Биология: учебник / Пехов А.П., -, 2010. - 664 с.
|
|
|
|
ГЛАВА IV РАЗМНОЖЕНИЕ, РОСТ И ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ ОРГАНИЗМОВ
Размножение - это свойство организмов производить потомство или способность организмов к самовоспроизведению. Являясь важнейшим свойством живого, размножение обеспечивает непрерывность жизни, продолжение видов.
Процесс размножения исключительно сложен и связан не только с передачей генетической информации от родителей к потомству, но и с анатомическими и физиологическими свойствами организмов, с их поведением, гормональным контролем. Размножение организмов сопровождается процессами их роста и развития.
Для живых существ характерно чрезвычайное разнообразие в способах размножения. Тем не менее различают два основных способа размножения - бесполое и половое (рис. 16). Бесполое размножение, или апомиксис (от греч. аро - без, mixis - смешение), представляет собой процесс, в котором участвует лишь один родитель (клетка или многоклеточный организм). Напротив, в половом размножении участвуют два родителя, каждый из которых имеет собственную репродуктивную систему и продуцирует половые клетки (гаметы), которые после слияния образуют зиготу (оплодотворенное яйцо), дифференцирующуюся затем в эмбрион. Следовательно, при половом размножении имеет место смешение наследственных факторов, т. е. процесс, называемый амфимиксисом (от греч. amphi - с обеих сторон, mixis - смешение).
БЕСПОЛОЕ РАЗМНОЖЕНИЕ
Бесполое размножение характерно для организмов многих видов как растений, так и животных. Оно встречается у вирусов, бактерий, водорослей, грибов, сосудистых растений, простейших, губок, кишечнополостных, мшанок и оболочников.
Наиболее простая форма бесполого размножения характерна для вирусов. Их репродуктивный процесс связан с молекулами нуклеиновых кислот, со способностью этих молекул к самоудвоению
и основан на специфичности относительно слабых водородных связей между нуклеотидами.
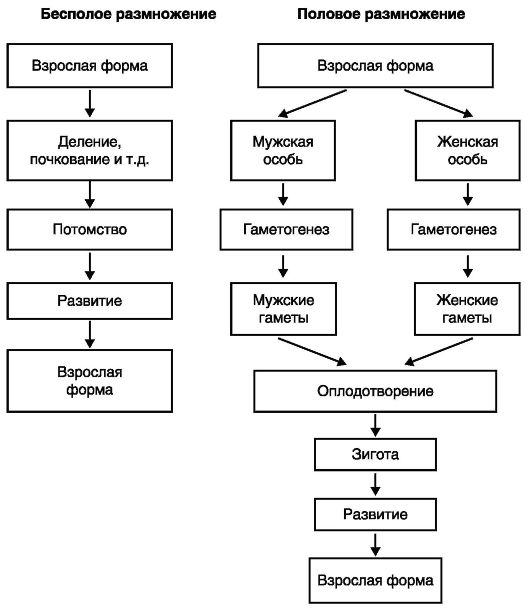
Рис. 16. Способы размножения организмов
Применительно к другим организмам, размножающимся бесполым путем, различают вегетативное размножение и размножение спорообразованием.
Вегетативное размножение - это размножение, при котором из части, отделившейся от материнского организма, развивается новый организм. Этот вид размножения характерен как для одноклеточных, так и многоклеточных организмов, но имеет у них разное проявление.
У одноклеточных организмов вегетативное размножение представлено такими формами, как деление, множественное деление и почкование. Деление путем простой перетяжки с образованием при этом из одного родительского организма двух дочерних присуще бактериям и синезеленым водорослям (цианобактериям). Напротив, размножение делением бурых и зеленых водорослей, а также одноклеточных животных (саркодовых, жгутиковых и инфузорий) происходит путем митотического деления ядра с последующей перетяжкой цитоплазмы.
Размножение путем множественного деления (шизогонии) заключается в делении ядра с последующим разделением цитоплазмы на части. В результате такого деления из одной клетки образуется несколько дочерних организмов. Примером множественного деления является размножение малярийного плазмодия (P. vivax) в эритроцитах человека. В этом случае у плазмодиев происходит повторяющееся много раз деление ядра без цитокинеза, после чего следует и цитокинез. В результате этого один плазмодий дает начало 12-24 дочерним организмам.
У многоклеточных растительных организмов вегетативное размножение путем деления осуществляется черенками, луковицами, листьями, корневищами. Но это, по существу, искусственное размножение, используемое в сельскохозяйственной практике. Размножение высших растений в искусственных условиях возможно и из одной клетки. Организмы, развивающиеся из одной клетки (клон), обладают всеми свойствами исходного многоклеточного организма. Это размножение получило название клонального микроразмножения. В качестве одной из форм вегетативного размножения могут служить прививки или трансплантации многих культурных растений, заключающиеся в пересадке почки или части побега от одного растения к другому. Конечно, это тоже способ размножения, который в природе не встречается, но в сельском хозяйстве используется очень широко.
У многоклеточных животных вегетативное размножение происходит путем фрагментации их тела на части, после чего каждая
часть развивается в новое животное. Такое размножение характерно для губок, кишечнополостных (гидр), немертин, плоских чер- вей, иглокожих (морских звезд) и некоторых других организмов. Близкой формой к вегетативному размножению животных фрагментацией является полиэмбриония животных, заключающаяся в том, что на определенной стадии развития эмбрион разделяется на несколько частей, каждая из которых развивается в самостоятель- ный организм. Полиэмбриония встречается у броненосцев. Однако последние размножаются половым путем. Поэтому полиэмбриония является скорее своеобразной стадией в половом размножении, а потомство, возникающее в результате полиэмбрионии, представлено монозиготными близнецами.
Почкование заключается в том, что на материнской клетке образуется бугорок (вырост) с ядром, который затем отделяется и становится самостоятельным организмом. Почкование встречается как у одноклеточных растений, например у дрожжей, так и у одноклеточных животных, например у инфузорий отдельных видов.
