Анатомия человека : Сапин М. Р., Билич Г. Л. учебник в 3 т. - изд. 3-е испр. 2007. - Т. 1, Т. 2, Т. 3.
|
|
|
|
Эндокринные железы анатомически и топографически разобщены (рис. 118), имеют различное происхождение. Они не имеют выводных протоков и выделяют вырабатываемые ими гормоны (от греч. hormao - побуждаю, привожу в движение), или инкреты, непосредственно в кровь и тканевую жидкость. Некоторые гормоны вырабатываются не отдельными железами, а группами компактно расположенных клеток (панкреатические островки, интерстициальные эндокриноциты яичка) или клеток, расположенных в различных органах и тканях (клетки APUD- системы).
Гормоны отличаются от других биологически активных веществ рядом свойств: 1) их действие носит дистантный характер, иными словами,
Таблица 13. Классификация эндокринных желез в зависимости от их происхождения (по А.А. Заварзину и С.И. Щелкунову, с изменениями)
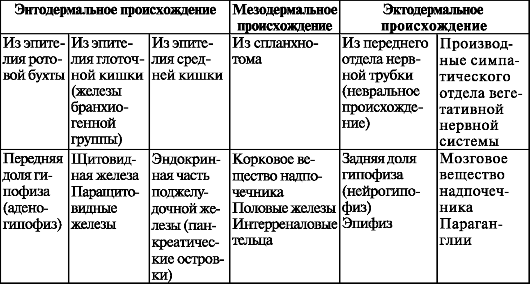
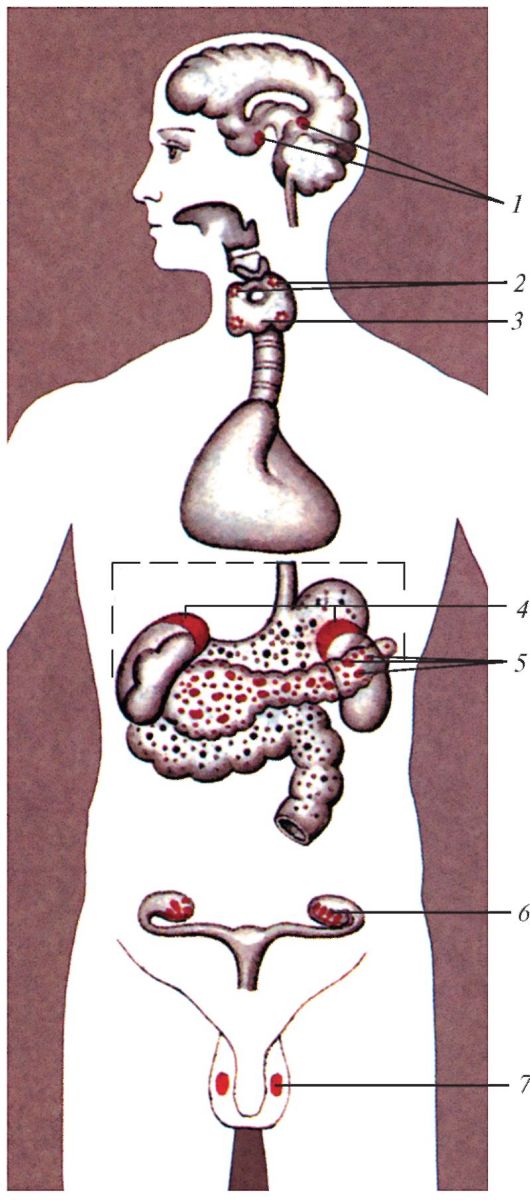
Рис. 118. Положение эндокринных желез в теле человека: 1 - гипофиз и эпифиз; 2 - паращитовидные железы; 3 - щитовидная железа; 4 - надпочечники; 5 - панкреатические островки; 6 - яичник; 7 - яичко
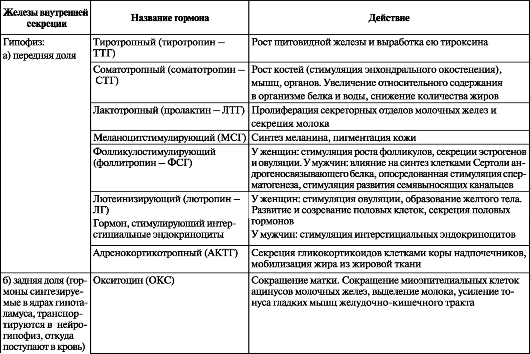
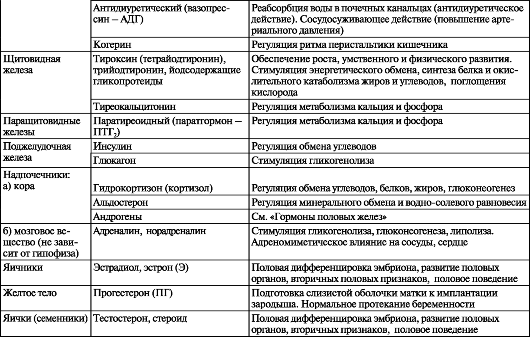
Таблица 14. Эндокринные железы и их гормоны
Окончаание таблицы 14.
органы, на которые гормоны действуют, расположены далеко от железы; 2) действие гормонов строго специфично, некоторые гормоны действуют лишь на определенные клетки-мишени, другие - на множество различных клеток; 3) гормоны обладают высокой биологической актив- ностью и присутствуют в очень малых концентрациях; 4) гормоны действуют только на живые клетки.
Гормоны контролируют жизнедеятельность организма, все функции клеток, активность генов, формирование клеточного фенотипа. Регулируя активность ферментов, гормоны влияют на метаболизм.
Все многообразие действия гормонов можно свести к трем важнейшим функциям: обеспечение роста и развития организма, обеспечение адаптации организма к постоянно меняющимся условиям среды, обес- печение гомеостаза.
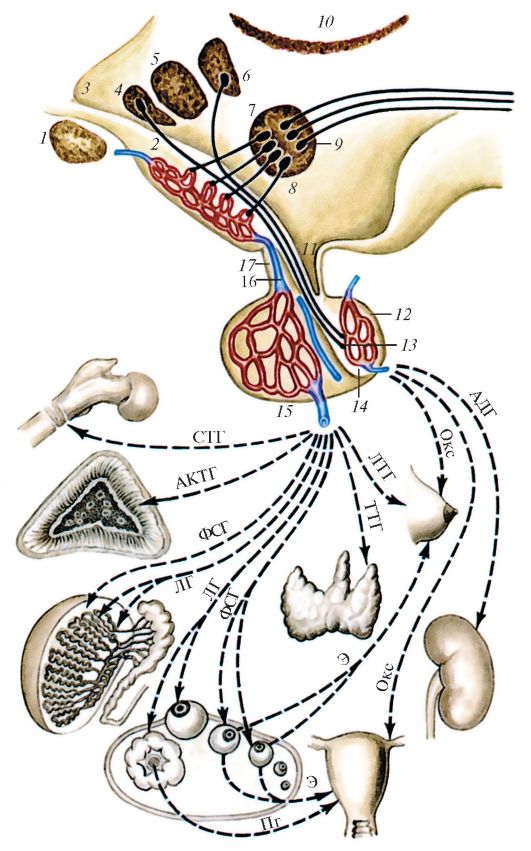
Рис. 119. Схема взаимовлияния органов гипоталамо-гипофизарной системы:
1 - зрительный перекрест; 2 - срединное возвышение с первичной капиллярной сетью; 3 - полость III желудочка; 4 - 7 - проекции некоторых ядер гипоталамуса на стенку III желудочка [4, 5, 6 - соответственно ядра супраоптическое, переднее (преоптическая зона гипоталамуса), паравентрикулярное; 7 - аркуатовентромедиальный комплекс медиобазального гипоталамуса]; 8 - гипоталамус; 9 - адренергические нейроны медиобазального гипоталамуса, дающие начало нисходящим эфферентным нервным путям; 10 - нейросекреторные пептидадренергические клетки медиобазального гипоталамуса, секретирующие адреногипофизарные гормоны в первичную капиллярную сеть срединного возвышения;
11 - углубление воронки III желудочка и гипофизарная ножка; 12 - задняя доля гипофиза; 13 - накопительные тельца Херринга (окончания аксонов нейросекреторных клеток переднего гипоталамуса - супраоптического и паравентрикулярного ядер на капиллярах задней доли гипофиза); 14 - промежуточная (средняя) доля гипофиза; 15 - передняя доля гипофиза со вторичной капиллярной
сетью; 16 - воротная вена гипофиза; 17 - туберальная часть аденогипофиза. Гормоны гипофиза и направление их действия: АДГ стимулирует реабсорбцию воды из первичной мочи в почках (уменьшает диурез) и одновременно повышает артериальное давление крови; ОКС вызывает сокращение матки и отдачу молока молочными железами; ЛТГ активирует выработку молока в молочных железах; ТТГ активирует продукцию и секрецию тиреоидного гормона щитовидной железой; ФСГ активирует рост фолликулов в яичниках и выработку ими эстрогенов, стимулирует сперматогенез в семенниках; ЛГ стимулирует овуляцию, образование желтого тела и продукцию в нем прогестерона, продукцию тестостерона в семенниках; АКТГ стимулирует функцию клеток пучковой и сетчатой зон коры надпочечников; СТГ стимулирует рост организма в целом и его отдельных органов (в том числе скелета); Э устанавливает половые циклы; Пг влияет на преобразование эндометрия в лютеиновой фазе менструального цикла (по Б.В. Алешину; В.Г. Елисееву и соавт.).
