Анатомия человека : Сапин М. Р., Билич Г. Л. учебник в 3 т. - изд. 3-е испр. 2007. - Т. 1, Т. 2, Т. 3.
|
|
|
|
Воздействия внешней среды воспринимаются органами чувств. В органах чувств образуется нервный импульс, который по нервам попадает в определенные отделы мозга, в том числе и в кору полушарий большого мозга, где происходит высший анализ. К органам чувств относятся органы зрения, слуха, чувства земного тяготения (гравитации), вкуса, обоняния, кожного чувства. С помощью органов чувств человек не только «ощущает» внешний мир. Благодаря особому развитию головного мозга, высокой дифференцировке его структур и органам чувств у человека сформировались особые, присущие только ему социальные формы восприятия внешнего мира и самосознание.
Под влиянием различных воздействий внешней среды в периферической части анализаторов (чувствительных приборах - нервных окончаниях) возникают нервные импульсы, которые по нервным волокнам (проводящим путям) передаются в корковые концы анализаторов - чувствительные центры в коре полушарий большого мозга. Нервные импульсы по проводящим путям поступают также в ядра (нервные центры) спинного мозга, ствола головного мозга, промежуточного мозга. Эти центры, в свою очередь, при участии восходящих (афферентных) и ассоциативных проводящих путей связаны с корой большого мозга и с другими центрами нервной системы.
ОРГАН ЗРЕНИЯ
Орган зрения состоит из глазного яблока, расположенного в глазнице, и вспомогательных органов глаза.
ГЛАЗ
Глазное яблоко (bulbus oculi) имеет шаровидную форму. У него выделяют передний и задний полюсы (рис. 117). Передний полюс (polus anterior) - это наиболее выступающая точка роговицы. Задний полюс (polus posterior) расположен латерально от места выхода из глазного яблока зрительного нерва. Соединяющая оба полюса условная линия называется наружной осью глазного яблока (axis bulbi externus). Эта ось равна примерно 24 мм и находится в плоскости меридиана глазного яблока. Расстояние от задней поверхности роговицы до сетчатки называется внутренней осью глазного яблока (axis bulbl internus), она равна 21,75 мм. Плоскость, перпендикулярная наружной и внутренней осям, разделяющая глазное яблоко на две половины - переднюю и заднюю, образует экватор (equator), равный 23,3 мм. Глазное яблоко относительно велико, его объем у взрослого человека в среднем равен 7,448 см3.
Глазное яблоко состоит из ядра, покрытого тремя оболочками: фиброзной, сосудистой и внутренней, или сетчатой. Снаружи глазное яблоко покрыто фиброзной оболочкой (tunica fibrosa bulbi), которая подразделяется на задний отдел - склеру и прозрачный передний - роговицу. Границей между склерой и роговицей служит борозда склеры (sulcus sclerae). Склера (sclera) - плотная соединительнотканная оболочка толщиной 0,3-0,4 мм в задней части и 0,6 мм вблизи роговицы. Склера образована пучками коллагеновых волокон различных размеров, между которыми залегают уплощенные фибробласты и небольшое количество эластических волокон. Сзади в склере находится решетчатая пластинка (lamina cribosa sclerae), через которую проходят волокна зрительного нерва. В толще склеры, в зоне ее соединения с роговицей имеются мелкие сообщающиеся между собой полости (фонтановы пространства), впадающие в венозный синус склеры (шлеммов канал - sinus venosus sclerae), через который обеспечивается отток жидкости из передней камеры глаза (рис. 118).
Роговица (cornea) - прозрачная выпуклая пластинка блюдцеобразной формы. Ее круговой край - лимб (limbus cornea) переходит в склеру. Толщина роговицы в центре 1-1,2 мм, по периферии - 0,8-0,9 мм. Роговица состоит из пяти слоев: переднего эпителия, передней пограничной пластинки, собственного вещества роговицы, задней пограничной пластинки, заднего эпителия (эндотелий роговицы).
Передний эпителий многослойный плоский неороговевающий, толщиной около 50 мкм. В эпителии множество свободных чувствительных
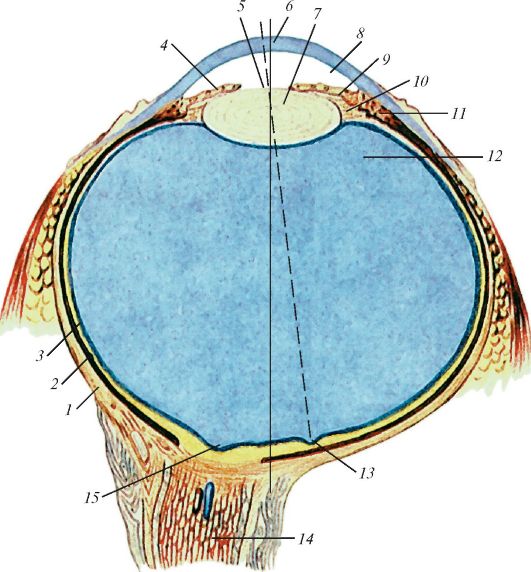 Рис. 117. Строение глазного яблока. Горизонтальный разрез:
Рис. 117. Строение глазного яблока. Горизонтальный разрез:
1 - фиброзная оболочка (склера); 2 - собственно сосудистая оболочка; 3 - сетчатка; 4 - радужка; 5 - зрачок; 6 - роговица; 7 - хрусталик; 8 - передняя камера глазного яблока; 9 - задняя камера глазного яблока; 10 - ресничный поясок;
11 - ресничное тело; 12 - стекловидное тело; 13 - центральная ямка; 14 - зрительный нерв; 15 - диск зрительного нерва. Сплошной линией показана наружная ось глаза, пунктирной - зрительная ось глаза
нервных окончаний. Клетки лежат на базальной мембране, на которой они укреплены множеством полудесмосом. Глубже лежит промежуточный (шиповатый) слой, образованный несколькими слоями скрепленных многочисленными десмосомами клеток. Свободная поверхность роговицы покрыта большим количеством узких микроворсинок и складками, которые удерживают на поверхности роговицы тонкую слезную пленку. Передняя пограничная пластинка (боуменова мембрана) образована переплетающимися тонкими коллагеновыми и ретикулярными фибриллами.
Собственное вещество роговицы толщиной около 0,5 мм, составляет большую часть роговицы. Оно образовано тонкими соединительнотканными (коллагеновыми) пластинками, между которыми лежат уплощенные фибробласты. Пластинки, в свою очередь, состоят из плотно
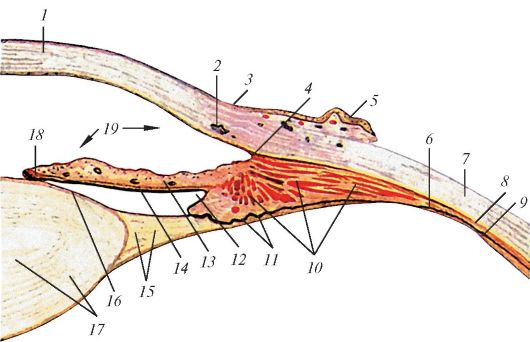 Рис. 118. Строение
передне-боковой части глазного яблока, разрез в горизонтальной
плоскости: 1 - роговица; 2 - венозный синус склеры; 3 - лимб (край
роговицы); 4 - радужно-роговичный угол; 5 - конъюнктива; 6 - ресничная
часть сетчатки; 7 - склера; 8 - сосудистая оболочка; 9 - зубчатый край
сетчатки; 10 - ресничная мышца; 11 - ресничные отростки; 12 - задняя
камера глазного яблока; 13 - радужка; 14 - задняя поверхность радужки;
15 - ресничный поясок; 16 - капсула хрусталика; 17 - хрусталик; 18 -
сфинктер зрачка (мышца, суживающая зрачок); 19 - передняя камера
глазного яблока
Рис. 118. Строение
передне-боковой части глазного яблока, разрез в горизонтальной
плоскости: 1 - роговица; 2 - венозный синус склеры; 3 - лимб (край
роговицы); 4 - радужно-роговичный угол; 5 - конъюнктива; 6 - ресничная
часть сетчатки; 7 - склера; 8 - сосудистая оболочка; 9 - зубчатый край
сетчатки; 10 - ресничная мышца; 11 - ресничные отростки; 12 - задняя
камера глазного яблока; 13 - радужка; 14 - задняя поверхность радужки;
15 - ресничный поясок; 16 - капсула хрусталика; 17 - хрусталик; 18 -
сфинктер зрачка (мышца, суживающая зрачок); 19 - передняя камера
глазного яблока
упакованных пучков тонких коллагеновых фибрилл одинакового диаметра. В передней части роговицы пучки ориентированы косо, в задней - параллельно ее поверхности. Клетки собственного вещества роговицы являются фиброцитами, они скрепляют между собой соединительнотканные пучки и пластинки. Этот слой также чрезвычайно богат нервными окончаниями. Задняя пограничная пластинка (десцеметова оболочка) толщиной 5-10 мкм представляет собой гомогенную эластическую мембрану, в которой встречаются плотные узкие поперечно исчерченные коллагеновые фибриллы. Фибриллы этого слоя погружены в богатое мукополисахаридами аморфное вещество. Десцеметова оболочка напоминает базальную мембрану эпителия. Задний эпителий - это один слой плоских гексагональных эпителиальных клеток. Эпителиоциты, скрепленные между собой простыми и сложными межклеточными соединениями, богаты органеллами, особенно митохондриями и микропиноцитозными пузырьками. Эпителиоциты осуществляют активный транспорт жидкости и ионов и участвуют в синтезе задней пограничной пластинки. Роговица лишена кровеносных сосудов, ее питание происходит путем диффузии из сосудов лимба и из жидкости передней камеры глаза.
Сосудистая оболочка глазного яблока (tunica vasculosa bulbi) расположена под склерой, ее толщина 0,1-0,22 мм. Эта оболочка богата кровеносными сосудами, она состоит из трех частей: собственно сосудистой оболочки, ресничного тела и радужки (рис. 119). Собственно сосудистая оболочка (choroidea) образует большую заднюю часть сосудистой оболочки, ее толщина 0,1-0,2 мм. Основу собственно сосудистой оболочки составляет сосудистая пластинка (lamina vasculosa) - густая сеть переплетающихся между собой тонких артерий и вен, между которыми располагается рыхлая волокнистая соединительная ткань, богатая крупными пигментными клетками и фибробластами, среди которых встречаются макрофагоциты и лаброциты. Сосудистая пластинка снаружи покрыта надсосудистой пластинкой (lamina suprachoroidea), образованной рыхлой волокнистой соединительной тканью, в которой преобладают эластические волокна и содержится множество пигментных клеток.
Под сосудистой пластинкой лежит тонкая сосудисто-капиллярная пластинка (lamina choroidocapillaris), образованная множеством крупных фенестрированных капилляров, в том числе и синусоидных, лежащих на тонкой базальной пластинке и начинающихся от артериол сосудистой
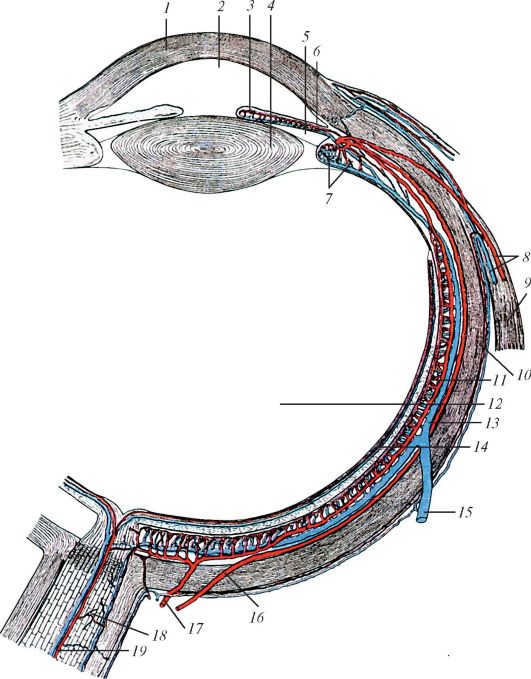 Рис. 119. Сосудистая
оболочка глазного яблока и образующие ее кровеносные сосуды: 1 -
роговица; 2 - передняя камера глаза; 3 - малый артериальный круг
радужки; 4 - хрусталик; 5 - задняя камера глаза; 6 - большой
артериальный круг радужки; 7 - ресничное тело; 8 - передние ресничные
артерия и вена; 9 - латеральная прямая мышца глаза; 10 - склера; 11 -
сосудистая оболочка глаза; 12 - стекловидное тело; 13 - длинная задняя
ресничная вена; 14 - сетчатка; 15 - вортикозная вена; 16 - длинная
задняя ресничная артерия; 17 - короткая задняя ресничная артерия; 18 -
зрительный нерв; 19 - центральная артерия
Рис. 119. Сосудистая
оболочка глазного яблока и образующие ее кровеносные сосуды: 1 -
роговица; 2 - передняя камера глаза; 3 - малый артериальный круг
радужки; 4 - хрусталик; 5 - задняя камера глаза; 6 - большой
артериальный круг радужки; 7 - ресничное тело; 8 - передние ресничные
артерия и вена; 9 - латеральная прямая мышца глаза; 10 - склера; 11 -
сосудистая оболочка глаза; 12 - стекловидное тело; 13 - длинная задняя
ресничная вена; 14 - сетчатка; 15 - вортикозная вена; 16 - длинная
задняя ресничная артерия; 17 - короткая задняя ресничная артерия; 18 -
зрительный нерв; 19 - центральная артерия
сетчатки
пластинки. Капилляры пластинки снабжают фоторецепторные клетки сетчатой оболочки кислородом и питательными веществами. Базальная пластинка (complexus, s. lamina basalis - мембрана Бруха), отделяющая пигментный слой сетчатки от сосудистой оболочки, толщиной 1-2 мкм, состоит из сети эластических волокон, окруженных тонкими коллагеновыми и ретикулярными фибриллами. Базальная пластинка лежит между базальной мембраной пигментного слоя сетчатки и эндотелием капилляров сосудисто-капиллярной пластинки. Между сосудистой оболочкой и склерой имеется система щелей - околососудистое пространство (spdatium perichoroiddale).
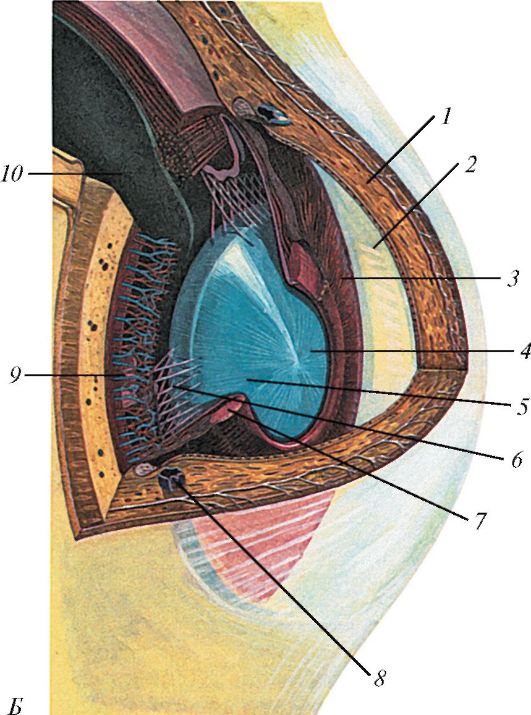
Впереди собственно сосудистая оболочка переходит в утолщенное ресничное (цилиарное) тело (cdrpus cilidre), имеющее кольцевидную форму (рис. 120). Ресничное тело участвует в аккомодации глаза, поддерживая, фиксируя и растягивая хрусталик. На разрезах, проведенных по меридиану глазного яблока, ресничное тело выглядит как треугольник, обращенный основанием к передней камере глаза, а кзади - вершиной, переходящей в собственно сосудистую оболочку (см. рис. 118). Ресничное тело делится на две части: внутреннюю - ресничный венец (cordona cilidaris) и наружную - ресничный кружок (orbiculus ciliaris). Ресничный кружок представляет собой утолщенную циркулярную полоску шириной 4 мм, переходящую в собственно сосудистую оболочку. От поверхности ресничного кружка по направлению к хрусталику отходит ресничный венец, образованный 70-75 ресничными отростками (procdssus ciliares) длиной около 2-3 мм каждый, содержащими в основном кровеносные сосуды (капилляры). К ресничным отросткам прикрепляются соединительнотканные волокна (циннова связка), идущие к хрусталику. Между волокнами связки имеются узкие щели, заполненные водянистой влагой. Из сосудов ресничных отростков (в области ресничного венца) выделяется жидкость - водянистая влага, заполняющая камеры глаза.
Рис. 120. Ресничное тело, ресничные отростки и ресничный венец: А - вид сзади: 1 - фиброзная оболочка (склера); 2 - ресничный венец; 3 - сосудистая оболочка; 4 - ресничный поясок; 5 - хрусталик; 6 - ресничные отростки; 7 - задняя поверхность ресничного тела; 8 - ресничная часть сетчатки; 9 - зубчатый край сетчатки; 10 - сетчатка; 11 - ресничный кружок; Б - вид сбоку, часть роговицы и ресничного тела отрезаны и удалены: 1 - роговица; 2 - передняя камера глазного яблока; 3 - радужка; 4 - зрачок; 5 - хрусталик; 6 - ресничный поясок; 7 - сфинктер зрачка; 8 - венозный синус склеры; 9 - циркулярные мышечные пучки; 10 - меридиональные (продольные) мышечные пучки
 Водянистая влага (humor aquosus) секретируется
сосудами ресничного тела и поступает в заднюю камеру глаза. Из
многочисленных капилляров ресничного тела жидкость и ионы диффундируют к
эпителию, покрывающему ресничное тело. Безпигментные эпителиоциты
особенно активно транспортируют жидкость и вещества, включая
аскорбиновую кислоту.
Водянистая влага (humor aquosus) секретируется
сосудами ресничного тела и поступает в заднюю камеру глаза. Из
многочисленных капилляров ресничного тела жидкость и ионы диффундируют к
эпителию, покрывающему ресничное тело. Безпигментные эпителиоциты
особенно активно транспортируют жидкость и вещества, включая
аскорбиновую кислоту.
Большая часть ресничного тела - ресничная мышца (musculus ciliaris), образованная пучками гладких миоцитов, среди которых различают так называемые меридиональные (продольные) волокна (fibrae meridionales), циркулярные волокна (fibrae circulares) и радиальные волокна (fibrae radiales). Ресничная мышца прикрепляется к выступу склеры - склеральной шпоре. Меридиональные (продольные) мышечные пуч- к и вплетаются в переднюю часть собственно сосудистой оболочки.
При их сокращении эта оболочка смещается кпереди, в результате чего уменьшается натяжение ресничного пояска, на котором укреплен хрусталик. При этом капсула хрусталика расслабляется, хрусталик изменяет кривизну, становится более выпуклым, его преломляющая способность увеличивается. Циркулярные пучки лежат кнутри от меридиональных. При сокращении они суживают цилиарное тело, приближая его к хрусталику, что также способствует расслаблению капсулы хрусталика. Радиальные пучки располагаются в радиарном направлении между меридиональными и циркулярными пучками, сближая их при сокращении. Присутствующие в толще ресничной мышцы эластические волокна расправляют цилиарное тело при расслаблении его мышцы. Миоциты в старческом возрасте частично атрофируются, развивается соединительная ткань; это приводит к нарушению аккомодации.
Строма ресничного тела образована соединительной тканью, пронизанной капиллярной сетью (фенестрированный эндотелий) и венулами. Внутренняя поверхность ресничного тела, обращенная в заднюю камеру глаза, покрыта двумя слоями кубических эпителиоцитов, лежащих на тонкой базальной пластинке (внутренней мембране). Внутренний слой эпителиоцитов образован безпигментными клетками. Наружный слой эпителиоцитов состоит из пигментных клеток, отделенных от стромы ресничного тела базальной мембраной (продолжение ,азальной пластинки). Толщина этой мембраны с возрастом увеличивается. Ресничные отростки, являющиеся продолжением сосудисто-капиллярной пластинки, окружены описанным выше двуслойным эпителием, который лежит на базальной пластинке. По существу ресничное тело и ресничные отростки покрыты со стороны задней камеры глаза ресничной частью сетчатки.
Ресничное тело кпереди продолжается в радужку, которая представляет собой круглый диск толщиной около 0,4 мм с отверстием в центре - зрачком (pupilla). Радужка расположена между роговицей спереди и хрусталиком сзади. Она отделяет переднюю камеру глаза (camera anterior bulbi) от задней камеры глаза (camera posterior bulbi), ограниченной сзади хрусталиком. Зрачковый край радужки (margo pupillaris) зазубрен, латеральный периферический ресничный край (margo ciliaris) переходит в ресничное тело.
Радужка (iris) состоит из пяти слоев. Передний слой образован эпителием, который является продолжением эпителия, покрывающего заднюю поверхность роговицы. Затем следуют наружный пограничный
слой, сосудистый слой, внутренний слой и пигментный слой, выстилающий заднюю поверхность радужки. Наружный пограничный слой образован основным веществом, в котором имеется множество фибробластов и пигментных клеток. Сосудистый слой состоит из рыхлой волокнистой соединительной ткани, в которой залегают многочисленные сосуды, пигментные клетки, фибробласты, лаброциты и гигантские макрофаги (70 - 100 мкм). В цитоплазме макрофагов содержится множество фагоцитированных гранул меланина. В толще сосудистого слоя проходят две мышцы. Циркулярно в зрачковой зоне расположены пучки миоцитов, которые образуют сфинктер (суживатель) зрачка (m. sphincter pupillae). Пучки миоцитов, расширяющих зрачок, образуют дилататор (расширитель) зрачка (m. dilatator pupillae). Миоциты - расширители зрачка, имеют радиальное направление и лежат в задней части сосудистого слоя. В радужке имеются многочисленные отдельные мышечные пучки, которые связывают между собой обе эти мышцы.
Внутренний (пограничный) слой радужки по строению сходен с наружным пограничным слоем. Пигментный слой радужной оболочки является продолжением эпителия, покрывающего ресничное тело и ресничные отростки (рис. 121). Различное количество и качество пигмента меланина, содержащегося в клетках этого слоя, обусловливает цвет глаз - карий, черный при большом количестве пигмента. Если меланоциты имеют мало пигмента, то глаза голубые, зеленые.
Внутренняя (светочувствительная) оболочка глазного яблока - сетчатка на всем протяжении изнутри прилежит к сосудистой оболочке.
Сетчатка (retina) состоит из двух частей: внутренней - светочувствительной (нервной части сетчатки - pars nervosa) и наружной - пигментной (pars pigmentosa). Пигментная часть сетчатки прилежит к сосудистой оболочке глазного яблока, она состоит из пигментных эпителиоцитов кубической формы, содержащих зерна меланина.
Анатомически сетчатку подразделяют на две части - заднюю (зрительную) и переднюю (ресничную и радужковую). Ресничная и радужковая части сетчатки (pars ciliaris et pars iridica retinae) покрывают сзади ресничное тело и радужку и не содержат светочувствительных клеток.
Зрительная часть сетчатки, или нервная часть (pars nervosa), занимает большую заднюю часть глазного яблока. Границей между ресничной частью спереди и зрительной частью сзади является зубчатый край (ora serrata), который находится на уровне перехода собственно сосудистой оболочки в ресничный кружок.