Размножение спорообразованием связано с образованием специализированных клеток - спор, которые содержат ядро, цитоплазму, покрыты плотной оболочкой и способны к длительному существованию в неблагоприятных условиях, что способствует, кроме того, и их расселению. Наиболее часто такое размножение встречается у бактерий, водорослей, грибов, мхов, папоротникообразных. У некоторых зеленых водорослей из отдельных клеток могут формироваться так называемые зооспоры.
Среди животных размножение спорообразованием отмечается у споровиков, в частности у малярийного плазмодия.
У организмов многих видов бесполое размножение может чередоваться с половым размножением.
ПОЛОВОЕ РАЗМНОЖЕНИЕ
Половое размножение встречается как у одноклеточных, так и у многоклеточных растений и животных.
Как отмечено в главах V и XIII, половое размножение у бактерий осуществляется путем конъюгации, служащей аналогом полового процесса и являющейся системой рекомбинации этих организмов, тогда как у простейших половое размножение происходит тоже путем конъюгации либо посредством сингамии и аутогамии.
У многоклеточных организмов (растений и животных) половое размножение связано с образованием зародышевых или половых клеток (гамет), оплодотворением и образованием зигот.
Половое размножение является значительным эволюционным приобретением организмов. С другой стороны, оно способствует пересортировке генов, появлению разнообразия организмов и повышению их конкурентоспособности в непрерывно меняющихся условиях окружающей среды.
У одноклеточных организмов половое размножение существует в нескольких формах. У бактерий половое размножение можно аналогизировать с имеющей место у них конъюгацией, заключающейся в передаче плазмидной или хромосомной ДНК от клеток-доноров (содержащих плазмиды) к клеткам-реципиентам (не содержащих плазмиды), а также с трансдукцией бактерий, заключающейся в передаче генетического материала от одних бактериальных клеток к другим фагам. Конъюгация встречается также у инфузорий, у которых во время этого процесса происходит переход ядер от одних особей к другим, после чего следует деление последних.
Одной из распространенных форм полового размножения у одноклеточных животных, например у паразитических споровиков, является копуляция. Половое размножение у них заключается в слиянии двух особей, которые являются гаметами, в одну, являющуюся споровой формой, из которой затем развивается новый организм. Независимо от способа слияния генетического материала у одноклеточных организмов существенной особенностью этого слияния является то, что оно сопровождается генетической рекомбинацией.
У многоклеточных растений и животных половое размножение клеток происходит через образование женских и мужских половых клеток (яйцеклеток и сперматозоидов), последующее оплодотворение яйцеклетки сперматозоидом и образование зиготы. У растений половые клетки продуцируются в специализированных репродуктивных структурах, у животных они продуцируются в половых железах, называемых гонадами (от греч. gone - семя).
Между соматическими и половыми клетками животных существует важное различие. Оно заключается в том, что соматические клетки способны к делению, т. е. репродуцируют себя, и, кроме того, из них образуются половые клетки. Напротив, половые клетки не делятся, но они «начинают» репродукцию целого организма.
Диплоидные соматические клетки, в которых образуются мужские половые клетки, называются сперматогониями, а в которых образуются женские половые клетки - овогониями. Процесс образования (роста и дифференциации) мужских и женских половых клеток носит название гаметогенеза.
Гаметогенез основан на мейозе (от греч. meiosis - понижать), который представляет собой редукционное деление ядер клеток, сопровождающееся понижением числа хромосом на ядро. Мейоз происходит в специализированных клетках репродуктивных органов живых существ, размножающихся половым путем (рис. 17). Например, у папоротникообразных мейоз встречается в специализированных клетках спорангий, располагающихся на нижней поверхности листьев этих растений и развивающихся в споры, а затем в гаметофиты. Последние существуют раздельно, продуцируя в конечном итоге мужские и женские гаметы. У цветковых растений мейоз осуществляется в специализированных клетках семяпочек, которые развиваются в споры. Последние продуцируют гаметофит с одной яйцеклеткой. Кроме того, у этих растений мейоз происходит и в специализированных клетках пыльников, которые также развиваются в споры, продуцирующие в конечном итоге пыльцу с двумя мужскими гаметами. У земляных червей, которые являются гермафродитами и содержат мужские половые органы в одном сегменте тела, а женские в другом и которые характеризу-
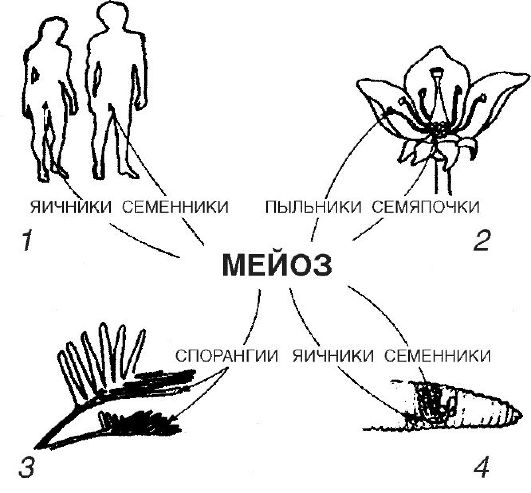
Рис. 17. Мейоз у разных организмов: 1 - человек; 2 - цветковые растения; 3 - папоротник; 4 - дождевой червь
ются способностью к перекрестному оплодотворению между разными особями, имеется способность одновременно к сперматогенезу и овогенезу.
Мейоз осуществляется в специализированных клетках семенников и яичников, в которых продуцируются мужские и женские гаметы соответственно. Выявлены белки - индукторы мейоза.
В процессе мейоза диплоидное число хромосом (2л), которое характерно для соматических клеток (клеточных ядер) и незрелых зародышей клеток, изменяется до гаплоидного числа (n), характерного для зрелых зародышевых клеток. Таким образом, в результате гаметогенеза половые клетки получают лишь половину хромосом соматических клеток (рис. 18).