В зависимости от происхождения эндокринные железы подразделяются на три группы (табл. 13).
В настоящее время эндокринные железы делят на зависимые и независимые от передней доли гипофиза. К первым относятся щитовидная железа, надпочечник (корковое вещество), половые железы. Другие железы (паращитовидная, эпифиз, панкреатические островки, мозговое вещество надпочечника, параганглии) не подчинены непосредственному влиянию передней доли гипофиза. Взаимоотношения между передней долей гипофиза и независимыми от нее железами строятся по типу прямых и обратных связей. Тропные гормоны (от греч. tropos - направление) пе- редней доли гипофиза активируют деятельность гипофизозависимых желез, а гормоны последних, в свою очередь, воздействуют на гипофиз, угнетая образование и выделение соответствующего тропного гормона. Эндокринные железы и их гормоны представлены в табл. 14.
Центром регуляции эндокринных функций является гипоталамус промежуточного мозга, который координирует нервные и гормональные механизмы регуляции функций внутренних органов, объединяет нервные и эндокринные регуляторные механизмы в общую нейроэндокринную систему. Гипоталамус образует с гипофизом единый функциональный комплекс, в котором первый играет регулирующую, а второй - эффекторную роль. Гипоталамус и гипофиз образуют единую гипоталамо-гипофизарную систему (рис. 119), которая включает такие подсистемы, как гипоталамус-нейрогипофиз (задняя доля гипофиза) и гипотала- мус-аденогипофиз (передняя доля гипофиза).
В гипоталамусе имеются нейроны обычного типа и нейросекреторные клетки. И те и другие вырабатывают белковые секреты и медиаторы, но в нейросекреторных клетках синтез белков преобладает, а нейросекрет выделяется в лимфу и кровь. Нейросекреторные клетки трансформируют нервный импульс в нейрогормональный. Подсистема гипоталамус-ней- рогипофиз функционирует следующим образом: нейроны супраоптического и паравентрикулярного ядер гипоталамуса синтезируют антидиуретический гормон (АДГ) и окситоцин, которые по аксонам транспортируются к капиллярам нейрогипофиза и выделяются в кровь.
В подсистеме гипоталамус-аденогипофиз нейроны мелкоклеточных ядер гипофизотропной зоны гипоталамуса, расположенной в срединном возвышении, выделяют рилизинг-гормоны, или либерины, а также ингибирующие гормоны, или статины, которые транспортируются по аксонам, окончания которых образуют контакты с воротными венами гипофиза. Из этих сосудов гормоны разносятся к клеткам-мишеням аденогипофиза, которые
синтезируют и секретируют тропные гормоны, направляющиеся к соответствующим клеткам-мишеням периферических эндокринных гипофизозависимых желез. Рилизинг-факторы способствуют высвобождению ти- рео-, лютео-, кортикотропина, пролактина, фолликулостимулирующего, соматотропного и меланоцитстимулирующего гормонов. Статины тормозят выделение последних двух гормонов и пролактина.
ГИПОФИЗ
Гипофиз (hypophysis) является важнейшей железой внутренней секреции, которая регулирует деятельность ряда эндокринных желез (щитовидной, половых, коры надпочечников). Гипофиз расположен в гипофизарной ямке турецкого седла клиновидной кости. Отросток твердой мозговой оболочки - диафрагма седла отделяет гипофиз от полости че- репа. Воронка, проходящая через диафрагму седла, соединяет гипофиз с гипоталамусом. Размеры гипофиза 10-17x5 - 15x5-10 мм, его масса у мужчин около 0,5-0,6 г, у женщин - 0,6-0,7 г.
Гипофиз снаружи покрыт соединительнотканной капсулой. Анатомически единый гипофиз делится на две доли - переднюю и заднюю. Передняя доля, или аденогипофиз (adenohypophysis, s. lobus anterior), крупнее задней. Аденогипофиз занимает 70-80% всей массы гипофиза и состоит из дистальной, бугорной и промежуточной частей. Меньшая зад- няя доля, или нейрогипофиз (lobus posterior, s. neurohypophysis), подразделяется на нервную долю и воронку. Между передней и задней долями расположена тонкая промежуточная часть (pars intermedia).
Для понимания функции гипофиза необходимо знать особенности его кровоснабжения. Нижние артерии гипофиза отходят от внутренних сонных артерий, верхние - от сосудов артериального круга большого мозга. Верхние артерии гипофиза направляются к серому бугру и воронке, где анастомозируют между собой и распадаются на капилляры, проникающие в ткань мозга (первичная гемокапиллярная сеть). Своеобразные петли этих капилляров близко подходят к окончаниям аксонов нейросекреторных клеток гипоталамуса. Здесь нейросекрет клеток ядер гипоталамуса выделяется в кровь. Из длинных и коротких петель этой капиллярной сети формируются воротные вены, которые из области бугра и воронки идут вдоль ножки гипофиза к его передней доле. В па- ренхиме передней доли гипофиза воротные вены переходят в широкие синусоидные капилляры, образующие вторичную гемокапиллярную сеть, оплетающую группы секреторных клеток. Капилляры вторичной
сети, сливаясь, образуют выносящие вены, по которым кровь (с гормонами передней доли) выносится из гипофиза. Фенестрированный эндотелий артериальных и венозных капилляров и синусоидов прилежит к тонкой базальной мембране секреторных клеток аденогипофиза.
Задняя доля гипофиза кровоснабжается преимущественно за счет нижних гипофизарных артерий, распадающихся на более мелкие ветви вплоть до капилляров, с которыми контактируют окончания про- никающих в нейрогипофиз аксонов нейросекреторных клеток супраоптического и паравентрикулярного ядер гипоталамуса. В утолщенном окончании аксона находится множество нейросекреторных гранул, содержащих нейрогормоны. Нейросекреторные гранулы из окончаний аксонов выделяются путем экзоцитоза в перикапиллярное пространство. В нейросекреторных гранулах гормоны связаны с белком-пере- носчиком - нейрофизином. В перикапиллярном пространстве гормоны, освобождающиеся от нейрофизина, транспортируются через стенки капилляров с фенестрированной цитоплазмой. Между верхними и нижними гипофизарными артериями имеются длинные артериальные анастомозы.