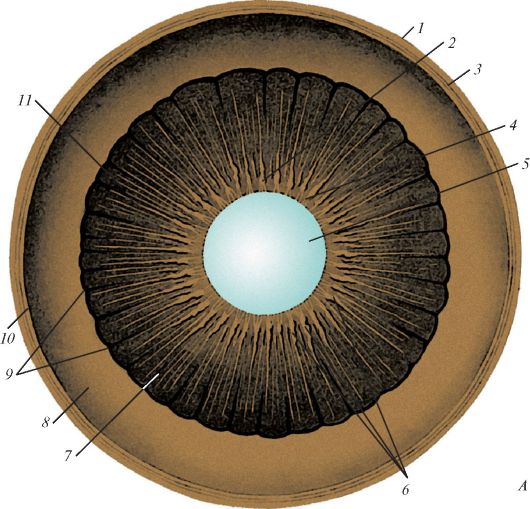
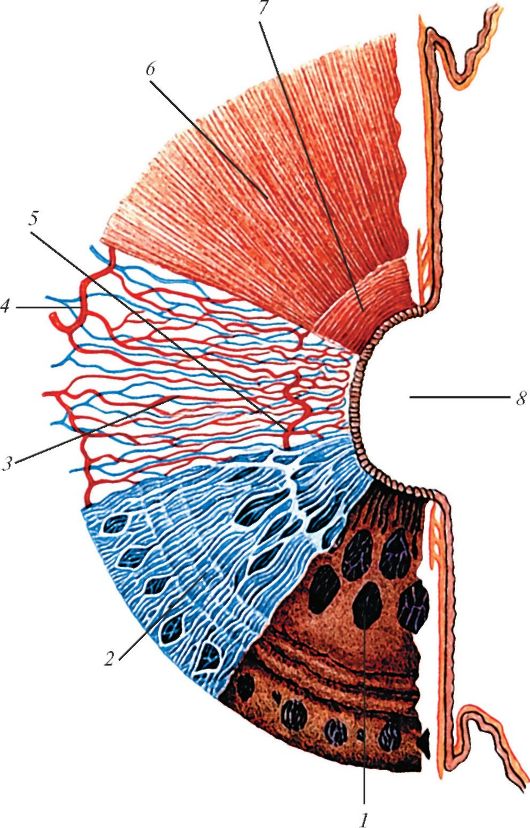 Рис. 121. Строение
радужки глазного яблока, вид спереди (схема): 1 - пигментный эпителий; 2
- внутренний пограничный слой; 3 - сосудистый слой; 4 - большой
артериальный круг радужки; 5 - малый артериальный круг радужки; 6 -
расширитель (дилататор) зрачка; 7 - сфинктер зрачка; 8 - зрачок
Рис. 121. Строение
радужки глазного яблока, вид спереди (схема): 1 - пигментный эпителий; 2
- внутренний пограничный слой; 3 - сосудистый слой; 4 - большой
артериальный круг радужки; 5 - малый артериальный круг радужки; 6 -
расширитель (дилататор) зрачка; 7 - сфинктер зрачка; 8 - зрачок
В задних отделах зрительной части сетчатки находится диск зрительного нерва (discus nervi optici) - место выхода из глазного яблока зрительного нерва. Это «слепое пятно», поскольку здесь нет светочувствительных фоторецепторных клеток. Диаметр диска около 1,7 мм, он имеет приподнятые в виде валика края и небольшое углубление в центре. В центре диска в сетчатку входит центральная артерия сетчатки (рис. 122). Зрительный нерв окружен оболочками (продолжение оболочек головного мозга) и направляется в сторону зрительного канала, открывающегося в полость черепа. Эти оболочки образуют наружное и внутренее влагалища зрительного нерва (vagina externa et vagina interna n. optici) Латеральнее диска на расстоянии около 4 мм имеется овальное углубление - желтое пятно (macula), место наилучшего видения. В области пятна сосуды отсутствуют.
Зрительная часть сетчатки имеет сложное строение. В ней выделяют 10 слоев (пигментный эпителий, фотосенсорный слой, наружный пограничный слой, наружный ядерный слой, наружный сетчатый слой,
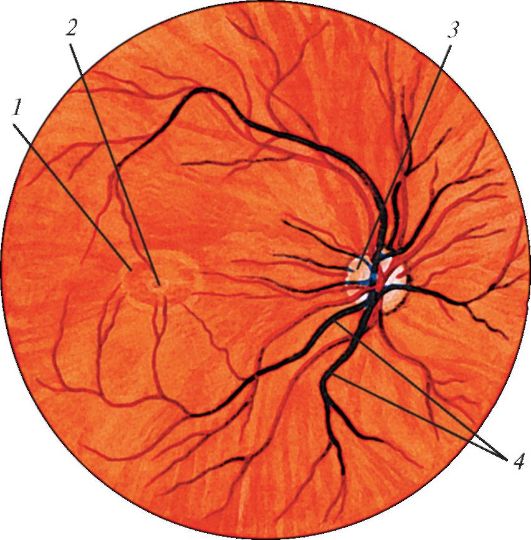 Рис. 122. Офтальмоскопическая
картина глазного дна (задняя часть сетчатки): 1 - пятно; 2 -
центральная ямка; 3 - диск зрительного нерва; 4 - кровеносные
Рис. 122. Офтальмоскопическая
картина глазного дна (задняя часть сетчатки): 1 - пятно; 2 -
центральная ямка; 3 - диск зрительного нерва; 4 - кровеносные
сосуды
внутренний ядерный слой, внутренний сетчатый слой, ганглиозный слой, слой нервных волокон и внутренний пограничный слой).
К пигментному эпителию (первый глубокий слой) прилежит фотосенсорный слой палочковидных (палочек) и колбочковидных (колбочек) зрительных клеток. И палочки, и колбочки представляют собой периферические отростки (наружные сегменты) фоторецепторных палочковидных и колбочковидных зрительных клеток, образующих фотосенсорный слой (второй слой сетчатки). Каждая палочка и колбочка окружена 30-45 отростками пигментоцитов. Третий слой сетчатки (наружный пограничный слой) образован периферическими концами глиоцитов. Этот слой соответствует зоне перехода палочек и колбочек (наружных сегментов) фоторецепторных клеток в их ядросодержащую часть (внутренний сегмент). Ядра палочковидных и колбочковидных фоторецепторных клеток лежат примерно на одном уровне, который (ядросодержащий слой) выделяют в качестве наружного ядерного слоя (четвертый слой сетчатки).
Каждая палочковидная зрительная клетка состоит из наружного и внутреннего сегментов, соединенных между собой связующим отделом (рис. 123). Наружный сегмент - светочувствительный, образован сдвоенными мембранными дисками, являющимися складками цитоплазматической мембраны, в которую встроен зрительный пурпур - родопсин. Внутренний сегмент состоит из двух частей. Ближе к наружному сегменту располагается эллипсоидная часть, заполненная длинными митохондриями, за ней следуют миоидная часть, содержащая эндоплазматическую сеть, свободные рибосомы и комплекс Гольджи. Кнутри клетка сужается, образуя заполненную нейрофибриллами переходную часть, связанную с телом клетки, в котором расположено овальное ядро. От тела клетки отходит короткий отросток (аксон), заканчивающийся на биполярных клетках.
Колбочковидные зрительные клетки отличаются от палочковидных клеток большей величиной и строением дисков, они имеют в дистальной части наружного сегмента впячивания цитоплазматической мембраны, которые образуют полудиски. В проксимальной части наружного сегмента диски колбочек похожи на диски палочек. В эллипсоидном внутреннем сегменте расположены многочисленные удлиненные митохондрии и элементы зернистой эндоплазматической сети. В расширенной базальной части колбочковидной клетки залегает крупное сферическое ядро. От тела клетки отходит короткий аксон, оканчивающийся
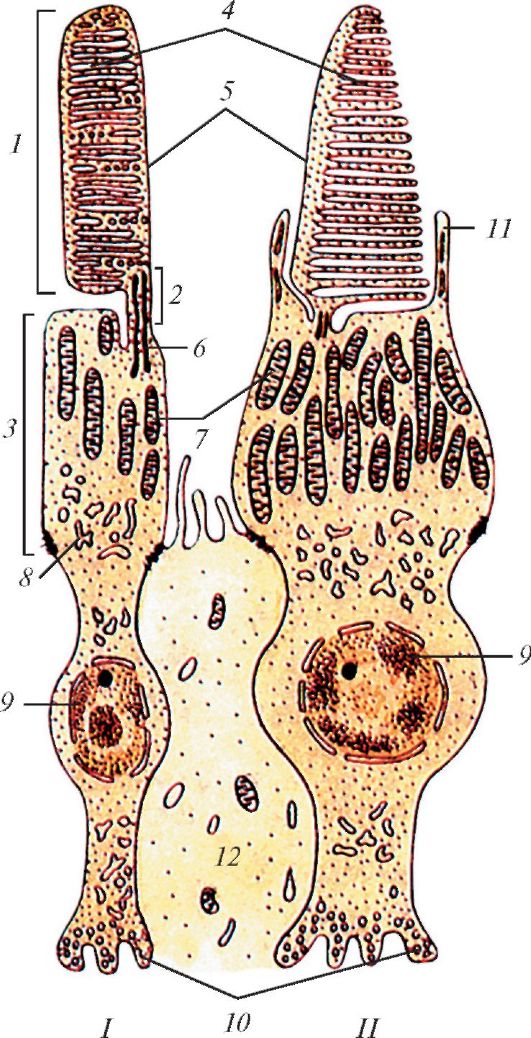 Рис. 123. Палочковидная
(I) и колбочковидная (II) зрительные клетки - фоторецепторные клетки.
Ультрамикроскопическое строение: 1 - наружный сегмент палочки; 2 -
связующий отдел между наружным и внутренним сегментами палочки; 3 -
внутренний сегмент палочки; 4 - диски; 5 - клеточная оболочка; 6 -
двойные микрофибриллы; 7 - митохондрии; 8 - пузырьки эндоплазматической
сети; 9 - ядро; 10 - область синапса с биполярным нейроцитом; 11 -
пальцевидные отростки внутреннего сегмента колбочковидной зрительной
клетки; 12 - лучевой глиоцит (мюллерово волокно) (по И.В. Алмазову и
Л.С. Сутулову)
Рис. 123. Палочковидная
(I) и колбочковидная (II) зрительные клетки - фоторецепторные клетки.
Ультрамикроскопическое строение: 1 - наружный сегмент палочки; 2 -
связующий отдел между наружным и внутренним сегментами палочки; 3 -
внутренний сегмент палочки; 4 - диски; 5 - клеточная оболочка; 6 -
двойные микрофибриллы; 7 - митохондрии; 8 - пузырьки эндоплазматической
сети; 9 - ядро; 10 - область синапса с биполярным нейроцитом; 11 -
пальцевидные отростки внутреннего сегмента колбочковидной зрительной
клетки; 12 - лучевой глиоцит (мюллерово волокно) (по И.В. Алмазову и
Л.С. Сутулову)
широкой ножкой, образующей синапсы с многочисленными дендритами биполярных нейронов и горизонтальных клеток.
Зрительный пигмент, располагающийся в мембранах наружного сегмента палочек и колбочек, под действием света изменяется, что приводит к возникновению импульса. В сетчатке глаза человека содержатся один тип палочек и три типа колбочек, каждый из которых воспринимает свет определенной длины волны. Глаз человека способен воспринимать волны длиной от 400 до 700 нм. Родопсин лучше всего воспринимает волны длиной около 510 нм (зеленая часть спектра), колбочки - около 430 нм (синяя часть спектра), 530 (зеленая) и 560 (красная часть спектра). Каждый рецептор воспринимает не только свет указанной длины волны, на волны этой длины он реагирует лучше.
Количество колбочек в сетчатке глаза человека достигает 6-7 млн, палочек в 10-20 раз больше. В области желтого пятна имеются лишь колбочковидные клетки, причем они уже и длиннее, чем на остальном протяжении сетчатки. Палочковидные зрительные клетки воспринимают слабый свет, колбочки активны при ярком свете. Цветовое зрение связано с функционированием колбочек разного типа.
Короткие отростки (аксоны) зрительных (фотосенсорных) клеток образуют наружный сетчатый слой (пятый), в котором они контактируют с ассоциативными биполярными нейронами, расположенными во внутреннем ядерном слое сетчатки (шестой слой). К ассоциативным нейронам относятся клетки нескольких разновидностей: биполярные, горизонтальные и амакриновые. Аксоны фотосенсорных клеток образуют синапсы с дендритами биполярных и горизонтальных клеток. Амакриновые нейроны, имеющие только дендриты, образуют синапсы с биполярными и ганглиозными клетками во внутреннем сетчатом слое (седьмой слой).
Ганглиозные невроциты, образующие ганглиозный слой (восьмой), по строению сходны с другими чувствительными нейронами. В их крупном перикарионе расположены элементы зернистой эндоплазматической сети (субстанции Ниссля), митохондрии, вторичные лизосомы, имеется развитый сетчатый аппарат (комплекс Гольджи). Немиелинизированные аксоны ганглиозных невроцитов (500 тыс - 1 млн) образуют слой нервных волокон (девятый слой), формирующих зрительный нерв. Десятый слой представляет собой внутренний пограничный слой сетчатки.
Следует обратить внимание на две важные закономерности: световая волна достигает колбочек и палочек лишь после того, как пройдет почти всю толщину сетчатки. Каждая ассоциативная клетка получает импульсы от нескольких фотосенсорных, каждый ганглиозный нейроцит - от нескольких ассоциативных клеток.
Сетчатка является нервной тканью. Помимо нейронов, в ней имеются клетки глии, радиальные глиоциты (мюллеровы клетки). Они расположены параллельно фотосенсорным клеткам на участке от внутренней пограничной мембраны до внутренних сегментов палочек и колбочек и перпендикулярно поверхности сетчатки. Глиоциты выполняют трофическую и поддерживающую функции. Лентовидные отростки глиоцитов окружают тела и отростки фотосенсорных клеток, биполярных и ганглиозных нейроцитов, образуя с ними лентовидные синаптические комплексы. От наружной поверхности глиоцитов отходят многочисленные тонкие микроворсинки, внедряющиеся между палочками и колбочками. Концы отростков глиоцитов и их базальная мембрана формируют тонкую внутреннюю пограничную мембрану (десятый слой), которая отделяет стекловидное тело от аксонов ганглиозных нейроцитов и отростков мюллеровых клеток.
Хрусталик (lens) представляет собой прозрачную двояковыпуклую линзу диаметром около 9 мм, имеющую переднюю и заднюю поверхности, которые переходят одна в другую в области экватора хрусталика. Линия, соединяющая наиболее выпуклые точки обеих поверхностей (полюсы), называется осью хрусталика (axis lentis). Ее размеры колеблются от 3,7 до 4,4 мм в зависимости от степени аккомодации. Коэффициент преломления хрусталика в поверхностных слоях равен 1,32, в центральных - 1,42. Хрусталик покрыт прозрачной капсулой ( capsula lentis) - гомогенной базальной мембраной толщиной около 10 нм на передней поверхности и 3-4 нм на задней поверхности хрусталика. Капсула хрусталика содержит множество ретикулярных волокон с типичной периодической исчерченностью. Под капсулой передняя поверхность хрусталика до его экватора образована эпителием (хрусталиковыми волокнами). Вблизи центра хрусталика эпителиоциты цилиндрические, по направлению к экватору их высота уменьшается. Вблизи экватора эпителиоциты плоские. Ядро хрусталика образовано прозрачными хрусталиковыми волокнами, состоящими большей частью из белка кристаллина. Эти волокна дифференцируются в эмбриональный период из эпителиальных клеток, покрывающих
заднюю поверхность образующегося хрусталика, и сохраняются в течение всей жизни человека. Хрусталиковые волокна представляют собой длинные шестигранные призмы, соединяющиеся между собой с помощью коротких отростков. Волокна заполнены аморфным умеренно осмиофильным материалом. Хрусталик не содержит сосудов и нервных волокон, его трофика осуществляется путем диффузии из водянистой влаги.
Хрусталик как бы подвешен на ресничном пояске (zonula ciliaris - циннова связка), между волокнами которого расположены пространства пояска (spatium zonulare - петитов канал). Этот канал сообщается с задней камерой глаза. Волокна цинновой связки передают хрусталику движения ресничной мышцы. При сокращении ресничной мышцы собственно сосудистая оболочка смещается вперед, ресничное тело приближается к экватору хрусталика, ресничный поясок ослабевает, хрусталик становится более выпуклым, его светопреломляющая способность возрастает. При расслаблении ресничной мышцы ресничное тело удаляется от экватора хрусталика, ресничный поясок натягивается, хрусталик уплощается. Его преломляющая способность уменьшается (рис. 124).
Стекловидное тело (corpus vitreum) заполняет пространство между сетчаткой сзади, хрусталиком и задней стороной ресничного пояска спереди. Стекловидное тело представляет собой аморфное межклеточное вещество желеобразной консистенции, индекс светопреломления равен 1,334. Стекловидное тело состоит из гигроскопичного белка витреина и гиалуроновой кислоты. На передней поверхности стекловидного тела имеется ямка, в которой располагается хрусталик.
Камеры глаза. Радужка разделяет пространство, расположенное между роговицей спереди и хрусталиком с цинновой связкой и ресничным телом сзади, на две камеры - переднюю и заднюю, которые играют важную роль в циркуляции водянистой влаги внутри глаза.
Передняя камера глазного яблока (camera anterior bulbi) находится между роговицей спереди и радужкой сзади. Задняя камера глазного яблока (camera posterior bulbi) расположена позади радужки. Задней стенкой этой камеры является передняя поверхность хрусталика и ресничного пояска. Обе камеры сообщаются друг с другом через зрачок. В обеих камерах глазного яблока находится водянистая влага (humor aquosus), представляющая собой жидкость с очень низкой вязкостью. Она содержит около 0,02% белка и наибольшое количество деполимеризованной
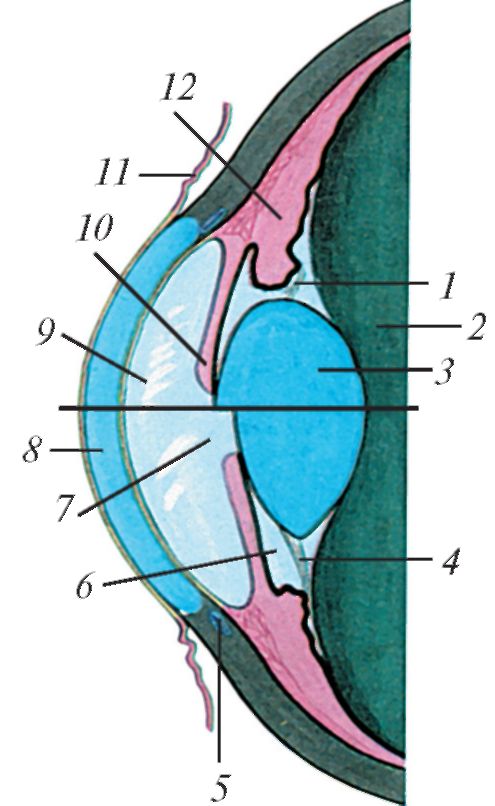 Рис. 124. Изменение формы хрусталика при натяжении и расслаблении ресничной мышцы (схема):
Рис. 124. Изменение формы хрусталика при натяжении и расслаблении ресничной мышцы (схема):
1 - ресничный поясок (расслаблен);
2 - стекловидное тело; 3 - хрусталик;
4 - ресничный поясок (натянут);
5 - венозный синус склеры; 6 - задняя камера глазного яблока; 7 - зрачок; 8 - роговица; 9 - передняя камера глазного яблока; 10 - радужка; 11 - конъюнктива; 12 - ресничное тело
гиалуроновой кислоты. Водянистая влага лишена фибриногена, поэтому она не свертывается. В физиологических условиях водянистая влага находится под давлением около 20-25 мм рт. ст. (внутриглазное давление). Постоянство этого давления зависит от равновесия между образованием и обратным всасыванием водянистой влаги внутри глазного яблока. У здорового человека этот процесс протекает со скоростью около 2 мм3/мин.
Водянистая влага вырабатывается капиллярами ресничных отростков и заднего отдела радужки и поступает в пространства пояска, представляющие собой круговую щель, которая расположена вокруг хрусталика между волокнами поясничного пояска, и в заднюю камеру глаза. Из задней камеры через зрачок водянистая влага оттекает в переднюю камеру. В углу передней камеры, образованном краем радужки и роговицы (радужно-роговичный угол - angulus iridocornealis), по окружности располагается зубчатая связка, между пучками волокон которой имеются выстланные эндотелием щели (фонтаново пространство). Из щелей фонтановых пространств влага проходит в просвет венозного синуса склеры (sinus venosus sclerae - шлеммов канал). Из этого венозного синуса водянистая влага направляется в собирательные сосуды, расположенные в склере, которые выходят под конъюнктиву (водоворотные вены), где вливаются в вены глаза.
ВСПОМОГАТЕЛЬНЫЕ ОРГАНЫ ГЛАЗА
Вспомогательными органами глаза являются мышцы глазного яблока, слезный аппарат, конъюнктива, веки.
Полость глазницы, в которой располагаются глазное яблоко и его вспомогательные органы, выстлана надкостницей глазницы, которая в области зрительного канала и верхней глазничной щели срастается с твердой оболочкой головного мозга. Глазное яблоко окутано его соединительнотканным влагалищем (vagina bulbi - тенонова капсула), которое соединяется со склерой рыхлой соединительной тканью. На задней поверхности глазного яблока влагалище сращено с наружным влагалищем зрительного нерва, спереди оно подходит к своду конъюнктивы. Сосуды, нервы и сухожилия глазодвигательных мышц прободают влагалище глазного яблока. Между глазным яблоком и его влагалищем находится узкое эписклеральное (теноново) пространство (spatium episclerale).
Между надкостницей глазницы и влагалищем глазного яблока залегает жировое тело глазницы (corpus adiposum orbitae). Спереди глазница (и ее содержимое) частично закрыта глазничной перегородкой (septum orbitale), начинающейся от надкостницы верхнего и нижнего краев глазницы и прикрепляющейся к хрящам верхнего и нижнего века. В области внутреннего угла глаза глазная перегородка соединяется с медиальной связкой века.
Веки (palpebrae) защищают глазное яблоко спереди. Они представляют собой кожные складки, ограничивающие глазную щель и закрывающие ее при смыкании век (рис. 125). По бокам веки соединены латеральной и медиальной спайками, замыкающими соответствующие углы глаза. Латеральный угол глаза (angulus oculi lateralis) острый, а медиальный угол (angulus oculi medialis) закругленный. Благодаря этому в области медиального угла имеется выемка - слезное озеро (lacus lacrimalis). Сверху верхнее веко ограничено бровью (supercilium) с короткими жесткими волосами. Нижнее веко при открывании глаз слегка опускается под действием силы тяжести. К верхнему веку подходит мышца, поднимающая верхнее веко (m. levator palpebrae), которая начинается вместе с прямыми мышцами от общего сухожильного кольца. Мышца проходит в верхней части глазницы и прикрепляется к верхнему хрящу века (tarsus superior) - пластинке плотной волокнистой соединительной ткани, выполняющей опорную функцию. В толще нижнего века имеется
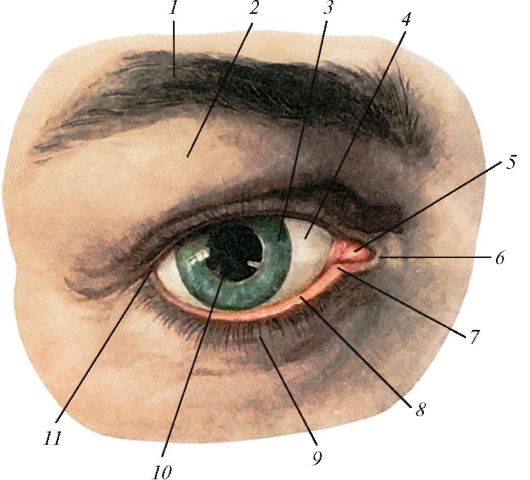 Рис. 125. Верхние
и нижние веки правого глаза, вид спереди: 1 - бровь; 2 - верхнее веко; 3
- радужка; 4 - фиброзная оболочка глазного яблока; 5 - слезное мясцо; 6
- медиальная спайка век; 7 - слезная точка; 8 - нижнее
Рис. 125. Верхние
и нижние веки правого глаза, вид спереди: 1 - бровь; 2 - верхнее веко; 3
- радужка; 4 - фиброзная оболочка глазного яблока; 5 - слезное мясцо; 6
- медиальная спайка век; 7 - слезная точка; 8 - нижнее
веко; 9 - ресницы; 10 - зрачок; 11 - латеральная спайка век
аналогичный хрящ нижнего века (tdrsus infdrior). В толще хрящей заложены открывающиеся по краям век разветвленные хрящевые железы (gldndulae tarsdles) - мейбомиевы железы. Ближе к передней поверхности в толще век залегает вековая часть круговой мышцы глаза. По краям век в 2-3 ряда располагаются ресницы (cdilia). В их волосяные сумки открываются выводные протоки сальных желез (gldandulae sebacdeae).
Выпуклая передняя поверхность век покрыта тонкой кожей с короткими пушковыми волосками. Вогнутая задняя поверхность век покрыта конъюнктивой.
Конъюнктива (tdnica conjunctiva) - тонкая соединительнотканная бледно-розовая оболочка, в которой выделяют конъюнктиву век, покрывающую изнутри веки, и конъюнктиву глазного яблока (рис. 126). В месте перехода одной части конъюнктивы в другую образуются верхний и
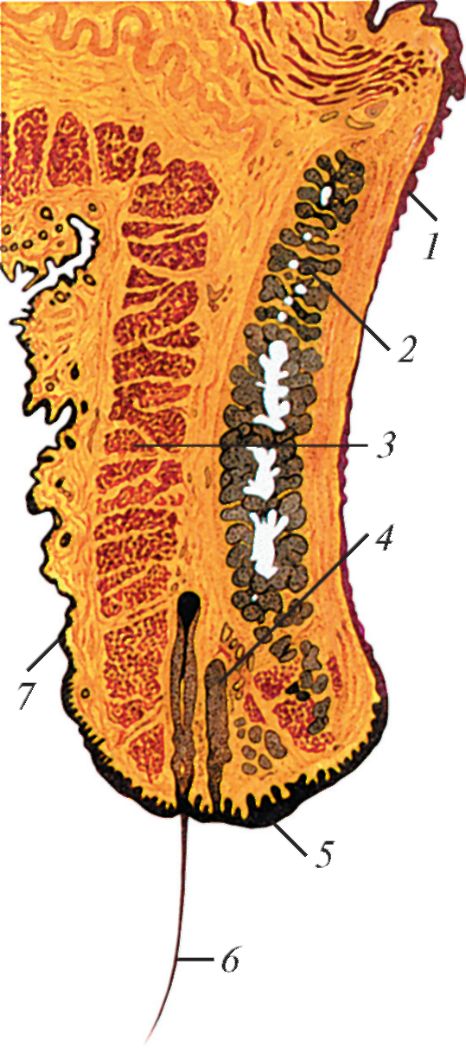 Рис. 126. Строение
века. Фронтальный разрез: 1 - конъюнктива; 2 - хрящ века; 3 - вековая
часть круговой мышцы глаза; 4 - ресничная железа; 5 - край века; 6 -
ресница; 7 - кожа
Рис. 126. Строение
века. Фронтальный разрез: 1 - конъюнктива; 2 - хрящ века; 3 - вековая
часть круговой мышцы глаза; 4 - ресничная железа; 5 - край века; 6 -
ресница; 7 - кожа
нижний своды конъюнктивы (fornix conjunctivae superior et fornix conjunctivae inferior). Пространство, расположенное спереди от глазного яблока и ограниченное конъюнктивой, образует конъюнктивальный мешок (sdccus conjunctivdlis), который при смыкании век закрывается. Закругленный медиальный угол глаза с медиальной стороны ограничивает слезное озеро. У медиального угла глаза находится небольшое возвышение - слезное мясцо (cardncula lacrimalis). Латеральнее от слезного мясца расположена полулунная складка конъюнктивы (plica semilunaris conjunctivae) - рудимент мигательного (третьего) века, имеющегося у позвоночных.