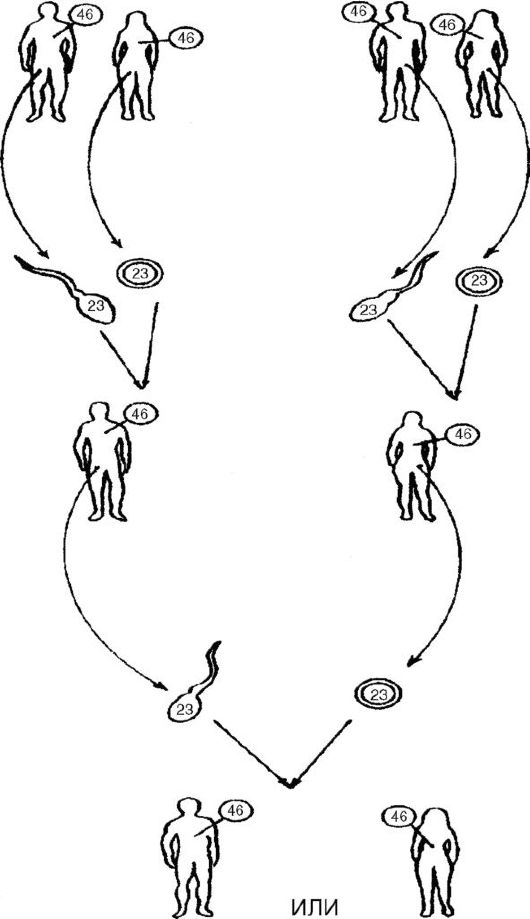
Рис. 18. Распределение хромосом при гаметогенезе
Поведение хромосом в период гаметогенеза у животных является одинаковым как у мужских, так и у женских особей. Однако полы различаются по времени происхождения разных стадий мейоза, что осо-
бенно заметно у человека. У мужских особей в постпубертатный период полный процесс мейоза завершается примерно в течение двух месяцев, в то время как у женских особей первое мейотическое деление начинается в фетальном яичнике и не завершается, пока не начнется овуляция, которая наступает у них примерно в 15 лет.
У высших животных в случае мужских особей мейоз сопровождается образованием четырех функционально активных гамет.
Напротив, у женских особей каждый овоцит II порядка дает лишь одну яйцеклетку. Другие ядерные продукты женского мейоза представляют собой три редукционных тельца, которые не участвуют в размножении и дегенерируют.
Мейоз состоит из двух ядерных делений. Первое меиотическое деление ядра разделяет членов каждой пары гомологичных хромосом после того, как они спарились одна с другой (синапсис) и обменялись генетическим материалом (кроссинговер). В результате этого разделения образуется два гаплоидных ядра. Второе мейотическое деление разделяет две продольные половины хромосом (хроматиды) в каждом из этих ядер, продуцируя четыре гаплоидных ядра.
В процессе гаметогенеза происходит также дифференциация яйцеклеток (овогенез) и сперматозоидов (сперматогенез), являю- щаяся пререквизитом их функций. Яйцеклетки животных намного крупнее сперматозоидов, обычно неподвижны и содержат питательный материал, который обеспечивает развитие эмбриона в начальном периоде после оплодотворения. Сперматозоиды большинства животных обладают жгутиком, обеспечивающим независимость их движения.
Мейоз имеет выдающееся биологическое значение. Благодаря мейозу в клетках организмов поддерживается постоянное число хромосом независимо от количества поколений. Следовательно, мейоз поддерживает постоянство видов. Наконец, в мейозе в результате кроссинговера происходит рекомбинация генов, которая является одним из факторов эволюции.
СПЕРМАТОГЕНЕЗ И ОВОГЕНЕЗ
Сперматогенез - это процесс образования зрелых мужских половых клеток. Сперматозоиды развиваются в мужских половых железах (семенниках, или тестисах) из специализированных соматических клеток (рис. 19). Такими специализированными клетками служат так
называемые примордиальные зародышевые клетки, которые мигрируют к тестисам в раннем периоде эмбриогенеза мужского индивида. Следовательно, примордиальные клетки являются прародителями (предшественниками) зрелых половых клеток.
Семенники человека состоят из многочисленных канальцев, стенки которых сформированы слоями клеток, находящихся на разных стадиях развития сперматозоидов. Наружный слой канальцев образован крупными клетками, называемыми сперматогониями. Эти клетки содержат диплоидный набор хромосом и являются в тестисах потомками примордиальных зародышевых клеток. В период половой зрелости индивида часть сперматогоний перемещается во внутрен- ний слой канальцев, где в результате мейоза они развиваются в клетки, получившие название сперматоцитов I порядка (сперматоциты I), затем в сперматоциты II порядка (сперматоциты II) и, наконец,
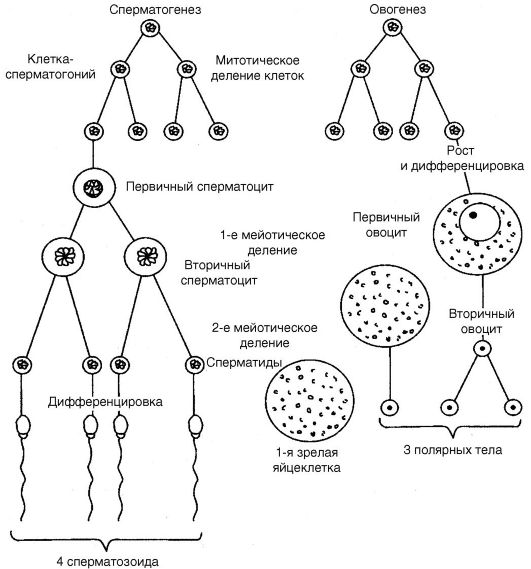
Рис. 19. Сперматогенез и овогенез
в сперматиды, являющиеся гаплоидными зародышевыми клетками, дифференцирующимися в конечном итоге в зрелые сперматозоиды. Таким образом, в обобщенном виде можно сказать, что сперматоге- нез инициируется в диплоидных соматических клетках (сперматогониях), после чего следует период созревания зародышевых клеток, в котором происходит два деления ядер путем мейоза, приводящего к образованию сперматид. Такова картина у человека.