Передняя доля гипофиза образована тяжами эпителиальных клеток - эндокриноцитов, между ними располагаются тонкие соединительнотканные прослойки, где проходят широкие (синусоидальные) капилляры. Между эндокриноцитами и базальной мембраной капилляров имеется перикапиллярное пространство. В передней доле есть крупные клетки - хорошо окрашивающиеся некоторыми красителями хромофильные аденоциты. Среди хромофильных аденоцитов различают ацидофильные (около 40% всех клеток аденогипофиза). Это округлые клетки средних размеров, в цитоплазме которых множество крупных гранул. Крупные базофильные клетки составляют около 10% всех клеток. Остальные клетки слабо окрашиваются. Это мелкие хромофобные аденоци- ты. Ацидофильные, или альфа-клетки, вырабатывают два гормона - пролактин и соматотропин. Это мелкие округлые клетки, с небольшим количеством митохондрий, слабо выраженным комплексом Гольджи, вырабатывающие пролактин. Зернистая эндоплазматическая сеть представлена уплощенными разбросанными цистернами. Крупные (до 6000-9000 нм) гранулы неправильной формы окрашиваются эритрозином. Соматотропные эндокриноциты, секретирующие соматотропный гормон, имеют средние размеры. Их крупное округлое ядро расположено в центре клетки. В цитоплазме находятся небольшое количество округлых или овальных митохондрий, хорошо развитый комплекс Гольджи, большое
количество длинных узких цистерн зернистой эндоплазматической сети и множество свободных рибосом. Многочисленные круглые секреторные гранулы имеют размеры 300-400 нм. Гранулы имеют электронноплотную сердцевину, окруженную мембраной.
Все базофильные эндокриноциты, или бета-клетки, окрашиваются основными красками. Круглые гонадотропные эндокриноциты секретируют фоллитропин и лютропин (ФСГ и ЛГ). Ядро клеток, продуцирующих ФСГ, круглое, ЛГ - складчатое. В цитоплазме располагаются хорошо развитый комплекс Гольджи и зернистая эндоплазматическая сеть, обра- зованная множеством круглых или полиморфных цистерн. Большое количество мелких электронно-плотных гранул разных размеров (от 100 до 300 нм). В цитоплазме иногда встречаются гликопротеидные капли.
Крупные неправильной формы тиротропные эндокриноциты, секретирующие тиротропный гормон (ТТГ), имеют крупное удлиненное ядро, компактный комплекс Гольджи, большое количество широких митохондрий. Зернистая эндоплазматическая сеть развита умеренно. Многочисленные мелкие гранулы неправильной формы размерами от 100 до 200 нм.
Кортикотропные эндокриноциты, продуцирующие адренокортикотропный гормон (АКТГ), неправильной формы, имеют широкие цитоп- лазматические отростки, оканчивающиеся вблизи синусоидных капилляров. Ядра этих клеток дольчатые, в электронно-светлой цитоплазме расположены развитые митохондрии, выраженный комплекс Гольджи, элементы которого распространяются по всей клетке, небольшое количество узких цистерн зернистой эндоплазматической сети. Немногочисленные секреторные гранулы диаметром 100-200 нм располагаются вблизи цитолеммы. При окрашивании по методу Шика в сочетании с тионинальдегидом гранулы приобретают красный цвет.
Аденоциты, которые не секретируют гормонов, представлены мелкими клетками с крупным ядром и небольшим количеством цитоплазмы, где только при электронной микроскопии выявляется небольшое количество маленьких гранул. Возможно, эти клетки являются предшественниками хромофильных аденоцитов (Rhodin A., 1974). Однако не исключено, что «хромофобные и хромофильные клетки - это клетки одного и того же типа, различающиеся по функциональной активности» (Хэм А., Кормак Д., 1983).
Бугорная (туберальная) часть передней доли гипофиза пронизана мелкими артериями, кровоснабжающими переднюю долю, венами портальной системы и синусоидными капиллярами. Между сосудами
располагаются тяжи эпителиальных клеток и изредка заполненные коллоидом псевдофолликулы. Клеточный состав бугорной части представлен хромофобными, ацидофильными и базофильными аденоцитами, аналогичными описанным клеткам.
Тонкая промежуточная часть гипофиза образована полоской аденоцитов, которые вырабатывают секрет, накапливающийся между клетками, в результате чего образуются псевдофолликулы. У человека в соеди- нительной ткани промежуточной части залегают полиморфные базофильные клетки, которые иногда по ходу сосудов амебоидно внедряются в паренхиму задней доли. С возрастом проникновение базофильных клеток увеличивается. В промежуточной части обнаруживаются меланостимулирующий (МСГ) и адренокортикотропный (АКТГ) гормоны, а также лютропин (ЛГ). ЛГ усиливает обмен липидов. Промежуточная часть гипофиза пронизана содержащими нейросекрет нервными волокнами, исходящими из нейрогипофиза, которые образуют синапсоподобные контакты с эпителиоцитами. Холинергические и аминергические волокна оказывают ингибирующее действие на секрецию клеток промежуточной части.
Нейрогипофиз не синтезирует гормоны. Вазопрессин и окситоцин, син- тезируемые супраоптическим и паравентрикулярными ядрами гипоталамуса, транспортируются по аксонам нейросекреторных клеток, имеющихся в составе гипоталамо-гипофизарного тракта, и выделяются в кровь, протекающую в капиллярах нейрогипофиза. В нейрогипофизе выявляются густая сеть безмиелиновых нервных волокон, глиоциты, множество кровеносных сосудов. Многоотростчатые глиоциты являются протоплазматическими астроцитами, обладают фагоцитарными способностями. Тонкая сеть, образованная аргирофильными соединительнотканными волокнами, окружает капилляры. В задней доле имеется множество телец Херринга.
Тропные гормоны гипофиза регулируют деятельность гипофизозависимых желез по принципу обратной связи. При снижении концентрации определенного гормона в крови соответствующие клетки передней доли гипофиза выделяют тропный гормон, который стимулирует образование гормона этой железой. Наоборот, повышение содержания гормона в крови становится сигналом для клеток гипофиза, которые отвечают замедлением секреции и освобождения тропного гормона, что приводит к подавлению секреции гормона.
Возрастные особенности гипофиза. Развитие гипофиза начинается на 9-й неделе эмбриогенеза дифференцировкой базофильных эндокриноцитов, на 4-м месяце - ацидофильных. Масса гипофиза новорожденного
равна
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа (glandula thyroidea) расположена на шее впереди гортани. В ней различают две доли и перешеек, который лежит на уровне дуги перстневидного хряща, а иногда - 1-3-го хряща трахеи (рис. 120). Щитовидная железа как бы охватывает гортань спереди и с боков. Задне-боковая поверхность каждой доли щитовидной железы прилежит к гортанной части пищевода и передней полуокружности общей сонной артерии. Масса железы у взрослого человека составляет 20-30 г.
Железа покрыта снаружи капсулой, образованной соединительной тканью, с пучками коллагеновых и эластических волокон. От капсулы внутрь железы отходят слабо выраженные перегородки - трабекулы, в которых проходят сосуды и нервы. Трабекулы делят железу на неполные дольки. Паренхима железы состоит из пузырьков-фолликулов, являющихся основными структурными и функциональными единицами (рис. 121).