Конъюнктива выстлана трехслойным неороговевающим эпителием, лежащим на базальной мембране. По направлению к краю века эпителий становится многослойным плоским. В эпителии конъюнктивы залегают бокаловидные гландулоциты. Собственная пластинка конъюнктивы образована рыхлой соединительной тканью, в которой находятся фибробласты, макрофаги, лаброциты, плазматические клетки, единичные меланоциты и лимфоциты. Конъюнктивальный мешок смачивается слезной жидкостью, которую секретирует слезная железа.
Слезный аппарат (apparatus lacrimalis) включает слезную железу и систему слезных путей (рис. 127).
Слезная железа (glandula lacrimalis), состоящая из нескольких альвеолярно-трубчатых серозных желез, расположена в ямке слезной железы лобной кости в верхнелатеральной части глазницы. Сухожилие мышцы, поднимающей верхнее веко, разделяет железу на две части: большую верхнюю орбитальную часть и меньшую нижнюю вековую, лежащую возле верхнего свода конъюнктивы. В своде конъюнктивы иногда встречаются
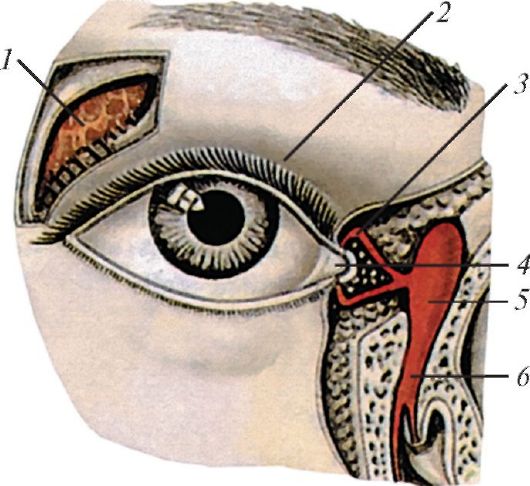 Рис. 127. Слезный
аппарат правого глаза, вид спереди: 1 - слезная железа; 2 - верхнее
веко; 3 - слезный каналец; 4 - слезное озеро; 5 - слезный мешок; 6 -
носослезный проток
Рис. 127. Слезный
аппарат правого глаза, вид спереди: 1 - слезная железа; 2 - верхнее
веко; 3 - слезный каналец; 4 - слезное озеро; 5 - слезный мешок; 6 -
носослезный проток
мелких размеров добавочные слезные железы. От 5 до 12 выводных канальцев слезной железы открываются в верхний свод конъюнктивы. Слеза омывает переднюю часть глазного яблока и по слезному ручью (rivus lacrimalis) - капиллярной щели, расположенной возле краев век, оттекает в слезное озеро (lacus lacrimalis), находящееся в медиальном углу глаза. У медиального угла глаза, на краях век, там, где они сходятся, окружая слезное озеро, расположены верхний и нижний слезные сосочки (papillae lacrimales). На вершине этих сосочков имеется по узкому отверстию - слезной точке (punctum lacrimalis). От слезной точки берет начало узкий слезный каналец (canaliculus lacrimalis) длиной около 1 см и диаметром около 0,5 мм. Верхний и нижний канальцы впадают в слезный мешок (saccus lacrimalis), который обращен слепым концом вверх. Нижний конец мешка переходит в носослезный проток (ductus nasolacrimalis), открывающийся в нижний носовой ход. Слезная часть круговой мышцы глаза, сращенная со стенкой слезного мешка, сокращаясь, расширяет его. Благодаря этому слеза всасывается в слезный мешок через слезные канальцы.
Мышцы глазного яблока. Глазное яблоко у человека может поворачиваться так, чтобы на рассматриваемом предмете сходились зрительные оси обоих глазных яблок. Движения глазных яблок осуществляют
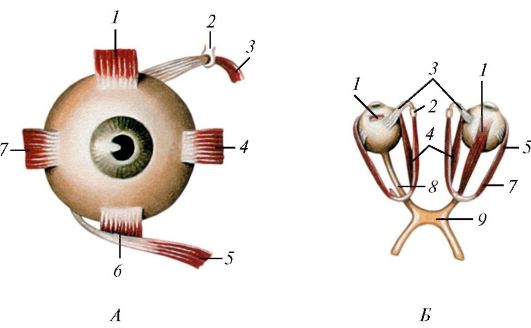 Рис. 128. Мышцы
глазного яблока (глазодвигательные мышцы), вид спереди (А) и сверху
(Б): 1 - верхняя прямая мышца; 2 - блок; 3 - верхняя косая мышца; 4 -
медиальная прямая мышца; 5 - нижняя косая мышца; 6 - нижняя прямая
мышца; 7 - латеральная прямая мышца; 8 - зрительный нерв; 9 - перекрест
зрительных нервов
Рис. 128. Мышцы
глазного яблока (глазодвигательные мышцы), вид спереди (А) и сверху
(Б): 1 - верхняя прямая мышца; 2 - блок; 3 - верхняя косая мышца; 4 -
медиальная прямая мышца; 5 - нижняя косая мышца; 6 - нижняя прямая
мышца; 7 - латеральная прямая мышца; 8 - зрительный нерв; 9 - перекрест
зрительных нервов
шесть поперечнополосатых глазодвигательных мышц: четыре прямые (верхняя, нижняя, медиальная, латеральная - musculi recti superior, inferior, medialis, lateralis) и две косые (верхняя и нижняя - musculi obliqui superior et inferior) (рис. 128, рис. 129). Нижняя косая мышца глаза начинается на нижней стенке глазницы возле отверстия носослезного канала. Остальные начинаются в глубине глазницы в окружности зрительного канала и прилегающей части верхней глазничной щели от общего сухожильного кольца (annulus tendineus communis), окружающего зрительный нерв и глазную артерию. Кольцо фиксировано к клиновидной кости, надкостнице вокруг зрительного канала и частично к краям верхней глазничной щели. Все прямые мышцы направляются вдоль соответствующих стенок глазниц, по сторонам от зрительного нерва, прободают влагалище глазного яблока и прикрепляются к склере впереди экватора в различных участках соответственно названиям.
Верхняя косая мышца глаза лежит в верхне-медиальной части глазницы между верхней и медиальной прямыми мышцами. Вблизи блоковой ямки глазницы она переходит в окутанное синовиальным влагалищем тонкое круглое сухожилие. Это сухожилие перекидывается через блок (trochlea) в верхне-медиальном углу глазницы, поворачивает кзади и вбок
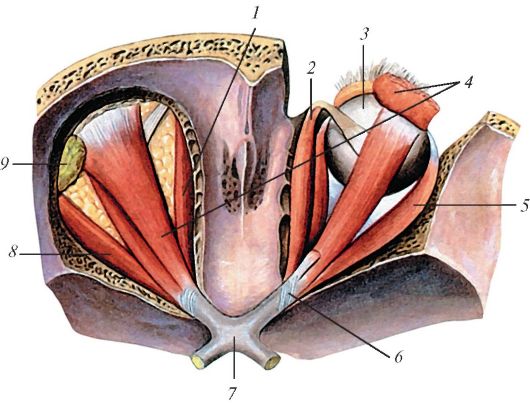 Рис. 129. Верхняя
косая и другие мышцы глазного яблока, вид сверху. На правой стороне
рисунка мышца, поднимающая верхнее веко, разрезана и частично удалена.
Верхняя стенка правой и левой глазниц удалена: 1 - медиальная прямая
мышца; 2 - блок (верхней косой мышцы); 3 - глазное яблоко; 4 - мышца,
поднимающая верхнее веко; 5 - латеральная прямая мышца; 6 - общее
сухожильное кольцо; 7 - зрительный перекрест; 8 - латеральная прямая
мышца; 9 - слезная железа
Рис. 129. Верхняя
косая и другие мышцы глазного яблока, вид сверху. На правой стороне
рисунка мышца, поднимающая верхнее веко, разрезана и частично удалена.
Верхняя стенка правой и левой глазниц удалена: 1 - медиальная прямая
мышца; 2 - блок (верхней косой мышцы); 3 - глазное яблоко; 4 - мышца,
поднимающая верхнее веко; 5 - латеральная прямая мышца; 6 - общее
сухожильное кольцо; 7 - зрительный перекрест; 8 - латеральная прямая
мышца; 9 - слезная железа
и прикрепляется к склере позади экватора глаза на верхне-латеральной поверхности глазного яблока. Нижняя косая мышца прикрепляется к глазному яблоку сбоку также позади экватора.
Прямые мышцы поворачивают глазное яблоко в соответствующем направлении вокруг двух взаимно пересекающихся осей: вертикальной и горизонтальной (поперечной). Латеральная и медиальная прямые мышцы поворачивают глазное яблоко кнаружи или кнутри вокруг вертикальной оси, каждая в свою сторону. Соответственно поворачивается и зрачок. Верхняя и нижняя прямые мышцы поворачивают глазное яблоко вокруг поперечной оси вверх или вниз. Косые мышцы поворачивают глазное яблоко вокруг сагиттальной оси: верхняя - вниз и кнаружи, нижняя - вверх и кнаружи. Благодаря содружественному действию указанных мышц движения обоих глазных яблок согласованы.
Кровоснабжение и иннервация органа зрения. Глазное яблоко и его вспомогательные органы получают кровь из ветвей глазной артерии (ветви внутренней сонной артерии). Одна из ветвей глазной артерии (центральная артерия сетчатки) кровоснабжает сетчатку и часть зрительного нерва, другая - склеру и сосудистую оболочку (рис. 130). Центральная артерия сетчатки проникает в толще зрительного нерва внутрь глазного яблока, в области диска отдает верхние и нижние ветви, которые распространяются в пределах внутреннего зернистого и ганглионарного слоев сетчатки. Капилляры, окруженные отростками глиальных и мюллеровых клеток, образованы фенестрированным эндотелием.
В сосудистой оболочке глазного яблока ветвятся короткие и длинные задние и передние ресничные артерии. Передние ресничные артерии в толще радужки анастомозируют между собой и образуют два артериальных круга: большой, расположенный у ресничного края радужки, и малый, прилежащий к ее зрачковому краю. Склера кровоснабжается задними короткими ресничными артериями. Из густой венозной сети собственно сосудистой оболочки формируются 4-6 вортикозных вен, которые прободают склеру и впадают в глазные вены. В передние ресничные вены оттекает кровь из ресничного тела, радужки и склеры. Веки и конъюнктива кровоснабжаются из медиальной и латеральной артерий, которые, анастомозируя, образуют в толще век дуги передних и задних конъюнктивных артерий. Одноименные вены впадают в глазную и лицевую вены. Слезная железа получает кровь из одноименной артерии.
Мышцы, фасции и жировое тело глазницы также кровоснабжаются ветвями глазной артерии.
Лимфатические сосуды от век и конъюнктивы направляются к поднижнечелюстным, а также к поверхностным и глубоким околоушным (предушным) лимфатическим узлам.
Иннервацию чувствительную содержимое глазницы получает из глазного нерва (первая ветвь тройничного нерва). От его ветви - носоресничного нерва к глазному яблоку отходят длинные ресничные нервы. Нижнее веко иннервируется подглазничным нервом (ветвь верхнечелюстного нерва). Сфинктер зрачка и ресничная мышца иннервируются парасимпатическими волокнами глазодвигательного нерва (от ресничного узла в составе коротких ресничных нервов). Дилататор зрачка получает иннервацию по симпатическим волокнам внутреннего сонного сплетения, которые подходят к глазному яблоку вместе с кровеносными сосудами. Верхняя, нижняя и
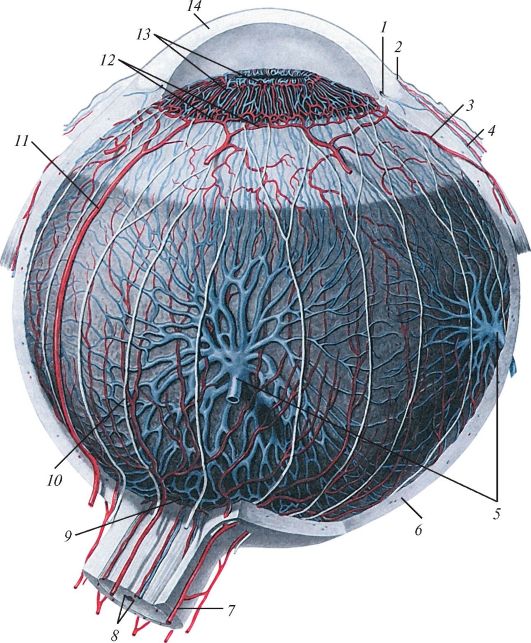 Рис. 130. Кровеносные сосуды и нервы глазного яблока:
Рис. 130. Кровеносные сосуды и нервы глазного яблока:
1 - венозный синус склеры; 2 - циркулярная борозда склеры; 3 - передняя ресничная артерия; 4 - конъюнктивальные сосуды; 5 - водоворотная вена (вена собственной сосудистой оболочки глаза); 6 - склера (белочная оболочка); 7 - короткие задние ресничные артерии; 8 - центральная артерия и вены сетчатки; 9 - артериальный круг зрительного нерва; 10 - длинные ресничные нервы;
11 - длинная задняя ресничная артерия; 12 - большой артериальный круг радужки;
13 - малый артериальный круг радужки; 14 - роговица
медиальная прямые, нижняя косая мышцы глаза и мышца, поднимающая верхнее веко, иннервируются ветвями глазодвигательного нерва, латеральная прямая - отводящим, верхняя косая - блоковым нервами.
Проводящий путь зрительного анализатора. Световой луч проходит через роговицу, водянистую влагу передней камеры, зрачок, который в зависимости от интенсивности света то расширяется, то суживается, водянистую влагу задней камеры, хрусталик, стекловидное тело и попадает на сетчатку. Пучок света благодаря светопреломляющим средам направляется на желтое пятно сетчатки - зону наилучшего видения. Важная роль в этом принадлежит хрусталику, который с помощью ресничной мышцы может увеличивать или уменьшать кривизну при аккомодации. Глазодвигательные мышцы направляют глазные яблоки в сторону рассматриваемого объекта, устанавливают оси обоих глаз параллельно при взгляде вдаль или сближают их при рассматривании предмета на близком расстоянии.
При попадании света на палочки и колбочки - отростки первых нейронов зрительного пути - в них генерируется нервный импульс, который передается биполярным нейроцитам, а от них - ганглиозным нейроцитам (рис. 131). Аксоны ганглиозных клеток формируют зрительный
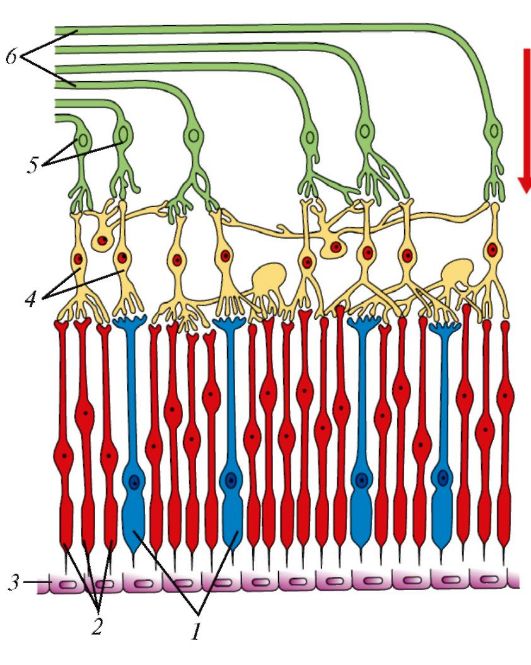 Рис. 131. Расположение нейронов в сетчатке глаза
Рис. 131. Расположение нейронов в сетчатке глаза
(схема): 1 - колбочки; 2 - палочки;
3 - пигментные клетки;
4 - биполярные клетки;
5 - ганглиозные клетки;
6 - нервные волокна. Стрелкой показано направление пучка света
нерв, который выходит из глазницы через канал зрительного нерва. На нижней поверхности мозга зрительные нервы образуют перекрест, но перекрещиваются лишь волокна, идущие от медиальной части сетчатки каждого глаза. В каждом зрительном тракте проходят волокна, несущие нервные импульсы от клеток медиальной половины сетчатки противоположного глаза и латеральной половины глаза своей стороны. Часть волокон зрительного тракта направляется в латеральное коленчатое тело, где они заканчиваются синапсами на залегающих здесь нейронах. Другая часть аксонов ганглиозных нейронов, не доходя до латерального коленчатого тела, направляется через ручки верхних холмиков четверохолмия в их ядра. Из верхних холмиков нервные импульсы следуют в ядра глазодвигательного нерва (двигательное и добавочное вегетативное), иннервирующие мышцы глаза, мышцу, суживающую зрачок, и ресничную мышцу. Таким образом, в ответ на попадание световых волн в глаз зрачок суживается, а глазные яблоки поворачиваются в направлении пучка света.
Аксоны нейронов латерального коленчатого тела направляются к клеткам зрительной коры, расположенной в затылочной доле полушария большого мозга, возле шпорной борозды (поле 17) (рис. 132).
РАЗВИТИЕ ОРГАНА ЗРЕНИЯ У ЧЕЛОВЕКА В ОНТОГЕНЕЗЕ
У человека (и у млекопитающих) части глаза развиваются из различных источников (рис. 133). Светочувствительная оболочка (сетчатка) происходит из боковой стенки мозгового пузыря (будущего промежуточного мозга), хрусталик развивается непосредственно из эктодермы, сосудистая и фиброзная оболочки - из мезенхимы. В конце 1-го месяца развития зародыша человека боковые стенки переднего мозгового пузыря выпячиваются, образуя первичные пузырьки. Ножки этих пузырьков - глазные стебельки - превращаются в зрительные нервы, а передняя часть пузырька впячивается в его полость. Таким образом формируется двухслойный бокал, который преобразуется в сетчатку. Из внутреннего слоя глазного бокала образуется внутренний светочувствительный (нервный) слой сетчатки, из наружного - наружный пигментный слой. Хрусталик формируется из утолщения эктодермы, расположенной впереди глазного пузырька. Вскоре будущий хрусталик отшнуровывается от эктодермы. На 2-м месяце
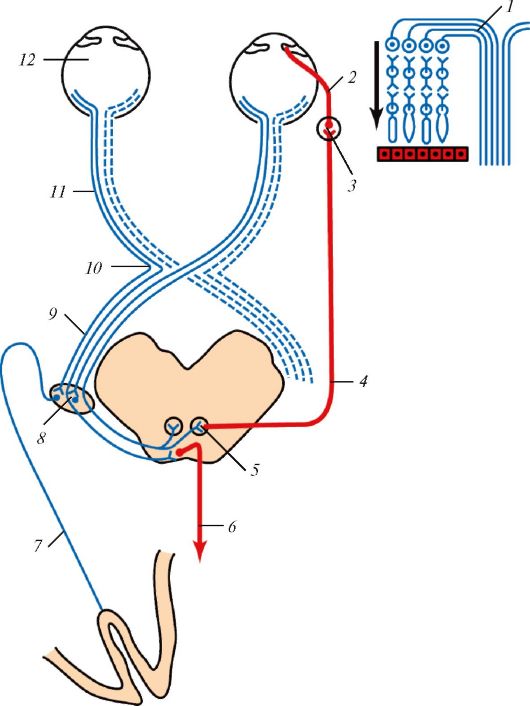 Рис. 132. Проводящий
путь зрительного анализатора: 1 - схема строения сетчатки и
формирования зрительного нерва (стрелка показывает направление света в
сетчатке); 2 - короткие ресничные нервы; 3 - ресничный узел; 4 -
глазодвигательный нерв; 5 - ядро глазодвигательного нерва; 6 -
покрышечно-спинномозговой путь; 7 - зрительная лучистость; 8 -
латеральное коленчатое тело; 9 - зрительный тракт; 10 - зрительный
перекрест; 11 - зрительный нерв; 12 - глазное яблоко
Рис. 132. Проводящий
путь зрительного анализатора: 1 - схема строения сетчатки и
формирования зрительного нерва (стрелка показывает направление света в
сетчатке); 2 - короткие ресничные нервы; 3 - ресничный узел; 4 -
глазодвигательный нерв; 5 - ядро глазодвигательного нерва; 6 -
покрышечно-спинномозговой путь; 7 - зрительная лучистость; 8 -
латеральное коленчатое тело; 9 - зрительный тракт; 10 - зрительный
перекрест; 11 - зрительный нерв; 12 - глазное яблоко
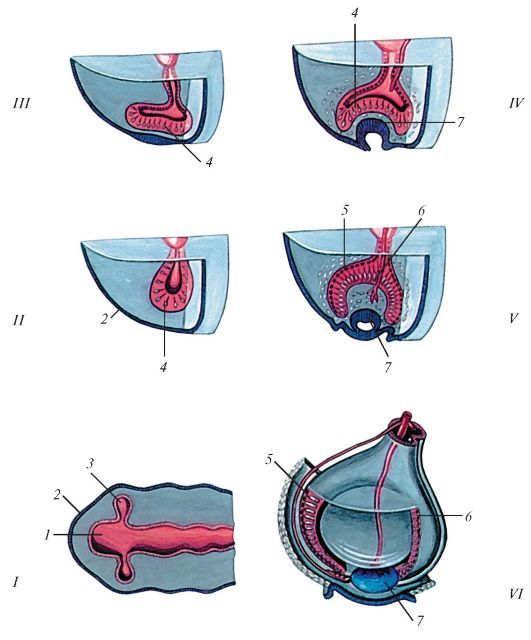 Рис. 133. Последовательные стадии развития (формирования) глазного яблока
Рис. 133. Последовательные стадии развития (формирования) глазного яблока
(схема):
1 - мозговой пузырь; 2 - эктодерма; 3 - глазной пузырь; 4 - формирование сетчатки; 5 - пигментная часть сетчатки; 6 - нервная часть сетчатки; 7 - формирующийся хрусталик
развития из мезенхимы, окружающей глазной бокал, начинают формироваться сосудистая оболочка, склера, роговица и стекловидное тело. В состав глазного стебелька входит также артерия, которая кровоснабжает эпителиальные элементы формирующегося глаза. Впоследствии эта артерия атрофируется. В течение 6-8 месяцев внутриутробного развития исчезают сосуды стекловидного тела и капсулы хрусталика, рассасывается зрачковая мембрана, закрывающая отверстие зрачка. Складки эктодермы, образующие веки, начинают формироваться на 3-м месяце развития. Из эктодермы происходит и эпителий конъюнктивы. Из выростов этого эпителия развивается слезная железа.
Глазное яблоко новорожденного относительно большое. Масса глазного яблока составляет в среднем 2,2 г, объем 3,25 см3, продольный диаметр 17,3 мм, поперечный - 16,7 мм. До двухлетнего возраста глазное яблоко увеличивается на 40%, по сравнению с его величиной у новорожденного, в 5 лет - на 70%, у взрослого - в 3 раза. В глазнице ребенка глазное яблоко расположено более поверхностно и латерально, а у взрослого оно лежит глубже. Это изменение положения связано с развитием носа и жировой клетчатки в глазнице. Широкая и толстая роговица у новорожденного, резко отграниченная от склеры, имеет сильно выраженную кривизну, которая практически не меняется после рождения. Толщина роговицы составляет 3-4 мм, диаметр по экватору 6-7 мм, радиус передней поверхности хрусталика равен 5 мм, задней - 4 мм. Хрусталик быстро растет в течение первого года жизни, после чего рост замедляется. У новорожденного радужка тонкая, бедная пигментом. По мере роста ребенка радужка утолщается, в ней увеличивается количество пигмента. Слабо развитое ресничное тело новорожденного быстро растет. Относительно тонкая сетчатка развита хорошо.