Мейоз (рис. 20) в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I -- первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) - интерфаза II (интерокинез) - второе мейотическое деление (профаза I, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а свойства хромосом на разных стадиях. Интерфаза I
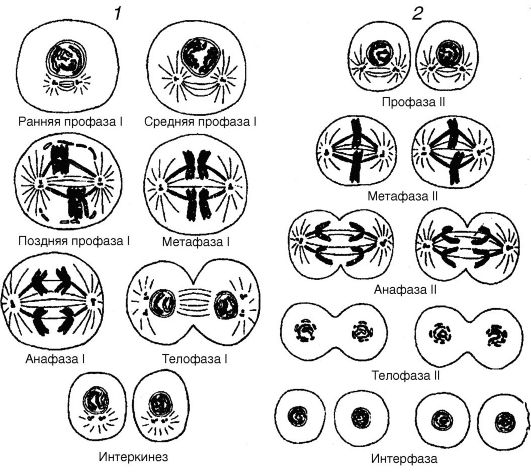
Рис. 20. Фазы мейоза: 1 - первое мейотическое деление; 2 - второе мейотическое деление
характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.
Первое мейотическое деление начинается в первичном сперматоците и характеризуется длинной профазой, которая состоит из переходящих одна в другую профазы I и профазы II. В профазе I различают пять главных стадий - лептонему, зигонему, пахинему, диплонему и диакинез.
На стадии лептонемы хромосомы в ядре представлены в виде тонких спирализованных нитей, содержащих многочисленные темноокрашенные гранулы (хромомеры). Расщепления хромомер и нитей не отмечают, но считают, что хромосомы на уровне этой стадии являются двойными, т. е. диплоидными. Гомологи каждой хромосомной пары объединяются хромомерами вдоль их длины по принципу застежки.
На стадии зигонемы характерно установление синапсов между гомологичными хромосомами, в результате чего образуются спарен- ные хромосомы (биваленты). Хромосомы X и Y ведут себя по сравнению с аутосомами несколько по-другому. Они конденсируются в темноокрашиваемые гетерохроматиновые тела, спариваемые в результате гомологических районов на их концах.
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии осуществляются обмены между гомологами и формирование хиазм.
На стадии диплонемы происходит укорачивание, утолщение и взаимное отталкивание сестринских хроматид, в результате чего хроматиды в биваленте почти разъединены. Разделение считается неполным по той причине, что в каждой паре хромосом еще не расщеплена центромера. Что касается бивалентов, то они удерживаются на различных местах вдоль их длины с помощью хиазм, которые являются структурами, сформированными между гомологичными хроматидами в результате
предыдущего кроссинговера между синаптически связанными гомологами. В хороших препаратах можно наблюдать от одной до нескольких хиазм в зависимости от длины бивалента. Каждая наблюдаемая на этой стадии хиазма представляет собой результат обмена, который встречался между несестринскими хроматидами в течение стадии пахинемы. Поскольку сжатие и отталкивание бивалентов усиливается, хиазмы двигаются к концам хромосом, т. е. происходит терминализация хромосом. В конце диплонемы наступает деспирализация хромосом; гомологи продолжают отталкиваться друг от друга.
На стадии диакинеза, которая сходна с диплонемой, продолжается укорочение бивалентов и наступает ослабление (уменьшение) хиазм, вследствие чего формируются дискретные единицы в виде хроматид (четыре). Непосредственно после завершения этой стадии происходит растворение ядерной мембраны.
В метафазе I биваленты достигают наибольшей концентрации. Становясь овальными, они располагаются в экваториальной части ядра, где формируют экваториальные пластинки мейотической метафазы I. Форма каждого бивалента определяется числом и локализацией хиазм. У мужчин число хиазм на бивалент в метафазе I составляет обычно 1-5. Бивалент XY становится палочковидным в результате одиночной терминально расположенной хиазмы.
В анафазе I начинается движение противоположных центромер к противоположным полюсам клетки. В результате этого происходит разделение гомологичных хромосом. Каждая хромосома состоит теперь из двух хроматид, удерживаемых центромерой, которая не делится и остается интактной. Этим анафаза I мейоза отличается от анафазы митоза, при котором центромера подвергается разделению. Важно заметить, что благодаря кроссинговеру каждая хроматида является генетически различной.
В стадии телофазы I хромосомы достигают полюсов, чем заканчивается первое мейотическое деление. После телофазы I наступает короткая интерфаза (интеркинез), в которой хромосомы деспи- рализуются и становятся диффузными, или телофаза I переходит прямо в профазу II второго меиотического деления. Ни в одном, ни в другом случае репликации ДНК не отмечается. После первого мейотического деления клетки называют сперматоцитами II порядка. Количество хромосом в каждой такой клетке снижается от 2n до 1n, но содержание ДНК еще не изменяется.
Второе мейотическое деление осуществляется в течение нескольких фаз (профаза II, метафаза II, анафаза II, телофаза II) и сходно с митотическим делением. В профазе II хромосомы вторичных спер- матоцитов остаются у полюсов. В метафазе II центромера каждой из двойных хромосом делится, обеспечивая каждую новую хромосому собственной центромерой. Начинается формирование веретена, к полюсу которого двигаются новые хромосомы. В телофазе II второе мейотическое деление заканчивается, в результате чего каждый сперматоцит II порядка дает два сперматида, из которых дифферен- цируются затем сперматозоиды. Как и во вторичном сперматоците, число хромосом в сперматиде является гаплоидным (1n). Однако хромосомы сперматид являются одиночными, тогда как хромосомы вторичных сперматоцитов II - двойными, будучи построенными из двух хроматид. Следовательно, ядро каждого сперматида имеет одиночный набор негомологичных хромосом. Вторичное мейотическое деление - это деление митотического типа (экваториальным делением). Оно разделяет двойные сестринские хроматиды и отличается от редукционного деления, в котором гомологичные хромосомы разделены. Единственное существенное отличие от классического митоза заключается в том, что здесь имеется гаплоидный набор хромосом.