В щитовидной железе человека имеется около 30 млн фолликулов. Стенки фолликулов образованы одним слоем кубических клеток - тироцитов, лежащих на базальной мембране, в которой имеются отверстия, где клетки соседних фолликулов
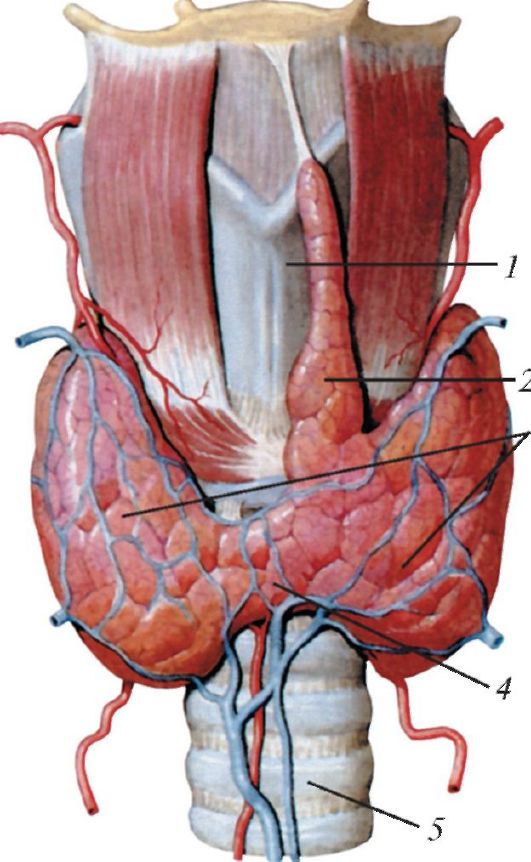
Рис. 120. Щитовидная железа, вид спереди: 1 - щитовидный хрящ; 2 - пирамидальная доля; 3 - правая и левая доли; 4 - перешеек щитовидной железы; 5 - трахея
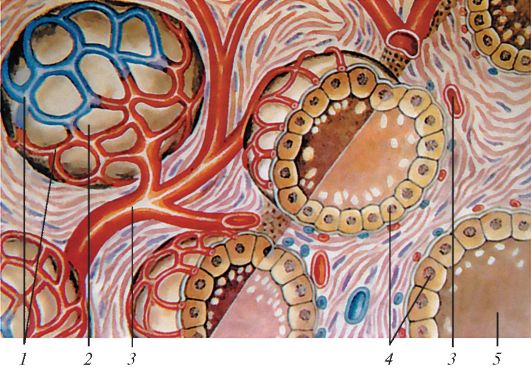
Рис. 121. Схема строения фолликулов щитовидной железы: 1 - кровеносные капилляры; 2 - фолликул; 3 - артерия; 4 - железистые клетки; 5 - коллоид, содержащий гормоны щитовидной железы (по Ю.И. Афанасьеву
и Е.Ф. Котовскому)
контактируют между собой. Встречаются разветвленные фолликулы. Размеры фолликулов колеблются от 0,05 до
В полости фолликула имеется густой вязкий коллоид щитовидной железы, содержащий тиреоглобулин, который синтезируется тироцитами. Крупное ядро расположено в центре тироцита. Цитоплазма тироцитов богата свободными рибосомами, разветвленными широкими цистернами зернистой эндоплазматической сети, развитый комплекс Гольджи расположен над ядром или вокруг ядра. Количество митохондрий невелико, лизосом - множество. Межклеточные щели расширены, кроме участков межклеточных контактов вблизи клеточной поверхности. Поверхность клеток несет на себе множество коротких тонких микроворсинок, погруженных в коллоид. Апикальная часть клеток заполнена многочисленными мелкими гранулами умеренной электронной плотности (апикальные везикулы), которые содержат вещество, похожее по ультраструктуре на коллоид.
Синтез и освобождение гормонов представляют собой сложный процесс. На рибосомах зернистой эндоплазматической сети синтезируется белок, который поступает в цистерны, где к нему присоединяются боковые углеводные цепи. Белок переносится транспортными пузырьками в комплекс Гольджи, где происходит его дальнейшее гликозилирование - образование тиреоглобулина. От комплекса Гольджи отпочковываются просекреторные гранулы, которые, созревая, превращаются в секреторные и затем в апикальные пузырьки, которые выходят в полость фолликула.
Йодирование тиреоглобулина происходит в полости фолликула, в коллоиде. Из крови капилляров в тиреоциты транспортируются ионы йода, откуда он поступает в коллоид. Капельки коллоида, отделенные псевдоподиями, имеющимися на апикальной поверхности тироцита, фагоцитируются. В цитоплазме эти капельки соединяются с лизосомами, ферменты которых расщепляют коллоид, в результате чего освобождаются гормоны трийодтиронин и тетрайодтиронин. Эти гормоны выделяются через базальную поверхность тироцита в перикапиллярное пространство, а из него в кровь. Основная функция этих гормонов - стимуляция окислительных процессов в клетке. Гормоны влияют на водный, белковый, углеводный, жировой, минеральный обмен, рост, развитие и дифференцировку тканей.
Секреция тиреоидных гормонов регулируется ТТГ гипофиза, который, в свою очередь, зависит от ТРГ-рилизинг-гормона. ТТГ действует на тироциты путем активации аденилатциклазы.
В стенках фолликулов между тироцитами и базальной мембраной имеются более крупные, светлые пара(около)фолликулярные клетки (их верхушка не достигает просвета фолликула). Парафолликулярные эн- докриноциты (кальцитониноциты) лежат на базальной мембране. Крупное округлое ядро расположено в центре клетки. В цитоплазме находятся множество пучков микрофиламентов, митохондрий, выраженный комплекс Гольджи. Зернистая эндоплазматическая сеть развита хорошо. В клетках имеется большое количество мембранных аргирофильных секреторных гранул высокой электронной плотности диаметром около 0,15 мкм, содержащих гормон тиреокальцитонин, который выделяется путем экзоцитоза в перикапиллярное пространство, откуда поступа- ет в кровеносные капилляры.
Тиреокальцитонин, участвующий в регуляции обмена кальция и фосфора, является антагонистом гормона паращитовидных желез. Он тормозит реабсорбцию кальция из костей и уменьшает содержание кальция в крови.
Кровоснабжение щитовидной железы. К верхним полюсам обоих долей подходят правая и левая верхние щитовидные артерии (ветви наружных сонных артерий), к нижним полюсам правой и левой долей - нижние щитовидные артерии (из щитошейных стволов подключичных артерий). Венозная кровь от щитовидной железы оттекает по верхней и средней щитовидным венам во внутреннюю яремную вену, по нижней щитовидной вене - в плечеголовную вену (или нижний отдел внутренней яремной вены).
Лимфатические сосуды щитовидной железы впадают в щитовидные, предгортанные, пред- и паратрахеальные лимфатические узлы.
Нервы щитовидной железы происходят из шейных узлов симпатического ствола и от блуждающего нерва.
Развитие и возрастные особенности щитовидной железы. Щитовидная железа начинает развиваться из эпителия передней кишки на 4-й неделе эмбрионального периода. Ее вырост - щитоязычный проток в дистальном отделе делится на два отростка - будущие правую и левую доли. У новорожденного масса железы равна 5-6 г, к 1 году она уменьшается до 2-2,5 г, затем постепенно возрастает, достигая к 12-14 годам 10-14 г, а к 25 годам - 18-24 г. После 60-65 лет масса железы уменьшается в связи с возрастной атрофией.
ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Две пары мелких паращитовидных (околощитовидньгх) желез (верхние и нижние) (glandulae parathyroideae) располагаются на задней поверхности долей щитовидной железы (рис. 122), но их число варьирует от 2 до 8. Размеры каждой железы 4-8x3 - 4x2-3 мм. Обычно одна железа расположена вверху, другая внизу, позади каждой доли щитовидной же- лезы. Общая масса всех паращитовидных желез у взрослого человека равна 0,2-0,35 г.