Зрительный нерв тонкий и короткий, он удлиняется по мере роста глазного яблока. К моменту рождения мышцы глазного яблока развиты хорошо. Относительно большие слезные железы новорожденного практически не функционируют. Слезоотделение начинается лишь в течение 2-го месяца жизни. Глазная щель новорожденного узкая, медиальный угол глаза закруглен. В дальнейшем глазная щель быстро увеличивается. У детей до 14-15 лет глазная щель широкая, поэтому глаз кажется большим, чем у взрослого человека.
АНОМАЛИИ РАЗВИТИЯ ГЛАЗНОГО ЯБЛОКА
Сложное эмбриональное развитие глазного яблока в ряде случаев обусловливает появление врожденных дефектов. Чаще всего встречается неправильная кривизна роговицы или хрусталика, из-за чего изображение на сетчатке искажается (астигматизм). При нарушенных пропорциях глазного яблока возникает врожденная близорукость (если зрительная ось удлинена) или дальнозоркость (если зрительная ось укорочена). Встречается дефект (щель) в радужке (колобома), чаще в передне-медиальном сегменте радужки. Остатки ветвей артерии внутри глазного яблока мешают прохождению света в стекловидном теле. Иногда встречается врожденная катаракта - нарушение прозрачности хрусталика. Недоразвитие шлеммова канала или фонтановых пространств вызывает врожденную глаукому. Крайне редко встречается недоразвитие или даже отсутствие век. Иногда глазные яблоки уменьшены в размерах. Редко наблюдается полное или частичное отсутствие радужки, которое обычно сочетается с другими пороками. Примерно у 1 из 150 000-200 000 новорожденных уменьшено содержание пигмента в сосудистой оболочке в сочетании с отсутствием пигмента в коже, волосах и т. д. (альбинизм). Крайне редко отсутствует один или оба глаза.
ОРГАНЫ СЛУХА И РАВНОВЕСИЯ
Органы слуха и равновесия (статического чувства) у человека объединены в систему, морфологически разделенную на три отдела (рис. 134): наружное ухо, включающее наружный слуховой проход и ушную раковину с мышцами и связками; среднее ухо, к которому относятся барабанная полость, сосцевидные придатки и слуховая труба; внутреннее ухо, которое объединяет перепончатый лабиринт, располагающийся в костном лабиринте внутри пирамиды височной кости. Если наружное и среднее ухо принадлежит только органу слуха, то внутреннее ухо включает в себя и внутреннюю часть слухового аппарата, и весь орган равновесия (статического чувства).
Наружное ухо (auris externa) начинается ушной раковиной, которая улавливает направление звуков. Ушная раковина (auricula) представляет собой эластический хрящ сложной формы, покрытый кожей (рис. 135). У человека ушная раковина удлинена, ее вогнутая поверхность обращена вперед и латерально, нижняя часть - долька ушной раковины, или мочка (lobus auriculae), лишена хряща и заполнена жиром. Свободный загнутый край - завиток (helix) ограничивает ушную раковину сзади и сверху. Завиток в передней части раковины над наружным слуховым проходом заканчивается в виде ножки завитка (crus helicis).
Рис. 134. Орган слуха и равновесия и его части: А - орган слуха и равновесия на продольном разрезе; Б - слуховые косточки; В - улитка (улитковые протоки «лестницы» вскрыты; Г - улитковый проток на поперечном разрезе; 1 - ушная раковина; 2 - наружный слуховой проход; 3 - барабанная перепонка; 4 - молоточек; 5 - наковальня; 6 - стремя; 7 - слуховая труба; 8 - улитка; 9 - лестница преддверия; 10 - барабанная лестница; 11 - улитковый проток; 12 - спиральный (кортиев) орган; 13 - покровная мембрана
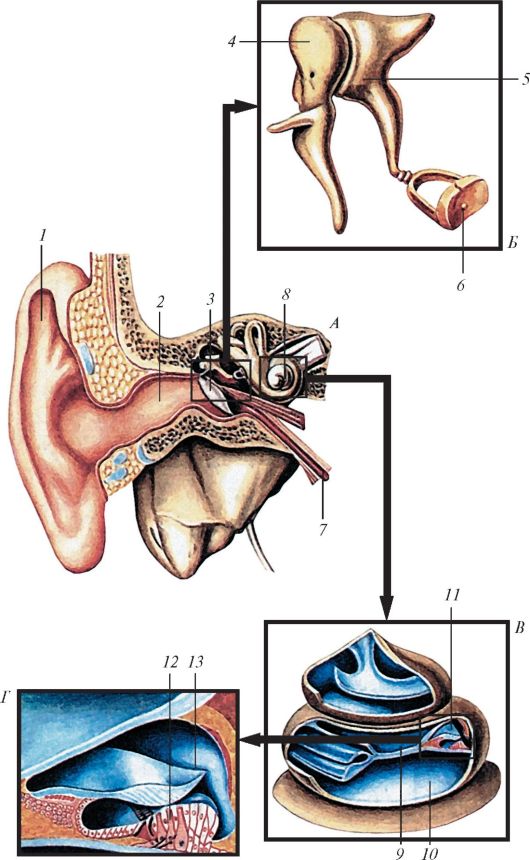
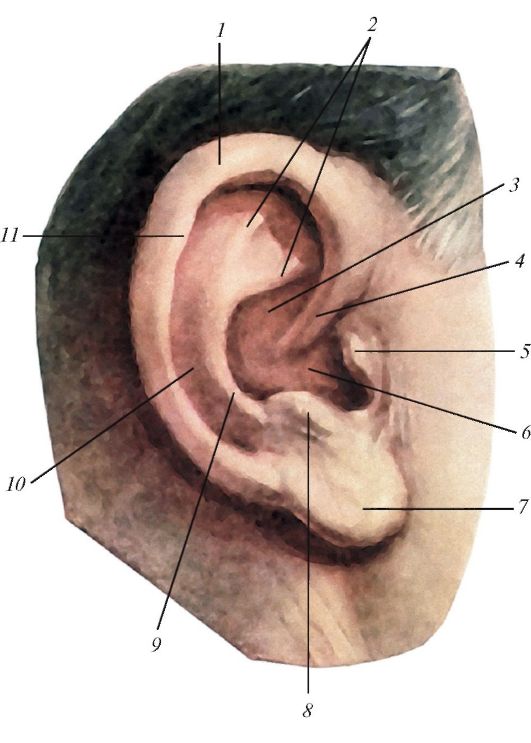 Рис. 135. Ушная
раковина, вид с латеральной стороны: 1 - завиток; 2 - ножки
противозавитка; 3 - челнок раковины; 4 - ножка завитка; 5 - козелок; 6 -
полость раковины; 7 - долька ушной раковины; 8 - противокозелок; 9 -
противозавиток; 10 - ладья; 11 - бугорок ушной раковины
Рис. 135. Ушная
раковина, вид с латеральной стороны: 1 - завиток; 2 - ножки
противозавитка; 3 - челнок раковины; 4 - ножка завитка; 5 - козелок; 6 -
полость раковины; 7 - долька ушной раковины; 8 - противокозелок; 9 -
противозавиток; 10 - ладья; 11 - бугорок ушной раковины
На внутренней стороне завитка, в задне-верхней его части, имеется не всегда четко выраженный выступ - бугорок ушной раковины (tuberculum auriculae; дарвинов бугорок).
На вогнутой поверхности параллельно завитку расположен противозавиток (anthielix). Кпереди от противозавитка имеется углубление - раковина уха (concha auriculae), на дне которой находится наружное слуховое отверстие (porus acusticus externus), ограниченное спереди козелком (triagus). Напротив него, в нижней части противозавитка, виден противокозелок (antitriagus).
Наружный слуховой проход (meatus acusticus externus) состоит из хрящевого и костного отделов, его длина у взрослого человека около 33-35 мм, диаметр просвета колеблется на разных участках от 0,6 до 0,9 см. По ходу наружного слухового прохода имеется S-образный изгиб в горизонтальной и вертикальной плоскостях. Если оттянуть ушную раковину вверх и кзади, проход выпрямляется. В многослойном плоском эпителии, выстилающем наружный слуховой проход, наряду с большим количеством сальных желез, имеются особые трубчатые железы (видоизмененные потовые), вырабатывающие вязкий желтоватый секрет - ушную серу. Протоки желез открываются непосредственно на поверхность кожи, выстилающей слуховой проход, или в выводные протоки сальных желез.
На границе наружного и среднего уха располагается тонкая полупрозрачная барабанная перепонка (membrana tympani), которая отделяет наружное ухо от среднего (рис. 136). Толщина перепонки около 0,1 мм, форма эллиптическая, размеры 9-11 мм. В центре имеется вдавление - пупок барабанной перепонки (umbo membranae tympani), место прикрепления к перепонке молоточка, одной из слуховых косточек. Барабанная перепонка вставлена в борозду барабанной части височной кости. У перепонки различают верхнюю (меньшую) - свободную, ненатянутую часть (pars flaccida) и нижнюю (бóльшую) натянутую часть (pars tensa). Барабанная перепонка расположена косо по отношению к оси слухового прохода. Она образует с горизонтальной плоскостью угол 45-55°, открытый в латеральную сторону. Натянутая часть представляет собой пластинку, состоящую из слоев коллагеновых волокон. Наряду с коллагеновыми имеется небольшое количество эластических волокон. В наружном слое волокна расположены радиально, во внутреннем - циркулярно. Свободная часть не содержит коллагеновых волокон. По периферии коллагеновые волокна образуют фиброзно-хрящевое кольцо, которое соединяет барабанную перепонку со стенкой наружного слухового прохода. Наружная
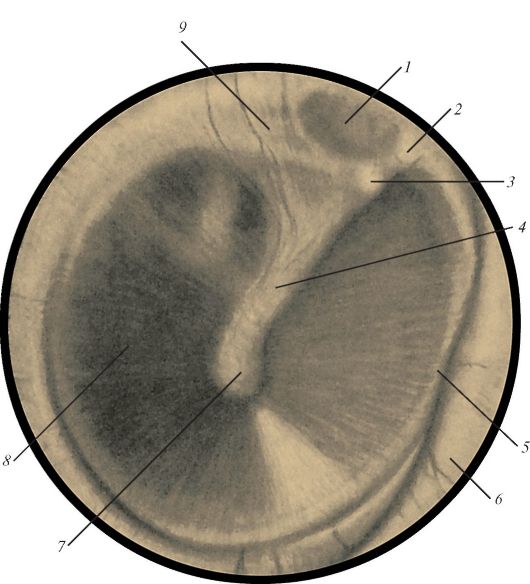 Рис. 136. Барабанная
перепонка: 1 - ненатянутая часть; 2 - передняя молоточковая складка; 3 -
молоточковый выступ; 4 - молоточковая полоска; 5 - волокнисто-хрящевое
кольцо; 6 - наружный слуховой проход; 7 - пупок барабанной перепонки; 8 -
натянутая часть;
Рис. 136. Барабанная
перепонка: 1 - ненатянутая часть; 2 - передняя молоточковая складка; 3 -
молоточковый выступ; 4 - молоточковая полоска; 5 - волокнисто-хрящевое
кольцо; 6 - наружный слуховой проход; 7 - пупок барабанной перепонки; 8 -
натянутая часть;
9 - задняя молоточковая складка
поверхность барабанной перепонки покрыта тонким слоем многослойного плоского эпителия, внутренняя - однослойным кубическим эпителием слизистой оболочкой среднего уха.
Среднее ухо (auris media) представляет собой воздухоносную барабанную полость объемом около 1 см3, которая расположена в основании пирамиды височной кости.
Барабанная полость имеет шесть стенок. Тонкая верхняя покрышечная стенка (paries tegmentalis) отделяет барабанную полость от полости черепа; нижняя яремная стенка (paries jugularis) отделяет барабанную полость от яремной вены, она соответствует нижней стенке пирамиды височной кости там, где располагается яремная ямка, медиальная лабиринтная стенка (paries labyrinthicus) отделяет барабанную полость от костного лабиринта внутреннего уха (рис. 137). В ней имеются разделенные мысом (promontorium) овальное окно преддверия и круглое окно улитки, ведущие в соответствующие отделы костного лабиринта; окно преддверия (fenestra vestibuli) закрыто основанием стремени, оно ведет в преддверие костного лабиринта; окно улитки (fenestra cochlea) закрыто вторичной
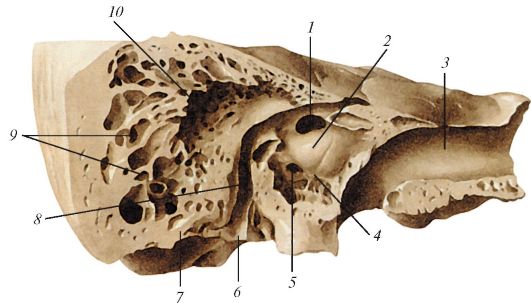 Рис. 137. Лабиринтная
(медиальная) стенка барабанной полости, вид с латеральной стороны, со
стороны барабанной полости. Распил сделан вдоль оси пирамиды: 1 - окно
преддверия; 2 - мыс; 3 - сонный канал; 4 - борозда мыса; 5 - окно
улитки; 6 - шило-сосцевидное отверстие; 7 - сосцевидный отросток; 8 -
канал лицевого нерва; 9 - сосцевидные ячейки; 10 - сосцевидная пещера
Рис. 137. Лабиринтная
(медиальная) стенка барабанной полости, вид с латеральной стороны, со
стороны барабанной полости. Распил сделан вдоль оси пирамиды: 1 - окно
преддверия; 2 - мыс; 3 - сонный канал; 4 - борозда мыса; 5 - окно
улитки; 6 - шило-сосцевидное отверстие; 7 - сосцевидный отросток; 8 -
канал лицевого нерва; 9 - сосцевидные ячейки; 10 - сосцевидная пещера
барабанной перепонкой (membrdana tdympani secunddaria), отделяющей барабанную полость от барабанной лестницы. Над окном преддверия в барабанную полость выступает стенка канала лицевого нерва.
Латеральная перепончатая стенка (paries membranaceus) образована барабанной перепонкой и окружающими ее отделами височной кости; передняя сонная стенка (pdaries cardoticus) отделяет барабанную полость от канала внутренней сонной артерии. В верхней части этой стенки открывается барабанное отверстие слуховой трубы (ostium tympanicum tubae auditivae); в верхней части задней сосцевидной стенки (paries mastoideus) расположен вход в сосцевидную пещеру (antrum mastoideum). Ниже входа в пещеру имеется пирамидальное возвышение (eminentia pyramidalis), внутри которого располагается стременная мышца.
В барабанной полости находятся три слуховые косточки, а также мышцы, натягивающие барабанную перепонку и стремя (обе мышцы поперечнополосатые). Здесь же проходит барабанная струна - ветвь лицевого нерва. Стенки барабанной полости изнутри, а также поверхности слуховых косточек покрыты однослойным плоским эпителием, который местами переходит в кубический или цилиндрический. В эпителиальном покрове, лежащем на тонкой соединительнотканной пластинке, имеются бокаловидные и отдельные реснитчатые клетки.
Барабанная полость кпереди продолжается в узкую слуховую трубу (евстахиеву), которая открывается в носовой части глотки глоточным отверстием слуховой трубы.
Слуховая труба (tuba auditiva) длиной около 3,5 см, диаметр просвета 1-2 мм. Через слуховую трубу происходит выравнивание давления воздуха внутри барабанной полости с давлением в наружной среде. Щелевидное глоточное отверстие слуховой трубы (ostium pharyngeum tubae auditivae) расположено на боковой стенке носовой части глотки. Слуховая труба состоит из костной (1/3) и хрящевой (2/3) частей (рис. 138).
В месте соединения обеих частей расположен наиболее узкий участок - перешеек (isthmus). Верхняя костная часть трубы находится в одноименном полуканале мышечно-трубного канала височной кости и открывается на передней стенке барабанной полости барабанным отверстием слуховой трубы (ostium tympanicum tubae auditivae). Нижняя хрящевая часть, на которую приходится 2/3 длины трубы, имеет вид желоба, открытого снизу, образованного медиальной и латеральной хрящевыми пластинками и соединяющей их перепончатой пластинкой. В том месте, где слуховая труба открывается на боковой стенке носоглотки глоточным отверстием слуховой трубы, медиальная (задняя) пластинка
эластического хряща трубы утолщается и выступает в полость глотки в виде трубного валика (torus tubarius). Продольная ось слуховой трубы от глоточного ее отверстия направлена вверх и латерально, образуя с горизонтальной и сагиттальной плоскостями угол 40-45°.
Складчатая слизистая оболочка слуховой трубы выстлана цилиндрическим многорядным реснитчатым эпителием, богатым бокаловидными гландулоцитами. Эпителий лежит на базальной мембране, под которой расположен подэпителиальный слой рыхлой волокнистой соединительной ткани, богатой лимфоидными элементами и железами. От хрящевой части слуховой трубы берут начало мышца, напрягающая небную занавеску, и мышца, поднимающая небную занавеску. При их сокращении канал слуховой трубы расширяется, и воздух из глотки в момент глотания поступает в барабанную полость.
Слуховые косточки - стремя, наковальня и молоточек, названные так благодаря своей форме, самые мелкие в человеческом организме (рис. 139). Молоточек (malleus) имеет округлую головку (caput mallei), которая переходит в длинную рукоятку молоточка (manubrium mallei). У молоточка имеется два отростка: латеральный и передний
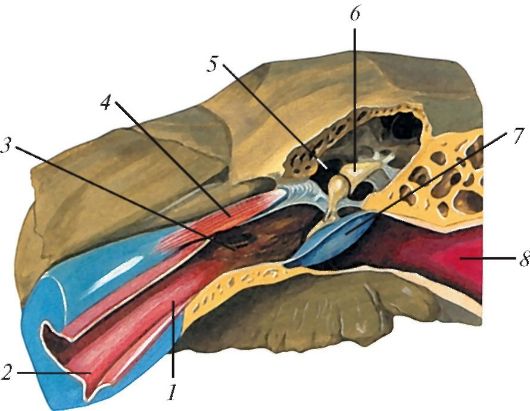 Рис. 138. Слуховая
труба, барабанная полость и наружный слуховой проход на фронтальном
разрезе: 1 - слуховая труба; 2 - хрящевая часть слуховой трубы; 3 -
костная часть слуховой трубы; 4 - мышца, напрягающая барабанную
перепонку; 5 - барабанная полость; 6 - слуховые косточки; 7 - барабанная
перепонка; 8 - наружный слуховой
Рис. 138. Слуховая
труба, барабанная полость и наружный слуховой проход на фронтальном
разрезе: 1 - слуховая труба; 2 - хрящевая часть слуховой трубы; 3 -
костная часть слуховой трубы; 4 - мышца, напрягающая барабанную
перепонку; 5 - барабанная полость; 6 - слуховые косточки; 7 - барабанная
перепонка; 8 - наружный слуховой
проход
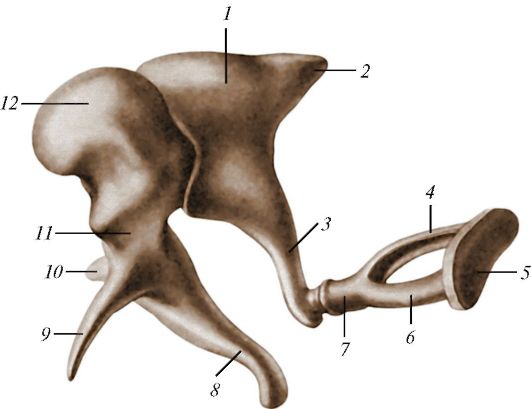 Рис. 139. Слуховые
косточки: 1 - наковальня; 2 - короткая ножка наковальни; 3 - длинная
ножка наковальни; 4 - задняя ножка стремени; 5 - основание стремени; 6 -
передняя ножка стремени; 7 - головка стремени; 8 - рукоятка молоточка; 9
- передний отросток молоточка; 10 - латеральный отросток молоточка; 11 -
шейка молоточка;
Рис. 139. Слуховые
косточки: 1 - наковальня; 2 - короткая ножка наковальни; 3 - длинная
ножка наковальни; 4 - задняя ножка стремени; 5 - основание стремени; 6 -
передняя ножка стремени; 7 - головка стремени; 8 - рукоятка молоточка; 9
- передний отросток молоточка; 10 - латеральный отросток молоточка; 11 -
шейка молоточка;
12 - головка молоточка
(processus lateralis et anterior). Наковальня (incus) состоит из тела, на котором находится суставная ямка для сочленения с головкой молоточка, и двух ножек - короткой (crus breve) и длинной (crus longum). На конце длинной ножки имеется утолщение - чечевицеобразный отросток (processus lentiformis). Стремя (stapes) имеет головку (caput stapedis) и две ножки - переднюю (crus anterius) и заднюю (crus posterius), соединенные между собой при помощи основания стремени (basis stapedis).
Слуховые косточки передают звуковые колебания от барабанной перепонки к окну преддверия. Рукоятка молоточка сращена с барабанной перепонкой. Головка молоточка и тело наковальни соединены между собой суставом (наковальне-молоточковый сустав, articulatio incudomallearis), укрепленным связками. Длинный отросток наковальни сочленяется с головкой стремечка (наковально-стременной сустав, articulatio incudostapedialis). Основание стремени входит в окно преддверия, соединяясь с его краем посредством кольцевой связки стремени (lig. anulare
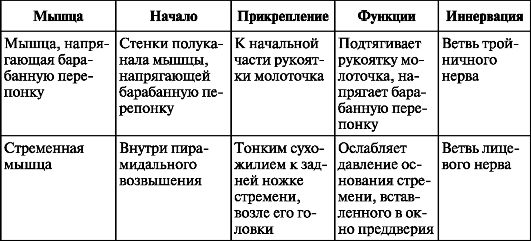
stapediale). Две мышцы регулируют движения косточек. Это мышца, напрягающая барабанную перепонку (musculus tensor tympani), прикрепляющаяся к рукоятке молоточка, и стременная мышца (muisculus stapedius), прикрепляющаяся к задней ножке стремени (табл. 12).
Внутреннее ухо (auris interna) расположено в толще пирамиды височной кости между барабанной полостью и внутренним слуховым проходом. Внутреннее ухо состоит из преддверия, трех полукружных каналов (протоков) и улитки (рис. 140). В костном лабиринте (labyrinthus (osseus), изнутри выстланном надкостницей, залегает перепончатый лабиринт (labyrinthus membranaceus), повторяющий формы костного (рис. 141). Между костным и перепончатым лабиринтами имеется узкая щель, заполненная перилимфой. Костное преддверие (vesiibulum) представляет собой овальную полость, имеющую на своей задней стенке пять тонких отверстий, которые ведут в полукружные каналы. Более крупное отверстие на передней стенке костного преддверия ведет в канал улитки. На латеральной стенке костного преддверия имеется окно преддверия (овальное), закрытое со стороны барабанной полости основанием стремени. У начала улитки находится окно улитки (круглое), открывающиееся на медиальной стенке барабанной полости среднего уха и закрытое вторичной барабанной перепонкой. На медиальной стенке преддверия расположен гребень, отделяющий друг от друга две ямки. Передняя ямка, округлая по форме, является сферическим углублением (recessus sphericus). Задняя ямка получила название эллиптического углубления (recessus ellipticus). В эллиптическом углублении находится внутреннее отверстие тонкого канальца - водопровода преддверия (agueductus vesiibuli),
Таблица 12. Мышцы барабанной полости
 в котором проходит эндолимфатический проток (ductus endolymphdticus), заканчивающийся на задней поверхности пирамиды височной кости (см. рис. 141, рис. 142).
в котором проходит эндолимфатический проток (ductus endolymphdticus), заканчивающийся на задней поверхности пирамиды височной кости (см. рис. 141, рис. 142).
Три дугообразно изогнутых костных полукружных канала (саnales semicirculdres dssei) лежат в трех взаимно перпендикулярных плоскостях. Ширина просвета каждого такого канала около 2 мм. Передний (сагиттальный, или верхний) полукружный канал (canalis semicirculdris anterior) ориентирован перпендикулярно продольной оси пирамиды височной кости. Латеральный (горизонтальный) полукружный канал (canalis semicirculdris lateralis) короче других полукружных каналов. Он образует на лабиринтной (медиальной) стенке барабанной полости возвышение - выступ латерального полукружного канала. Задний (фронтальный) полукружный канал (canalis semicirculdris posterior) - самый длинный из каналов. Он располагается почти параллельно задней поверхности пирамиды височной кости.