Итак, первое мейотическое деление сперматоцитов I порядка приводит к образованию двух вторичных сперматоцитов (II порядка). Обе хроматиды структур, образующихся в результате редукционного деления, являются сестринскими хроматидами. Последние возникают в результате репликации, предшествующей первому мейотическому делению. Второе мейотическое деление каждого вторичного сперматоцита приводит к образованию четырех сперматидов. Таким образом, в типичном мейозе клетки делятся дважды, тогда как хромосомы - только один раз (рис. 21).
Конечный этап в сперматогенезе связан с дифференциацией, которая заканчивается тем, что каждый из сравнительно больших, сферически неподвижных сперматидов превращается в небольшой вытянутый подвижный сперматозоид.
У большинства взрослых (сексуально зрелых) мужских особей животных сперматогенез происходит в семенниках постоянно или периодически (сезонно). Например, у насекомых для завершения цикла сперматогенеза требуется лишь несколько дней, тогда как у млекопитающих этот цикл затягивается на недели и даже
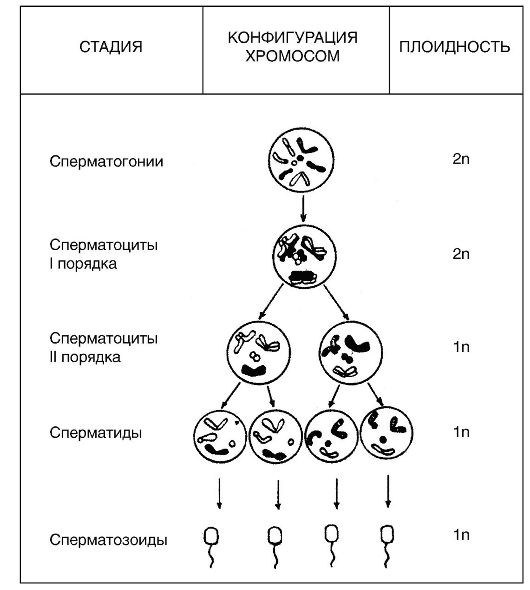
Рис. 21. Распределение хромосом при гаметогенезе
месяцы. У взрослого человека сперматогенез проходит в течение всего года. Время развития примитивных сперматогоний в зрелые сперматозоиды составляет около 74 дней.
Мужские половые клетки, продуцируемые организмами разной видовой принадлежности, характеризуются подвижностью и чрезвычайным разнообразием в размерах и по структуре (рис. 22). Каждый сперматозоид человека состоит из трех отделов - головки, средней части и хвоста (рис. 23). В головке сперматозоида располагается ядро, которое содержит гаплоидный набор хромосом.
Головка снабжена акросомой, которая содержит литические ферменты, необходимые содержимому сперматозоиду для вступления в яйцеклетку. В головке локализуются также две центриоли - прок-
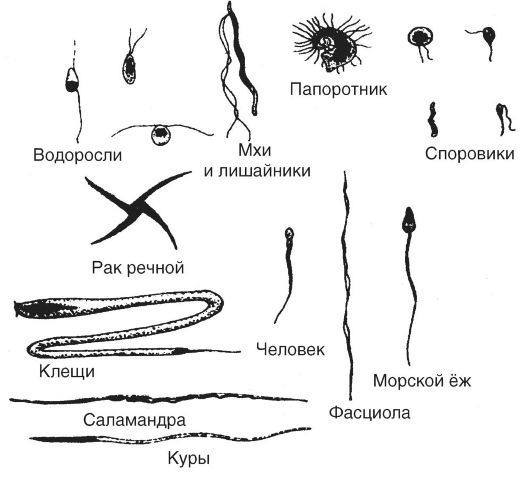
Рис. 22. Формы сперматозоидов

Рис. 23. Строение сперматозоида: А - светомикроскопическое изображение в разных плоскостях: 1 - головка, 2 - средняя часть, 3 - хвост; Б - схематическая реконструкция электронномикроскопического изображения: 1 - ядро, 2 - акросома, 3 - центросома (проксимальная центриоль), 4 - центральное кольцо, 5 - митохондриальная спираль,
6 - осевая нить
симальная, которая побуждает деление оплодотворенной сперматозоидом яйцеклетки, и дистальная, которая дает начало аксимальному стержню хвоста. В средней части сперматозоида располагаются базальное тельце хвоста и митохондрии. Хвост (отросток) сперматозоида образован внутренним аксимальным стержнем и внешним футляром, имеющим цитоплазматическое происхождение. Сперматозоиды человека характеризуются значительной подвижностью.
ОВОГЕНЕЗ И ЯЙЦЕКЛЕТКИ
Процесс формирования яйцеклеток носит название овогенеза. Его функциями являются обеспечение гаплоидного набора хромосом в ядре яйцеклетки и питательных потребностей зиготы. Овогенез в своем проявлении в основном сопоставим со сперматогенезом.
У млекопитающих и человека овогенез начинается еще во внутриутробном периоде (до рождения). Овогонии, представляющие собой мелкие клетки с довольно крупным ядром и локализованные в фолликулах яичников, начинают дифференцироваться в первичные овоциты. Последние формируются уже на третьем месяце внутриутробного развития, после чего они вступают в профазу первого мейотического деления. Ко времени рождения девочки все первичные овоциты уже находятся в профазе первого мейотического деле- ния. Первичные овоциты остаются в профазе вплоть до наступления половой зрелости женского индивида. В одном овоците содержится до 100 000 митоходрий. Когда при наступлении половой зрелости фолликулы яичника созревают, мейотическая профаза в первичных овоцитах возобновляется. Первое мейотическое деление для каждой развивающейся яйцеклетки завершается вскоре перед временем овуляции этой яйцеклетки. В результате первого мейотического деления и неравномерного распределения цитоплазмы одна образовавшаяся клетка становится вторичным овоцитом, другая - полярным (редукционным) тельцем.
