают фибробласты (форма, развитая зернистая эндоплазматическая сеть и комплекс Гольджи). В то же время они фагоцитируют коллаген и богаты лизосомами.
Ретикулярная клетка, или ретикулоцит, - удлиненная многоотрост- чатая клетка, которая соединяется отростками с другими ретикулоцитами, формируя сеть. Ретикулярные клетки вместе с ретикулярными волокнами образуют строму органов кроветворения и иммунной системы. Крупное округлое или эллипсоидное, относительно бедное хроматином ядро с четко выраженным ядрышком заполняет почти всю клетку. В цитоплазме содержится множество митохондрий, развитый комплекс Гольджи, свободные рибосомы, лизосомы, пучки микрофиламентов и частички гликогена. В зависимости от функциональной активности клетки
имеют более или менее расширенные цистерны зернистой эндоплазматической сети, заполненные осмиофильным веществом. Количество органелл зависит от степени дифференцировки клетки. В определенных условиях (инфекция, внедрение инородных частиц и т. д.) ретикулярные клетки округляются, отделяются от ретикулярных волокон и становятся способными к фагоцитозу. При этом клетки формируют ундулирующие мембраны, псевдоподии, множество микроворсинок и филоподий. Ретикулярные клетки способны связывать на своей поверхности антигены.
Жировые клетки, или адипоциты, накапливают жир. Различают два типа жировой ткани: белую и бурую, которые сформированы соответственно белыми и бурыми жировыми клетками - адипоцитами. Зрелый однокапельный адипоцит белой жировой ткани (диаметром 50-120 мкм) представляет собой шаровидную клетку, почти полностью занятую каплей жира (рис. 19). Ядро жировой клетки оттеснено к периферии, а цитоплазма располагается в виде ободка вокруг жировой капли. При электронной микроскопии в клетке определяется слабо осмиофильная гомогенная капля жира, состоящая из смеси нейтральных жиров, триглицеридов, фосфолипидов и холестерола. Эта капля оттесняет на периферию плоское ядро и узкий слой цитоплазмы, содержащей единичные округлые или овальные митохондрии, элементы зернистой и незернистой эндоплазматической сети, комплекс Гольджи. Цитоплаз- матическая мембрана адипоцитов (липоцитов) лишена микроворсинок, но формирует множество микропиноцитозных пузырьков, образующих розетки. Каждый адипоцит окружен базальной мембраной, которая снаружи укреплена сеточкой, состоящей из ретикулярных микрофибрилл.
В развивающейся жировой клетке органелл больше, чем в зрелой, имеется несколько капель жира, каждая из которых отделена от цитоплазмы тонкой внутренней мембраной, снаружи окруженной системой тонких параллельных меридионально расположенных филаментов. Элементы незернистой эндоплазматической сети в виде коротких трубочек и пузырьков окружают капли жира. Адипоциты поглощают жирные кислоты и синтезируют из них и глицерофосфата триглицериды. Необходимый глицерофосфат образуется жировыми клетками в процессе метаболизма глюкозы. Триглицериды могут синтезироваться также из глюкозы и аминокислот.
По мере потери жира адипоцитами капли уменьшаются или даже исчезают из цитоплазмы. Одновременно клетки формируют длинные отростки. Освобождающиеся жирные кислоты поступают в кровоток, связываются альбуминами, транспортируются и поглощаются другими клетками, которые используют их в качестве источника энергии.
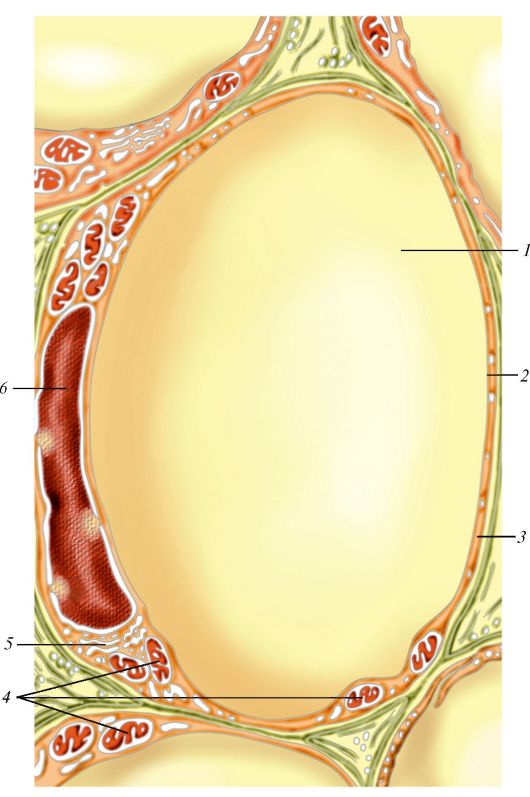
Рис. 19. Строение однокапельного адипоцита: 1 - капля жира; 2 - цитоплазма адипоцита; 3 - базальная мембрана; 4 - митохондрии; 5 - сетчатый аппарат; 6 - ядро фибробласта (по Р. Крстичу, с изменениями)
Жировые включения обычно полностью вымываются из клеток при проводке тканей через спирты, поэтому цитоплазма жировых клеток выглядит ячеистой, пустой, так как вместо жира содержит крупную светлую полость. После фиксации водным раствором формалина и окраски суданом III и гематоксилином видны скопления жировых клеток, располагающихся, как правило, по ходу кровеносных сосудов.
Многокапельный полигональный адипоцит бурой жировой ткани имеет небольшие размеры, содержит шаровидное ядро, расположенное в центре, множество крупных митохондрий, а также малочисленные элементы зернистой и незернистой эндоплазматической сети, большое количество липидных капель, окруженных описанными выше микрофиламентами. Содержимое капель умеренно осмофильно и гомогенно. Цитолемма формирует множество пиноцитозных пузырьков. Многокапельный адипоцит окружен собственной базальной мембраной, укрепленной снаружи сетью, образованной ретикулярными и коллагеновыми микрофибриллами.
Перициты (перикапиллярные клетки, или клетки Руже) располагаются кнаружи от эндотелия, внутри базального слоя капилляра. Перициты - это отростчатые клетки, соприкасающиеся отростками с эндотелиоцитами. Они передают последним нервное возбуждение, что способствует накоплению или потере эндотелиоцитом жидкости. Это приводит к расширению или сужению просвета капилляра. По строению перициты весьма напоминают фибробласты, но цитоплазма перицита богаче филаментами, а на цитоплазматической стороне цитолеммы имеются плотные тельца.
Пигментные клетки (меланоциты), содержащие пигмент, залегают в эпидермисе кожи. Их особенно много в эпидермисе наружных половых органов, в радужке и собственно сосудистой оболочке глазного яблока, в мягкой мозговой оболочке. Пигментные клетки имеют звездчатую форму и содержат множество зерен эндогенного меланина. В световом микроскопе видны клетки, имеющие неправильную форму, цитоплазма которых заполнена черно-коричневыми гранулами пигмента меланина. Ядро выглядит в виде светлой зоны, окруженной гранулами. Эллипсоидное ядро клетки имеет множество инвагинаций. Большинство окруженных мембраной гранул размерами 0,7-0,3 мкм (до 1 мкм) содержат зрелый пигмент, гранулы проникают и в отростки клетки. Промеланиновые гранулы, синтезируемые зернистой эндоплазматической сетью, транспортируются в сетчатый аппарат, от которого отщепляются содержащие спиральные молекулы тирозиназы пузырьки, которые превращаются в меланосомы (предшественники гранул). Тирозиназа под влиянием ди- гидроксифенилаланина образует зрелый электронноплотный меланин,
заключенный в мембрану. Цитоплазматические органеллы развиты хорошо, имеется небольшое количество микрофибрилл. В пигментной соединительной ткани пигментные клетки соединяются своими отрос- тками, образуя сеть. В эпидермисе пигментные клетки залегают между клетками базального слоя и в волосяных луковицах.
На 1 мм2 поверхности кожи приходится 1200-1500 пигментных клеток, у представителей черной и желтой рас количество их значительно больше. Цвет глаз зависит от количества пигментных клеток в радужке. Большое количество тесно расположенных клеток обусловливает черный или темно-карий цвет. Чем меньше пигментных клеток, тем светлее радужка.
Макрофаг - основной структурный элемент системы мононуклеар- ных фагоцитов, включающей группу клеток, объединенных общностью происхождения из моноцитов крови, строения и функции (активный фагоцитоз и пиноцитоз). К макрофагам относятся оседлые макрофагоциты (гистиоциты), звездчатые ретикулоэндотелиоциты (клетки Купфера), остеокласты, макрофаги нервной системы, свободные и фиксированные макрофаги лимфатических узлов и селезенки, очагов воспаления, альвеолярные макрофаги.
Циркулирующий моноцит покидает кровеносное русло, мигрирует в ткани, где превращается в макрофаг. Макрофаги подразделяют на свободные - кочующие (макрофаги соединительной ткани, легких и серозных полостей) и фиксированные - оседлые, которые постоянно находятся в печени, костном мозге и лимфоидных органах. Макрофаги отличаются высокой гетерогенностью. Их морфофункциональные особенности зависят от микроокружения. Так, например, альвеолярные макрофаги - аэробы, а преобладающий тип их метаболизма - окислительное фосфорилизирование. Перитонеальные макрофаги и клетки Купфера - факультативные анаэробы, у которых преобладает гликолиз.
Размеры макрофагов колеблются в пределах 10-23 мкм. Поверхность макрофага покрыта многочисленными микроворсинками, складками цитолеммы, округлыми выростами, ундулирующими мембранами. Имеются псевдоподии, число которых пропорционально функциональной активности макрофага. Макрофаг способен к амебоидному движению. Цитолемма образует множество пиноцитозных углублений и пузырьков. Ядро обычно шаровидное или овоидное, зачастую имеет глубокие инвагинации, бедно хроматином (преимущественно периферический гетерохроматин), однако ядрышко всегда выражено четко. Ядерно-цитоплазматическое отношение всегда сдвинуто в сторону цитоплазмы. Макрофаги человека содержат небольшое количество элементов зернистой эндоплазматической
сети и митохондрий, хорошо развитый сетчатый аппарат, множество рибосом. В макрофагах много различных функциональных форм лизосом: мелкие первичные в окружности комплекса Гольджи, фаголизосомы, остаточные тельца (рис. 20). По мере созревания от костномозгового промоноцита к макрофагу в нем увеличивается количество лизосом, микроворсинок, повышается активность большинства ферментов и снижается активность пероксидазы. В цитолемму макрофагов встроены рецепторы к иммуноглобулинам, комплементу, опсонинам, лимфокинам, Т-лимфоцитам, опухолевым и другим клеткам.
Благодаря активному фагоцитозу широко используется прижизненное (витальное) окрашивание макрофагов с помощью трипанового синего, коллоидного серебра, китайской туши. Прижизненное введение красителя животному позволяет проследить участие элементов соединительной ткани, в первую очередь макрофагов, в очищении внутренней среды организма. Частицы красителя захватываются макрофагами. При микроскопии можно отчетливо видеть макрофаги с четкими контурами, содержащие в цитоплазме темно-синие гранулы фагоцитированного красителя и гиперхромные ядра клеток, окрашенные кармином в красный цвет. При необходимости фагоцитировать крупные частицы нескольких макрофагов сливаются, образуя гигантские клетки инородных тел, содержащие два ядра и более.
Макрофаги развиваются из агранулоцитов крови - моноцитов, которые мигрируют в ткани из кровяного русла. Моноциты в свою очередь образуются в костном мозге из стволовых клеток крови, проходя следую- щие этапы: общая клетка-предшественница миелопоэза →клетка-пред- шественница гранулоцитомонопоэза → монобласт → промоноцит → моноцит костного мозга → циркулирующий моноцит крови → тканевый макрофагоцит.
Плазмоциты, или плазматические клетки (см. т. 2 «Органы иммунной системы»), вырабатывают иммуноглобулины, чем определяется их важ- нейшая роль в защите организма. Плазмоциты происходят из В-лимфоцитов. В рыхлой волокнистой неоформленной соединительной ткани при окраске метиловым зеленым и пиронином по методу Унны-Паппенгей- ма видны небольшие плазматические клетки овальной формы, величиной 15-17 мкм, с базофильной цитоплазмой, окрашенной пиронином в розовый цвет. Интенсивная базофилия цитоплазмы связана с большим содержанием рибосомальной РНК. Во многих клетках видны более светлые участки цитоплазмы около ядра («дворик»), не содержащие рибосом; здесь расположены клеточный центр и комплекс Гольджи. Шаровидное ядро, богатое хроматином, который образует характерную фигуру
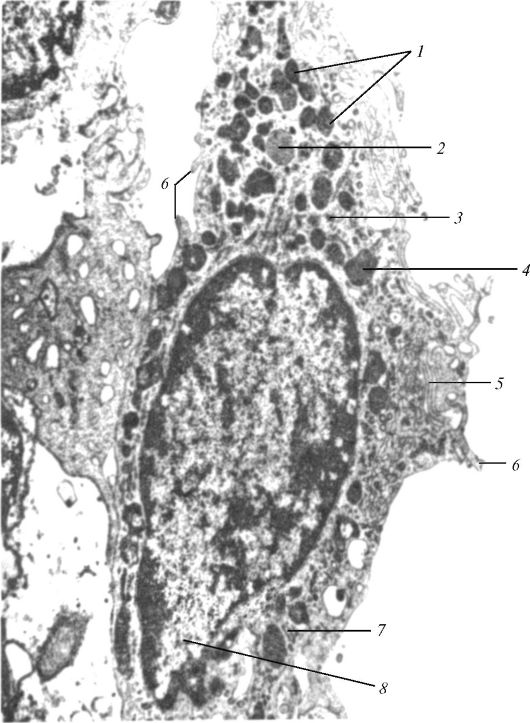
Рис. 20. Строение макрофага: 1 - первичные лизосомы; 2 - вторичная лизосома; 3 - остаточное тельце; 4 - митохондрия; 5 - сетчатый аппарат; 6 - микроворсинки; 7 - зернистая эндоплазматическая сеть; 8 - ядро
колеса со спицами, лежит в центре или эксцентрично, содержит крупные глыбки хроматина, расположенные радиально.