Подобно всем эндокринным железам, паращитовидные железы покрыты тонкой соединительнотканной капсулой, от которой вглубь отходят соединительнотканные прослойки, делящие ткань железы на неполные дольки. Соединительная ткань капсулы и перегородок богата нервными волокнами, кровеносными и лимфатическими сосудами. Паренхима железы образована паратироцитами, формирующими переплетающиеся между собой эпителиальные перекладины. У детей иногда видны фолликулоподобные образования, напоминающие фолликулы щитовидной железы. Различают два вида паратироцитов: светлые главные
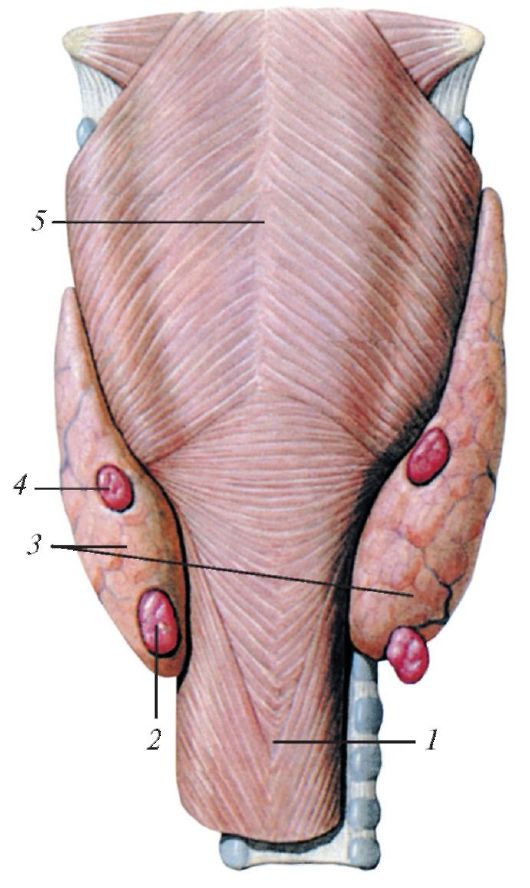
Рис. 122. Паращитовидные железы,
вид сзади: 1 - пищевод; 2 - нижняя левая паращитовидная железа; 3 - щитовидная железа; 4 - верхняя левая паращито- видная железа; 5 - глотка (нижний констриктор)
с базофильной цитоплазмой (у детей в возрасте до 8-10 лет других клеток нет) и ацидофильные (появляются после 8-10 лет). Каких-либо закономерностей расположения клеток обоих типов в паренхиме железы не выявлено. Мелкие полиэдрические главные клетки имеют базофильную цитоплазму, богатую свободными рибосомами, хорошо развитый ком- плекс Гольджи, от которого отшнуровываются окруженные мембранами электронно-плотные гранулы, размерами 15-20 нм, которые часто ассоциируются с липидными каплями различных размеров. Зернистая эндоплазматическая сеть представлена уплощенными цистернами. Среди главных клеток встречаются тем- ные и светлые паратироциты. В цитоплазме последних много частичек гликогена и мало секреторных гранул. По-видимому, обе разновидности главных клеток характеризуют различные фазы секреторной активности: темные активно секретируют, светлые неактивны, либо находятся в фазе подготовки к синтезу. Границы главных паратироцитов четко очерчены, клетки соединены между собой с помощью десмосом и интердигитаций плазмалеммы.
Крупные оксифильные клетки полиэдрической формы также имеют четкие контуры. Их цитоплазма заполнена мелкими митохондриями с ламеллярными кристами и частичками гликогена, комплекс Гольджи выражен слабо. В настоящее время считается, что оба вида клеток - по существу одни и те же клетки на разных этапах развития и функционального состояния. Главные клетки синтетически активны, оксифильные представляют стадии жизненного цикла паратироцитов. Между клетками
обоих типов существуют переходные формы. Возможно, что некоторые оксифильные клетки сохраняют способность синтезировать гормон.
Тяжи и скопления паратироцитов окружены базальной мембраной и сеточкой, образованной нежными ретикулярными фибриллами. Кровеносные капилляры проходят в соединительнотканных прослойках. Секреторные гранулы опорожняются по всей клеточной поверхности, гормон выделяется в перикапиллярное пространство, а затем в просвет капилляров. Главные клетки синтезируют и секретируют белковый паратиреоидный гормон, который регулирует концентрацию кальция и опосредованно фосфора в крови и тем самым влияет на возбудимость нервной и мышечной системы. После удаления паращитовидных желез содержание кальция в крови снижается, а фосфора повышается. Гормон действует на костную ткань, активируя остеокласты, которые обусловливают деминерализацию костной ткани, что приводит к выделению ионов кальция и фосфора в кровь. Избыток фосфора под влиянием гормона удаляется почками. Вместе с тем гормон усиливает реабсорбцию кальция в почке, что способствует уменьшению его выделения с мочой и повышению содержания в крови. Гормон повышает всасывание кальция в кишечнике при условии поступления в организм необходимого количества витамина D. Антагонистом паратгормона является тиреокальцитонин, вырабатываемый описанными выше околофолликулярными клетками щитовидной железы. Он тормозит выделение кальция из костей, ввиду чего снижается его содержание в крови. Клетки паращитовидных желез и парафолликулярные клетки реагируют на изменение содержания кальция в крови изменением синтеза и секреции своих гормонов, что и регулирует содержание кальция в крови.
Кровоснабжение паращитовидных желез. Железы снабжаются кровью из ветвей верхних и нижних щитовидных артерий, а также из пищеводных и трахеальных ветвей. Венозная кровь оттекает по венам, прилежащим к артериям.
Околощитовидные железы иннервируются симпатическими волокнами, отходящими от шейных узлов симпатического ствола, и от ветвей блуждающих нервов.
Развитие и возрастные особенности паращитовидных желез. Па- ращитовидные железы начинают развиваться на 7-й неделе эмбрионального периода из эпителия третьего и четвертого жаберных карманов. Эпителиальные зачатки желез обособляются от стенок жаберных карманов и в процессе роста перемещаются в каудальном направлении. В дальнейшем формирующиеся паращитовидные железы занимают
постоянное положение на задней поверхности правой и левой долей щитовидной железы.
У новорожденных масса паращитовидных желез не превышает 10 мг, к 1 году она достигает 20-30 мг, к 5 годам удваивается, к 10 годам возрастает в 3 раза, а к 20 годам достигает постоянной величины, не изменяясь в течение всей жизни человека.
НАДПОЧЕЧНИКИ
Надпочечник (glandula suprarenalis) состоит из двух функционально различных частей (желез), представленных корковым и мозговым веществом. Корковое вещество, или кора (cortex) развивается из мезодермы, расположенной между двумя первичными почками, на 5-й неделе внутриутробного развития (из того же источника возникают зачатки половых желез). Мозговое вещество (medulla) имеет эктодермальное происхождение. Оно закладывается на 6-7-й неделе развития из выселяющихся из закладки узлов симпатического ствола симпатобластов, которые превращаются в хромаффинобласты, дифференцирующиеся в хромаффиноциты. Наряду с мозговым веществом симпатобласты дают начало параганглиям. Зачаток мозгового вещества внедряется в зачаток коркового, в результате чего образуется единый надпочечник.
Надпочечник, напоминающий по форме уплощенную пирамиду со слегка закругленной вершиной, располагается забрюшинно в толще око- лопочечного жирового тела на уровне XI-XII грудных позвонков, непосредственно над верхним полюсом почки (рис. 123). Правый надпочечник лежит ниже левого. Масса одного надпочечника у взрослого человека около 12-13 г, размеры - 40-60x20 - 30x2-6 мм.
На передней поверхности каждого надпочечника видны ворота органа, через которые выходит центральная вена надпочечника. Надпочеч- ник покрыт соединительнотканной капсулой, от которой в глубь железы отходят тонкие прослойки, разделяющие его корковое вещество на множество эпителиальных тяжей, окутанных густой сетью капилляров.
В корковом веществе различают три зоны: клубочковую (наружную), пучковую (среднюю) и сетчатую (на границе с мозговым слоем) (рис. 124). Клубочковая зона (zona gromerulosa) образована мелкими полиэдрическими призматическими клетками, расположенными в виде клубочков. Извитые капилляры окружают клубочки. В поверхностных отделах капилляры имеют базальную мембрану, в более глубоких базальная мембрана почти полностью исчезает. Эндотелий капилляров фенестрированный.
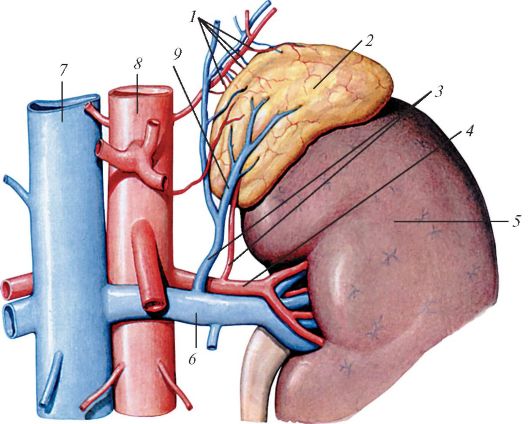
Рис. 123. Левый надпочечник: 1 - верхние надпочечниковые артерии и вены; 2 - надпочечник; 3 - нижние надпочечниковые артерия и вена; 4 - почечная артерия; 5 - почка; 6 - почечная вена; 7 - нижняя полая вена; 8 - аорта; 9 - центральная вена надпочечника
(по Р. Д. Синельникову)
Митохондрии с тубулярными кристами и электронно-плотным матриксом относительно малочисленны.