Вторичное мейотическое деление наступает, когда вторичный овоцит (развивающаяся яйцеклетка) проходит из яичника в фаллопиеву трубу. Однако это деление не завершается до тех пор, пока во вто- ричный овоцит не проникнет сперматозоид, что происходит обычно в фаллопиевой трубе. Когда сперматозоид проникает во вторичный овоцит, то последний делится, в результате чего образуется овотида (зрелая яйцеклетка) с пронуклеоусом, содержащим одиночный набор
из 23 материнских хромосом. Другая клетка, образующаяся в результате этого деления, является вторым полярным тельцем, не способным к дальнейшему развитию. В это время подвергается делению надвое и полярное (редукционное) тельце. Таким образом, развитие одного овоцита I порядка сопровождается образованием одной овотиды и трех редукционных телец. В яичниках таким путем на протяжении жизни обычно созревает 300-400 овоцитов, но в месяц созревает лишь один овоцит. В течение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро.
У некоторых видов животных овогенез протекает быстро и непрерывно и приводит к образованию большого количества яйцеклеток.
Вопреки сходству со сперматогенезом овогенез характеризуется некоторыми специфическими особенностями. Питательный материал (желток) первичного овоцита не распределяется поровну между четырьмя клетками, которые образуются в результате мейотических делений. Основное количество желтка сохраняется в одной большой клетке, тогда как полярные тельца содержат очень мало этого вещества. Первые и вторые полярные тельца получают в результате делений те же самые хромосомные наборы, что и вторичные овоциты, но они не становятся половыми клетками. Поэтому яйцеклетки намного богаче питательным материалом по сравнению со сперматозоидами. Особенно сильно это различие проявляется в случае яйцекладущих животных.
Яйцеклетки млекопитающих имеют овальную или несколько вытянутую форму (рис. 24) и характеризуются типичными чертами клеточного строения. Они содержат все структуры, характерные для соматических клеток, однако внутриклеточная организация яйцеклетки очень специфична и определяется тем, что яйцеклетка является также и средой, которая обеспечивает развитие зиготы. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. У очень многих животных различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется еще на стадии овоцита. Представляя собой поверхностный слой овоцита, она имеет сложную структуру, так как пронизана выростами прилегающих к ней фолликулярных клеток. Вторичная (средняя) оболочка полностью сформирована фолликулярными клетками, а третичная (наружная) образована веществами, представляющими собой продукты секреции желез яйцеводов, через которые проходят яйцеклетки. У птиц, например,
третичными оболочками яйцеклеток служат белковая, подскорлуповая и скорлуповая оболочки. Для яйцеклеток млекопитающих характерно наличие двух оболочек. Структура внутриклеточных компонентов яйцеклеток специфична в видовом отношении, а иногда имеет даже индивидуальные особенности.
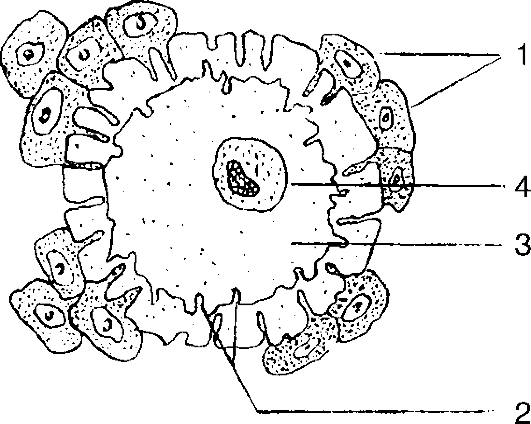
Рис. 24. Строение яйцеклетки: 1 - фолликулярные клетки; 2 - оболочка; 3 - цитоплазма; 4 - ядро
ОПЛОДОТВОРЕНИЕ
Оплодотворение - это процесс объединения мужской и женской гамет, который приводит к формированию зиготы и последующему развитию нового организма. В процессе оплодотворения происходит установление диплоидного набора хромосом в зиготе, что определяет выдающееся биологическое значение этого процесса.
В зависимости от видовой принадлежности организмов, размножающихся половым путем, различают наружное и внутреннее оплодотворение. Наружное оплодотворение происходит в окружающей среде, в которую поступают мужские и женские половые клетки. Например, наружным является оплодотворение у рыб. Выделяемые ими мужские (молока) и женские (икра) половые клетки поступают в воду, где и происходит их встреча и объединение.
Внутреннее оплодотворение обеспечивается переносом сперматозоидов из мужского организма в женский. Такое оплодотворение встречается у млекопитающих, центральным моментом которого является слияние ядер половых клеток. Считают, что в яйцеклетку проникает содержимое одного сперматозоида. В механизмах опло-
дотворения многое еще остается неясным. Данные об оплодотворении у морских ежей свидетельствуют о том, что уже через 2 секунды после соприкосновения сперматозоидов и яйцеклетки наступают изменения в электрических свойствах плазменной мембраны последней. Слияние гамет наступает через 7 секунд. Предполагают, что проникновение в яйцеклетку содержимого лишь одного из множества сперматозоидов объясняется изменениями электрических свойств ее плазменной мембраны. Оплодотворенная яйцеклетка дает начало зиготе. По вопросу о причинах активации сперматозоидом метаболизма яйцеклетки существует два мнения. Одни считают, что связывание сперматозоида с внешними рецепторами на поверхности клеток представляет собой сигнап, который через мембрану поступает вовнутрь яйцеклетки и активизирует там инозитолтрифосфат и ионы кальция. Другие полагают, что сперматозоиды содержат специальный инициирующий фактор.
Экспериментальные разработки, выполненные в последние годы, показали, что оплодотворение яйцеклеток млекопитающих, включая человека, возможно и в пробирке. Болеее того, зародыши, развившиеся в пробирке, могут быть имплантированы в матку женщины, где они подвергаются дальнейшему нормальному развитию. Известны многочисленные случаи рождения «пробирочных» детей.
В отличие от животных, размножающихся путем зигогенеза, многие организмы способны к размножению путем партеногенеза (от греч. parthenos - девственница и genos - рождение), под которым понимают размножение организмов из неоплодотворенной яйцеклетки. Различают облигатный и факультативный партеногенез. Облигатный партеногенез стал главным способом размножения организмов отдельных видов, например кавказской скальной ящерицы. Животные этого вида являются только женскими особями. Напротив, факультативный партеногенез заключается в том, что яйцеклетки способны развиваться как без оплодотворения, так и после оплодотворения. Факультативный партеногенез, в свою очередь, бывает женским и мужским. Женский партеногенез часто наблюдается у пчел, муравьев, коловраток, у которых из неоплодотворенных яйцеклеток развиваются самцы. Мужской партеногенез встречается у некоторых изогамных водорослей.