Плазмоциты находятся главным образом вблизи мелких кровеносных сосудов в органах иммунной системы, в собственной пластинке слизистой оболочки пищеварительной и дыхательной систем и т. д. Цитолемма плаз- моцита формирует небольшое количество мелких микроворсинок, пальцевидных и шаровидных выростов, а также микропиноцитозные пузырьки и ямки, часть из них окаймленные. Плазматическая клетка способна к незначительным амебоидным движениям. Цитоплазма богата элементами зернистой цитоплазматической сети с цистернами, расположенными параллельными рядами, заполненными мелкозернистым осмиофильным веществом. Между ними лежат единичные митохондрии и большое количество свободных рибосом. Некоторые цистерны отделены от цитолеммы очень тонкой прослойкой цитоплазмы. Вблизи ядра расположены центросома и комплекс Гольджи, молодая плазматическая клетка имеет округлую форму, ядро с крупноглыбчатым хроматином, цитоплазму, заполненную расширенными цистернами зернистой цитоплазматической сети и содержащую скопления мелких митохондрии и центросому.
Тканевые базофилы, или тучные клетки (лаброциты), располагаются группами в собственно коже и строме различных органов, вблизи мелких кровеносных сосудов и отличаются полиморфизмом и вариабельностью. Тучные клетки имеют размеры 12-15 мкм, округлую или овальную форму, небольшое эллипсоидное ядро и большое количество цитоплазмы, в которой находится множество крупных (до 2 мкм) мембранных гранул. В этих гранулах у человека выявляются цилиндрические закрученные ламеллярные структуры. Гранулы содержат гепарин, гистамин, гиалуронидазу. Гепарин тормозит свертывание крови. Гистамин - производное гистидина, вызывает сокращение гладких миоцитов, повышает проницаемость капилляров, способствуя расхождению эндотелиоцитов в тех зонах, где между их цитолеммами отсутствуют плотные контакты, вызывает аллергические и анафилактические симптомы.
В тучных клетках между гранулами располагаются элементы зернистой эндоплазматической сети, хорошо развитые комплекс Гольджи, рибосомы и единичные митохондрии. На гладкой поверхности неактивных тучных клеток имеется небольшое количество выпячиваний. Поверхность активных клеток покрыта множеством микроворсинок, округлых и пальцевидных выростов и складок, а также углублений, оставшихся после дегрануляции. Функция тучных клеток - секреция биологически активных веществ. Выделение гепарина происходит путем дегрануляции, гистамина - без повреждения
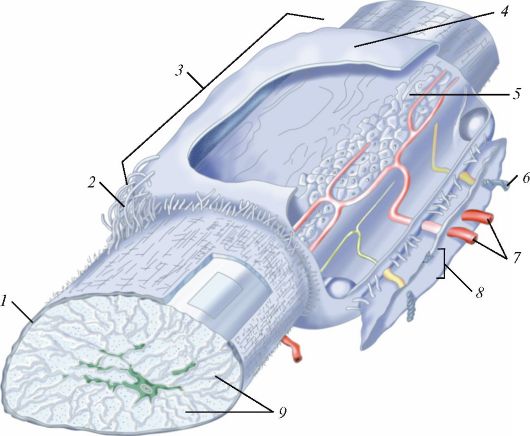
Рис. 21. Строение сухожилия: 1 - перитендиний; 2 - фиброзный слой; 3 - сухожильное влагалище; 4 - париетальная часть влагалища; 5 - сухожильная часть влагалища; 6 - нервное волокно; 7 - кровеносные сосуды; 8 - мезотендрий; 9 - пучки сухожильных волокон I порядка (по Р. Крстичу, с изменениями).
гранул. Тучные клетки не утратили способность к синтезу ДНК и митотическому делению. Близость тучных клеток к капиллярам облегчает проникновение вырабатываемых ими веществ в кровь.
Особенность тучных клеток - базофильная метахроматическая зернистость цитоплазмы. Краситель толуидиновый синий обладает ярко выраженной способностью выявлять метахромазию тучных клеток, т. е. их способность изменять цвет. Тулоидиновый синий окрашивает мета- хроматическую зернистость тучных клеток в красно-фиолетовый цвет.
Плотная волокнистая соединительная ткань (textus conjunctivusfibrosus compactus) образована переплетающимися в разных направлениях волокнами (неоформленная ткань) либо волокнами, имеющими упорядоченную ориентацию (оформленная ткань).
Неоформленная плотная волокнистая соединительная ткань образует капсулы органов и отходящие от них внутрь органов трабекулы, адвентицию, оболочки сосудов, склеру глаза, надкостницу и надхрящницу, суставные капсулы, сетчатый слой кожи (дермы), клапаны сердца, перикард, твердую оболочку мозга. Эта ткань бедна клетками (в основном присутствуют фиброциты), однако богата волокнами. Многочисленные отдельные волокна и пучки коллагеновых волокон проходят в различных направлениях, имеется небольшое количество переплетающихся эластических волокон, придающих ткани некоторую эластичность.
Оформленная плотная волокнистая соединительная ткань образует су- хожилия, связки, фасции, пластины. Между параллельно расположенными коллагеновыми волокнами находятся сухожильные клетки с характерными темными ядрами продолговатой формы. Пучки коллагеновых волокон I порядка объединены в пучки II порядка, которые разделены прослойками рыхлой волокнистой соединительной ткани, получившими название мезотендиний (рис. 21). Снаружи сухожилие покрыто перитенонием. Сухожильные клетки - тендиноциты (разновидность фиброцитов), находясь между коллагеновыми пучками, образуют множество тон- ких отростков, которые охватывают пучки коллагеновых волокон. Овоидное ядро тендиноцитов богато хроматином. Хорошо развитая зернистая эндоплазматическая сеть и множество свободных рибосом свидетельствует об активной белоксинтезирующей функции этих клеток. Тендиноциты синтезируют и секретируют коллаген. Коллагеновые микрофибриллы связаны между собой межфибриллярными мостиками.
Эластическая соединительная ткань участвует в образовании стенок артерий эластического типа, эластического конуса гортани и ее голосовых связок, желтых связок. Главными элементами этой ткани являются тесно прилежащие друг к другу эластические волокна, между которыми залегают малочисленные фиброциты. Тонкофибриллярная сеть, образованная коллагеновыми и ретикулярными микрофибриллами, окутывает эластические волокна.
СОЕДИНИТЕЛЬНЫЕ ТКАНИ СО СПЕЦИАЛЬНЫМИ СВОЙСТВАМИ
Соединительные ткани со специальными свойствами (жировая, ретикулярная, пигментная) расположены лишь в определенных органах и участках тела и имеют особые черты строения и своеобразную функцию.
Жировая ткань (textus adipdsus) выполняет трофическую, депонирующую, формообразующую и терморегулирующую функции. Жировая
ткань подразделяется на белую, образованную однокапельными адипоцитами, и бурую, образованную многокапельными адипоцитами. Группы жировых клеток объединены в дольки, отделенные друг от друга перегородками рыхлой волокнистой неоформленной соединительной ткани, в которой проходят сосуды и нервы. В свою очередь между отдельными адипоцитами проходят тонкие коллагеновые и ретикулярные волокна, вместе с которыми проходят кровеносные капилляры.
У человека преобладает белая жировая ткань. Она окружает некоторые органы, сохраняя их положение в теле человека (например, почки, лимфатические узлы, глазное яблоко и др.), заполняет пространства еще не функционирующих органов (например, молочная железа), замещает красный костный мозг в диафизах длинных трубчатых костей. Большая часть жировой ткани является резервной (подкожная основа, сальники, сальниковые отростки толстой кишки).
Количество бурой жировой ткани у взрослого человека невелико. Она имеется главным образом у новорожденного. Бурая жировая ткань расположена в области шеи, в подмышечной полости, под кожей спины и боковых отделов туловища, в средостении и в брыжейках. Подобно белой, бурая жировая ткань также сформирована дольками, образованными многокапельными адипоцитами. Бурый цвет обусловлен множеством кровеносных капилляров, обилием митохондрий и лизосом в многокапельных адипоцитах. Главная функция бурой жировой ткни - теплоизоляция. Бурая жировая ткань поддерживает температуру тела животных во время зимней спячки и температуру тела новорожденных.
Ретикулярная соединительная ткань (tdxtus conjunctivus reticularis), образующая строму органов кроветворения и иммунной системы, сформирована ретикулярными клетками, соединяющимися отростками и ретикулярными волокнами. При импрегнации серебром в поле зрения микроскопа видна сеть, состоящая из тонких черного цвета расщепленных ретикулярных волокон, которые образуют сетчатый каркас. В петлях этой сети располагаются клетки иммунной системы, главным образом лимфоциты, ретикулярные клетки, макрофаги.
Пигментная соединительная ткань (textus conjunctivus pigmentosus) при- сутствует в радужке и собственно сосудистой оболочке глазного яблока, мягкой мозговой оболочке, коже наружных половых органов, сосков, окружности ануса. Многоотростчатые клетки, соединяясь между собой отростками, формируют трехмерную сеть, в ячейках которой проходят коллагеновые и эластические волокна, кровеносные и лимфатические капилляры и нервные волокна.
СКЕЛЕТНЫЕ ТКАНИ (ХРЯЩ, КОСТЬ)
К скелетной соединительной ткани относятся хрящевая и костная ткани.
Хрящевая ткань (textus cartilagineus) содержит около 70-80% воды, 10-15% органических веществ, 4-7% солей. Около 50-70% сухого вещества хряща составляет коллаген. У взрослого человека масса хряща составляет около 2% массы тела. Это суставные хрящи, межпозвоночные диски, хрящи носа и уха, гортани, трахеи, бронхов и ребер. Хрящи покрывают суставные поверхности костей и обусловливают их высокую устойчивость к износу. Суставные хрящи выполняют роль амортизаторов. Хрящи воздухоносных путей и наружного уха формируют стенки полостей. Хрящи не имеют нервов, лимфатических и кровеносных сосудов, их питание осуществляется путем диффузии из окружающих тканей.
В хрящевой ткани присутствуют клетки двух типов: хондробласты (от греч. chondros - хрящ, blastos - зачаток) и хондроциты. Хондробласты - это молодые, способные к митотическому делению клетки, продуцирующие компоненты матрикса: протеогликаны, коллаген, эластин и др. Хондробласты имеют небольшие размеры, на гистологических срезах они округлые или овальные. Цитолемма хондробластов образует множество микроворсинок. У этих клеток хорошо развиты эндоплазматическая сеть (зернистая и незернистая), комплекс Гольджи, митохондрии, лизосомы, гранулы гликогена. Ядро хондробласта, богатое активным эухроматином, имеет 1-2 ядрышка.
Хондроциты - это зрелые клетки хрящевой ткани, которые вырабаты- вают все компоненты хрящевого матрикса одновременно. Это крупные клетки округлой, овальной или полигональной формы с отростками. Хондроциты, богатые элементами незернистой и зернистой эндоплазматической сети, имеют хорошо выраженный комплекс Гольджи, в котором содержится много гранул, окрашивающихся рутениевым красным (протеогликаны). Цитоплазма базофильная, богата включениями гликогена и липидов. Макромолекулы протеогликанов синтезируются клеткой, а их самосборка в агрегаты и комплексирование с коллагеном происходят в матриксе. При этом на разных стадиях развития хондроциты синтезируют различные протеогликаны. Так, молодые хондроциты син- тезируют протеогликаны, в которых цепи хондроитинсульфата в 2 раза длиннее, чем синтезируемые старыми клетками. Ядро хондроцита, расположенное в центре клетки или эксцентрично, богато хроматином. Цитолемма образует множество микроворсинок, которые контактируют
с матриксом. Однако коллагеновые и эластические волокна непосредственно не соприкасаются с цитолеммой и находятся от нее на расстоянии 1-2 мкм. По мере старения человека уменьшаются размеры протеогликанов хряща и количество удерживаемой ими воды, в результате чего хрящ становится менее упругим.