Незернистая эндоплазматическая сеть представлена различными пузырьками, количество элементов зернистой эндоплазматической сети невелико, комплекс Гольджи развит умеренно, цитоплазма заполнена каплями одинакового размера диаметром около 0,5 мкм. Неровная поверхность клеток покрыта микроворсинками, которые находятся в широких межклеточных пространствах.
Самая широкая часть коры надпочечников представлена пучковой зоной (zona fasciculata). Она сформирована крупными светлыми многогранными клетками, располагающимися длинными тяжами, ориентированными перпендикулярно поверхности органа. Между тяжами проходят прямые капилляры, которые анастомозируют между собой. Каждый эндокриноцит контактирует с соседним капилляром. Эндотелиоциты,
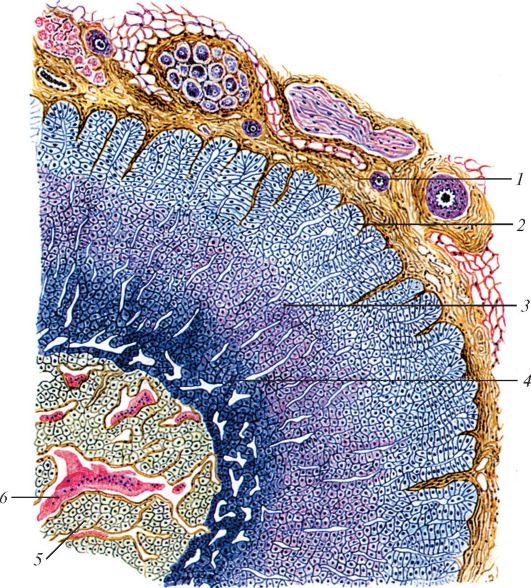
Рис. 124. Микроскопическое строение надпочечника: 1 - капсула надпочечника; 2 - клубочковая зона; 3 - пучковая зона; 4 - сетчатая зона; 5 - мозговое вещество; 6 - синусоидный капилляр (по И.В. Алмазову
и Л.С. Сутулову)
образующие стенки капилляров, фенестрированны. Эндокриноциты заполнены липидными каплями разных размеров. Хорошо развитая незернистая эндоплазматическая сеть образует пузырьки и трубочки, которые располагаются слоями вокруг крупных сферических митохондрий с везикулярными кристами и липидных капель. В цитоплазме много свободных рибосом и частичек гликогена. Клетки пучковой зоны богаты холестерином и аскорбиновой кислотой.
В сетчатой зоне (zona reticularis) мелкие полиэдрические или кубические клетки с хорошо развитой незернистой эндоплазматической сетью
образуют небольшие скопления (группы клеток), имеющие различное направление. Между группами клеток проходят капилляры. Клетки и капилляры формируют широкопетлистую сеть. Посткапиллярные венулы в сетчатой зоне несколько шире, чем в пучковой. Ядра эндокриноцитов сетчатой зоны мелкие. В цитоплазме имеется небольшое количество крупных липидных капель высокой электронной плотности. Крупные митохондрии имеют кристы тубулярного и везикулярного типов. Развитая эндоплазматическая сеть представлена множеством везикулярных элементов. В цитоплазме большое количество первичных и вторичных лизосом, а также остаточных телец, заполненных гранулами липофусцина. В пери- капиллярном пространстве встречаются макрофаги.
Указанные зоны коркового вещества анатомически достаточно четко отделены друг от друга. По современным воззрениям, их клетки синтезируют и секретируют различные гормоны: клубочковая - минералокортикоиды (альдостерон), средняя пучковая - глюкокортикоиды (гидрокортизон и кортикостерон), внутренняя сетчатая - андрогены, эс- трогены и прогестерон (последние в небольшом количестве).
Минералокортикоиды участвуют в регуляции электролитного обмена и опосредованно - в регуляции водного. Альдостерон усиливает реабсорбцию натрия в почках, слюнных железах, желудочно-кишечном тракте и тем самым задерживает натрий в организме. Альдостерон изменяет проницаемость клеточных мембран для натрия и калия, усиливает воспалительные процессы и образование коллагена. Секреция альдостерона регулируется концентрацией ионов натрия и калия в плазме крови и ангиотензином II. Увеличение содержания калия и уменьшение натрия, уменьшение объема плазмы приводят к усилению секреции альдостерона. Сходный эффект дает ангиотензин II, который образуется из плазменного ангиотензина I под влиянием ренина, вырабатываемого юкстагломерулярными клетками почки.
Глюкокортикоиды оказывают мощное и разностороннее действие на организм. Они оказывают катаболическое действие на белковый обмен и стимулируют глюконеогенез, что приводит к повышению уровня глюкозы в крови и гликогена в печени, скелетных мышцах и миокарде. Глюкокортикоиды регулируют липолиз, нормализуют выделение воды из организма, повышая клубочковую фильтрацию и уменьшая реабсорбцию воды в дистальных извитых канальцах почки. Известно выраженное противовоспалительное действие глюкокортикоидов. Введение гидрокортизона вызывает инволюцию иммунной системы, выраженную иммуносупрес- сию, что связано с разрушением лимфоидной ткани.
Глюкокортикоиды избирательно тормозят развитие основного вещества соединительной ткани и пролиферацию фибробластов, уменьшают количество тканевых базофилов, подавляют действие фермента гиа- луронидазы и тем самым уменьшают проницаемость капилляров. Стресс-факторы вызывают увеличение секреции глюкокортикоидов вплоть до истощения секреторных клеток коркового вещества надпочечников. Секреция глюкокортикоидов эндокриноцитами пучковой и частично сетчатой зон регулируется АКТГ.
Мозговое вещество надпочечников образовано скоплениями крупных округлых и многоугольных клеток, разделенных синусоидными капиллярами и венулами. Клетки хорошо окрашиваются солями хрома в коричневый цвет, поэтому они названы хромаффинными. К клеткам подходят преганглионарные симпатические нервные волокна, которые образуют на них синапсы. В мозговом веществе различают два вида клеток: эпинефроциты, вырабатывающие адреналин, и норэпинефроциты, вырабатывающие норадреналин. Клетки богаты гранулами умеренной электронной плотности, заключенными в мембрану, их количество непостоянно и зависит от активности клетки. В цитоплазме эпинефроцитов находятся развитый комплекс Гольджи, небольшое количество мелких митохондрий, множество свободных рибосом и частичек гликогена. Норэпинефроциты располагаются группами, они отличаются от эпинефроцитов лишь гранулами высокой электронной плотности.
Мозговое вещество продуцирует небольшое количество гормонов, и лишь при воздействии на организм сильных раздражителей секреция резко усиливается. У взрослого человека мозговое вещество надпочеч- ников выделяет около 70-90% адреналина и 10-30% норадреналина. Оба гормона являются метаболическими. Молекулы катехоламинов связываются со специфическими рецепторами цитолеммы клеток-мишеней. Норадреналин вызывает сужение сосудов (кроме сосудов головного мозга), адреналин суживает одни (сосуды кожи) и расслабляет другие сосуды (коронарные и сосуды скелетных мышц). Адреналин повышает систолическое артериальное давление, не изменяя или несколько снижая диастолическое, норадреналин повышает и систолическое, и диастолическое давление. Оба гормона вызывают расслабление гладких мышц бронхов, что приводит к углублению дыхания.
Адреналин повышает уровень сахара в крови, вызывая распад гликогена. Эффект норадреналина более слабый. Оба гормона усиливают распад жиров, активируют энергетический обмен. Секреция катехола- минов, которая в обычных физиологических условиях невелика и равна
8-10 (нгмин), резко возрастает при возбуждении симпатической нервной системы, в стрессовых ситуациях.