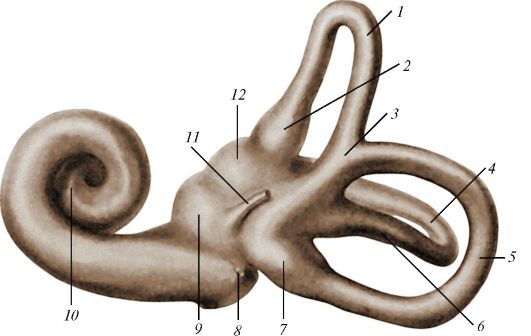 Рис. 140. Слепок
костного лабиринта внутреннего уха: 1 - передний полукружный канал; 2 -
передняя костная ампула; 3 - общая костная ножка; 4 - латеральный
полукружный канал; 5 - задний полукружный канал; 6 - простая ножка
латерального полукружного канала; 7 - задняя костная ампула; 8 - каналец
улитки; 9 - сферическое углубление; 10 - улитка; 11 - водопровод
преддверия; 12 - эллиптическое углубление (маточка)
Рис. 140. Слепок
костного лабиринта внутреннего уха: 1 - передний полукружный канал; 2 -
передняя костная ампула; 3 - общая костная ножка; 4 - латеральный
полукружный канал; 5 - задний полукружный канал; 6 - простая ножка
латерального полукружного канала; 7 - задняя костная ампула; 8 - каналец
улитки; 9 - сферическое углубление; 10 - улитка; 11 - водопровод
преддверия; 12 - эллиптическое углубление (маточка)
 Рис. 141. Строение перепончатого лабиринта и его положение в костном лабиринте (схема): 1
- эндолимфатический мешочек; 2 - эндолимфатический проток; 3 - проток
эллиптического мешочка; 4 - сферический мешочек; 5 - улитковый проток; 6
- лестница преддверия; 7 - барабанная лестница; 8 - наружное отверстие
канальца (водопровода) улитки; 9 - перилимфатический проток (водопровод)
улитки; 10 - окно улитки; 11 - слуховая труба; 12 - барабанная
перепонка; 13 - барабанная полость; 14 - молоточек; 15 - основание
стремени; 16 - наковальня; 17 - ампула заднего полукружного протока; 18 -
ампула латерального полукружного протока; 19 - перилимфатическое
пространство заднего (костного) полукружного канала; 20 - перепончатая
ножка (латеральный полукружный проток); 21 - эллиптический мешочек; 22 -
общая перепончатая ножка; 23 - передняя (верхняя) перепончатая ампула
Рис. 141. Строение перепончатого лабиринта и его положение в костном лабиринте (схема): 1
- эндолимфатический мешочек; 2 - эндолимфатический проток; 3 - проток
эллиптического мешочка; 4 - сферический мешочек; 5 - улитковый проток; 6
- лестница преддверия; 7 - барабанная лестница; 8 - наружное отверстие
канальца (водопровода) улитки; 9 - перилимфатический проток (водопровод)
улитки; 10 - окно улитки; 11 - слуховая труба; 12 - барабанная
перепонка; 13 - барабанная полость; 14 - молоточек; 15 - основание
стремени; 16 - наковальня; 17 - ампула заднего полукружного протока; 18 -
ампула латерального полукружного протока; 19 - перилимфатическое
пространство заднего (костного) полукружного канала; 20 - перепончатая
ножка (латеральный полукружный проток); 21 - эллиптический мешочек; 22 -
общая перепончатая ножка; 23 - передняя (верхняя) перепончатая ампула
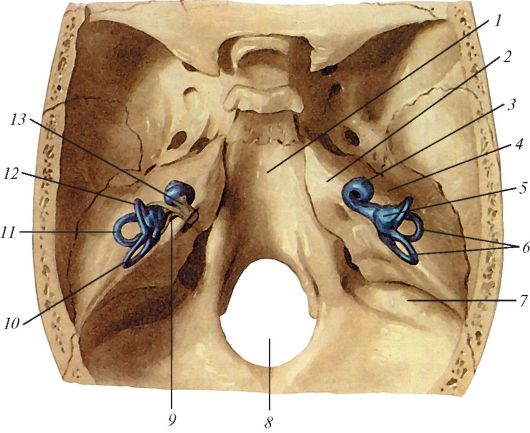 Рис. 142. Расположение
внутреннего уха по отношению к пирамиде височной кости, вид сверху
(схема): 1 - скат; 2 - тройничное вдавление; 3 - улитка; 4 - пирамида
(каменистая часть); 5 - преддверие лабиринта; 6 - костные полукружные
каналы; 7 - борозда сигмовидного синуса; 8 - большое затылочное
отверстие; 9 - преддверный нерв; 10 - задний полукружный канал; 11 -
латеральный полукружный канал;
Рис. 142. Расположение
внутреннего уха по отношению к пирамиде височной кости, вид сверху
(схема): 1 - скат; 2 - тройничное вдавление; 3 - улитка; 4 - пирамида
(каменистая часть); 5 - преддверие лабиринта; 6 - костные полукружные
каналы; 7 - борозда сигмовидного синуса; 8 - большое затылочное
отверстие; 9 - преддверный нерв; 10 - задний полукружный канал; 11 -
латеральный полукружный канал;
12 - передний полукружный канал; 13 - улитковый нерв
Каждый полукружный канал имеет по две ножки, одна из которых (ампулярная костная ножка - crus osseum ampullare) перед впадением в преддверие расширяется, образуя костную ампулу (ampulla ossea). Соседние костные ножки, переднего и заднего полукружных каналов соединяются, образуя общую костную ножку (crus osseum commune), поэтому три канала открываются в преддверие пятью отверстиями.
Костная улитка (cochlea) является передней частью костного лабиринта. Она представляет собой извитой спиральный канал улитки (canalis spiralis cochleae), образующий вокруг горизонтально лежащего стержня - костной оси улитки - 2,5 завитка. Основание улитки (basis cochleae) обращено медиально, в сторону внутреннего слухового прохода, а вершина
(купол улитки - cupula cochleae) направлена к барабанной полости. Вокруг костного стержня наподобие винта закручена костная спиральная пластина (lamina spiralis ossea), которая не полностью перегораживает спиральный канал улитки. Стержень (modiolus) пронизан продольными канальцами, в которых располагаются волокна улитковой части преддверно-улиткового нерва (рис. 143). В основании костной спиральной пластинки расположен спиральный канал стержня (canalis spiralis modioli), в котором лежит нервный узел улитки. Костная спиральная пластинка вместе с соединяющимся с ней перепончатым улитковым каналом (протоком) делит полость канала улитки на две «лестницы». Это лестница преддверия (scala vestibuli) и барабанная лестница (scala tympani), сообщающиеся между собой в области купола через отверстие улитки. В основании улитки имеется внутреннее отверстие канальца улитки, который выходит из пирамиды височной кости в углублении рядом с яремной ямкой и заканчивается наружной апертурой канальца улитки.
Перепончатый лабиринт (labyrinthus membranaceus) располагается внутри костного и в основном повторяет его очертания. Между внутренней поверхностью костного лабиринта и перепончатым лабиринтом имеется узкая щель - перилимфатическое пространство (spatium
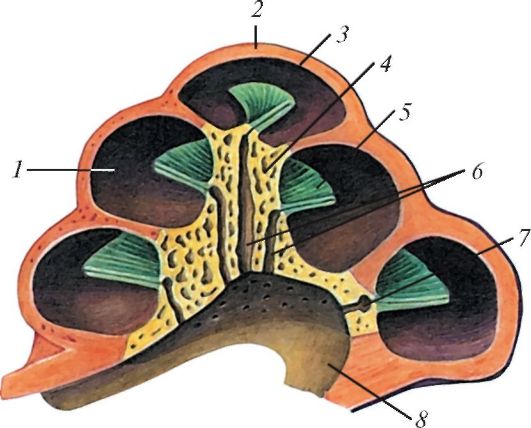 Рис. 143. Костная
улитка на предельном ее разрезе: 1 - спиральный канал улитки; 2 - купол
улитки; 3 - геликотрема; 4 - стержень улитки; 5 - костная спиральная
пластинка; 6 - продольные каналы стержня; 7 - спиральный канал стержня; 8
- основание улитки
Рис. 143. Костная
улитка на предельном ее разрезе: 1 - спиральный канал улитки; 2 - купол
улитки; 3 - геликотрема; 4 - стержень улитки; 5 - костная спиральная
пластинка; 6 - продольные каналы стержня; 7 - спиральный канал стержня; 8
- основание улитки
perilymphaticus). Оно сообщается с подпутинным пространством на нижней поверхности пирамиды височной кости через перилимфатический проток (ductus perilymphaticus), проходящий в костном канальце улитки. Стенки перепончатого лабиринта образованы плотной соединительной тканью. Изнутри перепончатый лабиринт выстлан однослойным плоским эпителием, лежащим на базальной мембране, и заполнен эндолимфой. Эндолимфа (endolympha) из перепончатого лабиринта может оттекать в эндолимфатический мешок (saccus endolymphaticus), находящийся в толще твердой мозговой оболочки на задней поверхности пирамиды. Отток эндолимфы происходит через эндолимфатический проток (ductus endolymphaticus), проходящий в канале водопровода преддверия (см. рис. 141).
У перепончатого лабиринта выделяют преддверную часть (эллиптический и сферический мешочки), три полукружных протока и улитковый проток, расположенные в соответствующих частях костного лабиринта внутреннего уха. Продолговатый эллиптический мешочек (utriculus - маточка) и сферический мешочек (sacculus) сообщаются друг с другом через тонкий каналец (проток эллиптического и сферического мешочков - ductus utriculosaccularis), от которого отходит эндолимфатический проток (ductus endolymphaticus). В нижней части сферический мешочек сообщается с перепончатым протоком улитки через соединяющий проток (ductus reuniens).
В эллиптический мешочек открываются пять отверстий перепончатых полукружных протоков (ductus semicirculares), расположенных в одноименных костных каналах. Одна из ножек каждого перепончатого протока, как и костного, расширяясь, образует перепончатую ампулу (переднюю, заднюю и латеральную - ampullae membranaceae anterior, posterior et lateralis). На внутренней поверхности эллиптического и сферического мешочков, а также на внутренней поверхности стенок перепончатых ампул полукружных протоков имеются покрытые желеподобным веществом образования, содержащие рецепторные (сенсорные) волосковые клетки, улавливающие изменения положения тела (головы) в пространстве (рис. 144). У эллиптического и сферического мешочков эти образования выглядят беловатыми пятнами (macula) размером 2 * 3 мм. Рецепторные клетки, расположенные здесь, воспринимают статические положения головы и прямолинейные движения, возникающие при колебаниях эндолимфы. В перепончатых ампулах имеются ампулярные гребешки (cristae ampulares), улавливающие любые повороты головы благодаря
ориентации трех полукружных протоков в трех взаимно перпендикулярных плоскостях.
Чувствительный эпителий пятен и ампулярных гребешков состоит из рецепторных волосковых (сенсорных) и поддерживающих эпителиальных клеток, лежащих на базальной мембране (рис. 145). На апикальных поверхностях волосковых клеток имеется по 60-80 ресничек (стереоцилий) длиной 35-40 мкм каждая, которые обращены в полость перепончатого лабиринта. Каждая рецепторная клетка снабжена также одной более длинной подвижной ресничкой. Реснички внедряются в богатую гликопротеидами тонковолокнистую желеобразную мембрану. В мембране пятен расположены многочисленные кристаллы углекислого кальция размерами 2-5 мкм - статолиты. Различают два вида рецепторных волосковых клеток: грушевидные и столбчатые.
Грушевидные рецепторные клетки у пятен и гребешков имеют короткую апикальную часть и широкое закругленное основание, окруженное чашеобразным футляром, образованным нервными окончаниями. Между аксолеммой нервного окончания и цитолеммой рецепторной клетки имеются синапсы. Столбчатые рецепторные клетки имеют цилиндрическую форму. К цитолемме в области основания клетки подходят нервные окончания, формирующие синапсы. Пятно сферического мешочка содержит около 18 000 рецепторных клеток, эллиптического мешочка - около 33 000. Рецепторные клетки иннервируются афферентными волокнами нейронов вестибулярного ганглия. Поддерживающие (опорные) клетки расположены между рецепторными волосковыми клетками. Апикальная поверхность поддерживающих клеток несет большое число коротких тонких микроворсинок, в цитоплазме находятся секреторные гранулы и множество митохондрий.
Проводящие пути органа равновесия. Рецепторные волосковые клетки пятен воспринимают изменение силы тяжести и линейного ускорения, поэтому при воздействии линейного ускорения мембрана сдвигается на поверхности эпителия, увлекая за собой реснички, что и становится стимулом для возникновения нервного импульса. При вертикальном положении головы пятно эллиптического мешочка расположено горизонтально, и мембрана не сдвигается. При наклонах головы пятно располагается под углом, мембрана сдвигается, благодаря чему ресничка изгибается, что также становится стимулом. Аналогичным образом функционирует и пятно сферического мешочка, с той лишь разницей,
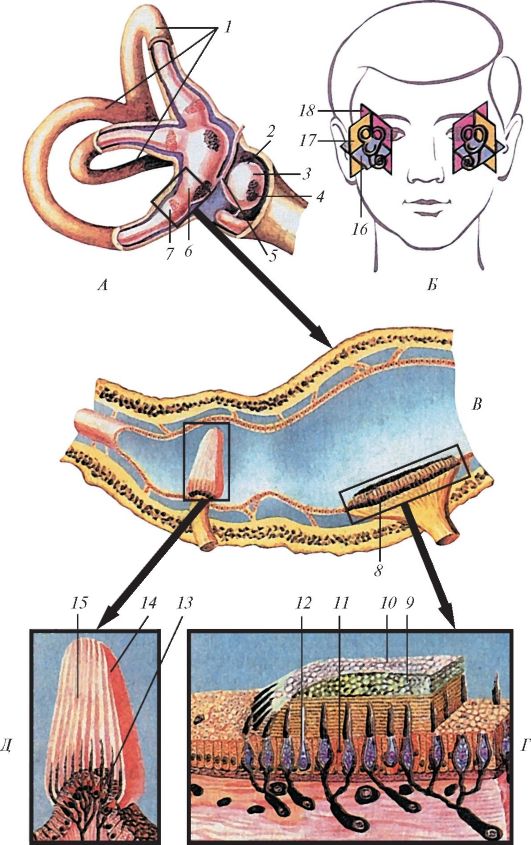 Рис. 144. Орган
равновесия и детали его строения: А - полукружные протоки и
эндолимфатические мешочки преддверного лабиринта (костная стенка
лабиринта частично удалена); Б - ориентация полукружных протоков
преддверного лабиринта внутреннего уха; В - расположение ампулярного
гребешка и пятна в стенке перепончатого лабиринта; Г - строение пятна
(на продольном разрезе); Д - строение ампулярного гребешка (на
продольном разрезе). Стрелками показаны места расположения структур в
лабиринте; 1 - полукружные каналы; 2 - преддверие; 3 - сферический
мешочек; 4 - пятно сферического мешочка; 5 - эндолимфатический проток; 6
- эллиптический мешочек; 7 - ампула; 8 - отолитовый аппарат; 9 -
статоконии; 10 - мембрана статоконий; 11 - поддерживающие клетки; 12, 13
- волосковые сенсорные клетки; 14 - ампулярный гребешок; 15 - купол; 16
- латеральный полукружный канал; 17 - передний полукружный канал; 18 -
задний полукружный канал
Рис. 144. Орган
равновесия и детали его строения: А - полукружные протоки и
эндолимфатические мешочки преддверного лабиринта (костная стенка
лабиринта частично удалена); Б - ориентация полукружных протоков
преддверного лабиринта внутреннего уха; В - расположение ампулярного
гребешка и пятна в стенке перепончатого лабиринта; Г - строение пятна
(на продольном разрезе); Д - строение ампулярного гребешка (на
продольном разрезе). Стрелками показаны места расположения структур в
лабиринте; 1 - полукружные каналы; 2 - преддверие; 3 - сферический
мешочек; 4 - пятно сферического мешочка; 5 - эндолимфатический проток; 6
- эллиптический мешочек; 7 - ампула; 8 - отолитовый аппарат; 9 -
статоконии; 10 - мембрана статоконий; 11 - поддерживающие клетки; 12, 13
- волосковые сенсорные клетки; 14 - ампулярный гребешок; 15 - купол; 16
- латеральный полукружный канал; 17 - передний полукружный канал; 18 -
задний полукружный канал
что при вертикальном положении головы пятна расположены вертикально.
Чувствительные аппараты полукружных протоков - ампулярные гребешки в виде складок располагаются в стенках каждой ампулы. Гребешки имеют такие же волосковые и поддерживающие клетки, как и пятна преддверия. На этих клетках лежит желатинообразный купол, куда проникают стереоцилии и реснички. Ампулярные гребешки воспринимают изменение углового ускорения. При поворотах головы желатиновый купол движется в противоположном направлении, сдвигая реснички. В результате изменяется положение ресничек, что, в свою очередь, вызывает изменение активности различных ферментов волосковых клеток и приводит к возникновению импульса. Импульс через синапсы передается афферентным нервным волокнам и по ним к телам нейронов преддверного нервного узла, лежащего на дне внутреннего слухового прохода. Аксоны клеток преддверного узла образуют преддверную часть преддверно-улиткового нерва (VIII черепной нерв), который выходит вместе с улитковой частью через внутреннее слуховое отверстие в полость черепа. В мосто-мозжечковом углу волокна нерва входят в вещество мозга и подходят к вестибулярным ядрам, расположенным в области вестибулярного поля ромбовидной ямки (рис. 146). Это верхнее ядро (Бехтерева), медиальное (Швальбе), латеральное (Дейтерса) и нижнее ядро (Роллера). Аксоны нейронов вестибулярных ядер образуют связи со многими структурами центральной нервной системы.
От вестибулярных ядер часть волокон, перекрещиваясь, идет в таламус, где расположены нейроны, откуда импульсы направляются к коре постцентральной извилины теменной и височной долей (корковые
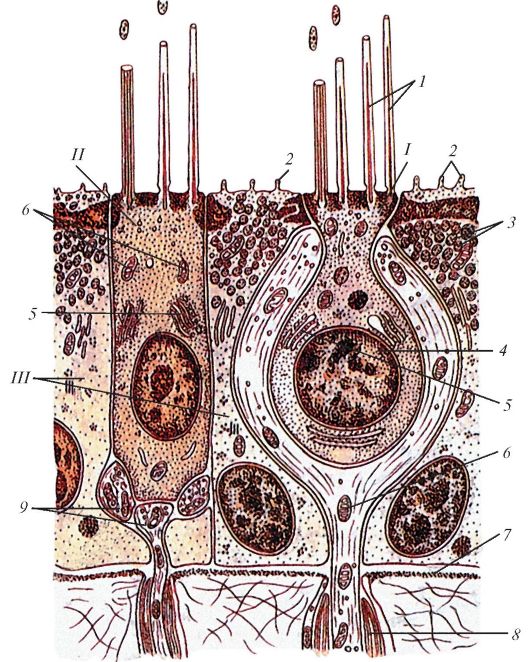 Рис. 145. Схема
ультрамикроскопического строения волосковых и поддерживающих клеток: I -
волосковая клетка первого типа; II - волосковая клетка второго типа;
III - поддерживающая клетка; 1 - статические волоски; 2 - микроворсинки
поддерживающей клетки; 3 - гранулы в поддерживающих клетках; 4 - ядро; 5
- внутренний сетчатый аппарат (комплекс Гольджи); 6 - митохондрия; 7 -
базальная мембрана; 8 - миелиновое нервное волокно; 9 - нервные
окончания (по И.В. Ал-
Рис. 145. Схема
ультрамикроскопического строения волосковых и поддерживающих клеток: I -
волосковая клетка первого типа; II - волосковая клетка второго типа;
III - поддерживающая клетка; 1 - статические волоски; 2 - микроворсинки
поддерживающей клетки; 3 - гранулы в поддерживающих клетках; 4 - ядро; 5
- внутренний сетчатый аппарат (комплекс Гольджи); 6 - митохондрия; 7 -
базальная мембрана; 8 - миелиновое нервное волокно; 9 - нервные
окончания (по И.В. Ал-
мазову и Л.С. Сутулову)
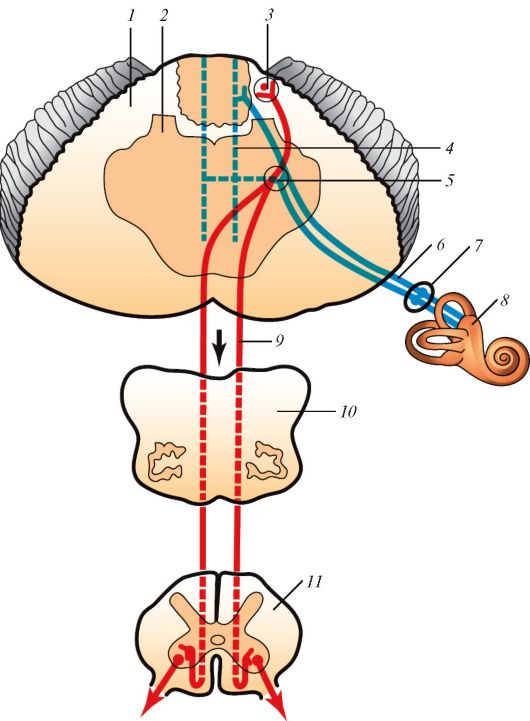 Рис. 146. Проводящий путь статического (вестибулярного) анализатора:
Рис. 146. Проводящий путь статического (вестибулярного) анализатора:
1 - мозжечок; 2 - мост; 3 - ядро шатра; 4 - задний продольный пучок; 5 - преддверные (вестибулярные) ядра; 6 - преддверная (вестибулярная) часть преддверно-улиткового нерва; 7 - вестибулярный узел; 8 - внутреннее ухо; 9 - преддверно-спинномозговой путь; 10 - разрез продолговатого мозга;
11 - разрез спинного мозга. Стрелки показывают направление следования
нервных импульсов
центры статокинетического анализатора). Эти связи обеспечивают сознательную ориентацию в пространстве. Часть нервных волокон преддверной части преддверно-улиткового нерва направляется непосредственно в мозжечок (в узелок и в клочок мозжечка). По волокнам преддверно-спинномозгового пути от вестибулярных ядер нервные импульсы идут к двигательным ядрам передних рогов спинного мозга. Эти связи регулируют вестибулярные рефлексы. Связи вестибулярных ядер с ядрами глазодвигательных нервов, которые осуществляют движения глаз, имеют отношение к изменениям положения головы и тела в пространстве.
Перепончатый лабиринт улитки, или улитковый лабиринт (labyrinthus cochlearis), начинается слепо в преддверии внутреннего уха, позади впадающего в него соединяющего протока, и продолжается внутри костного спирального канала улитки. В области верхушки улитки улитковый проток заканчивается также слепо. Длина улиткового протока около 3,5 см. На поперечном разрезе он имеет форму треугольника (рис. 147), занимающего лишь центральную часть спирального канала улитки. Наружная стенка улиткового протока срастается с надкостницей наружной стенки костной улитки и представлена спиральной связкой ( ligamentum spirale), которая имеется на всем протяжении улиткового протока. На связке расположена богатая капиллярами сосудистая полоска (stria vascularis), покрытая кубическими эпителиоцитами. Эпителиоциты сосудистой полоски образуют 1-3 слоя клеток. Более мелкие базальные клетки лежат на базальной мембране. Поверхностный слой образуют высокие призматические клетки, очень богатые митохондриями. Между клетками обоих типов располагаются светлые клетки с крупными ядрами и малым числом митохондрий. Между эпителиоцитами проходят многочисленные гемокапилляры со сплошным слоем эндотелия, окруженные собственной базальной пластинкой, в которой расположены перициты. В образовании эндолимфы участвуют гемокапилляры и эпителиоциты сосудистой полоски.
Верхняя стенка улиткового протока - преддверная стенка (мембрана Рейснера, преддверная мембрана - paries vestibularis ductus cochlearis), обращенная к лестнице преддверия, простирается от свободного края спиральной костной пластинки косо вверх к наружной стенке улиткового протока. Изнутри преддверная стенка покрыта однослойным плоским эпителием. Нижняя - барабанная стенка улиткового протока (paries tympanicus ductus cochlearis), обращенная к барабанной лестнице улитки,
устроена наиболее сложно. Она одним краем прикреплена к спиральной костной пластинке и является как бы ее продолжением, а другим краем срастается с наружной стенкой улиткового протока. Барабанная стенка представляет собой базилярную (спиральную) пластинку (мембрану) (lamina basilaris), на которой располагается спиральный орган (ganglion spirale - кортиев орган), осуществляющий восприятия звуковой волны. Таким образом, улитковый проток, расположенный между свободным краем спиральной костной пластинки и наружной стенкой костной улитки, отделяет барабанную лестницу (scala tympani), граничащую со спиральной мембраной, от лестницы преддверия (scala vestibuli), прилежащей к преддверной мембране. В области купола улитки обе лестницы сообщаются друг с другом при помощи отверстия улитки - геликотремы (helicotrema). В основании улитки барабанная лестница заканчивается у окна улитки, закрытого вторичной барабанной перепонкой.