Различают три типа хондроцитов. Меньше всего хондроцитов I типа. Это отростчатые клетки, у которых ядерно-цитоплазматическое отно- шение сдвинуто в сторону ядра, эндоплазматическая сеть развита слабо, в отличие от выраженного комплекса Гольджи, в котором преобладают вакуоли и крупные везикулы. В клетке имеются митохондрии и множество свободных рибосом. Клетки I типа делятся митотически, их много в молодом хряще. Они принимают участие в восстановительных процессах. Среди хрящевых клеток преобладают хондроциты II типа. Это крупные (15-20 мкм), округлые или овальные клетки с крупным ядром, богатым эу- и гетерохроматином. У хондроцитов II типа хорошо выражены зернистая эндоплазматическая сеть и комплекс Гольджи, мало митохондрий, много мелких отростков. Эти клетки не делятся. Хондроциты III типа имеют небольшое ядро, зернистая эндоплазматическая сеть развита хорошо. Подобно клеткам II типа, они активно секретируют белок (в меньшей степени гликозаминогликаны) и также не делятся. Синтетическая активность хондроцитов стимулируется тироксином, тестостероном, соматотропным гормоном и угнетается глюкокортикоидами и эстрогеном.
Хондроциты располагаются в полостях - лакунах, окруженные межклеточным веществом. Чаще всего это две клетки в одной лакуне. Это изогенные группы. Стенки лакуны состоят из двух слоев; наружный слой образован коллагеновыми волокнами, внутренний - агрегатами протеогликанов, которые входят в контакт с гликокаликсом клетки. Структур- ной и функциональной единицей хрящей является хондрон, образованный клеткой или изогенной группой клеток, околоклеточным матриксом и капсулой лакуны.
Различают три вида хрящевой ткани: гиалиновый, эластический и волокнистый хрящ.
Гиалиновый хрящ (от греч. hyalos - стекло) гладкий, блестящий, голубоватого цвета, в его основном хрящевом веществе располагаются колла- геновые волокна. Хрящевые клетки имеют разнообразную форму и строение в зависимости от степени дифференцировки и места расположения в хряще. По периферии расположены хондробласты, в центре - хондроциты в виде изогенных групп (рис. 22). Из гиалинового хряща построены
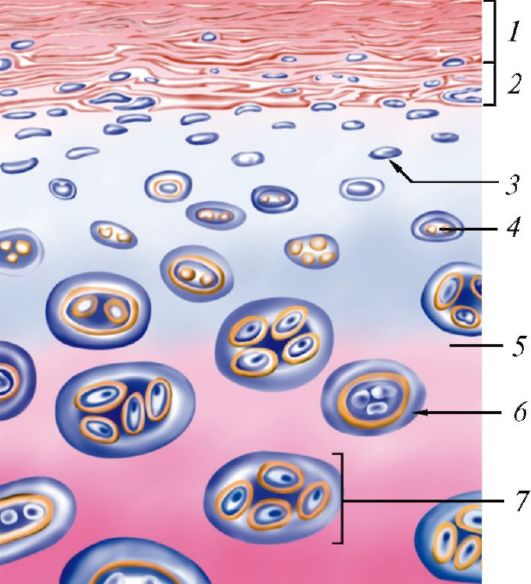
Рис. 22. Строение гиалинового хряща, покрытого надхрящницей:
1 - волокнистый слой надхрящницы;
2 - клеточный слой надхрящницы; 3 - мо- лодые хондроциты; 4 - хондроцит в лакуне; 5 - межклеточное вещество (хрящевой матрикс); 6 - интерстициальный рост; 7 - изогенные группы хондроцитов (зрелые хрящевые клетки) (по А. Хэму и
Д. Кормаку, 1983)
суставные, реберные хрящи, хрящи носа, бронхов и большинство хрящей гортани. С возрастом гиалиновый хрящ кальцифицируется.
Эластический хрящ, имеющий желтоватый цвет, отличается упругостью. В матриксе эластического хряща наряду с коллагеновыми содержится большое количество сложно переплетающихся разветвленных эластических волокон, образованных белком эластином и микрофибриллярными гликопротеинами. Округлые хондроциты расположены в лакунах, стенки которых образованы эластическими во- локнами. Из эластического хряща построены надгортанник, клиновидные и рожковидные хрящи гортани, голосовой отросток черпаловидных хрящей, хрящ ушной раковины, хрящевая часть слуховой трубы.
Волокнистый хрящ содержит в основном веществе много коллагеновых волокон, придающих хрящу повышенную прочность. Клетки, расположенные между коллагеновыми волокнами, имеют вытянутую форму, длинное палочковидное ядро и узкий ободок базофильной цитоплазмы. В матриксе волокнистого хряща большая часть коллагена (около 90%) относится к I типу, меньшая - ко II типу. Коллагеновые волокна расположены упорядоченными параллельными рядами, которые хорошо видны на специально окрашенных микропрепаратах. Хондроцитов значительно меньше, чем в гиалиновом и эластическом хрящах. Хондроциты также расположены в лакунах, реже встречаются изогенные группы. В хондроцитах волокнистого хряща хорошо развита зернистая эндоплазматическая сеть. Из волокнистого хряща построены фиброзные кольца межпозвоночных дисков, суставные диски и мениски. Этим хрящом покрыты суставные поверхности височно-нижнечелюстного и грудино-ключичного
суставов. Этот хрящ имеется в зонах прикрепления связок и сухожилий к костям и хрящам.
Костная ткань (textus osseus) имеет особые механические свойства. Она состоит из костных клеток, замурованных в костное основное вещество, содержащее коллагеновые волокна и пропитанное неорганическими соединениями. Различают клетки двух типов: остеобласты и остеоциты (рис. 23). В костной ткани имеется еще одна категория клеток - остеокласты, которые не являются костными, а имеют моноцитарное происхождение и относятся к системе макрофагов.
Остеобласты - это отростчатые молодые удлиненные костные клетки, залегающие во внутреннем слое надкостницы и эндоста и в местах регенерации (восстановления) костной ткани после ее повреждения. Остеобласты богаты элементами зернистой эндоплазматической сети, рибосомами, хорошо развитым сетчатым аппаратом (Гольджи) и резко базофильной цитоплазмой. Многочисленные отростки этих клеток с множеством актиноподобных микрофиламентов контактируют между
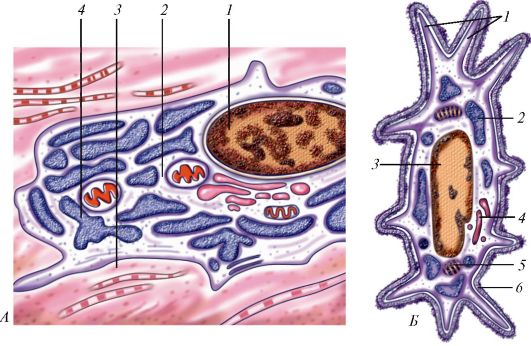
Рис. 23. Костные клетки (схема): А - строение остеобласта: 1 - ядро; 2 - цитоплазма; 3 - остеоид; 4 - развитая зернистая эндоплазматическая сеть; Б - строение остеоцита: 1 - отростки остеоцита; 2 - эндоплазматическая сеть; 3 - ядро; 4 - внутренний сетчатый аппарат; 5 - митохондрия; 6 - остеоидное (необызвествленное) вещество кости по краям лакуны, в которой расположен остеоцит (по В.Г. Елисееву и др.)
собой и с отростками остеоцитов. Округлое или овальное ядро с большим количеством хроматина содержит одно крупное ядрышко, обычно расположенное на периферии. Расширенные полости эндоплазматической сети содержат осмиофильное вещество. Вблизи комплекса Гольджи имеется множество везикул. Остеобласты окружены тонкими коллагеновыми микрофибриллами, образующими микроокружение клеток. Вещества, синтезируемые остеобластами, выделяются через всю их поверхность в различных направлениях, что приводит к образованию стенок лакун, в которых эти клетки залегают. Остеобласты синтезируют компоненты межклеточного вещества и выделяют их из клетки. Активно синтезируется коллаген I типа, который во внеклеточном пространстве собирается в коллагеновые волокна. В промежутках между волокнами располагается амор- фное вещество, состоящее из протеогликанов, сульфатированных гликозаминогликанов, органических кислот. Этот комплекс представляет собой остеоидную ткань, или предкость, которая кальцифицируется. Органический матрикс кости содержит кристаллы гидроксиапатита и аморфный фосфат кальция, которые поступают в костную ткань из крови.
Матрикс кости, пропитанный кристаллами гидроксиапатита, расположен на расстоянии 1-2 мкм от клеточной мембраны костных клеток. Кристаллы гидроксиапатита окутывают коллагеновые фибриллы в виде тонкого слоя, а также расположены внутри фибрилл. Мостики, образованные кристаллами, соединяют фибриллы между собой. В процессе кальцификации важную роль играют мембранные матриксные пузырьки (диаметром 30-100 нм), содержащие липиды, щелочную фосфатазу и пирофосфатазу. Считается, что матриксные пузырьки инициируют процесс кальцификации.
Остеоциты - это зрелые многоотростчатые веретенообразной формы костные клетки с крупным округлым ядром, в котором четко видно ядрышко. Количество органелл невелико - это митохондрии, элементы зернистой эндоплазматической сети и комплекс Гольджи. Остеоциты располагаются в лакунах, однако тела клеток не соприкасаются непосредственно с кальцифицированным матриксом. Между поверхностью клетки и стенками лакуны имеется тонкий слой тканевой (костной) жидкости. Очень длинные (до 50 мкм) отростки остеоцитов, богатые актиноподобными микрофиламентами, проходят в канальцах, причем они отделены от кальцифицированного матрикса пространством шириной около 0,1 мкм, где также находится тканевая (костная) жидкость, за счет которой осуществляется трофика остеоцитов. В то же время расстояние между каждым остеоцитом и ближайшим капилляром не превышает 100-200 мкм.
Щелевые контакты между отростками остеоцитов создают возможность обмена веществами между клетками.
Остеокласты - это крупные многоядерные (5-100 ядер) клетки моноцитарного происхождения размерами до 190 мкм. Эти клетки разрушают кость и хрящ, осуществляют резорбцию костной ткани в процессе ее физиологической и репаративной регенерации. Благодаря разрушению в кости образуются лакуны, в которых залегают остеокласты. Ядра остеокластов богаты хроматином и имеют хорошо видимые ядрышки. В цитоплазме содержится множество митохондрий, элементов зернистой эндоплазматической сети и комплекса Гольджи, свободных рибосом, различных функциональных форм лизосом. Вокруг ядра остеокласта нахо- дится базальная область цитоплазмы, где расположено большинство органелл. Остеокласты имеют ворсинкообразные цитоплазматические отростки, которых особенно много на поверхности, прилежащей к разрушаемой кости. Это гофрированная, или щеточная, каемка остеокласта, увеличивающая площадь его соприкосновения с костью. Отростки остеокластов, в свою очередь, покрыты микроворсинками, между которыми находятся кристаллы гидроксиапатита. Эти кристаллы обнаруживаются в фаголизосомах клетки, где они подвергаются фагоцитозу.
Деятельность остеокластов зависит от действия паратгормона, увеличение синтеза и секреции которого приводит к активации функции остеокластов и разрушению кости.
Типы костной ткани. Различают два типа костной ткани - грубоволок- нистую (ретикулофиброзную) и пластинчатую. Пластинчатая костная ткань образована костными пластинками толщиной от 4 до 20 мкм, которые состоят из остеоцитов и тонковолокнистого основного костного вещества. Волокна, участвующие в образовании пластинки, лежат параллельно друг другу и ориентированы в определенном направлении. Волокна соседних пластинок разнонаправлены и перекрещиваются почти под прямым углом, что обеспечивает большую прочность кости. Грубоволокнистая костная ткань у взрослого человека располагается в зонах прикрепления сухожилий к костям, в швах черепа после их зарастания. Грубоволокнистая костная ткань содержит наряду с остеоцитами толстые неупорядоченные пучки коллагеновых волокон, между которыми находится аморфное вещество.
КРОВЬ
Одной из разновидностей соединительной ткани является кровь с ее жидким основным веществом.
Кровь (sanguis, haima) состоит
из клеток, взвешенных в жидком межклеточном веществе (плазме) сложного
состава. На долю плазмы приходится около 54% объема крови, на долю
форменных элементов (клеток) - около 44%. Кровь выполняет трофическую,
транспортную, защитную функции. Кроме того, кровь участвует в
сохранении постоянного состава и свойств внутренней среды организма -
гомеостаза (от греч. homdoios - одинаковый, stdasis -
состояние, неподвижность). Общее количество крови у взрослого человека
4-6 л, что составляет 6-8% массы его тела (у мужчин в среднем около
Плазма - это жидкая часть крови, в которой содержится около 91% воды, 6,5-8,0% белков, 2% низкомолекулярных соединений. рН плазмы крови колеблется в пределах от 7,37 до 7,43, а удельная плотность равна 1,025-1,029. Плазма богата как электролитами, так и неэлектролитами. Среди катионов преобладают натрий (143 мэкв/л), калий и кальций (по 5 мэкв/л каждый), среди анионов - хлор (103 мэкв/л), бикарбонаты (27 мэкв/л), фосфаты (2 мэкв/л), органические кислоты (6 мэкв/л). В плазме крови содержатся глюкоза (5 мэкв/л) и мочевина (7 мэкв/л). Белки крови (6,5-8 г/л) (альбумины и глобулины) выполняют трофическую, транс- портную, защитную, буферную функции, они также участвуют в процессе свертывания крови и создании коллоидно-осмотического давления.