Кровоснабжение надпочечников. Каждый надпочечник получает 25-30 артерий. Наиболее крупные из них верхние надпочечниковые (из нижней диафрагмальной артерии), средняя надпочечниковая (из брюшной части аорты) и нижняя надпочечниковая (из почечной артерии) артерии. Одни ветви этих артерий кровоснабжают только корковое вещество, другие прободают корковое вещество надпочечника и разветв- ляются в мозговом веществе. Из синусоидных кровеносных капилляров формируются притоки центральной вены, которая у правого надпочечника впадает в нижнюю полую вену, у левого - в левую почечную вену. Из надпочечника (особенно левого) выходят многочисленные мелкие вены, впадающие в притоки воротной вены.
Лимфатические сосуды надпочечников впадают в поясничные лимфатические узлы.
В иннервации надпочечников участвуют блуждающие нервы, а также нервы, происходящие из чревного сплетения, которые содержат преганглионарные симпатические волокна для мозгового вещества.
Возрастные особенности надпочечников. Масса
одного надпочечника у новорожденного составляет около 8-9 г и
значительно превышает массу надпочечника ребенка 1-го года жизни. В
первые 3 мес после рождения масса надпочечника резко уменьшается (до
3-4 г), главным образом в результате истончения и перестройки коркового
вещества, а затем постепенно восстанавливается (к 5 годам) и
продолжает нарастать. Формирование коркового вещества надпочечников
завершается в период второго детства (8-12 лет). К 20 годам масса
каждого надпочечника увеличивается в 1,5 раза (по сравнению с массой у
новорожденного) и достигает максимума (в среднем 12-13 г). В
последующие возрастные периоды размеры и масса надпочечников почти не
изменяются. Надпочечники у женщин в среднем несколько большие, чем у
мужчин. Во время беременности масса каждого надпочечника увеличивается
примерно на
ПАРАГАНГЛИИ
Кроме мозгового вещества надпочечников, хромаффиноциты находятся также в параганглиях (paraganglia), которые образуются из зачатка симпатической нервной системы и тесно связаны с симпатическими
узлами. Большинство параганглиев расположено ретроперитонеально вблизи симпатического ствола в виде обособленных анатомических структур. Это межсонный параганглий (сонный клубочек), расположен- ный у начала наружной и внутренней сонных артерий, пояснично-аортальные на передней поверхности брюшной части аорты. Множество мелких параганглиев расположено ретроперитонеально на участке от надпочечников до половых желез и по ходу симпатических нервов. Параганглии всегда обнаруживаются вблизи семенных пузырьков, в маточно-влагалищном нервном сплетении. Количество и величина параганглиев широко варьируют у разных людей. У новорожденных их число достигает 40. Кроме того, у новорожденных имеются множественные скопления хромаффиноцитов и отдельные клетки в ганглиях и ветвях симпатической части вегетативной нервной системы. Мелкие группы хромаффиноцитов или даже отдельные клетки располагаются внутри вегетативных нервных узлов. Параганглии, окруженные тонкой соединительнотканной капсулой, состоят из соединенных между собой хромаффиноцитов, образующих сеть, в просветах которой проходят многочисленные кровеносные капилляры. Внутри нервов и симпатических ганглиев хромаффиноциты залегают в виде клеточных скоплений. Хромаффиноциты окружены густой сетью нервных волокон. Клетки аналогичны описанным в мозговом ве- ществе надпочечников. Они содержат множество мелких гранул, окруженных мембраной. В настоящее время установлено, что клетки параганглиев секретируют катехоламины.
Инволюция
анатомически обособленных параганглиев начинается в 1,5-2 года и
заканчивается после завершения полового созревания. Возникают
гиперемия, дегенерация хромаффиноцитов, разрастание со- единительной
ткани. Пояснично-аортальные параганглии хорошо выражены у новорожденных
и грудных детей. Это небольшие тонкие полоски, расположенные по обеим
сторонам аорты на уровне начала нижней брыжеечной артерии. У
новорожденных их размеры составляют 8-15x2-3 мм, у грудных - около
ЭНДОКРИННАЯ ЧАСТЬ ПОЛОВЫХ ЖЕЛЕЗ
Половые железы (яичко и яичник) вырабатывают половые гормоны, которые всасываются в кровь. Как указывалось ранее, эту функцию в яичке осуществляют интерстициальные эндокриноциты, или клетки Лейдига. Это крупные клетки, которые располагаются скоплениями между семенными канальцами около кровеносных капилляров. Они богаты элементами незернистой цитоплазматической сети. В настоящее время считают, что эти клетки активно участвуют в образовании мужского полового гормона - тестостерона (андроген), который оказывает разностороннее действие на различные андрогенчувствительные клетки мужского организма, стимулируя их рост и функциональную активность. К таким клеткам относятся клетки предстательной железы, семенных пузырьков, желез крайней плоти, почек и кожи. Под влиянием андрогенов происходят эмбриональная дифференцировка и развитие половых органов, половое созревание и развитие вторичных половых признаков, опорно-двигательного аппарата. Вместе с другими гормонами андрогены регулируют рост костей в толщину и длину и после наступления половой зрелости прекращают его. Андрогены стимулируют синтез белка и ускоряют рост тканей. Очень важно их воздействие на сперматогенез: низкая концентрация гормона активирует этот процесс, высокая - тормозит. Андрогены обусловливают половое поведение муж- чины, его либидо и потенцию. Следует указать, что в яичках синтезируется и небольшое количество эстрогенов (женских половых гормонов).
Женские половые гормоны вырабатываются в яичнике. Клетки фолликулярного эпителия синтезируют эстрогены (фолликулин), лютеоциты, клетки желтого тела - гестагены (прогестерон). Кроме того, в яичниках образуется небольшое количество андрогенов. Эстрогены, как и андрогены у мужчин, влияют на эмбриональную дифференцировку и на развитие половых органов, вторичных половых признаков, рост и развитие опорно-двигательного аппарата; обеспечивая развитие тела по женскому типу, влияют на половое поведение. Женские половые гормоны обусловливают описанные выше изменения эндометрия во время менструального цикла. Эстрогены стимулируют пролиферацию эндометрия. Прогестерон влияет на слизистую оболочку матки, готовя ее к имплантации оплодотворенной яйцеклетки и развитию плода, образованию плаценты, увеличению размеров молочных желез, а также задерживает рост новых фолликулов. Наряду с этим половые гормоны оказывают и экстрагенитальное действие. Так, прогестерон вызывает повышение
утренней температуры тела в покое (базальной температуры), оказывает катаболическое действие.
ШИШКОВИДНОЕ ТЕЛО
Шишковидное тело, или эпифиз (corpus pineale, epiphysis cerebri), располагается
в бороздке между верхними холмиками четверохолмия сред- него мозга и
прикреплен поводками к обоим зрительным буграм промежуточного мозга.
Эпифиз имеет овоидную форму, его масса у взрослого человека не превышает
Большинство пинеалоцитов имеют электронно-плотную цитоплазму, богатую рибосомами, остальные - светлую с небольшим количеством рибосом. Большое округлое ядро содержит много периферического гетерохроматина. В цитоплазме есть свободные рибосомы и полисомы, короткие цистерны зернистой эндоплазматической сети, большое ко- личество трубчатых элементов незернистой эндоплазматической сети, хорошо выраженный комплекс Гольджи. Число митохондрий, первичных и вторичных лизосом варьирует. Вблизи комплекса Гольджи располагаются крупные электронно-плотные гранулы, которые имеются в отростках и их окончаниях. В пинеалоцитах мало цитоплазматических филаментов и большое количество микротрубочек. Иногда в пинеалоцитах встречаются необычные органеллы, которые образованы группами микротрубочек и перфорированных пластинок, связанных с элементами
зернистой эндоплазматической сети и липидными каплями, - так называемые аннулярные ламеллы.
Глиоциты эпифиза представляют собой астроциты, которые окру- жают пинеалоциты. Глиоциты богаты цитоплазматическими филаментами толщиной 5-6 нм, их пучки находятся в перикарионе и проникают в отростки. Многочисленные отростки глиоцитов окружают пинеалоциты и их отростки, а также симпатические нервы и нервные окончания. Многие отростки оканчиваются на поверхности долек паренхимы вблизи базальной пластинки, окружающей дольку. Зернистая эндоплаз- матическая сеть глиоцитов представлена множеством коротких цистерн. Количество свободных рибосом невелико. Диктиосомы комплекса Гольджи разбросаны по клетке. В цитоплазме имеются мелкие митохондрии с электронно-плотным матриксом, большое количество первичных и вторичных лизосом, частичек гликогена.