Партеногенез бывает как естественным, так и искусственным (индуцированным). Механизм искусственного партеногенеза заключается в раздражении яйцеклеток с помощью физических или хими-
ческих факторов, что ведет к активации яйцеклеток и, как следствие, к развитию неоплодотворенных яиц. Искусственный партеногенез наблюдали в случае животных многих систематических групп - иглокожих, червей, моллюсков и даже млекопитающих.
Известна форма партеногенеза, получившая название андрогенеза (от греч. andros - мужчина, genesis - зарождение). Если в яйцеклетке инактивировать ядро и после этого в нее проникнет несколько сперматозоидов, то из такой яйцеклетки в результате слияния мужских (сперматозоидных) ядер развивается мужской организм.
Известны случаи, когда партеногенез проявляется циклически, в зависимости от сезона. Например, коловратки, дафнии и тли летом размножаются путем партеногенеза, осенью - оплодотворения яйце- клеток и образования зигот, т. е. путем зигогенеза.
Роль партеногенеза и его форм в природе невелика, так как он не обеспечивает широких адаптивных возможностей организмов.
В отличие от зигогенеза и партеногенеза существует гиногенез, представляющий собой псевдогамию, когда сперматозоид вступает в яйцеклетку и активизирует ее, но ядро сперматозоида не сливается с ядром яйцеклетки. В этом случае появляющееся потомство состоит только из женских особей. Гиногенез встречается в естественных условиях у нематод и рыб, но его можно вызвать и искусственно. В частности, описаны случаи искусственного гиногенеза у тутового шелкопряда, рыб и амфибий.
ЧЕРЕДОВАНИЕ ГАПЛОИДИИ И ДИПЛОИДИИ.
ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Организмам, размножающимся половым путем, свойственно чередование гаплоидной и диплоидной фаз в их развитии. У многих организмов, включая млекопитающих, это чередование имеет регулярный характер, и на нем основано сохранение видовых признаков организмов.
Для многих организмов характерна также смена поколений, когда поколения особей, размножающихся бесполым путем, сменяются поколениями особей, размножающихся половым путем с образованием гамет. Это получило название первичной смены поколений. Она встречается у споровиков, жгутиконосцев и многих растений. Первичная смена поколений имеет регулярный характер, и ее нали-
чие в природе свидетельствует о сохранении в филогенезе многих организмов как бесполого, так и полового размножения. У других организмов встречается чередование полового размножения с парте- ногенезом. Это называют вторичной сменой поколений. Например, у трематод половое размножение регулярно сменяется партеногенезом. Такое вторичное чередование поколений называют гетерогонией. У кишечнополостных на некоторых стадиях развития осуществляется переход от полового размножения к бесполому (вегетативному). Эту форму вторичного чередования поколений называют метагенезом.
ПОЛОВОЙ ДИМОРФИЗМ. ГЕРМАФРОДИТИЗМ
Для мужских и женских особей характерны специфические фенотипические черты. Различия между самками и самцами по их свойствам называют половым диморфизмом. У животных он встречается уже на низших ступенях эволюционного развития, например у круглых гельминтов, членистоногих, и достигает наибольшего выражения у позвоночных животных.
Если мужские и женские половые клетки продуцируются одной и той же особью, имеющей как мужские, так и женские половые железы, то это явление носит название истинного гермафродитизма. Он встречается у плоских червей, кольчатых червей, моллюсков. У плоских червей мужские и женские половые железы функционируют на протяжении всей жизни особей. Напротив, у моллюсков половые железы продуцируют яйцеклетки и сперматозоиды попеременно.
Истинный гермафродитизм встречается и у человека, являясь результатом нарушения развития. Генотипами гермафродитов являются 46 XX или 46 XY, причем большинство случаев относится к XX (около 60%). Генотип XX чаще всего встречается у гермафродитов негроидных афри- канских популяций, тогда как XY в большинстве случаев - среди японцев. У гермафродитов обоих типов отмечена тенденция в сторону билатеральной асимметрии гонад. Среди истинных гермафродитов встречаются также хромосомные мозаики, которые в одних соматических клетках имеют пару хромосом XX, в других - пару XY.
Известен и ложный гермафродитизм, когда индивиды обладают наружными половыми органами и вторичными половыми признака- ми, характерными для обоих полов, но продуцируют лишь половые клетки одного типа - мужские или женские.
ПРОИСХОЖДЕНИЕ СПОСОБОВ РАЗМНОЖЕНИЯ
Предполагают, что наиболее древним является бесполое размножение, в частности вегетативное размножение. Из последнего развилось размножение спорообразованием, несомненное преимущество которого заключается в том, что оно обеспечивает лучшие возможности в сохранении видов и особенно в их расселении.
Половое размножение - наиболее эффективный путь воспроизводства организмов. Предполагают, что оно развилось из бесполого, возникнув около 1 млрд лет назад, причем первые этапы были связаны с усложнением в развитии гамет. Примитивные гаметы характеризовались недостаточной морфологической дифференцировкой, в результате чего для многих организмов ведущей была изогамия (от греч. isos - равный, gamos - брак), когда половые клетки были подвижными изогаметами, не дифференцированными на мужские и женские формы.
В последующем получила развитие анизогамия (от греч. anisos - неравный, gamos - брак), характеризующаяся наличием дифференцированных гамет, различающихся лишь величиной. На более поздних этапах эволюции возникли резкие различия в подвижности, форме и размерах гамет. В процессе эволюции у позвоночных выработался также ряд добавочных приспособлений, облегчающих перенос спермы самца в половые пути самки и создающих условия для развития оплодотворенной яйцеклетки. Эти приспособления в ходе эволюции развились из выделительной системы, что привело к формированию мочеполовой системы.