Часть объема крови, занимаемая его форменными элементами (точнее, эритроцитами), называется гематокритом. У мужчин он равен 44-46 об% общей массы крови, у женщин - 41-43 об%. В крови содержатся безъя- дерные клетки - эритроциты (4,0-5,0)*1012/л крови, лейкоциты (4,0- 6,0)*109/л крови, среди которых выделяют зернистые, или гранулоциты (нейтрофильные, ацидофильные и базофильные), а также незернистые, или агранулоциты (моноциты) (рис. 24). В крови имеются кровяные пластинки (тромбоциты), число которых составляет (180,0-320,0)*109/л, а также лимфоциты - основные морфофункциональные элементы иммунной системы (см. «Органы кроветворения и иммунной системы»).
Эритроциты (от греч. erythros - красный), или красные кровяные тельца, имеют форму двояковогнутых дисков диаметром от 7 до 10 мкм. Эритроциты содержат гемоглобин, осуществляющий перенос кислорода и двуокиси углерода. Содержание эритроцитов у мужчин составляет (4-5)-1012/л, у женщин (3,9-4,7)-1012/л. Общее количество эритроцитов
Рис. 24. Клетки крови: I - базофильный гранулоцит; II - ацидофильный гранулоцит; III - сегментоядерный нейтрофильный гранулоцит; IV - эритроцит; V - моноцит; VI - тромбоциты; VII - лимфоцит
у мужчин достигает 27?1012 клеток, у женщин 18?1012 клеток, а общая площадь поверхности всех эритроцитов равна приблизительно
Примерно 1-2% эритроцитов имеют голубоватый цвет, их называют полихроматофильными эритроцитами (ретикулоцитами). Цитоплазма полихромная, при окраске крезиловым синим в них видна синяя сеточка неправильной формы, обусловленная наличием рРНК, которая сохраняется в течение 24 ч. Гемоглобина в этих клетках меньше, чем в зрелых эритроцитах. В них имеется небольшое количество митохондрий, элементов комплекса Гольджи и рибосом, а также гранулы гемосидерина (сидеросомы). Ретикулоциты поглощают ферритин посредством пиноцитоза. Через 24-36 ч ретикулоциты превращаются в зрелые эритроциты. Увеличение количества ретикулоцитов свидетельствует об усиленном образовании эритроцитов в ответ на их более интенсивное разрушение.
Лейкоциты (от греч. ldikos - белый) представляют собой ядросодержащие шаровидные клетки, обладающие амебоидной подвижностью.
В лейкоцитах имеются митохондрии, лизосомы, комплекс Гольджи, эндоплазматическая сеть, рибосомы и другие органеллы. В отличие от эритроцитов, которые выполняют присущие им функции в просвете кровеносных сосудов, большая часть лейкоцитов осуществляет свои функции в тканях, куда они мигрируют посредством диапедеза (от греч. dia - сквозь, peddesis - прыжок). Для этого они входят в контакт с эндотелием. Затем образуют псевдоподии (от греч. psdudes - ложный, pus - нога) - лишенные органелл выросты цитоплазмы, покрытые цитолеммой, которые внедряются в межклеточные щели между эндотелиоцитами и проникают в соединительную ткань. После этого содержимое клетки как бы перетекает в псевдоподию. Это происходит благодаря растворимым сократительным белкам (актину и миозину), которые полимеризуются и взаимодействуют между собой при участии АТР, в результате чего возникает сила, необходимая для движения. Лишь 20% лейкоцитов циркулирует в крови, около 50% находится в тканях, а 30% - в костном мозге. Продолжительность жизни лейкоцитов колеблется в широких пределах - от нескольких дней до нескольких лет.
Лейкоциты, содержащие различные по форме и окраске гранулы, подразделяются на нейтрофильные, эозинофильные и базофильные, полиморфно-ядерные гранулоциты, которые составляют от 93 до 96% всех гранулоцитов. Общее количество гранулоцитов в крови взрослого человека составляет 3?1012. Время циркуляции гранулоцитов в крови не пре- вышает 8-12 ч, затем они мигрируют в соединительную ткань.
Зрелый нейтрофильный гранулоцит представляет собой сферическую клетку диаметром 10-12 мкм. Дольчатое (сегментированное) ядро клетки, в котором не видно ядрышек, содержит гетерохроматин и при световой микроскопии выглядит окрашенным в темно-синий или сине-фиолетовый цвет. Гетерохроматин расположен по периферии ядра, вблизи нуклеолеммы, а эухроматин - в его центре, ядрышек нет. Цитоплазматическая мембрана образует небольшое количество коротких микроворсинок.
Слабо оксифильная цитоплазма клетки богата нейтрофильными и азурофильными мембранными гранулами. Мелкие вторичные специ- фические нейтрофильные гранулы (диаметром 0,1-0,3 мкм) составляют 80-90% всех гранул. При световой микроскопии они имеют лиловый цвет. При электронно-микроскопическом исследовании гранулы округлые или удлиненные, видны окружающая их элементарная мембрана и содержимое умеренной электронной плотности, богатое щелочной фосфатазой и бактерицидными веществами. Гранулы содержат катионные белки, аминопептидазу, коллагеназу, лизоцим. Более крупные
(диаметром до 0,4 мкм) красновато-фиолетовые азурофильные гранулы (первичные лизосомы) составляют около 10-20% всех гранул. В их электронноплотном материале содержатся кислая фосфатаза и другие лизосо- мальные ферменты, лизоцим, миелопероксидаза. Последняя является маркером этих гранул. Гранулы обоих типов участвуют в фагоцитозе и инактивации фагоцитированного материала. Цитоплазма нейтрофильных гранулоцитов богата частичками гликогена, липидами и бедна органеллами. Внутренний сетчатый аппарат (Гольджи) развит слабо, центриоли мелкие и обнаруживаются с трудом, мелких округлых митохондрий немного.
Около 3-5% лейкоцитов составляют палочкоядерные нейтрофильные гранулоциты с ядром, напоминающим изогнутую палочку или S-образ- ным. До 0,5% составляют юные (метамиелоциты), имеющие бобовидное ядро. Палочкоядерные и юные нейтрофилы имеют описанные нейтро- фильные гранулы и лишь небольшое количество азурофильных гранул.
Осуществляя фагоцитоз продуктов распада и микроорганизмов, нейтрофильные гранулоциты погибают, а освобождающиеся при этом лизосомальные ферменты разрушают окружающие ткани, способствуя формированию гнойника.
Эозинофильные (ацидофильные) гранулоциты составляют от 0,5 до 5% циркулирующих лейкоцитов. В 1 мм3 крови их число колеблется от 120 до 350. Между числом эозинофильных гранулоцитов и уровнем глюкокортикоидов в крови существует обратно пропорциональная связь. В полночь количество клеток достигает максимума, рано утром - минимума. Эозинофилы циркулируют в крови не более 8 дней, после чего покидают кровеносное русло через мелкие венулы и проникают в рыхлую соединительную ткань. Их особенно много в собственной пластинке слизистой оболочки кишечника и дыхательных путей. Диаметр эозинофильных гранулоцитов составляет 10-15 мкм. Двудольчатое ядро, напоминающее по форме гантелю, богато гетерохроматином и содержит 1-2 ядрышка. Цитолемма формирует небольшое количество мелких микроворсинок. В цитоплазме имеется множество крупных мембранных ацидофильных (красных или оранжевых) светопреломляющих, несколько удлиненных гранул диаметром около 1 мкм. Гранулы представляют собой лизосомы, содержащие пероксидазу, кислую фосфатазу и другие лизосомальные ферменты. Кроме того, имеются мелкие (диаметром 0,1-0,3 мкм) округлые гомогенные гранулы, которые содержат ферменты кислую фосфатазу и арилсульфатазу. Хорошо развитый внутренний сетчатый аппарат (Гольджи) располагается в углублении между долями ядра. В цитолемме имеются митохондрии, элементы зернистой и незернистой эндоплазматической сети, частички гликогена.
Эозинофильные гранулоциты участвуют в фагоцитозе, но менее активно, чем нейтрофильные. Эозинофильные гранулоциты фагоцитируют комплекс антиген-антитело, участвуют в разрушении гистамина, уменьшают альтернативные процессы при местных аллергических реакциях. Количество эозинофильных гранулоцитов в циркулирующей крови (эозинофилия) увеличивается при паразитарных заболеваниях, аллергических и аутоиммунных процессах.
Количество базофильных гранулоцитов в циркулирующей крови невелико - около 0,5% всех лейкоцитов (40-50 клеток в 1 мм3 крови), а время их циркуляции в крови не превышает 12-15 ч. Диаметр клетки 10-12 мкм, при световой микроскопии в клетке видно множество темно-синих округлых или овальных метахроматичных гранул размерами до 2 мкм. Количество гранул столь велико, что они маскируют крупное двудольчатое или S-образное ядро, которое занимает около половины объема клетки. Базофильные гранулы окружены мембранами и заполнены частичками размерами около 15 нм. Гранулы содержат гистамин и гепарин. В цитоплазме находятся рибосомы, небольшое количество митохондрий и элементов зернистой эндоплазматической сети, хорошо развитый комплекс Гольджи, множество частиц гликогена размерами 25-30 нм, небольшое количество азурофильных гранул (лизосомы).
Цитоплазматическая мембрана базофильных гранулоцитов формирует небольшое количество коротких микроворсинок. В цитолемму встроены рецепторы IgE, которые связывают этот иммуноглобулин. В результате образования иммунного комплекса на поверхности клетки происходит дегрануляция и освобождается гистамин, вызывающий аллергическую реакцию. Гепарин, выделяющийся базофильными гранулоцитами, активирует липолиз в сыворотке крови. В клетках содержится и небольшое количество азурофильных гранул (лизосомы). Базофильные гранулоциты осуществляют синтез (и метаболизм) гепарина и гистамина, участвуют в аллергических и воспалительных реакциях, обладают фагоцитарной активностью.
В крови постоянно присутствуют также клетки лимфоидного ряда (лимфоциты), которые являются структурными элементами иммунной системы. В научной и учебной литературе лимфоциты все еще причисляют к незернистым лейкоцитам (агранулоцитам) крови, что явно не- правильно. Лимфоциты (см. т. 2 «Органы кроветворения и иммунной системы») содержатся в большом количестве в крови (25-40% всех лейкоцитов, 1000-4000 клеток в 1 мм3), преобладают в лимфе и ответственны за иммунитет. В организме взрослого человека число лимфоцитов
достигает 6?1012 («Человек», 1977). Большая часть лимфоцитов постоянно циркулирует в организме, что способствует обеспечению иммунной защиты организма.
Моноциты составляют от 3 до 11% циркулирующих клеток крови (200-600 клеток в 1 мм3). Они пребывают в кровеносной системе 2-3 сут, после чего мигрируют в ткани, где превращаются в макрофаги. В течение суток у человека присутствует (0,6-1,0)?109 моноцитов, причем в циркулирующей крови их примерно в 20 раз меньше, чем в тканях. Моноцит - довольно крупная овальная клетка, ее диаметр достигает 15 мкм. Цитолемма формирует небольшое число коротких микроворсинок. Крупное почкообразное богатое хроматином ядро содержит 1-2 ядрышка, окружено оно большим количеством голубовато-серой базофильной цитоплазмы, в которой присутствуют мелкие азурофильные гранулы (первичные лизосомы). В клетке имеется умеренное количество свободных рибосом, элементов гранулярной эндоплазматической сети, а также митохондрии, множество фагоцитарных вакуолей и пиноцитозных пузырьков. В ядре расположен развитый комплекс Гольджи, вблизи которого видны центриоли.
Тромбоциты, или кровяные пластинки, - это уплощенные овальные двояковыпуклые безъядерные фрагменты крупных клеток мегакариоцитов диаметром 2-4 мкм и толщиной 0,5-0,75 мкм. Их число достигает 250 000-350 000 в 1 мм3 крови. Время их циркуляции в крови не превышает 7-10 дней, после чего они попадают в селезенку, где разрушаются. Под цитолеммой тромбоцитов, которая снаружи покрыта слоем гликокаликса толщиной около 50 нм, располагается бледно-голубой мелкозернистый матрикс (гиаломер), не имеющий органелл. По периферии матрикса образуется кольцо микротрубочек, оно и обусловливает форму пластинки. В центре пластинки находится грануломер, в котором располагается небольшое количество митохондрий, элементов эндоплазматической сети, лизосом, частичек гликогена, секреторных гранул (α-гранул) диаметром 0,2-0,3 мкм, содержащих гидролитические ферменты (в том числе кислую фосфатазу, глюкуронидазу, катепсин). В пластинках имеется система трубочек, связанных с цитолеммой, внутри которых находятся мукополисахариды, аналогичные гликокаликсу. Кроме того, обнаруживаются плотные трубочки, содержащие материал умеренной электронной плотности.