В эпифизе человека встречаются характерные округлые тельца, которые образованы концентрическими слоями солей кальция и органическим матриксом (мозговой песок). Эпифиз выполняет нейросекреторную функцию, которая подчинена циркадным ритмам и каким-то образом имеет обратную связь с освещенностью. Пинеалоциты синтезируют мелатонин, серотонин и ряд полипептидов, оказывающих гормональное действие. Мелатонин является антагонистом МСГ, вырабатываемого гипофизом.
Функция пинеалоцитов имеет четкий суточный ритм: ночью синтезируется мелатонин, днем - серотонин. Этот ритм связан с освещенностью, свет вызывает угнетение синтеза и секреции мелатонина. В последние годы показано, что мелатонин имеет обратную связь и с рилизинг-фактором ЛГ и тормозит его выделение. В настоящее время есть все основания считать, что эпифиз влияет на половое созревание, функцию половых желез и такие общие состояния, как сон-бодрствование. Кроме того, пинеалоциты вырабатывают гормон, участвующий в регуляции содержания ионов калия в крови (он повышает их концентрацию в плазме). Пептидные гормоны ингибируют продукцию АКТТ и тироксина.
Итак, эпифиз является важнейшей железой, которая моделирует активность адено- и нейрогипофиза, панкреатических островков поджелудочной железы, паращитовидных желез, надпочечников, половых желез, щитовидной железы. Действие гормонов эпифиза может быть как прямым - на клетки-мишени соответствующих желез, так и опосредованным через рили- зинг-факторы гипоталамуса. Гормоны эпифиза достигают клетки-мишени либо гематогенным путем, либо через спинномозговую жидкость.
Кровоснабжение эпифиза осуществляется ветвями задней мозговой и верхней мозжечковой артерий. Кровь оттекает по венам, которые впадают в большую вену мозга или в ее притоки.
Иннервация эпифиза осуществляется симпатическими нервными волокнами, отходящими от верхних шейных симпатических узлов.
Развитие и возрастные особенности эпифиза. Эпифиз развивается из выпячивания крыши будущего III желудочка головного мозга на 5-6-й неделе внутриутробного периода. Клетки нейроэктодермы, образующей выпячивание, дают начало пинеалоцитам и глиоцитам. Покрывающая выпячивание мягкая мозговая оболочка образует соединительнотканные элементы железы.
У новорожденного масса эпифиза около 7 мг. В течение 1-го года после рождения она достигает 100 мг и удваивается к 10 годам, после чего практически не меняется. В пожилом возрасте в эпифизе могут воз- никать кисты, откладываться мозговой песок, поэтому его масса увеличивается.
ПАНКРЕАТИЧЕСКИЕ ОСТРОВКИ (ЭНДОКРИННАЯ
ЧАСТЬ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ)
Поджелудочная железа состоит из эндокринной и экзокринной частей. Экзокринная часть описана в разделе «Органы пищеварения». Эндокринная часть образована панкреатическими островками (insulae pancreatici) (островки Лангерганса), которые сформированы клеточными скоплениями, окруженными густыми капиллярными сетями (рис. 125). Общее количество панкреатических островков составляет 1-2 млн, а диаметр каждого островка 100-300 мкм. Клетки островков содержат множество покрытых мембранами гранул, отличающихся от гранул зимогена ацинозных клеток. Преобладают бета-клетки (60-80%), которые секретируют инсулин, альфа-клетки (10-30%) вырабатывают глюкагон, дельта-клетки (около 10%) синтезируют соматостатин. Последний угнетает выработку гипофизом гормона роста и синтез ферментов ацинозными клетками, а также выделение инсулина и глюкагона бета- и альфа-клетками. РР-клетки, расположенные по периферии островков, синтезируют полипептид, который стимулирует выделение желудочного сока и панкреатического сока экзокринной частью железы. Многочисленные бета-клетки занимают внутреннюю зону островка. Они содержат множество мелких гранул, которые окрашиваются нейтральным этиловым, фиолетовым и шарлахом в синий цвет. Ядра бета-клеток
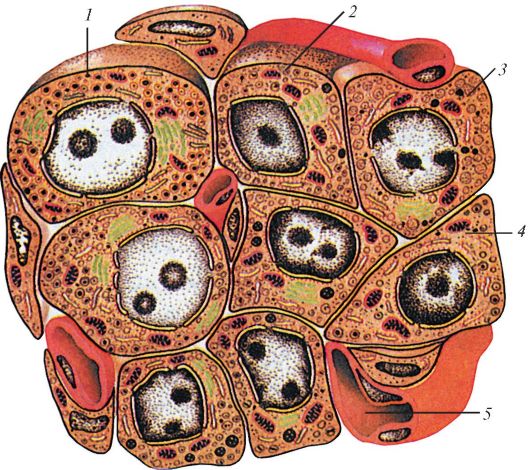
Рис. 125. Схема строения панкреатического островка: 1 - альфа-клетка; 2 - бета-клетка; 3 - дельта-клетка; 4 - РР-клетка; 5 - крове- носный капилляр (по В.Г. Елисееву и др.)
имеют неправильную форму и расположены в центре. В некоторых бета-клетках зернистая эндоплазматическая сеть развита хорошо, но количество гранул в них невелико. Крупные округлые митохондрии многочисленны, выраженный комплекс Гольджи окружен множеством пресекреторных гранул. Гранулы у человека окружены мембраной, в них заключены один или несколько прямоугольных или полигональных кристаллоидов, погруженных в матрикс. На рибосомах зернистой эндоплазматической сети синтезируется проинсулин, который подвергается процессингу в комплексе Гольджи. От него отщепляются секреторные гранулы, содержащие инсулин.
Альфа-клетки располагаются группами по всему островку. Ядра не- правильной формы занимают центральную часть клетки. Многочисленные круглые (около 300 нм) электронно-плотные гранулы окружены мембранами. В цитоплазме находится небольшое количество мелких
митохондрий, слабо развитый комплекс Гольджи и умеренное количество коротких цистерн зернистой эндоплазматической сети.
Дельта-клетки имеют большие размеры, низкую электронную плотность и крупные (320-350 нм) секреторные гранулы высокой электронной плотности. Органеллы развиты слабее, чем в альфа- и бета-клетках.
РР-клетки имеют мелкие гранулы размерами около 140 нм. Кроме описанных, в островках человека встречается небольшое количество дельта-клеток, содержащих самые мелкие аргирофильные электронноплотные гранулы (около 100 нм) со светлым ободком. Эти клетки, возможно, вырабатывают вазоактивный полипептид.
Кровоснабжение панкреатических островков. Каждый островок кровоснабжается 1-3 артериолами, которые отходят от мелких артериальных ветвей. Венозная кровь поступает в нижнюю полую вену и в воротную вену печени.
Вегетативные нервы, иннервирующие поджелудочную железу, образуют вокруг островков сплетения, веточки которых проникают в островки.
ДИФФУЗНАЯ НЕЙРОЭНДОКРИННАЯ СИСТЕМА (APUD-СИСТЕМА)
В организме человека имеются гормонпродуцирующие клетки, происходящие из нейробластов нервного гребешка, а также экто- и энтодермы, и продуцирующие нейроамины и олигопептиды, которые оказывают гормональное действие. Эти клетки разбросаны в организме в виде клеточных групп или отдельных клеток. Все они объединены в APUD-систему (Amine Precursors Uptake and Decarboxylation - поглощение и декарбоксилирование предшественников аминов).
К APUD-системе относятся парафолликулярные клетки щитовидной железы, клетки мозгового вещества надпочечников, нейросекреторные клетки гипоталамуса, пинеалоциты эпифиза, главные паратироциты паращитовидных желез, эндокриноциты аденогипофиза, плаценты, поджелудочной железы, желудочно-кишечного тракта. APUD-система дополняет и связывает между собой нервную и эндокринную системы, осуществляя весьма чувствительный контроль гомеостаза.