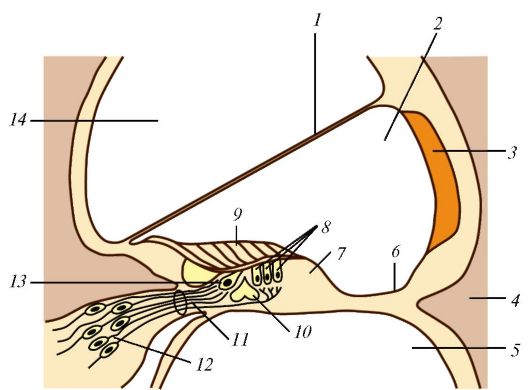 Рис. 147. Строение
улиткового протока внутреннего уха (схема): 1 - преддверная мембрана; 2
- улитковый проток; 3 - сосудистая полоска; 4 - кость; 5 - барабанная
лестница; 6 - базилярная пластинка; 7 - спиральный (кортиев) орган; 8 -
наружные волосковые клетки спирального (кортиева) органа; 9 - покровная
мембрана; 10 - внутренний туннель; 11 - нервные волокна; 12 - узел
(спиральный) улитки; 13 - внутренняя волосковая клетка; 14 - лестница
преддверия
Рис. 147. Строение
улиткового протока внутреннего уха (схема): 1 - преддверная мембрана; 2
- улитковый проток; 3 - сосудистая полоска; 4 - кость; 5 - барабанная
лестница; 6 - базилярная пластинка; 7 - спиральный (кортиев) орган; 8 -
наружные волосковые клетки спирального (кортиева) органа; 9 - покровная
мембрана; 10 - внутренний туннель; 11 - нервные волокна; 12 - узел
(спиральный) улитки; 13 - внутренняя волосковая клетка; 14 - лестница
преддверия
Лестница преддверия переходит в перилимфатическое пространство преддверия, в овальное окно которого со стороны барабанной полости вставлено основание стремени.
Функционально важна для работы спирального (кортиева) органа базилярная пластинка, которая образована тонкими радиальными коллагеновыми волокнами (около 24 000), погруженными в аморфное вещество. Длина этих волокон возрастает от основания улитки к ее вершине. Толщина базилярной пластинки увеличивается от 0,21 мм в основании улитки до 0,36 мм у ее верхушки. Кнаружи от спирального (кортиева) органа базилярная пластинка со стороны улиткового протока покрыта однослойным кубическим эпителием, который переходит в эпителиоциты, покрывающие сосудистую полоску. Со стороны барабанной лестницы находится однослойный плоский эпителий, под базальной мембраной которого проходят кровеносные капилляры, окруженные тонким слоем рыхлой соединительной ткани.
На базилярной пластинке (мембране) расположен спиральный (кортиев) орган, образованный чувствительными (сенсорными) клетками и поддерживающими эпителиоцитами. Среди клеток обоих типов различают внутренние, расположенные кнутри от внутреннего туннеля (ближе к спиральному ганглию), и наружные, лежащие кнаружи от туннеля (ближе к сосудистой полоске). Непосредственно на базилярной мембране лежат наружные и внутренние опорные (поддерживающие) клетки, а на них - рецепторные волосковые (сенсорные) клетки (рис. 148). Внутренние и наружные рецепторные волосковые клетки сходятся вершинами под острым углом друг к другу и образуют канал треугольной формы - внутренний (кортиев) туннель. Этот туннель, заполненный эндолимфой, проходит спирально вдоль всего кортиева органа. В туннеле расположены нервные волокна, идущие от нейронов спирального ганглия. Внутренние волосковые сенсорные клетки грушевидной формы расположены в один ряд, рядом с внутренними поддерживающими эпителиоцитами. Между мембранами обеих клеток имеется узкая щель. В базальной части волосковых клеток находится округлое ядро, в цитоплазме имеются митохондрии, актиновые и миозиновые микрофиламенты, зернистая эндоплазматическая сеть, центриоли, комплекс Гольджи. От апикальной поверхности каждой сенсорной клетки отходит 30-60 стереоцилий, расположенных в 3-4 ряда. Стереоцилии имеют длину от 20 до 40 мкм. Апикальная часть сенсорной клетки окружена отростками (фалангами) внутренних поддерживающих (фаланговых) эпителиоцитов.
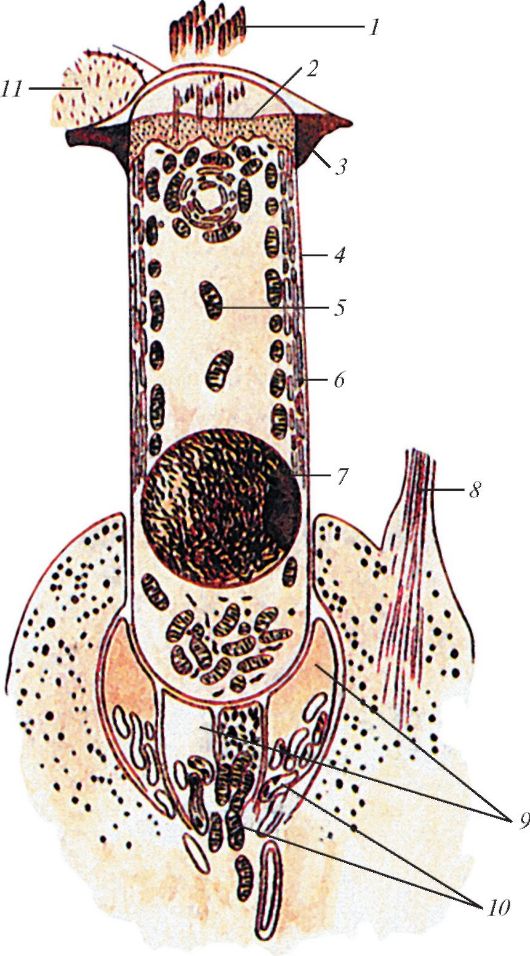 Рис. 148. Схема
ультрамикроскопического строения волосковой сенсорной клетки
спирального (кортиева) органа: 1 - слуховые волоски; 2 - кутикула; 3 -
сетчатая мембрана; 4 - цитолемма клетки; 5 - митохондрия; 6 -
эндоплазматическая сеть; 7 - ядро; 8 - фаланговая пластинка наружной
поддерживающей клетки; 9 - нервные окончания; 10 - митохондрии в нервном
окончании; 11 - микроворсинки на опорных клетках (по В.Г. Елисееву и
др.)
Рис. 148. Схема
ультрамикроскопического строения волосковой сенсорной клетки
спирального (кортиева) органа: 1 - слуховые волоски; 2 - кутикула; 3 -
сетчатая мембрана; 4 - цитолемма клетки; 5 - митохондрия; 6 -
эндоплазматическая сеть; 7 - ядро; 8 - фаланговая пластинка наружной
поддерживающей клетки; 9 - нервные окончания; 10 - митохондрии в нервном
окончании; 11 - микроворсинки на опорных клетках (по В.Г. Елисееву и
др.)
Наружные волосковые сенсорные клетки (столбовые) лежат в 3-4 параллельных ряда. Каждая из них погружена в тело наружной поддерживающей клетки наподобие описанных выше внутренних сенсорных клеток. Наружные сенсорные клетки имеют цилиндрическую форму, в их цитоплазме содержатся такие же органеллы, как и у внутренних. Цитоплазма сенсорных клеток обоих типов богата различными окислительными ферментами. Активность ферментов возрастает при коротких звуковых сигналах и снижается при длительных. Стереоцилии наружных сенсорных клеток, аналогичные стереоцилиям внутренних, располагаются несколькими рядами в виде латинской цифры V.
У человека около 3500 внутренних и 12 000 наружных сенсорных волосковых клеток. Основания сенсорных клеток контактируют с нервными окончаниями, образуя с ними синапсы. Около 30 000-40 000 нервных волокон являются дендритами биполярных клеток, залегающих в спиральном ганглии, расположенном в толще (в канале) спиральной костной пластинки. Около 90% дендритов нейронов спирального ганглия контактируют с внутренними сенсорными клетками и лишь 10% - с наружными, поэтому каждое из этих волокон разветвляется и образует синапсы со многими клетками.
Над стереоцилиями сенсорных клеток по всей длине спирального (кортиева) органа расположена желатинозная покровная мембрана ( membrtana tecttoria), которая состоит из тонкого мелкозернистого слоя, обращенного к сенсорным клеткам, и грубоволокнистого слоя, лежащего над ним. В образовании покровной мембраны участвуют также радиально ориентированные коллагеновые волокна, залегающие в аморфном веществе, богатом гликозаминогликанами. Один край покровной мембраны прикрепляется к костной спиральной пластинке, а другой свободно оканчивается в полости улиткового протока над рецепторными клетками.
ФУНКЦИИ СЛУХОВОГО АНАЛИЗАТОРА. ПРОВОДЯЩИЕ ПУТИ СЛУХОВОГО АНАЛИЗАТОРА
Здоровый человек способен различать звуки в диапазоне от 20 до 16 000 Гц (герц). Однако понимание речи (речевая зона) находится в пределах от 300 до 3500 Гц. Высота звука зависит от частоты звуковых колебаний. Звуковые волны передаются через наружный слуховой проход и
достигают барабанной перепонки, колебания которой передаются через цепь слуховых косточек на окно преддверия (рис. 149). Движения стремени в окне преддверия вызывают колебания перилимфы лестницы преддверия, которые через отверстие в области верхушки улитки передаются перилимфе барабанной лестницы. Колебания перилимфы воспринимаются также эндолимфой. При этом происходят волнообразные движения базилярной мембраны, которая в зависимости от частоты и интенсивности звука колеблется по всей своей длине. Благодаря этому волосковые сенсорные клетки колеблются, их реснички касаются покровной мембраны. В результате этих касаний ресничек сенсорных клеток с покровной мембраной возникает нервный импульс. Эти импульсы проводятся по нервным волокнам клеток спирального узла (ганглия) в составе улитковой части преддверно-улиткового нерва (VIII черепной нерв). Аксоны клеток спирального узла заканчиваются синапсами на нейронах переднего (вентрального) и заднего (дорсального) улитковых
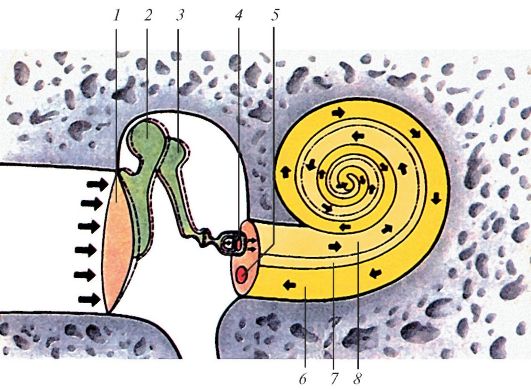 Рис. 149. Распространение
звуковой волны (показано стрелками) в наружном, среднем и внутреннем
ухе: 1 - барабанная перепонка; 2 - молоточек; 3 - наковальня; 4 -
стремя; 5 - круглое окно; 6 - барабанная лестница; 7 - улитковый проток;
8 - лестница преддверия
Рис. 149. Распространение
звуковой волны (показано стрелками) в наружном, среднем и внутреннем
ухе: 1 - барабанная перепонка; 2 - молоточек; 3 - наковальня; 4 -
стремя; 5 - круглое окно; 6 - барабанная лестница; 7 - улитковый проток;
8 - лестница преддверия
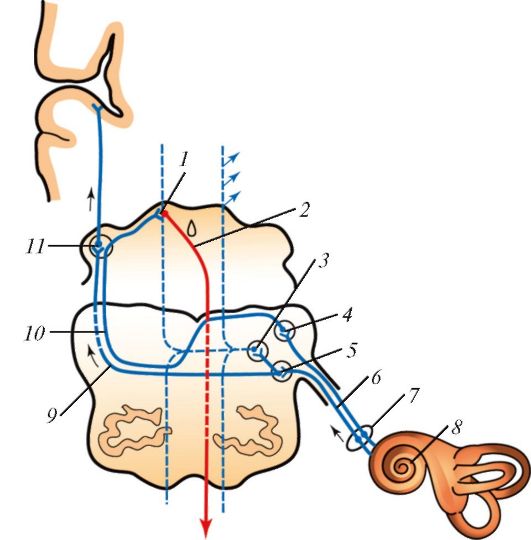 Рис. 150. Проводящий
путь слухового анализатора: 1 - нижний холмик среднего мозга; 2 -
покрышечно-спинномозговой путь; 3 - ядро трапециевидного тела; 4 -
заднее улитковое ядро; 5 - переднее улитковое ядро; 6 - улитковая часть
преддверно-улиткового нерва (VIII); 7 - спиральный узел; 8 - улитка; 9 -
трапециевидное тело; 10 - латеральная петля; 11 - медиальное коленчатое
тело. Стрелками показан путь распространения
Рис. 150. Проводящий
путь слухового анализатора: 1 - нижний холмик среднего мозга; 2 -
покрышечно-спинномозговой путь; 3 - ядро трапециевидного тела; 4 -
заднее улитковое ядро; 5 - переднее улитковое ядро; 6 - улитковая часть
преддверно-улиткового нерва (VIII); 7 - спиральный узел; 8 - улитка; 9 -
трапециевидное тело; 10 - латеральная петля; 11 - медиальное коленчатое
тело. Стрелками показан путь распространения
(слуховых) ядер, которые лежат в продолговатом мозге в области вестибулярного поля ромбовидной ямки (рис. 150). Отростки клеток переднего ядра направляются на противоположную сторону, образуя трапециевидное тело. Аксоны заднего ядра выходят на поверхность ромбовидной ямки и в виде мозговых полосок IV желудочка мозга направляются к срединной борозде ромбовидной ямки, затем погружаются внутрь вещества мозга и присоединяются к волокнам трапециевидного тела.
На противоположной стороне моста волокна трапециевидного тела изгибаются, давая начало латеральной петле. После этого нервные волокна направляются далее к подкорковым центрам слуха: медиальному коленчатому телу и нижнему холмику пластинки крыши среднего мозга.
Часть аксонов улитковых ядер (слухового пути) заканчивается на клетках медиального коленчатого тела, где передают импульс следующему нейрону. Отростки этих нейронов, пройдя через подчечевицеобразную часть задней ножки внутренней капсулы, направляются к слуховому центру (корковому концу слухового анализатора), который находится в коре верхней височной извилины (поле 41, по Бродману). Здесь осуществляются высший анализ и синтез нервных импульсов, поступающих из звуковоспринимающего аппарата. Другая часть аксонов улитковых ядер проходит транзитом через медиальное коленчатое тело, а затем через ручку нижнего холмика вступает в его ядро и заканчивается на клетках этого ядра. Аксоны этих клеток образуют экстрапирамидный покрышечно-спинномозговой путь, который передает импульсы из нижних холмиков пластинки крыши среднего мозга клеткам двигательных ядер передних рогов спинного мозга.
Кровоснабжение и иннервация органа слуха и равновесия. Орган слуха и равновесия кровоснабжается из нескольких источников (табл.13). К наружному уху подходят ветви из системы наружной сонной артерии: передние ушные ветви поверхностной височной артерии, ушные ветви затылочной артерии и задняя ушная артерия. В стенках наружного
Таблица 13. Артерии, разветвляющиеся в слизистой оболочке и мышцах барабанной полости и в барабанной перепонке
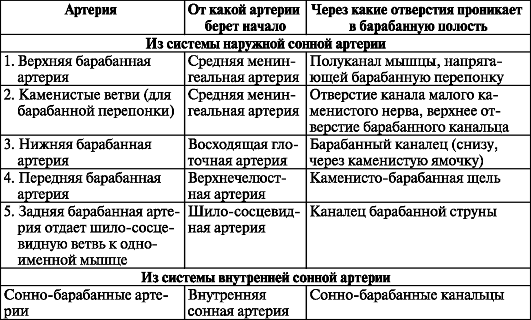 слухового
прохода разветвляется глубокая ушная артерия (из верхнечелюстной
артерии). Эта же артерия участвует в кровоснабжении барабанной
перепонки, которая получает кровь также из артерий, кровоснабжающих
слизистую оболочку барабанной полости. В результате в перепонке
образуются две сосудистые сети: одна в кожном слое, другая - в слизистой
оболочке. Венозная кровь от наружного уха по одноименным венам оттекает
в занижнечелюстную вену, а из нее - в наружную яремную вену.
слухового
прохода разветвляется глубокая ушная артерия (из верхнечелюстной
артерии). Эта же артерия участвует в кровоснабжении барабанной
перепонки, которая получает кровь также из артерий, кровоснабжающих
слизистую оболочку барабанной полости. В результате в перепонке
образуются две сосудистые сети: одна в кожном слое, другая - в слизистой
оболочке. Венозная кровь от наружного уха по одноименным венам оттекает
в занижнечелюстную вену, а из нее - в наружную яремную вену.
В слизистой оболочке барабанной полости разветвляются передняя барабанная артерия (ветвь верхнечелюстной артерии), верхняя барабанная артерия (ветвь средней менингеальной артерии), задняя барабанная артерия (ветви шилососцевидной артерии), нижняя барабанная артерия (из восходящей глоточной артерии), сонно-барабанная артерия (из внутренней сонной артерии).
Стенки слуховой трубы кровоснабжают переднюю барабанную артерию и глоточные ветви (из восходящей глоточной артерии), а также каменистая ветвь средней менингеальной артерии. К слуховой трубе отдает веточки артерия крыловидного канала (ветвь верхнечелюстной артерии). Вены среднего уха сопровождают одноименные артерии и впадают в глоточное венозное сплетение, в менингеальные вены (притоки внутренней яремной вены) и в занижнечелюстную вену.
К внутреннему уху подходит лабиринтная артерия (ветвь базилярной артерии), сопровождающая преддверно-улитковый нерв и отдающая две ветви: вестибулярную и общую улитковую. От первой отходят ветви к эллиптическому и сферическому мешочкам и полукружным каналам, где они разветвляются до капилляров. Улитковая ветвь кровоснабжает спиральный ганглий, спиральный орган и другие структуры улитки. Венозная кровь оттекает по лабиринтной вене в верхний каменистый синус.
Лимфа от наружного и среднего уха оттекает в сосцевидные, околоушные, глубокие латеральные шейные (внутренние яремные) лимфатические узлы, от слуховой трубы - в заглоточные лимфатические узлы.
Чувствительную иннервацию наружное ухо получает из большого ушного, блуждающего и ушно-височного нервов, барабанная перепонка - от ушно-височного и блуждающего нервов, а также от барабанного сплетения барабанной полости. В слизистой оболочке барабанной полости нервное сплетение образовано ветвями барабанного
нерва (из языкоглоточного нерва), соединительной ветвью лицевого нерва с барабанным сплетением и симпатическими волокнами сонно-барабанных нервов (от внутреннего сонного сплетения). Барабанное сплетение продолжается в слизистой оболочке слуховой трубы, куда проникают также ветви от глоточного сплетения. Барабанная струна проходит через барабанную полость транзитом, в ее иннервации не участвует.
РАЗВИТИЕ ОРГАНОВ СЛУХА И РАВНОВЕСИЯ В ОНТОГЕНЕЗЕ
Образование перепончатого лабиринта в онтогенезе у человека начинается с утолщения эктодермы на поверхности головного отдела зародыша по бокам от нервной пластинки. На 4-й неделе внутриутробного развития эктодермальное утолщение прогибается, образует слуховую ямку, которая превращается в слуховой пузырек, обособляющийся от эктодермы и погружающийся внутрь головного отдела зародыша (на 6- й неделе). Пузырек состоит из многорядного эпителия, секретирующего эндолимфу, заполняющую просвет пузырька. Затем пузырек делится на две части. Одна часть (вестибулярная) превращается в эллиптический мешочек с полукружными протоками, вторая часть образует сферический мешочек и улитковый лабиринт. Размеры завитков увеличиваются, улитка растет и отделяется от сферического мешочка. В полукружных протоках развиваются гребешки, в маточке и сферическом мешочке - пятна, в которых расположены нейросенсорные клетки.
В течение 3-го месяца внутриутробного развития в основном заканчивается формирование перепончатого лабиринта. Одновременно начинается образование спирального органа. Из эпителия улиткового протока формируется покровная мембрана, под которой дифференцируются волосковые рецепторные (сенсорные) клетки. Разветвления периферической части преддверно-улиткового нерва (VIII черепной нерв) соединяются с указанными рецепторными (волосковыми) клетками. Одновременно с развитием перепончатого лабиринта вокруг него из мезенхимы сначала образуется слуховая капсула, которая замещается хрящом, а затем костью.
Полость среднего уха развивается из первого глоточного кармана и боковой части верхней стенки глотки. Слуховые косточки происходят из хряща первой (молоточек и наковальня) и второй (стремя) висцеральных дуг. Проксимальная часть первого (висцерального) кармана суживается и превращается в слуховую трубу. Появляющееся напротив
формирующейся барабанной полости впячивание эктодермы - жаберная борозда в дальнейшем преобразуется в наружный слуховой проход. Наружное ухо начинает формироваться у зародыша на 2-м месяце внутриутробной жизни в виде шести бугорков, окружающих первую жаберную щель.
Ушная раковина у новорожденного уплощена, ее хрящ мягкий, покрывающая его кожа тонкая. Наружный слуховой проход у новорожденного узкий, длинный (около 15 мм), круто изогнут, имеет сужение на границе расширенных медиального и латерального отделов. Наружный слуховой проход, за исключением барабанного кольца, имеет хрящевые стенки. Барабанная перепонка у новорожденного относительно велика и почти достигает размеров перепонки взрослого человека - 9 х 8 мм. Она наклонена сильнее, чем у взрослого, угол наклона равен 35-40° (у взрослого 45-55°). Размеры слуховых косточек и барабанной полости у новорожденного и взрослого различаются мало. Стенки барабанной полости тонкие, особенно верхняя. Нижняя стенка местами представлена соединительной тканью. Задняя стенка имеет широкое отверстие, ведущее в сосцевидную пещеру. Сосцевидные ячейки у новорожденного отсутствуют из-за слабого развития сосцевидного отростка.
Слуховая труба у новорожденного прямая, широкая, короткая (17-21 мм). В течение 1-го года жизни ребенка слуховая труба растет медленно, на 2-м году быстрее. Длина слуховой трубы у ребенка в 1-й год жизни равна 20 мм, в 2 года - 30 мм, в 5 лет - 35 мм, у взрослого человека - 35-38 мм. Просвет слуховой трубы постепенно суживается от 2,5 мм у 6-месячного ребенка до 1-2 мм у 6-летнего.
Внутреннее ухо к моменту рождения развито хорошо, его размеры близки к таковым у взрослого человека. Костные стенки полукружных каналов тонкие, постепенно утолщаются в результате слияния ядер окостенения в пирамиде височной кости.
АНОМАЛИИ РАЗВИТИЯ ОРГАНОВ СЛУХА И РАВНОВЕСИЯ
Нарушения развития рецепторного аппарата (спирального органа), недоразвитие слуховых косточек, препятствующее их движению, ведут к врожденной глухоте. Иногда встречаются дефекты положения, формы и строения наружного уха, которые, как правило, связаны с недоразвитием нижней челюсти (микрогнатия) или даже ее отсутствием (агнатия).
ОРГАН ОБОНЯНИЯ
Орган обоняния (organum olfactorium) располагается в обонятельной области слизистой оболочки полости носа площадью 250-500 мм2 у взрослого человека. Верхняя носовая раковина и лежащая на этом же уровне зона носовой перегородки покрыты многорядным столбчатым обонятельным эпителием высотой около 60 мкм, лежащим на базальной мембране. Благодаря накоплению в некоторых клетках пигмента эпителий имеет желтоватый цвет. Эпителий представлен обонятельными нейросенсорными клетками, которые расположены среди поддерживающих эпителиоцитов, а также базальными клетками (рис. 151). Поддерживающие клетки лежат на базальной мембране между нейросенсорными, разделяя их. Это высокие призматические клетки с суженной базальной частью. Круглые ядра поддерживающих клеток образуют верхний ряд ядер обонятельного эпителия. Обонятельные нейросенсорные клетки имеют на апикальной поверхности множество длинных тонких микроворсинок.
Мелкие базальные клетки лежат глубже, на базальной мембране, окружают пучки аксонов обонятельных нейросенсорных клеток. Различают две разновидности базальных клеток. Одни из них постоянно находятся в базальном слое. Это различные полигональные клетки, содержащие крупные ядра неправильной формы, окруженные узким ободком цитоплазмы. Многочисленные тонофибриллы прикрепляются к десмосомам, которые соединяют базальные и поддерживавшие эпителиоциты. Базальные клетки второй разновидности окружают основания нейросекреторных клеток.