Тромбоциты участвуют в свертывании крови, остановке кровотечений и защите организма благодаря способности фагоцитировать вирусы, иммунные комплексы и неорганические частички (Вейсс У., 1986),
в депонировании серотонина и гистамина. При повреждении эндотелия и соприкосновении с обнаженными коллагеновыми волокнами тромбоциты прилипают к ним и агрегируют, в результате чего увеличивается проницаемость мембран тромбоцитов. Из них высвобождаются серотонин, катехоламины, АТР, АДФ и фосфолипид - тромбоцитарный фактор (ТФ-3). Серотонин и катехоламины вызывают сужение сосудов, а АДФ усиливает адгезию пластинок. Под действием ТФ-3 в присутствии ионов кальция белок плазмы протромбин, образующийся в печени, превращается в тромбин, который вызывает переход плазменного белка фибриногена, также образующегося в печени, в фибрин. Последний и формирует основную часть тромба (Вейсс У., 1986).
Свертываемость крови. После повреждения кровеносного сосуда вытекающая из него кровь свертывается через 3-4 мин, а через 5-6 мин превращается в плотный сгусток - тромб. Это важное свойство крови свертываться предохраняет организм от кровопотери.
Процесс свертывания крови протекает с участием веществ, освобождающихся при разрушении тромбоцитов и повреждении тканей. Из поврежденных тромбоцитов и клеток тканей выделяется белок, который, взаимодействуя с белками плазмы крови, преобразуется в активный тромбопластин. Для образования тромбопластина необходимо, в частности, присутствие в крови антигемолитического фактора. Если в крови антигемолитический фактор отсутствует или его мало, то свертываемость крови низкая, кровь не свертывается. Это состояние получило название гемофилии. Далее при участии образовавшегося тромбопластина белок плазмы крови протромбин превращается в активный фермент тромбин. При взаимодействии образовавшегося тромбина растворенный в плазме белок фибриноген превращается в нерастворимый фибрин. Для предупреждения свертывания крови в кровеносных сосудах в организме имеется противосвертывающая система. В печени и легких образуется вещество гепарин, препятствующий свертыванию крови путем превращения тромбина в неактивное состояние.
Группы крови. Переливание крови. При кровопотерях в результате травмы и при некоторых других состояниях практикуется переливание человеку (называемому реципиентом) крови другого человека (донорской крови). Важно, чтобы донорская кровь была совместима с кровью реципиента. Дело в том, что при смешивании крови от разных лиц эритроциты, оказавшиеся в плазме крови другого человека, могут склеиваться (агглютинироваться), а затем разрушаться (гемолизироваться). Гемолизом называют процесс разрушения цитолеммы эритроцитов и выхода из них
Таблица 8. Классификация групп крови человека

гемоглобина в окружающую их плазму крови. Гемолиз эритроцитов может произойти при смешивании несовместимых групп крови или при введении в кровь гипотонического раствора, при действии некоторых химических ядовитых веществ, а также в результате действия яда некоторых змей. Как известно, в крови каждого человека имеются особые белки, которые способны взаимодействовать с такими же белками крови другого человека. В эритроцитах такие белковые вещества получили название агглютиногенов, обозначаемых заглавными буквами А и В. В плазме крови также имеются белковые вещества, получившие название агглютининов α (альфа) и β (бета). Свертывание крови (агглютинация и гемолиз эритроцитов) происходит в том случае, если встречаются одноименные агглютиноген и агглютинин (α и A; В и β). С учетом наличия тех или иных агглютининов и агглютиногенов кровь людей делят на 4 группы (табл. 8.).
В крови первой (I) группы, в ее плазме, содержатся оба агглютинина α и β, а в эритроцитах этой группы агглютиногенов нет вообще. В плазме крови второй (II) группы имеется агглютинин β, а в эритроцитах присутствует агглютиноген А. В крови третьей (III) группы, в ее плазме, имеется агглютинин α, а в эритроцитах содержится агглютиноген В.
Плазма крови четвертой (IV) группы агглютининов не имеет, а эритроциты содержат оба агглютиногена - А и В.
Кровь всех 4 групп одинаково полноценная и различается только содержанием агглютиногенов и агглютининов. Группа крови человека постоянна. Она не изменяется в течение жизни и передается по наследству. При переливании крови нужно обязательно учитывать совместимость групп крови. Важно, чтобы в результате переливания крови эритроциты донора не склеивались в крови реципиента.
С учетом наличия в крови агглютининов и агглютиногенов людей с кровью I группы называют универсальными донорами, с IV - универсальными реципиентами: им можно переливать кровь любой другой группы, поскольку в плазме их крови нет агглютининов.
Кроме агтлютиногенов А и В эритроциты крови некоторых людей могут содержать агглютиноген, получивший название резус-фактора (Rh). Этот фактор впервые был обнаружен в крови обезьян макак-резусов. Резус-фактор обнаруживается в крови примерно у 85% людей. Кровь таких людей называют резус-положительной (Rh+). Кровь, в которой резус-фактора нет, называют резус-отрицательной (Rh-). Феномен резус-фактора заключается в том, что в крови людей с резус-отрицательным фактором отсутствуют вещества, получившие название антирезусагглютининов. Если человеку с резус-отрицательной кровью повторно перелить резус-положительную кровь, то под влиянием резус-агглютиногена донора в крови реципиента образуются антирезус-агглютинины и гемолизирующие вещества. Это может вызвать агглютинацию и гемолиз эритроцитов.
МЫШЕЧНАЯ ТКАНЬ
Мышечная ткань (textus muscularis) обладает способностью сокращаться, укорачиваться, она осуществляет функции движения. Существуют три разновидности мышечной ткани: исчерченная (поперечнополосатая, скелетная), неисчерченная (гладкая) и сердечная. Наряду с этими разновидностями в организме человека выделяют мышечную ткань эпидер- мального происхождения (миоэпителиальные клетки) и нейтрального происхождения (миоциты мышцы, расширяющей и суживающей зрачок).
Исчерченная (поперечнополосатая, скелетная) мышечная ткань (textus muscularis stridtus, s. skeletdlis) образована цилиндрическими мышечными волокнами длиной от 1 до
Рис. 25. Исчерченная (поперечнополосатая, скелетная) мышечная ткань: 1 - мышечное волокно; 2 - сарколемма; 3 - миофибриллы; 4 - ядра
В зависимости от толщины волокон и содержания в них миофибрилл и саркоплазмы различают красные и белые поперечнополосатые мышечные волокна. Красные волокна богаты саркоплазмой, миоглобином и митохондриями. Однако они самые тонкие, миофибрилл в них мало, они расположены группами. В красных волокнах окислительные процессы более интенсивны, чем в белых, выше активность сукцинатдегидрогеназы и больше гликогена. Белые волокна толстые, содержат меньше саркоплазмы, миоглобина и митохондрий, но миофибрилл в них больше и располагаются они равномерно. Структура и функция волокон неразрывно связаны. Так, белые волокна сокращаются быстрее, но быстрее устают. Красные способны сокращаться длительнее, долго оставаться в сокращенном (рабочем) состоянии. У человека мышцы содержат оба типа волокон. В зависимости от функции мышцы в ней преобладает тот или иной тип волокон.
Мышечные волокна имеют поперечную исчерченность: темные анизотропные диски (полоски А) чередуются со светлыми изотропными дисками (полоски I). Диск А разделен светлой зоной (полоска Н), в центре которой проходит мезофрагма (линия М). Диск I разделен темной линией Z (телофрагма). Мышечные волокна содержат сократительные элементы - миофибриллы, среди которых различают толстые (миозиновые) диаметром 10-15 нм и длиной 1,5 мкм, занимающие диск А, и тонкие (актиновые) диаметром 5-8 нм и длиной 1 мкм, лежащие в диске I и прикрепляющиеся к телофрагмам. Участок миофибриллы, расположенный между двумя телофрагмами, представляет собой саркомер - сократительную единицу длиной около 2,5 мкм (рис. 26). Благодаря тому
Рис. 26. Схема строения двух миофибрилл мышечного волокна: 1 - саркомер; 2 - полоска А (диск А); 3 - полоска H; 4 - линия М (мезофрагма) в середине диска А; 5 - полоска I (диск I); 6 - линия (телофрагма) в середине диска I; 7 - митохондрия; 8 - конечная цистерна; 9 - саркоплазматический ретикулум; 10 - поперечные трубочки (по В.Г. Елисееву и др.)
что границы саркомеров всех миофибрилл одного волокна совпадают, возникает регулярная поперечная исчерченность, которая хорошо видна на продольных срезах мышечного волокна. На поперечных срезах мышечного волокна хорошо видны миофибриллы (myofibrilla) в виде темных округлых точек (пятен) на фоне светлой цитоплазмы.
На электронограмме хорошо видны более электронноплотные анизотропные и светлые изотропные диски, в них продольно идущие миофиламенты, осмиофильная линия Z и светлая зона (полоска Н), разделенная мезофрагмой, многочисленные митохондрии, элементы незернистой эндоплазматической сети. В расслабленной миофибрилле концы актиновых филаментов входят между миозиновыми, в сокращенной зоне перекрытия актиновых и миозиновых филаментов увеличиваются вплоть до полного исчезновения изотропного диска. Каждая миофибрилла окружена незернистой эндоплазматической сетью, состоящей из сетчатого и трубчатого элементов. Первые окружают центральную часть саркомера в виде ажурной сеточки, вторые охватывают большую часть саркомера в виде параллельных трубочек и расположены по обеим сторонам от сетчатых. Трубчатые элементы эндоплазматической сети переходят по обеим сторонам диска А в терминальные цистерны. На границе между дисками А и I сарколемма впячивается, образуя Т-трубочки (поперечные трубочки), которые разветвляются внутри волокна и анастомозируют только в горизонтальном направлении.
На поверхности сарколеммы видны отверстия Т-трубочек. Две терминальные цистерны и поперечная трубочка контактируют между собой, образуя триады. Сети, окружающие саркомеры, сообщаются между собой.
Мышечное сокращение - это результат скольжения тонких (актиновых) филаментов относительно толстых (миозиновых), в результате чего длина филаментов изменяется.
В состав мышечного волокна, помимо миосимпласта, входят сателлитомиоциты (satellitomyocytus). Это уплощенные клетки, которые лежат на поверхности волокна под базальной мембраной. Крупное ядро этих клеток богаче хроматином, чем ядра миосимпластов. В отличие от последних, в клетке сателлитомиоцита имеется центросома, органелл немного. Сателлитомиоциты способны к синтезу ДНК и митотическому делению. Благодаря этому они являются стволовыми клетками поперечнополосатой мышечной ткани, которые участвуют в гистогенезе скелетной мускулатуры и ее регенерации.
Неисчерченная (гладкая) мышечная ткань (textus musculdris nonstriatus) состоит из гладкомышечных клеток - миоцитов, которые располагаются
в стенках кровеносных, лимфатических сосудов и полых внутренних органов, в сосудистой оболочке глаза, в собственно коже. Гладкие миоциты - это удлиненные веретенообразные клетки длиной от 50 до 200 мкм, толщиной от 5 до 15 мкм, не имеющие поперечной исчерченности (рис. 27). Миоциты располагаются группами так, что их заостренные концы внедряются между двумя соседними клетками. Каждый миоцит окружен базальной мембраной, коллагеновыми и ретикулярными микрофибриллами, среди которых проходят эластические волокна. В зонах межклеточных контактов - нексусов базальная мембрана отсутствует. Удлиненное палочковидное ядро с четко видимым ядрышком достигает 10-25 мкм в длину, при сокращении клетки оно принимает форму што- пора. Клетка содержит продольно ориентированные миофиламенты. Лишь вблизи обоих полюсов ядра расположена лишенная миофиламентов цитоплазма, в которой залегают органеллы. Изнутри к цитолемме прилежат веретенообразные клеточные тельца (тельца прикрепления). Они располагаются и в цитоплазме миоцита. Прикрепительные тельца
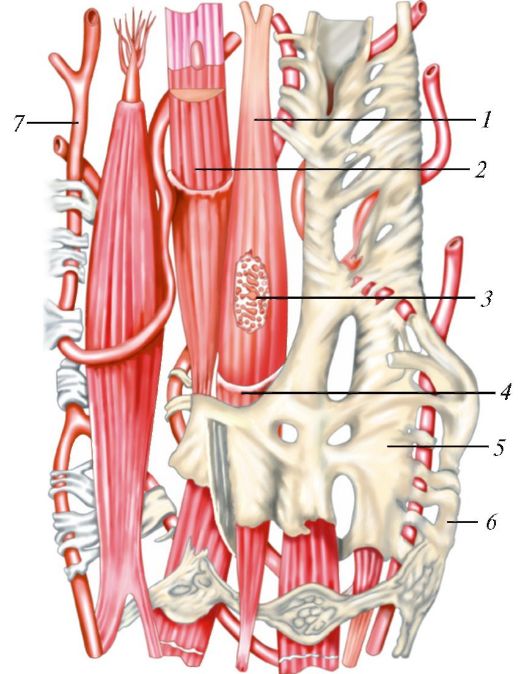
Рис. 27. Строение неисчерченной (гладкой) мышечной ткани: 1 - миоцит; 2 - миофибриллы в саркоплазме; 3 - ядро миоцита; 4 - сарколемма; 5 - эндомизий; 6 - нерв; 7 - кровеносный капилляр (по И.В. Алмазову и Л.С. Сутулову)
(пластинки) являются эквивалентами Z-пластинок поперечнополосатых мышечных волокон, они образованы белком α-актинином. Пластинки представляют собой эллипсоидные тельца длиной до 3 мкм, толщиной 0,2-0,5 мкм, удаленные друг от друга на расстояние 1-3 мкм. Там, где находятся плотные прикрепительные тельца, микропиноцитозные пузырьки отсутствуют.