Нейросенсорных обонятельных клеток у человека около 10-40 млн, а у макросмических животных их число достигает 200 млн и более. Обонятельные нейросенсорные клетки представляют собой видоизмененные
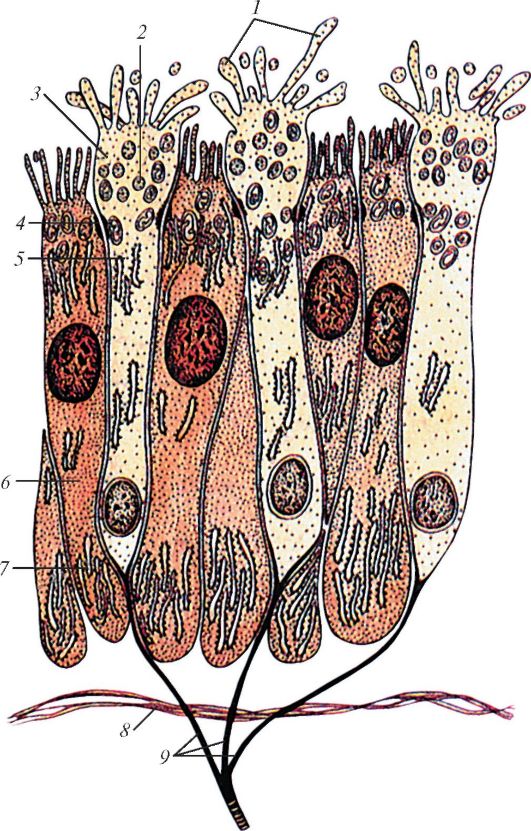 Рис. 151. Схема
ультрамикроскопического строения обонятельного эпителия: 1 -
микроворсинки; 2 - пузырьки; 3 - обонятельная булава; 4 - замыкательная
пластинка (десмосома); 5 - тело обонятельной нейросенсорной клетки; 6 -
поддерживающая клетка; 7 - эндоплазматическая сеть; 8 - базальная
мембрана; 9 - аксоны обонятельных нейросенсорных клеток, образующие
обонятельные нити (по В.Г. Елисееву и др.)
Рис. 151. Схема
ультрамикроскопического строения обонятельного эпителия: 1 -
микроворсинки; 2 - пузырьки; 3 - обонятельная булава; 4 - замыкательная
пластинка (десмосома); 5 - тело обонятельной нейросенсорной клетки; 6 -
поддерживающая клетка; 7 - эндоплазматическая сеть; 8 - базальная
мембрана; 9 - аксоны обонятельных нейросенсорных клеток, образующие
обонятельные нити (по В.Г. Елисееву и др.)
биполярные нейроны, имеющие два отростка: длинный центральный (аксон) и короткий периферический (дендрит). Округлые светлые ядра этих клеток, содержащие одно или два ядрышка, расположены в нижней части эпителиального слоя. Базальная часть клетки, суживаясь, переходит в длинный узкий центральный отросток диаметром около 0,1 мкм, который прободает базальную мембрану и, соединяясь с аксонами других обонятельных клеток, формирует безмиелиновые обонятельные нервы. Тонкий дендрит направляется к поверхности эпителия, где заканчивается утолщением - дендритической луковицей (обонятельная булава). От боковых поверхностей этих луковиц отходит по 10-15 обонятельных ресничек. Каждая ресничка содержит девять пар периферических микротрубочек и две центральные микротрубочки, отходящие от базальных телец. Периферическая часть каждой реснички тоньше и содержит 2-3 единичные микротрубочки. Реснички погружены в слизь, покрывающую эпителиальный пласт. Под очень тонкой базальной мембраной расположена собственная пластинка слизистой оболочки, образованная рыхлой волокнистой соединительной тканью, богатой коллагеновыми и эластическими волокнами, среди которых проходит много кровеносных сосудов. В собственной пластинке слизистой оболочки залегают начальные отделы смешанных белково-слизистых (боуменовых) желез, выделяющих секрет, богатый мукополисахаридами. Гландулоциты секреторных отделов этих желез окружены миоэпителиоцитами. Молекулы пахучих веществ, распределенные в секрете обонятельных желез, взаимодействуют с рецепторными белками цитолеммы ресничек нейросекреторных обонятельных клеток, что вызывает нервный импульс. Этот импульс по обонятельным нервам передается к обонятельным луковицам головного мозга, где залегают митральные клетки (рис. 152). Около 1000 аксонов нейросенсорных клеток заканчиваются на дендритах одного митрального нейрона обонятельной луковицы, образуя обонятельные клубочки. Аксоны этих клеток образуют обонятельный тракт и направляются в обонятельный треугольник, а затем в составе промежуточной и медиальной обонятельных полосок вступают в переднее продырявленное вещество, в подмозолистое поле и в диагональную полоску (Брока). В составе латеральной полоски отростки митральных клеток следуют в парагиппокампальную извилину и в крючок, где находится корковый центр обоняния. Обонятельный анализатор связан с лимбической системой, поэтому обонятельные ощущения имеют эмоциональную окраску.
РАЗВИТИЕ ОРГАНА ОБОНЯНИЯ В ОНТОГЕНЕЗЕ
Орган обоняния в онтогенезе человека закладывается в эмбриональной нервной пластинке на границе с эктодермой. Периферическая часть будущего обонятельного органа отделена от зачатка центральной нервной системы и имеет вид парных обонятельных ямок. Затем периферический обонятельный орган вторично соединяется с центральными частями анализатора при помощи обонятельного нерва (нитей). Клетки обонятельной ямки дифференцируются в нейроглиальные, опорные и обонятельные.
У новорожденного обонятельная область слизистой оболочки лишена пигмента, поэтому имеет розовый цвет. Пигмент липофусцин появляется в клетках на шестом году жизни. К моменту рождения обонятельные нейросенсорные клетки и обонятельные нервы развиты достаточно хорошо.
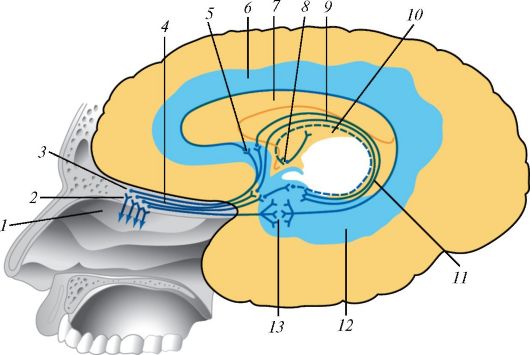 Рис. 152. Проводящий путь органа обоняния:
Рис. 152. Проводящий путь органа обоняния:
1 - верхняя носовая раковина; 2 - обонятельные нервы (I ); 3 - обонятельная луковица; 4 - обонятельный тракт; 5 - подмозолистое поле; 6 - поясная извилина; 7 - мозолистое тело; 8 - сосцевидное тело; 9 - свод; 10 - задний таламус;
11 - зубчатая извилина; 12 - парагиппокампальная извилина (извилина
гиппокампа); 13 - крючок
ОРГАН ВКУСА
Орган вкуса (orgdnum gustus), вкусовые почки имеются в стенках ротовой полости, главным образом в слизистой оболочке языка и неба. Вкусовые почки (calliculi gustatorii) имеют эктодермальное происхождение, развиваются они из элементов эмбрионального многослойного эпителия сосочков языка. Уже в периоде образования вкусовые почки связаны с окончаниями соответствующих нервов (язычный, языкоглоточный, блуждающий). Зачатки вкусовых почек вдаются в подлежащий эпителий сосочков и постепенно принимают вид луковиц.
Орган вкуса у человека представлен множеством (около 2000-3000) вкусовых почек, расположенных в многослойном эпителии боковых поверхностей желобовидных, листовидных и грибовидных сосочков языка, а также в слизистой оболочке неба, зева, глотки и надгортанника. В эпителии каждого сосочка, окруженного валом, расположено до 200 вкусовых почек, у остальных - по нескольку почек. Между сосочками, а также в ровиках сосочков, окруженных валом, открываются выводные протоки мелких слюнных желез языка, выделяющих секрет, омывающий вкусовые почки. Вкусовые почки занимают всю толщину эпителиального покрова сосочков языка.
Вкусовые почки имеют эллипсоидную норму, состоят из 20-30 плотно прилежащих друг к другу вкусовых сенсорных и поддерживающих клеток (рис. 153), в основании которых находятся базальные клетки. На вершине каждой вкусовой почки имеется вкусовое отверстие (porus gustatorius - вкусовая пора), которое ведет в маленькую вкусовую ямку, образованную верхушками вкусовых клеток. Большинство клеток проходят через всю вкусовую почку от базальной мембраны до вкусовой
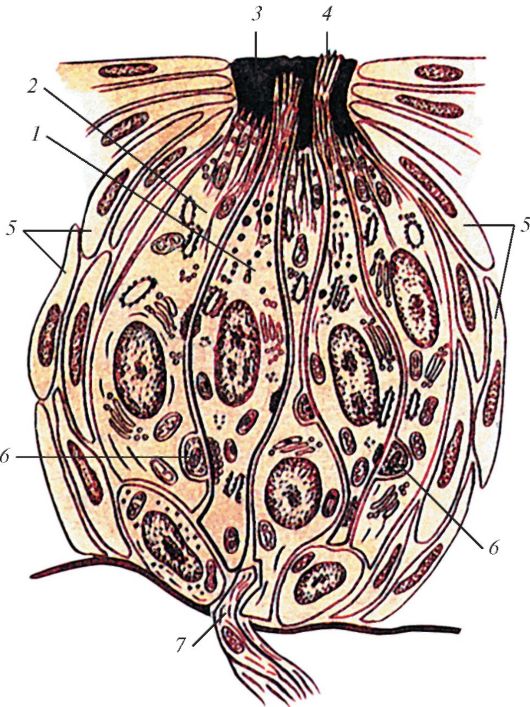 Рис. 153. Строение вкусовой почки (схема): 1
- вкусовая клетка; 2 - поддерживающая клетка; 3 - вкусовая пора; 4 -
микроворсинки; 5 - эпителиальные клетки; 6 - нервное окончание; 7 -
нервное волокно
Рис. 153. Строение вкусовой почки (схема): 1
- вкусовая клетка; 2 - поддерживающая клетка; 3 - вкусовая пора; 4 -
микроворсинки; 5 - эпителиальные клетки; 6 - нервное окончание; 7 -
нервное волокно
ямки, к которой конвергируют апикальные части этих клеток. Мелкие базальные клетки полиэдрической формы, расположенные на базальной мембране по периферии вкусовой почки, не достигают вкусовой ямки. В них обнаруживаются фигуры митоза. Базальные клетки являются стволовыми. Удлиненные вкусовые сенсорные клетки имеют овальное ядро, расположенное в базальной части клеток. Их цитоплазма бедна рибосомами и элементами зернистой эндоплазматической сети, поэтому она выглядит электронно-светлой («светлые» клетки). Различают два типа вкусовых сенсорных клеток. В клетках первого типа находится большое
количество везикул диаметром около 70 нм с электронно-плотной сердцевиной, содержащей катехоламины. В клетках второго типа везикулы отсутствуют. Возможно, они представляют разные стадии дифференцировки сенсорных клеток. На апикальной поверхности каждой вкусовой клетки, обращенной в сторону вкусовой ямки, имеются микроворсинки, вступающие в контакт с растворенными веществами. Большая часть микроворсинок, видимых при световой микроскопии, принадлежит поддерживающим эпителиоцитам, которые окружают сенсорные клетки со всех сторон, кроме апикальной. Продолжительность жизни сенсорных эпителиоцитов не превышает 10 дней. Новые клетки образуются из делящихся базальных клеток.
Среди поддерживающих эпителиоцитов различают клетки двух типов. Удлиненные клетки, лишенные микроворсинок, располагаются по периферии вкусовой почки, отделяя ее от окружающего эпителия. Другие поддерживающие эпителиоциты цилиндрической формы окружают вкусовые сенсорные клетки. Апикальная поверхность поддерживающих клеток покрыта длинными микроворсинками, в которых проходят пучки микрофиламентов. Переплетающиеся между собой микроворсинки клеток обоих типов погружены в электронно-плотное вещество, богатое белками и мукопротеинами с высокой активностью фосфатаз. Нервные окончания лицевого и языкоглоточного нервов образуют множественные синапсы с цитолеммой вкусовых сенсорных клеток.
Растворенные в слюне вкусовые вещества, адсорбированные на гликокаликсе микроворсинок, реагируют с рецепторными белками, встроенными в цитолемму микроворсинок сенсорных клеток. Различают четыре вкусовых ощущения: горькое, соленое, кислое и сладкое. Сенсорные клетки обладают очень высокой чувствительностью. Так, порог восприятия поваренной соли составляет 0,01 моль/л, глюкозы - 0,08 моль/л, сахарозы - 0,01 моль/л, соляной кислоты - 0,0009 моль/л, лимонной кислоты - 0,0023 моль/л, сульфата хинина - 0,000008 моль/л.
На слизистой оболочке языка различают области восприятия вкусовых ощущений, перекрывающих друг друга. Однако горький вкус воспринимается главным образом сосочками основания языка. Одна сенсорная клетка воспринимает несколько вкусовых раздражений.
Взаимодействие молекул с рецепторами вызывает нервный импульс (рецепторный потенциал), который через синапсы передается нервным волокнам. Каждое нервное волокно ответвляется и иннервирует множество
нейросенсорных клеток разных вкусовых почек. Афферентные нервные окончания по-разному реагируют на вкусовые воздействия. Так, многие окончания языкоглоточного нерва особенно сильно возбуждаются под действием горьких веществ, а лицевого нерва - под действием кислых, соленых и сладких. Одни окончания активнее реагируют на соленые вещества, другие - на сладкие. Нервный импульс от передних 2/3 языка передается по нервным волокнам язычного нерва, а затем барабанной струны лицевого нерва. От желобовидных сосочков, мягкого неба и небных дужек импульс идет по волокнам языкоглоточного нерва, от надгортанника - по блуждающему нерву. Тела нейронов этих нервов залегают в соответствующих нервных узлах (чувствительных), их аксоны направляются в составе указанных нервов в чувствительное ядро одиночного пути, расположенное в продолговатом мозге. Центральные отростки клеток этих ядер направляются через медиальную петлю в таламус, в вентральное заднелатеральное ядро. Аксоны этих нейронов идут к корковому концу вкусового анализатора, расположенному в коре парагиппокампальной извилины, крючка и гиппокампа (аммонова рога) (рис. 154).
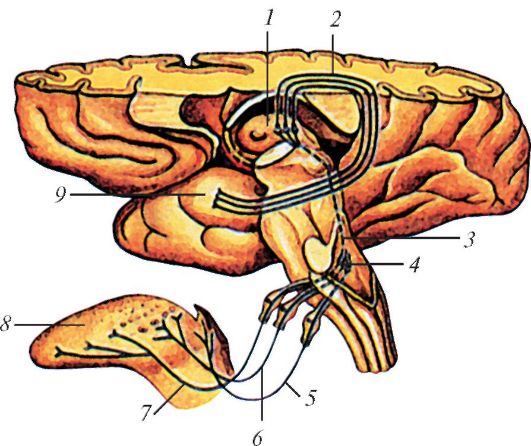 Рис. 154. Проводящий
путь органа вкуса: 1 - таламус; 2 - волокна, соединяющие таламус и
крючок; 3 - волокна, соединяющие ядро одиночного пути и таламус; 4 -
ядро одиночного пути; 5 - вкусовые волокна в составе верхнего гортанного
нерва (блуждающий нерв); 6 - вкусовые волокна в составе языкоглоточного
нерва; 7 - вкусовые волокна в составе
Рис. 154. Проводящий
путь органа вкуса: 1 - таламус; 2 - волокна, соединяющие таламус и
крючок; 3 - волокна, соединяющие ядро одиночного пути и таламус; 4 -
ядро одиночного пути; 5 - вкусовые волокна в составе верхнего гортанного
нерва (блуждающий нерв); 6 - вкусовые волокна в составе языкоглоточного
нерва; 7 - вкусовые волокна в составе
барабанной струны; 8 - язык; 9 - крючок
ОБЩИЙ ПОКРОВ (КОЖА)
Кожа (cutis) выполняет многообразные функции: защитную, терморегуляторную, дыхательную, обменную, рецепторную. Железы кожи вырабатывают пот, кожное сало. С пóтом у человека в течение суток в обычных условиях выделяется около 500 мл воды, а также соли и конечные продукты азотистого обмена. Кожа активно участвует в обмене витаминов. Особенно важен синтез витамина D под влиянием ультрафиолетовых лучей. Кожа является одним из важнейших депо крови - в ней депонируется до 1 л крови. Площадь кожного покрова взрослого человека достигает 1,5-2 м2. Эта поверхность является обширным рецепторным полем тактильной, болевой, температурной кожной чувствительности.
Кожа состоит из эпидермиса, который развивается из эктодермы, и дермы, образующейся из дерматомов (мезодермальное происхождение). Эпидермис (epidermis) - это многослойный плоский ороговевающий эпителий (рис. 155), толщина которого (0,03-1,5 мм) зависит от выполняемой функции. Так, на участках, подвергающихся постоянному механическому давлению (ладони, подошвы), толщина эпидермиса больше, чем на груди, животе, бедре, плече, предплечье, шее. Эпидермис, у которого выделяют базальный, шиповатый, зернистый, блестящий и роговой слои, расположен на базальной мембране (рис. 156). Непосредственно на ней лежит базальный слой, образованный мелкими призматическими клетками, базальная часть которых образует множество отростков, инвагинирующих в базальную мембрану. Базальные клетки (базальные эпидермоциты) прикреплены к базальной мембране с помощью полудесмосом. В углубления базальной мембраны внедряются тонкие ретикулярные и коллагеновые волоконца подлежащей соединительной ткани. В цитоплазме апикальной части многих базальных эпидермоцитов присутствуют
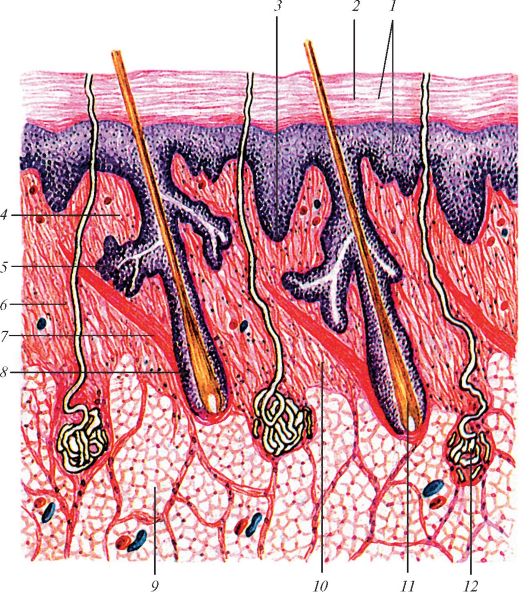 Рис. 155. Строение
кожи: 1 - эпидермис; 2 - роговой слой; 3 - базальный слой; 4 -
сосочковый слой; 5 - сальная железа; 6 - соединительнотканные волокна
(коллагеновые, эластические, ретикулярные) и клетки; 7 - мышца,
поднимающая волос; 8 - волосяная луковица; 9 - дольки жировой ткани; 10 -
сетчатый слой; 11 - корень волоса;
Рис. 155. Строение
кожи: 1 - эпидермис; 2 - роговой слой; 3 - базальный слой; 4 -
сосочковый слой; 5 - сальная железа; 6 - соединительнотканные волокна
(коллагеновые, эластические, ретикулярные) и клетки; 7 - мышца,
поднимающая волос; 8 - волосяная луковица; 9 - дольки жировой ткани; 10 -
сетчатый слой; 11 - корень волоса;
12 - потовая железа
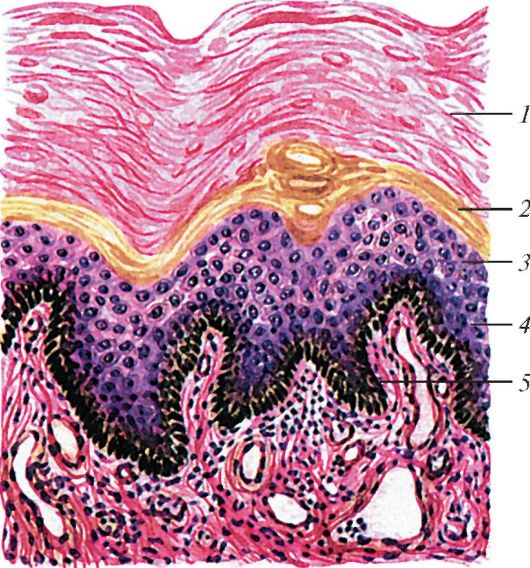 Рис. 156. Строение
эпидермиса: 1 - роговой слой; 2 - блестящий слой; 3 - зернистый слой; 4
- шиповатый слой; 5 - базальный (цилиндрический слой) (по В. Баргману)
Рис. 156. Строение
эпидермиса: 1 - роговой слой; 2 - блестящий слой; 3 - зернистый слой; 4
- шиповатый слой; 5 - базальный (цилиндрический слой) (по В. Баргману)
поглощенные клеткой зерна меланина. Цитоплазма богата рибосомами, тонофиламентами толщиной 75 нм, образующими пучки. Митохондрии и элементы зернистой эндоплазматической сети малочисленны, комплекс Гольджи развит слабо. Тонофиламенты являются ранними предшественниками кератина, который синтезируется рибосомами. Крупное ядро овальной или неправильной формы богато хроматином. Базальные клетки соединены друг с другом и с клетками шиповатого слоя десмосомами. Между клетками имеются широкие извилистые межклеточные щели. Часть базальных клеток стволовые, что подтверждается отсутствием митотически делящихся клеток. Дочерние клетки дифференцируются и перемещаются в вышележащие слои.
Среди базальных клеток имеются пигментные эпителиоциты (меланоциты), богатые зернами пигмента меланина, от содержания которого зависит цвет кожи. Меланин защищает кожу от ультрафиолетовых лучей. Меланоциты, происходящие из ганглиозной пластинки, в эмбриональный период мигрируют в эпидермис и располагаются на базальной
мембране. Это крупные светлые многоотростчатые клетки с круглым ядром, хорошо развитыми органеллами, умеренным количеством свободных рибосом, небольшим числом микрофибрилл и большим числом меланосом, являющихся предшественниками гранул меланина. Зрелые гранулы меланина имеют размеры 0,3-0,7 мкм, овальной формы, окружены мембраной, проникают в отростки меланоцитов. Длинные отростки меланоцитов внедряются между клетками базального слоя эпидермиса. Меланоциты пролиферируют в течение всей жизни человека. Их число достигает 1200-1500 в 1 мм2 эпидермиса. В эпидермисе альбиносов меланоцитов нет - в связи с генетически детерминированным отсутствием фермента тирозиназы образующиеся меланосомы не созревают в гранулы меланина.
Над базальным слоем эпидермиса расположен шиповатый слой, включающий от 2 до 10 слоев полиэдрических клеток. Некоторые из них также митотически делятся. Однако их митотическая активность ниже, чем у клеток базального слоя. В цитоплазме имеются мелкие сферические или овальные мембранные тельца - кератиносомы. Множество отростков этих клеток соединяются между собой с помощью десмосом.
Базальный слой и расположенные в глубине клетки шиповатого слоя функционально объединены в ростковый слой благодаря их способности к митотическому делению и дальнейшей дифференцировке в клетки других слоев.
Выше расположен зернистый слой, состоящий из нескольких слоев уплощенных чешуйчатых эпителиоцитов, содержащих крупные зерна кератогиалина, с сильным лучепреломлением. Эти зерна по мере продвижения клеток в верхние слои эпидермиса превращаются в кератин. В толстой коже (ладони, подошвы) зернистый слой состоит из 2-3 слоев клеток, в тонкой коже (бедра, шея, живот, грудь) клетки на образуют сплошного слоя. Кератогиалин состоит из белка, содержащего большое количество пролина, гистидина, аргинина и цистина, полисахаридов и липидов. Многочисленные тонофиламенты связаны с гранулами кератогиалина. В цитоплазме эпителиоцитов расположены митохондрии, рибосомы, лизосомы (часть лизосом представлена пластинчатыми гранулами).
Над зернистым слоем лежит блестящий слой, который характерен только для толстой кожи, лишенной волос. Он образован 3-4 слоями плоских (чешуйчатых) клеток, не имеющих ядер, богатых белком элеидином, хорошо преломляющим свет. Элеидин образуется из кератогиалина
и белков тонофибрилл. Цитолемма чешуйчатых эпителиоцитов блестящего слоя утолщена (до 11-12 нм).
Поверхностный роговой слой представляет собой множество слоев роговых чешуек, содержащих белок кератинин и пузырьки воздуха. Этот водонепроницаемый слой плотный, упругий и, что особенно важно, через него не проникают микроорганизмы. Особая структура рогового слоя обеспечивает выполнение указанных функций. Кератин в роговых чешуйках образует плотно упакованные кератиновые филаменты толщиной 7-9 нм, погруженные в электронно-плотное вещество, которое, повидимому, происходит из гранул кератогиалина. Роговые чешуйки постоянно слущиваются и заменяются новыми, которые подходят к поверхности из глубжележащих слоев клеток. Эти клетки в процессе миграции на поверхность постепенно ороговевают. Полная смена клеток в эпидермисе человека происходит в течение 30-90 дней.
Дерма, или собственно кожа (derma, s. corium), толщиной 0,5-5 мм, образована соединительной тканью. Толщина дермы варьирует у одного и того же человека, она наибольшая на плечах, спине, бедрах, ягодицах, наименьшая на груди, мошонке. В дерме различают сосочковый и сетчатый слои, которые переходят один в другой без резкой разницы.