В цитоплазме гладких миоцитов находятся миофиламенты трех типов: тонкие актиновые диаметром 3-8 нм, которые прикрепляются к плотным тельцам; промежуточные миофиламенты толщиной около 10 нм, образующие пучки, которые соединяют между собой соседние плотные тельца; толстые короткие миозиновые филаменты диаметром около 15-17 нм.
Группа миоцитов, окруженных соединительной тканью, иннервируются обычно одним нервным волокном. Нервный импульс передается с одной мышечной клетки на другую по межклеточным контактам. Воз- буждение передается от одной клетки к другой через нексусы со скоростью 8-10 см/с. Однако в некоторых гладких мышцах (например, сфинктер зрачка) иннервируется каждый миоцит.
В расслабленном миоците между актиновыми филаментами расположены единичные короткие миозиновые. При сокращении актиновые
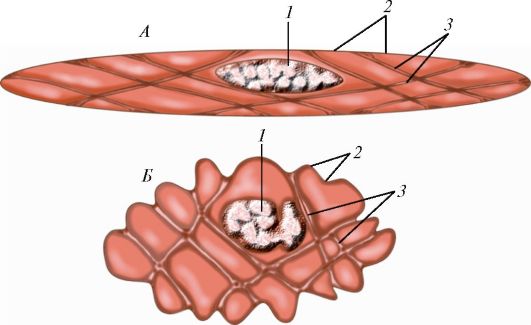
Рис. 28. Гладкая мышечная клетка (миоцит) в расслабленном (А) и сокращенном (Б) состояниях: 1 - ядро; 2 - плотные поля (прикрепительные тельца), прикрепленные к цитолемме; 3 - промежуточные филаменты (по А. Хэму и Д. Кормаку)
филаменты скользят по отношению друг к другу под влиянием миозина, подтягивая прикрепительные тельца, в результате чего цитолемма деформируется, плотные тельца сближаются, а участки, расположенные между ними, вздуваются (рис. 28). Движения одних плотных прикрепительных телец передаются другим промежуточными филаментами, что вызывает синхронное сокращение миоцита.
Гладкие мышцы совершают длительные тонические сокращения (например, сфинктеры полых органов, гладкие мышцы кровеносных сосудов) и относительно медленные движения, которые зачастую ритмичны. Глад- кие мышцы отличаются высокой пластичностью - после растяжения они долго сохраняют длину, которую получили в связи с растяжением.
Сердечная исчерченная мышечная ткань (textus muscularis cardiacus) которая по строению и функции отличается от скелетных мышц, состоит из сердечных миоцитов (кардиомиоцитов). По микроскопическому строению сердечная мышечная ткань похожа на скелетную (поперечнополосатая исчерченность). Однако сокращения сердечной мышцы
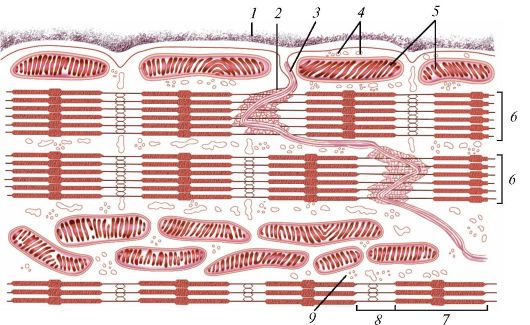
Рис. 29. Схема строения кардиомиоцита: 1 - базальная мембрана; 2 - окончание миопротофибрилл на цитолемме кардиомиоцита; 3 - вставочный диск между кардиомиоцитами; 4 - саркоплазматическая сеть; 5 - саркосомы (митохондрии); 6 - миопротофибриллы; 7 - диск А (анизотропный диск); 8 - диск I (изотропный диск); 9 - саркоплазма
(по В.Г. Елисееву и др.)
не подконтрольны сознанию человека, она иннервируется вегетативной нервной системой, подобно неисчерченной мышечной ткани.
Кардиомиоциты (myocytus cardiacus) - это клетки неправильной цилиндрической формы, длиной 100-150 мкм и диаметром 10-20 мкм (рис. 29). Каждый кардиомиоцит имеет 1-2 овальных удлиненных ядра, лежащих в центре и окруженных микрофибриллами, расположенными на периферии строго прямолинейно. На обоих полюсах ядра видны удлиненные зоны цитоплазмы, лишенной миофибрилл. Весьма характерны контакты двух соседних кардиомиоцитов, имеющих вид извилистых темных полосок, вставочных дисков, которые активно участвуют в передаче возбуждения от клетки к клетке. Клетки богаты митохондриями. Сарколемма кардиомиоцитов толщиной около 9 нм имеет множество микропиноцитозных инвагинаций, пузырьков. По мере старения человека в его кардиомиоцитах накапливается липофусцин.
Строение миофибрилл кардиомиоцитов аналогично таковому скелетных мышц. В периферических отделах кардиомиоцитов и между митохондриями находится множество частичек гликогена и элементов незернис- той эндоплазматической сети. В кардиомиоцитах имеется очень большое количество крупных митохондрий с хорошо развитыми кристами, которые располагаются группами между миофибриллами. На уровне Z-линий цитолемма кардиомиоцитов также формирует Т-трубочки, вблизи которых сосредоточены скопления цистерн незернистой эндоплазматической сети. Однако триады выражены менее четко, чем в скелетных мышцах.
Кардиомиоциты соединены между собой вставочными дисками, которые на продольном разрезе имеют вид ступенек. В этих участках кардиомиоциты соединяются между собой наподобие зубчатых швов чере- па. Сарколемма соседних клеток соединена с помощью десмосом, лентовидных поясков или пятен сцепления, к которым с обеих сторон прикрепляются актиновые филаменты. Поперечные участки расположены на месте Z-линий. Между кардиомиоцитами (в эндомизиуме) располагаются кровеносные капилляры.
Миоэпителиоциты (эктодермального происхождения) - многоотростчатые клетки, в цитоплазме которых имеются способные сокращаться филаменты, состоящие из мышечных белков. Миоэпителиоциты окружают начальные отделы молочных, потовых, слезных, слюнных желез и, сокращаясь, способствуют выведению секрета из клетки. Мионевроциты радужной оболочки глаза, образующие мышцы, суживающие и расширяющие зрачок, являются производными нейроэктодермы. Миоэпителиоциты и мионевроциты иннервируются вегетативной нервной системой.
НЕРВНАЯ ТКАНЬ
Нервная ткань (textus nervosus) образует центральную нервную систему (головной и спинной мозг) и периферическую нервную систему - нервы, нервные волокна с их концевыми приборами, нервные узлы (ганглии). Нервная ткань состоит из нервных клеток - нейронов (нейроцитов) с особым строением и функцией, и нейроглии, которая выполняет опорную, трофическую, защитную и разграничительную функции.
Нейроцит (нейрон) (neurocdtus) с отходящими от него отростками является структурно-функциональной единицей нервной системы. Ос- новная функция нейрона - это получение, переработка, хранение, проведение и передача информации, закодированной в виде электрических или химических сигналов (нервных импульсов). В нейроне различают тело, где информация обрабатывается, и отростки, отходящие от тела клетки. Один или несколько древовидно ветвящихся отростков, по которым нервный импульс приносится к телу нейрона, называется дендритом. Единственный отросток, по которому нервный импульс направляется от нервной клетки, - аксон, или нейрит. Нервная клетка динамически поляризована, т. е. способна пропускать нервный импульс только в одном направлении - от дендрита к аксону.
В зависимости от числа отростков различают униполярные, или одноотростчатые, биполярные, или двухотростчатые, и мультиполярные, или многоотростчатые нейроны (рис. 30). К биполярным относятся и ложноуниполярные нейроны (рецепторные нейроны спинномозговых и черепных ганглиев), у которых короткий отросток вскоре Т-образно разветвляет- ся на аксон и дендрит.
Размеры тела нервных клеток колеблются в пределах от 4-5 до 130-140 мкм, а длина отростков может достигать
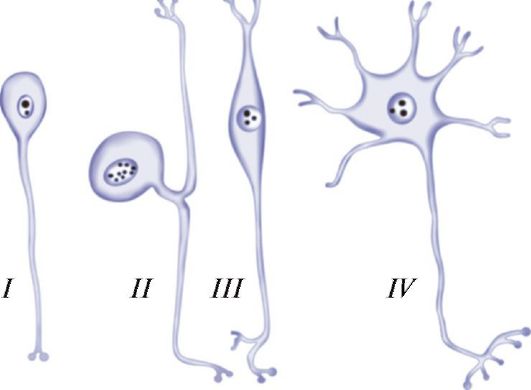
Рис. 30. Типы нейронов: I - униполярный; II - ложноуниполярный; III - биполярный; IV - мультиполярный (по В.Г. Елисееву и др.)
ложноуниполярные клетки спинномозговых узлов имеют округлое тело, ганглионарные мультиполярные нейроны спинного мозга имеют тело неправильной формы, множество слабо ветвящихся дендритов, отходящих в разные стороны, и длинный аксон, от которого отходят коллатерали. От треугольных тел больших пирамидных нейронов коры головного (большого) мозга отходит большое число коротких горизонтальных слабоветвящихся дендритов. Аксон у этих нейронов отходит от основания клетки. В отличие от дендритов, диаметр аксона не меняется на всем протяжении. Грушевидные нейроны коры мозжечка имеют два крупных дендрита, сильно ветвящихся наподобие кроны дерева. Длинный аксон отходит от вершины клетки.
Различают два типа мультиполярных нейронов: длинноаксонные с большим количеством дендритов (клетки типа Гольджи I) и короткоаксонные (клетки типа Гольджи II) с множеством сильно ветвящихся дендритов. К типу I относятся нейроны симпатических и парасимпатических узлов, большие пирамидные нейроны коры головного мозга, грушевидные нейроны коры мозжечка, двигательные нейроны спинного мозга. Эти нейроны передают нервные импульсы на большие расстояния. К типу II относится множество клеток (нейронов) центральной нервной системы, которые передают нервные импульсы многим соседним нейронам.
В сером веществе полушарий большого мозга и мозжечка нейроны располагаются слоями, в других отделах нервной системы образуют скопления (ядра).
Как правило, нейроны являются одноядерными клетками; два ядра имеют некоторые нейроны ганглиев вегетативной нервной системы. Сферическое ядро диаметром около 17 мкм в большинстве нейронов занимает центральное положение (рис. 31). Гетерохроматин располагается по всему ядру. Хорошо выражено базофильное ядрышко, иногда их несколько. В перикарионе находятся многочисленные сферические или удлиненные митохондрии диаметром около 0,1 мкм. Вблизи ядра располагаются десмосомы, связанные между собой элементами комплекса Гольджи. Часто в зоне Гольджи обнаруживаются мультивезикулярные тельца.
Основными особенностями строения нейронов является наличие многочисленных нейрофибрилл и скоплений хроматофильной субстанции (вещество Ниссля), которые представляют собой группы параллельных цистерн зернистой цитоплазматической сети и полирибосомы, богатые РНК. Элементы незернистой эндоплазматической сети в теле клетки малочисленны. Они имеются лишь в аксонах и дендритах в виде трубочек, цистерн и пузырьков. Хроматофильная субстанция и свободные
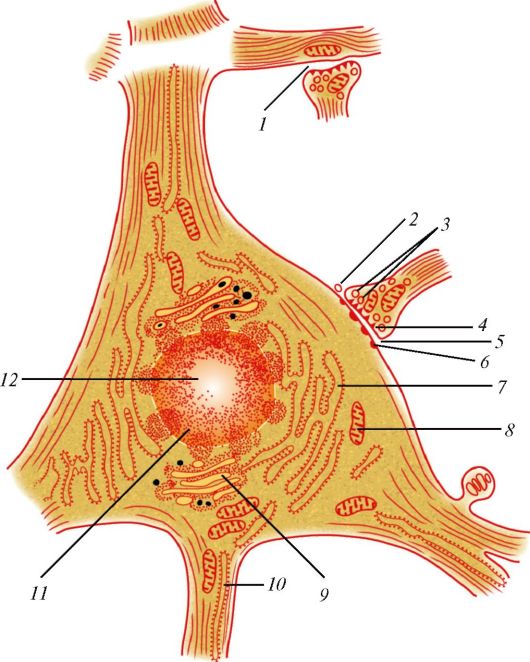
Рис. 31. Схема ультрамикроскопического строения нервной клетки: 1 - аксонодендритический синапс; 2 - аксоносоматический синапс; 3 - преси- наптические пузырьки; 4 - пресинаптическая мембрана; 5 - синаптическая щель; 6 - постсинаптическая мембрана; 7 - эндоплазматическая сеть; 8 - митохондрия; 9 - внутренний сетчатый аппарат (комплекс Гольджи); 10 - нейрофибриллы; 11 - ядро; 12 - ядрышко
рибосомы располагаются по всей цитоплазме клетки и в дендритах, она отсутствует в основании (холмике) аксона и в самом аксоне.
Между элементами эндоплазматической сети расположены многочисленные митохондрии, лизосомы, гранулы липофусцина. Нейрофибриллы, переходящие из тела нейрона в его отростки, образованы микротрубочками диаметром около 20 нм и нейрофиламентами толщиной 7-10 нм.
Нейрофибриллы формируют в перикарионе густую трехмерную сеть, в ячейках которой лежат лизосомы, а также пронизывают цитоплазму отростков. Нейрофибриллы обеспечивают прочность перикариона и от- ростков, осуществляют химическую интеграцию клетки.
Макромолекулы, синтезируемые в перикарионе, направляются в самые отдаленные участки отростков. Посредством постоянного медлен- ного транспорта макромолекул со скоростью 1-3 мм/сут доставляются ферменты, участвующие в синтезе медиаторов в пресинаптической части синапсов, и белки цитоскелета. Быстрый антероградный транспорт со скоростью около 400 мм/сут доставляет пузырьки в синаптические окончания. Кроме того, существует ретроградный транспорт от окончаний аксона к перикариону со скоростью 200-300 мм/сут, с помощью которого более крупные везикулы переносят частицы структур и веще- ства, подлежащие перевариванию в лизосомах. В дендритах также происходит медленный и быстрый транспорт.
В направлении длинной оси дендрита проходит множество нейротрубочек и небольшое количество нейрофиламентов. В цитоплазме дендритов находятся удлиненные митохондрии и небольшое количество цистерн незернистой эндоплазматической сети. Субстанция Ниссля имеется в крупных дендритах, в мелких дендритах элементы незернистой эндоплазматической сети встречаются реже. Конечный отдел дендритов часто колбообразно расширен.
Диаметр аксонов различных клеток (вместе с оболочками) колеблется в широких пределах (от 1 до 20 мкм). Толстые аксоны проводят нервные импульсы быстрее, чем тонкие. Аксоны отходят от конического аксонного холмика, вблизи которого от аксона ответвляются коллатерали. Аксон заканчивается телодендроном - множеством концевых разветв- лений, которые образуют синапсы (контакты). Поверхность цитолеммы аксона (аксолеммы) гладкая. Аксолемма начального сегмента аксона и участка аксона в области узла нервного волокна (перехвата Ранвье) утолщена. В аксоплазме находятся тонкие удлиненные митохондрии, большое количество нейротрубочек и нейрофиламентов, пузырьки и трубочки незернистой эндоплазматической сети, единичные мультивезикулярные тельца. Рибосомы и элементы зернистой эндоплазматической сети отсутствуют в аксоплазме и имеются только в цитоплазме холмика аксона, где расположены пучки микротрубочек, в то время как количество нейрофиламентов здесь невелико.
Итак, нейроны воспринимают, проводят и передают электрические сигналы (нервные импульсы). В основе передачи нервных импульсов лежит изменение мембранного потенциала, вызванного перемещением через
мембрану ионов натрия и калия благодаря функционированию натрийкалиевого насоса.
Нейроны, которые передают возбуждение от точки восприятия раздражения в центральную нервную систему и далее к рабочему органу, связаны между собой с помощью множества межклеточных контактов - синапсов (от греческого syndapsis - связь), передающими нервный импульс от одного нейрона к другому. В синапсах происходит преобразование электрических сигналов в химические и обратно - химических в электрические.
В зависимости от того, какие части нейрона соединены между собой, различают синапсы аксосоматические, когда окончание аксона одного нейрона образует контакты с телом другого; аксодендрические, когда аксоны вступают в контакт с дендритами; а также аксоаксональные, когда контактируют одноименные отростки-аксоны. Такое устройство цепочек нейронов создает возможность для проведения возбуждения от места образования нервных импульсов в спинной или головной мозг и из мозга к рабочим органам (мышцам, железам). При этом передача импульса осуществляется с помощью биологически активных веществ (химическая передача), а сами вещества, осуществляющие передачу, называются нейромедиаторами (от лат. medidator - посредник). Роль медиаторов выполняют норадреналин, ацетилхолин, некоторые моноамины (адреналин, серотонин, дофамин и аминокислоты - глицин, глутаминовая кислота, ГАМК), нейропептиды (энкефалины, нейротензин, ангиотензин II, вазоактивный кишечный пептид, соматостатин, вещество Р и т. д.).
В каждом межнейронном синапсе различают пресинаптическую и постсинаптическую части, разделенные синаптической щелью (рис. 32). Нервный импульс поступает по нервному окончанию в булавовидную пресинаптическую часть нервного окончания, которая ограничена пресинапти- ческой мембраной. Цитоплазма пресинаптической части содержит большое количество округлых мембранных синаптических пузырьков диаметром от 4 до 20 нм, содержащих медиатор. Когда нервный импульс достигает пресинаптической части, открываются кальциевые каналы. Ионы кальция проникает в цитоплазму пресинаптической части, в результате чего его концентрация кратковременно возрастает. При повышении содержания кальция синаптические пузырьки, содержащие нейромедиатор, внедряются в описанные ячейки, сливаются с мембраной, и нейромедиатор выделяется в синаптическую щель, заполненную аморфным веществом умеренной электронной плотности. Чем выше содержание ионов кальция, тем больше синаптических пузырьков выделяют нейромедиаторы. Нейромедиа - тор связывается с рецептором постсинаптической мембраны, что ведет к изменению ее потенциала - возникает постсинаптический потенциал.
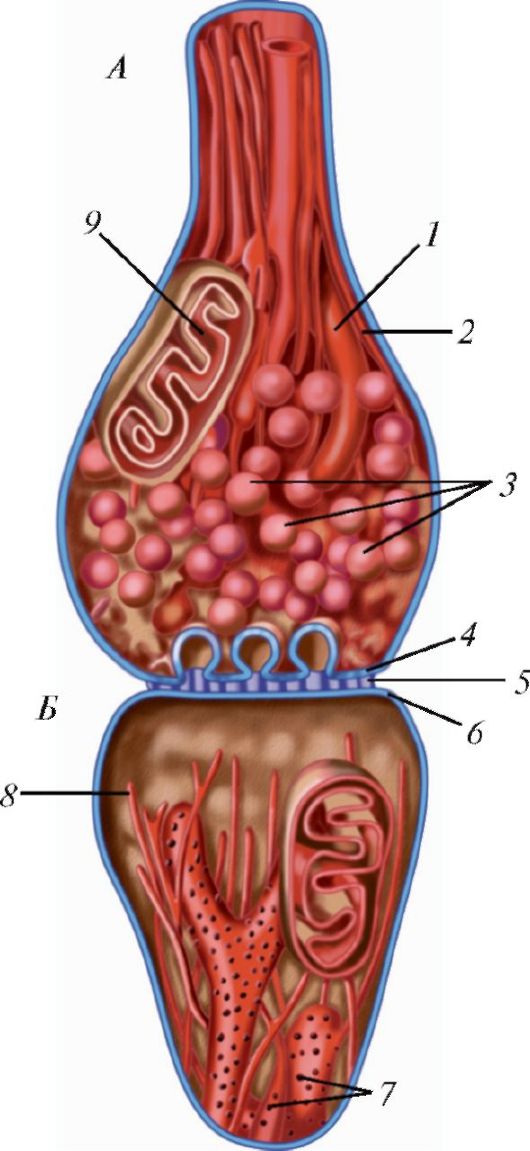
Рис. 32. Схема строения синапса: А - пресинаптическая часть; Б - постсинаптическая часть; 1 - гладкий эндоплазматический ретикулум; 2 - нейротрубочка; 3 - синаптические пузырьки; 4 - пресинаптическая мембрана с гексагональной сетью; 5 - синаптическая щель; 6 - постсинаптическая мембрана; 7 - зернистая эндоплазматическая сеть; 8 - нейрофиламенты; 9 - митохондрия
Таким образом, постсинаптическая мембрана преобразует химический стимул в электрический сигнал. Открываются Na+, К+-каналы, ионы натрия устремляются в постсинаптический полюс, а ионы калия выходят в синаптическую щель, в результате чего происходит деполяризация постсинаптической мембраны. Это ведет к изменению мембранного по- тенциала и возникновению электрического сигнала, величина которого прямо пропорциональна количеству нейромедиатора. Как только прекращается выделение нейромедиатора, пресинаптическое окончание поглощает медиатор из синаптической щели. После этого рецепторы постсинаптической мембраны блокируются антагонистом и возвращаются в исходное состояние.
Нейроглия. Кроме нейронов в нервной системе имеются клетки ней- роглии (neuroglia), выполняющие многообразные функции: опорную,
Рис. 33. Виды нейроглии: I - эпендимоциты; II - протоплаз- матические астроциты; III - волокнистые астроциты; IV - олигоденд- роглиоциты; V - микроглия (по В.Г. Елисееву и др.)
трофическую, защитную, изолирующую, секреторную (рис. 33). Различают две группы нейроглии: глиоциты, или макроглию (эпендимоциты, олигодендроциты и астроциты), и микроглию.
Макроглия. Эпендимоциты (ependymocytus) кубической или призматической формы выстилают изнутри желудочки мозга и спинномозговой канал. Эпендимоциты соединены между собой запирающими зонами и лентовидными десмосомами. От базальной поверхности некоторых эпендимоцитах отходит отросток, который проходит между подлежащими клетками, разветвляется и контактирует с базальным слоем капилляров. Под слоем эпендимоцитов лежит слой недифференцированных глиоцитов.
Астроциты (astrocytus) являются основными глиальными элементами центральной нервной системы. Различают протоплазматические и волокнистые астроциты. Протоплазматические астроциты имеют звездчатую форму, на их телах образуются многочисленные короткие выпячивания, служащие как бы опорой для отростков нейронов, отде- ленных от цитолеммы астроцита щелью шириной около 20 нм. Многочисленные отростки протоплазматических астроцитов заканчиваются на нейронах и на капиллярах. Отростки астроцитов образуют сеть, в которой залегают нейроны. Указанные отростки расширяются на концах, переходя в широкие ножки, которые контактируют между собой. Эти ножки со всех сторон окружают нейроны и кровеносные капилляры, покрывая около 80% их поверхности (вокругсосудистая глиальная пограничная мембрана, membrana limitans glialis perivascularis). He покрыты этой мембраной лишь участки синапсов. Глиальная мембрана, образованная расширенными концами отростков астроцитов, изолирует нейроны, создавая для них специфическое микроокружение. Отростки, достигающие расширенными окончаниями поверхности мозга, соединяясь между собой нексусами, образуют на ней сплошную поверхностную глиальную пограничную мембрану. На этой пограничной мембране лежит базальная мембрана, отграничивающая ее от мягкой мозговой оболочки.
Волокнистые астроциты преобладают в белом веществе центральной нервной системы. Это многоотростчатые (20-40 отростков) клетки, тела которых имеют размеры около 10 мкм. Отростки располагаются между нервными волокнами, некоторые отростки достигают кровеносных капилляров.
Олигодендроциты (oligodendrocytus) - это мелкие клетки овоидной формы (6-8 мкм) с крупным, богатым хроматином ядром, окруженным тонким ободком цитоплазмы, в которой находятся умеренно развитые
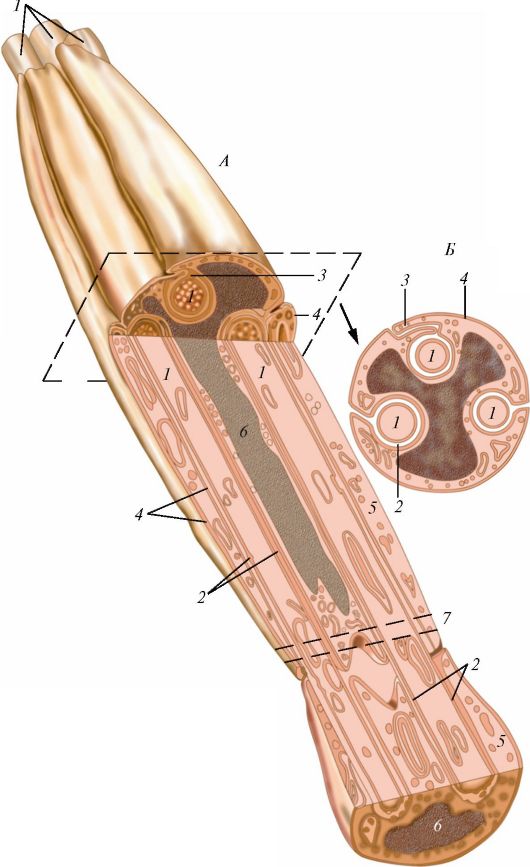
Рис. 34. Схема строения безмиелинового (безмякотного) нервного волокна: А - продольный срез; Б - поперечный срез; 1 - осевые цилиндры; 2 - аксолемма; 3 - мезаксон; 4 - клеточная оболочка нейролеммоцита (шванновской клетки); 5 - цитоплазма нейролеммоцита; 6 - ядро нейролеммоцита; 7 - контакт двух нейролеммоцитов (по В.Г. Елисееву и др.)