Сосочковый слой (stratum papillare) находится под базальной мембраной эпидермиса. Он образован рыхлой волокнистой неоформленной соединительной тканью, которая формирует конические сосочки, внедряющиеся в эпидермис и как бы прогибающие его базальную мембрану. Соединительная ткань состоит из переплетающихся пучков коллагеновых и небольшого числа эластических волокон и богата клеточными элементами, большинство из которых фибробласты. Наряду с фибробластами имеются макрофаги. Плазматические клетки и лимфоциты встречаются реже. Богато снабженный кровеносными сосудами сосочковый слой осуществляет питание эпидермиса, который не имеет сосудов. Капиллярные петли расположены перпендикулярно поверхности эпидермиса, венулы расположены в основании сосочков. В сосочках присутствуют нервные окончания.
Благодаря сосочкам на поверхности кожи видны гребешки, разделенные бороздками. Гребешки, соответствующие возвышениям сосочков дермы, и бороздки между ними формируют строго индивидуальный сложный рисунок кожной поверхности, особенно на ладонях и стопах, сохраняющийся в течение всей жизни человека и нарушающийся при некоторых наследственных заболеваниях (хромосомных аномалиях).
Строение кожного рельефа широко используется в медицине для изучения наследственности человека и в криминалистике для идентификации личности. Изучение деталей рельефа кожи (папиллярных линий и узоров) получило название дерматоглифики (от греч. glypho - вырезаю, гравирую).
В сосочковом слое имеются миоциты, связанные с волосяными луковицами. В дерме лица, мошонки, соска молочной железы, тыльной поверхности конечностей имеются самостоятельные пучки миоцитов, не связанные с луковицами волос. При их сокращении возникает так называемая гусиная кожа.
Под сосочковым слоем находится сетчатый слой (stratum reticulare), который состоит из плотной неоформленной соединительной ткани, содержащей крупные пучки коллагеновых волокон, расположенных под углом друг к другу и образующих сеть. Одни пучки лежат параллельно кожной поверхности, другие проходят косо. В дерме тех областей, которые при движениях растягиваются (например, над суставами), ячейки этой сети узкопетлистые. На стопе, локтях, концевых фалангах пальцев, подвергающихся постоянному давлению, ячейки сети широкопетлистые. Наряду с коллагеновыми волокнами в сетчатом слое имеется сеть эластических и небольшое количество ретикулярных волокон. Количество клеток невелико. Это, в основном, фиброциты. В сетчатом слое залегают корни волос, потовые и сальные железы.
Пучки коллагеновых волокон сетчатого слоя проходят в подкожную клетчатку (tela subcutanea), содержащую жировую ткань. Этот слой играет важную роль в терморегуляции и является жировым депо организма. Жировая ткань достигает наибольшего развития в области ягодиц и подошв, где выполняет механическую функцию. У век, мошонки жировой слой отсутствует. Как правило, жировой слой больше развит у женщин.
Волосы (pili) являются производным эпидермиса. Почти вся кожа покрыта волосами. Исключение составляют ладони, подошвы, переходная часть губ, головка полового члена и малые половые губы. Наибольшее число волос обычно бывает на голове. Характер оволосения зависит от пола и возраста, и относится ко вторичным половым признакам. В период полового созревания начинается усиленный рост волос в подмышечных впадинах, на лобке, у мужчин также на лице, конечностях, груди, животе. Различают волосы трех типов - длинные, щетинистые и пушковые. Длинные волосы покрывают голову, лобок и подмышечные
впадины; щетинистые принадлежат бровям, ресницам, преддверию полости носа и наружному слуховому проходу; пушковые волосы располагаются на остальных поверхностях тела.
Волос имеет выступающий над поверхностью кожи стержень (scapus) и корень (radix), лежащий в толще кожи (рис. 157). Длина стержня колеблется от 1-2 мм до 1,5 м, а толщина - от 0,005 до 0,6 мм. Корень волоса находится в волосяном мешке (фолликуле - folliculus pili), образованном эпителиальным (корневым) влагалищем и соединительнотканной сумкой волоса (bursa рili). К сумке прикрепляется мышца-поднима- тель волоса (m. arrector pili). Волосы на подбородке и в области лобка лишены этой мышцы. В сумку волоса открывается сальная железа. Сокращаясь, мышца поднимает волос, сдавливает сальную железу, благодаря чему выделяется ее секрет. Наружное корневое влагалище продолжается в эпидермис. В области сосочка волоса оно истончается, в нем остается лишь ростковый слой, окружающий сосочек. Внутреннее корневое влагалище, расположенное между волосом и наружным корневым влагалищем, образовано эпителиальными клетками, которые окружают корень волоса наподобие муфты.
Корень волоса переходит в расширенную волосяную луковицу (bulbus pili), в которую впячивается соединительнотканный сосочек волоса, богатый кровеносными капиллярами, питающими луковицу. Над сосочком расположен матрикс - ростковая часть волоса. В результате деления клеток волос растет. Между эпителиоцитами матрикса залегают меланоциты, синтезирующие пигмент меланин. Как и в клетках эпидермиса, зерна меланина выделяются отростками меланоцитов, захватываются эпителиальными клетками, где по мере ороговения меланин входит в состав кератина, окрашивая тем самым волос.
Стержень волоса состоит из мозгового и коркового вещества, которое преобладает. Корковое вещество (кора волоса) образовано плоскими роговыми чешуйками, заполненными, в основном, кератином. Кроме того, в этих чешуйках содержатся зерна пигмента и пузырьки воздуха. Клетки мозгового вещества лежат друг на друге, они богаты трихогиалином, который превращается в кератин, а также содержат пузырьки воздуха и зерна пигмента. С возрастом количество пузырьков воздуха увеличивается, а синтез пигмента постепенно прекращается, волосы седеют. Корковое вещество снаружи покрыто кутикулой, образованной плоскими кутикулярными клетками. Волосы сменяются в течение 2-3 мес - 2-3 лет.
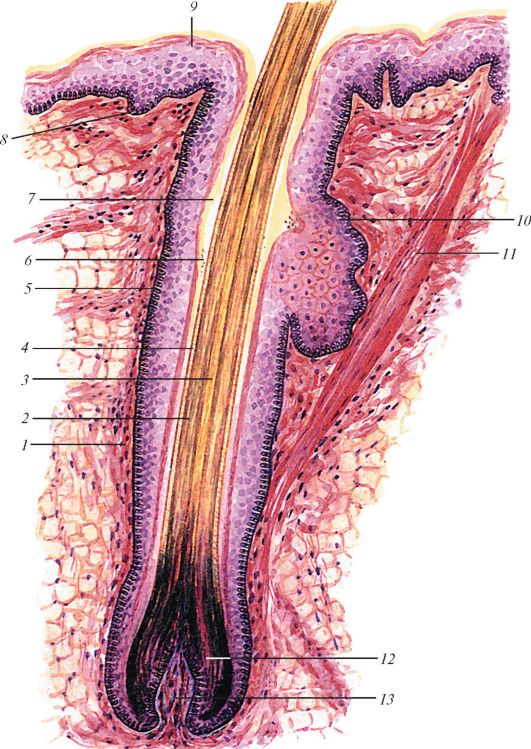 Рис. 157. Схема
строения волоса: 1 - волосяная сумка; 2 - кора волоса; 3 - мозговое
вещество волоса; 4 - кутикула; 5 - наружное корневое влагалище; 6 - два
слоя внутреннего корневого влагалища; 7 - волосяная воронка; 8 -
базальный (ростковый) слой эпидермиса; 9 - роговой слой эпидермиса; 10 -
сальная железа; 11 - мышца, поднимающая волос; 12 - луковица волоса; 13
- сосочек волоса (по В.Г. Елисееву и др.).
Рис. 157. Схема
строения волоса: 1 - волосяная сумка; 2 - кора волоса; 3 - мозговое
вещество волоса; 4 - кутикула; 5 - наружное корневое влагалище; 6 - два
слоя внутреннего корневого влагалища; 7 - волосяная воронка; 8 -
базальный (ростковый) слой эпидермиса; 9 - роговой слой эпидермиса; 10 -
сальная железа; 11 - мышца, поднимающая волос; 12 - луковица волоса; 13
- сосочек волоса (по В.Г. Елисееву и др.).
Ноготь (unguis). Ногти, как и волосы, являются производными эпидермиса кожи. Ноготь представляет собой роговую пластинку, лежащую на соединительнотканном ложе ногтя (matrix unguis), ограниченном у основания и с боков ногтевыми валиками. Ноготь впячивается в щели, расположенные между ложем и валиками. В задней ногтевой щели залегает корень ногтя (radix unguis). Тело ногтя (corpus unguis) лежит на ногтевом ложе, а свободный край (margo liber) выступает за его пределы. Ноготь растет в результате деления клеток росткового слоя ногтя - эпителия ногтевого ложа в области корня. Делящиеся клетки, подобно эпителиоцитам эпидермиса, продвигаясь вперед, ороговевают.
Железы кожи. К ним относятся потовые, сальные и молочные железы. Потовых желез (glandula sudorifera) около 2-2,5 млн, они представляют собой простые трубчатые железы. Различают мерокриновые и апокриновые потовые железы.
Секреторные отделы мерокриновых желез закручиваются, образуя клубочки. Секреторные отделы сформированы лежащими на базальной мембране экзокриноцитами, среди которых различают темные и светлые клетки. Темные экзокриноциты вырабатывают мукоид. Их цитоплазма богата рибосомами и секреторными гранулами, содержащими мукопротеины и мукополисахариды. Светлые экзокриноциты бедны рибосомами, но в них много частичек гликогена. Между двумя соседними светлыми клетками имеются широкие межклеточные щели, куда проникают узкие цитоплазматические отростки клеток, которые переплетаются между собой. Светлые клетки выделяют воду и ионы. Кнаружи от экзокриноцитов на базальной мембране спирально располагаются богатые микрофиламентами веретенообразные миоэпителиоциты. Длинный выводной проток, извиваясь, прободает кожу и открывается на поверхности потовой порой. Выводной проток выстлан 2-3 слоями мелких кубических клеток. Апикальные отделы поверхностных клеток содержат большое количество тонофиламентов, рибосом и частичек гликогена. Эта зона при световой микроскопии определяется как кутикула. Она образована множеством мелких плотно прилежащих
друг к другу микроворсинок, через которые проходят микрофиламенты, спускающиеся в апикальную часть клетки. Эпителий протока лежит на базальной мембране.
Апокриновые потовые железы развиваются лишь в период полового созревания в коже лба, лобка, больших половых губ, окружности заднего прохода, подмышечных ямок. Их секрет содержит больше белковых веществ, которые при разложении дают специфический запах. Секреторные отделы апокриновых желез крупнее, чем мерокриновых. Они представляют собой извитые, иногда разветвленные трубочки, образованные одним слоем кубических или цилиндрических эпителиоцитов, апикальные части которых выбухают в просвете железы. В отличие от мерокриновых, у апокриновых желез обнаружены только темные клетки. Круглое ядро расположено в центре клетки, цитоплазма богата крупными митохондриями, вторичными лизосомами, включая гранулы липофусцина, секреторными гранулами. Апикальная поверхность клетки покрыта большим количеством мелких выростов и микроворсинок. Экзокриноциты окружены веретеновидными миоэпителиоцитами, которые лежат на базальной мембране. Потовые протоки прободают эпидермис и открываются на его поверхности вблизи волосяных фолликулов. Потовые протоки выстланы многослойным кубическим эпителием, лежащим на базальной мембране. Миоэпителиоциты отсутствуют. При испарении пота теплоотдача увеличивается, что составляет один из важных механизмов терморегуляции.
Сальные железы (glandulae sebaceae), простые альвеолярные, располагаются на границе между сосочковым и сетчатым слоями дермы. Сальные железы отсутствуют лишь на ладонях и подошвах, наибольшее их количество на голове, лбу, щеках, подбородке. Общая масса выделяемого железами кожного сала может достигать 20 г/сут. Железа состоит из альвеолярного секреторного отдела диаметром 0,2 - 2,0 мм и короткого выводного протока, который открывается в волосяной мешочек (сальная железа волоса). На участках кожи, лишенных волос (головка полового члена, переходная часть губы), протоки открываются на поверхность кожи (свободная сальная железа). Секреторные отделы образованы малодифференцированными себоцитами в состоянии жирового перерождения. Малодифференцированные клетки, располагающиеся непосредственно на базальной мембране, митотически делятся и, постепенно обогащаясь каплями жира, передвигаются в сторону выводного протока. Себоциты богаты элементами гладкой эндоплазматической сети
и липидными каплями. Постепенно ядра становятся пикнотическими, связи между клетками нарушаются, мембраны эндоплазматической сети, цитолемма и митохондрии разрушаются. Клетки, насыщенные жиром, гибнут, образуя кожное сало. Кожное сало имеет бактерицидные свойства и не только смазывает волосы и эпидермис, но и в известной мере предохраняет его от микроорганизмов. В период полового созревания функция сальных желез активизируется, что связано с влиянием половых гормонов.
КРОВОСНАБЖЕНИЕ КОЖИ
Кожа обильно кровоснабжается ветвями регионарных артерий. Кожные артерии отходят от фасциальной артериальной сети. На границе между кожной и подкожной основой артерии формируют подкожное сплетение, снабжающее железы и волосы. От этой сети отходят ветви, направляющиеся к сосочковому слою. В основании сосочков они образуют подсосочковую сеть, от которой отходят веточки, распадающейся на капилляры, питающие сосочки, а также веточки к корням волос и сальным железам. Венозная кровь оттекает в два сосочковых венозных сплетения, а из них - в кожное венозное сплетение, расположенное между кожей и подкожной основой. Сюда же оттекает кровь и от потовых желез. Из кожного венозного сплетения кровь оттекает в фасциальное сплетение, из которого начинаются крупные вены.
Лимфа от кожи оттекает в поверхностное и глубокое лимфатические сплетения, расположенные в глубоких слоях кожи (в подкожной основе).
МОЛОЧНАЯ ЖЕЛЕЗА
Молочная железа (mamma) является измененной потовой железой. Она расположена на передней поверхности грудной мышцы (рис. 158). У девственниц ее масса составляет примерно 150-200 г, у кормящей женщины - 300-400 г. У мужчин железа недоразвита. На передней поверхности железы находится гиперпигментированный сосок (papilla mammaria), окруженный пигментированным околососковым кружком (areola mammae) (рис. 159). На поверхности соска открываются 10-15 точечных отверстий - млечных пор. В коже соска и околососкового кружка множество миоцитов, при их сокращении сосок напрягается (становится
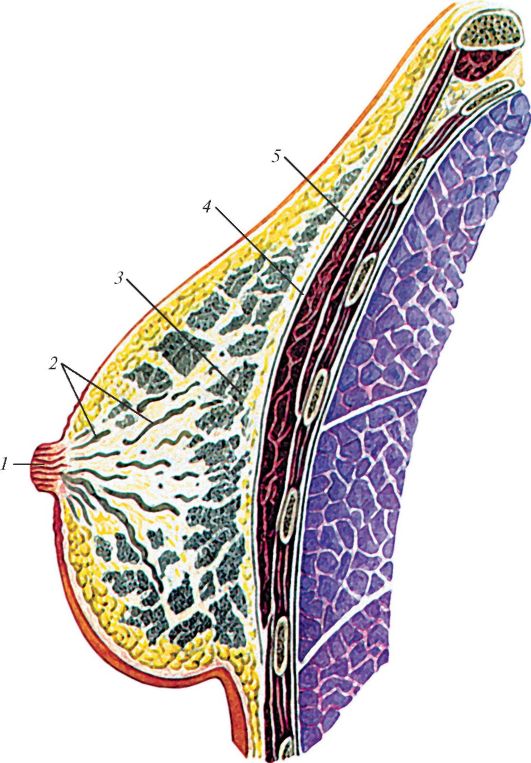 Рис. 158. Молочная
(грудная) железа, сагиттальный разрез: 1 - сосок молочной железы; 2 -
млечные протоки; 3 - тело молочной железы; 4 - грудная фасция; 5 -
большая грудная мышца
Рис. 158. Молочная
(грудная) железа, сагиттальный разрез: 1 - сосок молочной железы; 2 -
млечные протоки; 3 - тело молочной железы; 4 - грудная фасция; 5 -
большая грудная мышца
 Рис. 159. Молочная
(грудная) железа, вид спереди: 1 - тело молочной железы; 2 -
околососковый кружок молочной железы; 3 - сосок молочной железы
Рис. 159. Молочная
(грудная) железа, вид спереди: 1 - тело молочной железы; 2 -
околососковый кружок молочной железы; 3 - сосок молочной железы
упругим). На неровной коже кружка видны бугорки, на поверхности которых открываются протоки потовых желез околососкового кружка (железы Монтгомери), рядом с ними располагаются сальные железы. У взрослой женщины железа состоит из 15-20 долей, между которыми располагается жировая и рыхлая волокнистая соединительная ткань. Доли делятся междольковыми перегородками на дольки. Система протоков начинается с мелких внутридольковых, которые вливаются в междольковые. Последние, сливаясь, образуют 15-20 млечных протоков (ductuli lactiferi), которые направляются радиально к соску и открываются на его поверхности. Не доходя до соска, каждый проток расширяется, образуя млечный синус (sinus lactiferi). Этим синусам отводится роль небольших резервуаров для молока.
Начальные отделы желез представляют собой слепые млечные альвеолярные протоки, выстланные однорядным кубическим эпителием,
лежащим на базальной мембране. Местами между эпителиоцитами и базальной мембраной расположены миоэпителиоциты. Млечные протоки выстланы многослойным эпителием, состоящим из кубических и призматических клеток, которые вблизи соска сменяются многослойным плоским эпителием.
У беременной женщины железа пролиферирует. У внутридольковых млечных протоков растут новые протоки, на которых формируются альвеолы, находящиеся в спавшемся состоянии. Альвеолы образованы одним слоем цилиндрических клеток, лежащих на базальной мембране. Перед родами клетки секретируют молозиво, образуемая ими жидкость растягивает альвеолы, в результате чего железа продолжает набухать. Этот процесс продолжается и в первые 1-2 дня после родов. Молозиво - богатая белком беловатая жидкость с небольшим содержанием жира. Она содержит молозивные тельца, которые представляют собой макрофагоциты, заполненные липидными каплями.
В период кормления альвеолы молочных желез продуцируют молоко. Альвеолы образованы кубическими клетками - лактоцитами, лежащими на базальной мембране. Лактоциты богаты элементами зернистой эндоплазматической сети и комплекса Гольджи, свободными рибосомами. От мешочков комплекса Гольджи отщепляются мембранные пузырьки, содержащие белковые частицы. Эти пузырьки движутся к поверхности клетки, сливаясь с цитолеммой, после чего их содержимое выделяется в просвет альвеолы (секреция белка по мерокриновому типу) и входит в состав молока. Липидные капли выделяются по апокриновому типу. Лактоциты окружены корзинчатыми миоэпителиоцитами, расположенными на базальной мембране. Их сокращение приводит к выдавливанию молока в протоки. Секреция молока стимулируется лактотропным гормоном гипофиза. После окончания кормления ребенка постепенно происходит обратное развитие молочной железы, сохраняются лишь некоторые альвеолы.
КРОВОСНАБЖЕНИЕ И ИННЕРВАЦИЯ МОЛОЧНОЙ ЖЕЛЕЗЫ
К молочной железе подходят ветви из III-VII задних межреберных артерий, из III-V прободающих ветвей внутренней грудной артерии. Артерии разветвляются в соединительнотканных прослойках. Развитые капиллярные сети окружают альвеолы функционирующей железы. Капилляры собираются в венулы, которые вливаются в вены, проходящие
в соединительнотканных прослойках. Глубокие вены прилежат к одноименным артериям, поверхностные располагаются под кожей, где образуют широкопетлистое сплетение.
Лимфатические сосуды из молочной железы направляются к подмышечным, окологрудинным (своей и противоположной стороны), глубоким нижним шейным (надключичным) лимфатическим узлам. Чувствительная иннервация железы (кожи) осуществляется ветвями межреберных нервов (II-VI), надключичных нервов (из шейного сплетения). Вместе с чувствительными нервами с кровеносными сосудами в железу проникают секреторные (симпатические) волокна.
РАЗВИТИЕ И ВОЗРАСТНЫЕ ОСОБЕННОСТИ
МОЛОЧНОЙ ЖЕЛЕЗЫ
Молочные железы закладываются у зародыша длиной около 10 см в виде двух продольных уплотненных эпидермальных молочных линий, проходящих вдоль всего туловища. От них в глубь мезенхимы врастают эпителиальные тяжи, которые дают начало железе. У новорожденной девочки секреторные отделы почти не развиты, имеется лишь недоразвитая система протоков. В препубератном периоде жировая ткань быстро растет. К началу половой зрелости железа становится округлой, но ее увеличение происходит в основном за счет жировой ткани.
Молочная железа девочки начинает развиваться во время полового созревания под влиянием женских половых гормонов (эстрогенов). После наступления беременности под влиянием эстрогенов, прогестерона, пролактина и плацентарного лактогена начинается развитие альвеол молочной железы. Концентрация пролактина в цитоплазме эпителиоцитов беременной женщины резко возрастает (до 300 мкг/л, у небеременной - 2-15 мкг/л). Главную роль играет пролактин, который стимулирует образование альвеол, рост желез и секрецию молока. Окситоцин вызывает сокращение звездчатых миоэпителиоцитов, оплетающих лактоциты, в результате чего выделяется молоко. При сосании возбуждаются рецепторы соска и околососкового поля, нервный импульс достигает нейросекреторных клеток супраоптического ядра гипоталамуса, секрет которых усиливают синтез и секрецию окситоцина. Таким образом, в выделении молока участвуют нервный и гуморальный компоненты. Обратное развитие железы и замещение ее жировой тканью происходят в климактерическом периоде.
Иногда встречается недоразвитие одной или обеих желез, появляются добавочные (кроме одной пары) железы (полимастия) или только добавочные соски. У мужчин иногда железы развиваются по женскому типу (гинекомастия).
КОЖНАЯ ЧУВСТВИТЕЛЬНОСТЬ
Кожа иннервируется чувствительными нервами, отходящими от спинномозговых и черепных нервов, а также волокнами вегетативных нервов, иннервирующих сосуды, гладкие глиоциты и железы. Густое нервное сплетение залегает в подкожной основе. От него отходят ветви, формирующие сплетения вокруг корней волос, желез, жировых долек, в сосочковом слое кожи. От последнего отходят волокна, иннервирующие сосочковый и сетчатый слои, а также эпидермис.
Кожа богато снабжена рецепторами. В зависимости от характера раздражителя различают терморецепторы, механорецепторы и ноцирецепторы. Первые воспринимают изменения температуры, вторые - прикосновение к коже, ее сдавление, третьи - болевые раздражения. Афферентные нервные волокна, по которым распространяются импульсы от указанных рецепторов, являются дендритами чувствительных псевдоуниполярных клеток, залегающих в спинномозговых узлах и чувствительных узлах черепных нервов.
Осязание (механорецепция) - это способность ощущать давление, прикосновение, вибрацию, которые воспринимаются лишь в определенных осязательных точках кожи. Плотность осязательных точек наибольшая в коже губ и подушечек пальцев, наименьшая - в коже спины, плеч, бедра. Современные исследования позволили дифференцировать функцию рецепторов кожи. Так, тельца Фатера-Пачини являются рецепторами вибрации, тельца Мейснера - прикосновения (тактильными), диски Меркеля, тактильные тельца Пинкуса-Игго и тельца Руффини - давления. Ко всем этим рецепторам подходят миелиновые нервные волокна диаметром 5-10 мкм, проводящие нервные импульсы со скоростью 30-70 м/с. Строение и положение рецепторов на участках кожи, покрытых волосами и лишенных волос, различны.
В коже человека преобладают рецепторы прикосновения. По-видимому, каждый отдельный рецептор воспринимает определенное осязательное ощущение, но при воздействии на кожу различных механических
стимулов одновременно реагируют рецепторы нескольких типов. Терморецепторы воспринимают температурные раздражения, причем различные точки кожи воспринимают тепло или холод, последние преобладают. Наиболее чувствительна к температурным раздражителям кожа лица, на 1 см2 которой приходится от 16 до 19 холодовых точек. На 10 см2 кожи кисти приходится 10-15 холодовых и 3-4 тепловые точки, морфологически терморецепторы окончательно не идентифицированы. Свободные нервные окончания безмиелиновых нервных волокон считают терморецепторами. Однако известно, что свободные окончания тонких миелинизированных волокон диаметром менее 3 мкм со скоростью проведения нервного импульса 10-25 м/с также воспринимают холод. Эти рецепторы чувствительны только к температуре, причем каждое афферентное волокно иннервирует одну или несколько температурных точек.
Болевых точек кожи значительно больше, чем тактильных (примерно в 9 раз) и температурных (примерно в 10 раз). Болевые ощущения воспринимаются ноцирецепторами. Это свободные нервные окончания, которые функционально подразделяются на механочувствительные, термочувствительные и полимодальные (механо- и термочувствительные). Последние в коже преобладают. Ноцирецепторы являются окончаниями тонких миелиновых нервных волокон диаметром около 3 мкм со средней скоростью проведения импульса от 10 до 25 м/с и безмиелиновых нервных волокон со скоростью проведения импульса около 1 м/с.