органеллы. Олигодендроциты располагаются вблизи нейронов и их отростков. От тел олигодендроцитов отходит небольшое число коротких конусовидных и широких плоских трапециевидных миелинобразующих отростков. Эти отростки формируют миелиновый слой нервных волокон в центральной нервной системе. Миелинобразующие отростки каким-то образом спирально накручиваются на аксоны.
Микроглия (microglia), составляющая около 5% клеток глии в белом веществе мозга и около 18% в сером, состоит из мелких удлиненных клеток угловатой или неправильной формы, рассеянных в белом и сером веществе центральной нервной системы (клетки Ортега). От тела клетки отходят многочисленные отростки, напоминающие кустики. Основания некоторых клеток микроглии как бы распластаны на капилляре.
Нервные волокна (neurofibrae) представляют собой отростки нервных клеток, окруженные оболочками, образованными олигодендроцитами (нейролеммоциты, или шванновские клетки). Различают миелиновые и безмиелиновые волокна. У безмиелиновых волокон, встречающихся преимущественно в нервах вегетативной нервной системы, отростки нейронов (осевые цилиндры) прогибают цитоплазматическую мембрану олигодендроцита (нейролеммоцита), которая окутывает (окружает) эти отростки (рис. 34). В одну шванновскую клетку может быть погружено несколько осевых цилиндров.
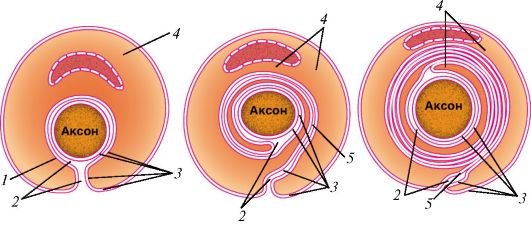
Рис. 35. Формирование миелинового (мякотного) нервного волокна:
1 - контакт аксолеммы и цитолеммы нейролеммоцита (шванновской клетки);
2 - межклеточная щель; 3 - аксолемма и цитолемма нейролеммоцита; 4 - цито-
плазма нейролеммоцита; 5 - мезаксон (по В.Г. Елисееву и др.)
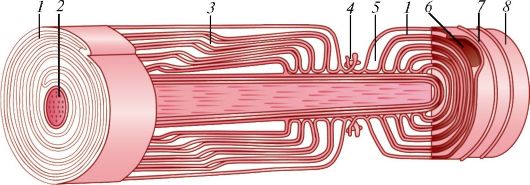
Рис. 36. Строение миелинового (мякотного) нервного волокна: 1 - мезаксон; 2 - осевой цилиндр; 3 - насечки нейролеммы; 4 - узел нервного волокна; 5 - цитоплазма нейролеммоцита; 6 - ядро нейролеммоцита; 7 - нейро- лемма; 8 - эндоневрий (по В.Г. Елисееву и др.)
Миелиновые нервные волокна толще
безмиелиновых, образуются благодаря тому, что нейролеммоцит
(шванновская клетка) спирально накручивается на осевой цилиндр (аксон)
нервной клетки (рис. 35). Миелин - это многократно закрученный двойной
слой цитоплазматической мембраны нейролеммоцита. Толстая и плотная
миелиновая оболочка, богатая липидами, изолирует нервное волокно и
предотвращает утечку нервного импульса из осевого цилиндра. Каждый
нейролеммоцит окутывает только часть осевого цилиндра длиной около
В зависимости от функции выделяют три основных типа нейронов: афферентные, ассоциативные и эфферентные. Чувствительные, рецепторные, или афферентные, нейроны (от лат. afferdns - приносящий) проводят нервные импульсы от органов и тканей в мозг. Тела таких нейронов у человека лежат вне центральной нервной системы. Как правило, это би- полярные (ложноуниполярные) нейроны. Один из отростков, отходящих от тела чувствительной нервной клетки, следует на периферию и заканчивается тем или иным чувствительном окончанием (рецептором), который способен трансформировать энергию внешнего раздражителя в нервный импульс. Второй отросток направляется в головной или спинной мозг. В зависимости от локализации различают: 1) экстерорецеп- торы, воспринимающие раздражение внешней среды. Они расположены
в наружных покровах тела, в коже и слизистых оболочках, в органах чувств; 2) интерорецепторы, получающие раздражение главным образом при изменении химического состава внутренней среды (хеморецепто- ры) и давления в тканях и органах (барорецепторы, механорецепторы); 3) проприорецепторы, воспринимающие раздражение в мышцах, сухожилиях, связках, фасциях, костях, суставных капсулах и т. д.
В зависимости от характера восприятия раздражения различают терморецепторы, механорецепторы и ноцирецепторы. Терморецепторы воспринимают изменение температуры, механорецепторы улавливают различные виды механических воздействий (прикосновение к коже, ее сдавление), ноцирецепторы воспринимают болевые раздражения.
Среди нервных окончаний различают свободные нервные окончания, не имеющие какой-либо оболочки, и несвободные (инкапсулированные), которые покрыты тонкой соединительнотканной пластинкой или слоем клеток нейроглии (рис. 37).
Свободные (неинкапсулированные) нервные окончания (terminatio nervi libera) имеются в коже. Подходя к эпидермису, нервное волокно теряет миелин, проникает через базальную мембрану в эпителиальный слой кожи. Нервные волокна разветвляются между эпителиоцитами вплоть до зернистого слоя, их веточки диаметром менее 0,2 мкм колбообразно расширяются на концах. Аналогичные концевые нервные окончания имеются в эпителии слизистой оболочки и роговицы глаза. Концевые свободные нервные окончания воспринимают боль, тепло и холод.
В эпидермисе свободные нервные окончания подходят также к измененным эпителиоцитам (клеткам Меркеля), которые имеют пальцевидные выросты. Эти окончания являются механорецепторами, воспри- нимающими давление.
Рис. 37. Схема строения рецепторов: А: 1 - свободное нервное окончание; 2 - граница между дермой и эпидермисом; Б: 1 - измененная клетка эпидермиса (клетка Меркеля); 2 - базальная мембрана; 3 - конечный диск афферентного волокна; 4 - миелин; 5 - нейролеммоцит; В: 1 - субкапсулярное пространство; 2 - капсула; 3 - миелин; 4 - нейролеммоцит; 5 - наружная колба; 6 - базальная мембрана; 7 - внутренняя колба; 8 - терминальный отросток афферентного волокна; Г: 1 - уплощенные нейролеммоциты; 2 - капсула; 3 - базальная мембрана; 4 - спиральные терминали афферентного волокна; 5 - нейролеммоциты; 6 - миелин; Д: 1 - пучки коллагеновых волокон в ядре тельца; 2 - терминальные ветви афферентного волокна; 3 - капсула; Е: 1 - терминальные ветви афферентного волокна; 2 - капсула (по А. Хэму
и Д. Кормаку)
Барорецепторы представляют собой ветвящиеся свободные нервные окончания, залегающие в адвентициальном слое крупных артерий грудной полости и шеи. Наиболее важные из них - рецепторы, залегающие в стенках дуги аорты и сонного синуса. При растяжении стенки артерии под влиянием изменяющегося артериального давления эти окончания возбуждаются. Участие барорецепторов в регуляции артериального давления осуществляется по принципу обратной связи.
Несвободные (инкапсулированные) нервные окончания - это осязательные тельца (Мейсснера), пластинчатые (Фатера-Пачини), луковицеобразные (Гольджи-Маццони). Все эти нервные окончания являются механорецепторами.
Пластинчатые тельца Фатера-Пачини - самые крупные из всех инкапсулированных нервных окончаний. Они имеют овальную форму, длину 3-4 мм и толщину
Осязательные тельца (Мейсснера) длиной 50-160 мкм, шириной около 60 мкм, овальные или цилиндрические, особенно многочисленны в сосочковом слое кожи пальцев рук, а также в коже стопы. Тельце образовано удлиненными, уплощенными шванновскими клетками, лежащими одна на другой. Нервное волокно, входя в тельце, теряет миелин и оканчивается колбообразным расширением. Периневрий переходит в окружающую тельце капсулу, образованную несколькими слоями эпителиоидных периневральных клеток. Сдавление этих клеток передается осязательному тельцу, вызывая деформацию его клеток, что приводит к возникновению импульса в нервных волокнах. Таким образом, мейсснеровские тельца являются механорецепторами, воспринимающими прикосновение, сдавление кожи.
Концевые колбы (Краузе) расположены в коже, конъюнктиве глаз, слизистой оболочке ротовой полости. Сферические тельца окружены толстой соединительнотканной капсулой, богатой коллагеновыми волокнами и фибробластами. Войдя в капсулу, нервное волокно теряет миелиновую оболочку и разветвляется в центре колбы, образуя множество веточек. Колбы Краузе воспринимают холод, возможно, они являются и механорецепторами.
Рис. 38. Схема пластинчатого тельца Фатера - Пачини: А - продольный срез: 1 - соединительнотканная капсула; 2, 5, 8 - гемокапилляры; 3 - пластинки наружной луковицы; 4 - коллагеновые микрофибриллы; 6 - периневрий; 7 - миелиновое нервное волокно; 9 - аксон; 10 - миелин; 11 - внутренняя луковица; 12 - окончание нервного волокна; Б - поперечный срез: 1 - интердигитирующие полулуковицы внутренней луковицы; 2 - радиальная щель; 3 - нервное волокно (по Р. Крстичу, с изменениями)
Проприорецепторы воспринимают тягу сокращения мышц, натяжения сухожилий и суставных капсул, возникающих при выполнении определенного движения или удержания частей тела в определенном положении. Это нервно-мышечные и нервно-сухожильные веретена, которые находятся в брюшке мышц или в их сухожилиях. Сухожильные органы (Гольджи) расположены внутри сухожилий, поблизости от мышц.
Нервно-мышечное веретено крупное (длиной 3-5 мм и толщиной до
Вставочный (ассоциативный), или кондукторный, нейрон осуществляет передачу нервного импульса с чувствительного (центростремительного) нейрона на двигательный или секреторный (центробежный).
Тела эфферентных (эффекторных, двигательных или секреторных) нейронов (от лат. efferens - выносящий) находятся в центральной нервной системе или в симпатических и парасимпатических нервных узлах. Аксоны эфферентных нейронов несут нервные импульсы к рабочим органам (мышцам или железам), а также ко всем органам и тканям для иннервации (регуляции) обмена веществ. Выделяют также двигательные и секреторные
окончания аксонов выносящих (эфферентных) нейронов. Аксоны двигательных нейронов разветвляются и каждый из них ин- нервирует большое количество мышечных волокон. Двигательные (моторные) нервные окончания располагаются на мышечных волокнах. Окончание одного двигательного нервного окончания и иннервируемое им поперечнополосатое мышечное волокно образуют двигательную единицу - мион.
Каждое мышечное волокно иннервируется веточкой аксона двигательного нейрона, который, оканчиваясь на волокне,
Рис. 39. Нервно-мышечная пластинка: 1 - мышечные волокна; 2 - моторные бляшки
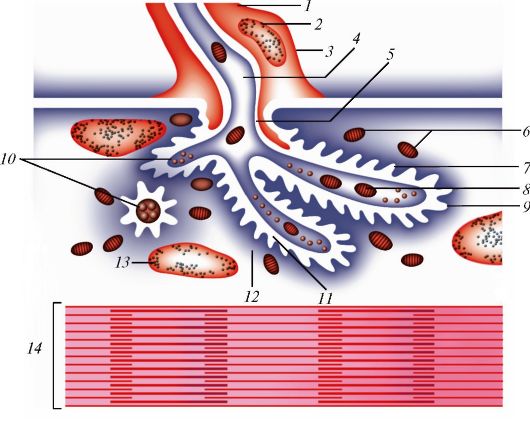
Рис. 40. Строение аксомышечного синапса: 1 - нейролеммоцит; 2 - ядро нейролеммоцита; 3 - нейролемма (цитоплазматическая мембрана нейролеммоцита); 4 - аксоплазма; 5, 11 - аксолемма (пресинаптическая мембрана); 6 - митохондрии мышечного волокна; 7 - синаптическая щель; 8 - митохондрия нервного окончания; 9 - сарколемма (постсинаптическая мембрана); 10 - синаптические пузырьки; 12 - сарколемма; 13 - ядро мышечного волокна; 14 - миофибриллы (по В.Г. Елисееву и др.)
образует нейромышечное окончание, или двигательную концевую пластинку (рис. 39), у которой различают пресинаптическую и постсинаптическую мембраны, разделенные синаптической щелью (рис. 40).
Нервный импульс, достигающий нервно-мышечного окончания, приводит к выходу в синаптическую щель ацетилхолина, который связывается со специфическими рецепторами мембраны мышечного волокна.
Нервные окончания неисчерченной (гладкой) мышечной ткани располагаются между гладкомышечными клетками, где образуют расширения - синаптические пузырьки, плотно прилежащие к базальной мембране миоцитов.