Диплоидное состояние предоставляет чрезвычайные преимущества для организмов, поскольку в этом состоянии происходит накопление различных аллелей. Поэтому половое размножение имеет еще и то преимущество, что оно предоставляет организмам большую возможность изменчивости по сравнению с бесполым, а это играет важнейшую роль в эволюции.
Явные репродуктивные преимущества имеет партеногенез, так как он продуцирует лишь потомство женского рода. Однако он редок. Для объяснения низкой частоты замещения партеногенезом полового размножения в естественных популяциях организмов используют две гипотезы. В соответствии с одной из них (мутационно-аккумулятивной) пол является адаптивным приспособлением, поскольку он «очищает» геном от повторяющихся во времени мутаций, тогда как по другой гипотезе (экологической) пол является
адаптивным приспособлением в меняющихся условиях среды, в частности, когда вариации организмов генерируются коэволюционным взаимодействием с паразитами, которые способствуют сохранению пола. В то же время наметилась тенденция считать обе гипотезы справедливыми.
Первые
представления о росте и развитии восходят к античному миру. Еще
Гиппократ (460-377 гг. до н. э.) предполагал, что яйцеклетки уже
содержат полностью сформированный организм, но в очень уменьшенном
виде. Это представление затем было развито в учение о преформизме (от
лат. preformatio - предобразование), которое особенно популярным
оказалось в XVII-XVIII вв. Сторонниками преформизма были Гарвей,
Мальпиги и многие другие видные биологи и медики того времени. Для
преформистов спорный вопрос заключался лишь в том, в каких половых
клетках преформирован орга- низм - женских или мужских. Тех, кто
отдавал предпочтение яйцеклеткам, называли овистами, а тех, кто большее
значение придавал мужским половым клеткам, - анималькулистами.
Преформизм - это метафизическое учение от начала и до конца, ибо оно
отрицало развитие. Решающий удар преформизму нанес Ш. Боннэ
(1720-1793), который открыл в
В античном мире возникло и другое учение, противоположное преформизму и получившее впоследствии название эпигенеза (от греч. epi - после, genesis - развитие).
Как и преформизм, эпи- генез большое распространение получил также в
XVII-XVIII вв. В распространении эпигенеза большое значение имели
взгляды К.Ф. Вольфа (1733-1794), обобщенные в его книге «Теория
развития» (1759). К.Ф. Вольф считал, что в яйце нет ни
преформированного организма, ни его частей и что оно состоит из первона-
чально однородной массы. В отличие от преформистов взгляды К.Ф. Вольфа
и других сторонников эпигенеза были прогрессивны применительно для
своего времени, так как содержали мысль о развитии. Однако в дальнейшем
появились новые моменты. В частности, в
Рост и развитие - важнейшие свойства живого. Рост - это повышение массы всего организма в результате увеличения количества клеток, тогда как развитие - это качественные изменения организма, которые определяются дифференциацией клеток и морфогенезом, обеспечивающими прогрессивные изменения индивидуумов, начиная с яйцеклеток и заканчивая их взрослым состоянием.
Онтогенез (от греч. ontos - существо, genesis - развитие) - это история (цикл) развития индивидуума, начинающаяся с обра- зования давших ему начало половых клеток и заканчивающаяся его смертью. Представления об онтогенезе основаны на данных о росте, развитии и дифференцировке. Изучение фундаментальных основ онтогенеза имеет важное значение для понимания биологии человека.
Современные данные о развитии организмов отвергают как преформизм, так и эпигенез. В рамках современных представлений развитие организма понимают в качестве процесса, при котором структуры, образовавшиеся ранее, побуждают развитие последующих структур. Процесс развития детерминирован генетически и теснейшим образом связан со средой. Следовательно, развитие определяется единством внутренних и внешних факторов.
ЕДИНСТВО РОСТА И РАЗВИТИЯ
Рост организма - это постепенное возрастание его массы и изменение формы в результате увеличения количества клеток и их дифференцировки, формирования тканей и органов, биохимических изменений в клетках и тканях. Таким образом, рост является результатом количественных изменений в виде увеличения количества клеток (массы тела) и качественных в виде дифференцировки клеток и морфогенеза. Дифференциация клеток - это процесс, благодаря которому одни клетки становятся морфологически, биохимически и функционально отличающимися от других клеток. Размножение и дифференцировка одних клеток всегда координированы с ростом и дифференцировкой других. Оба эти процесса происходят на протяжении всего жизненного цикла организма. Поскольку дифференцирующиеся клетки изменяют свою форму, а в изменения формы вовлекаются группы клеток, то это сопровождается морфогенезом, представляющим собой совокупность процессов, определяющих структурную организацию клеток и тканей, а также общую морфологию организмов.
Рост можно измерить, построив на основе результатов измерений кривые размеров организма, массы, сухой массы, количества клеток, содержания азота и других показателей.
ОНТОГЕНЕЗ И ЕГО ТИПЫ. ПЕРИОДИЗАЦИЯ ОНТОГЕНЕЗА
Онтогенез в зависимости от характера развития организмов типируют на прямой и непрямой, в связи с чем различают прямое и непрямое развитие. Прямое развитие организмов в природе встречается в виде неличиночного и внутриутробного развития, тогда как непрямое развитие наблюдается в форме личиночного развития. В противоположность онтогенезу видовой категорией является филогенез.
Личиночное развитие. Под этим развитием понимают непрямое развитие, поскольку организмы в своем развитии имеют одну или несколько личиночных стадий. Личиночное развитие характерно для насекомых, амфибий, иглокожих. Личинки этих животных ведут самостоятельный образ жизни, подвергаясь затем превращениям. Поэтому это развитие называют еще развитием с метаморфозами (см. ниже).
Неличиночное развитие. Такая форма развития характерна для организмов, развивающихся прямым образом, например для рыб, пресмыкающихся и птиц, яйца которых богаты желтком (питательным материалом). Благодаря этому в яйцах, откладываемых во внешнюю среду, проходит значительная часть онтогенеза. Метаболизм зародышей обеспечивается развивающимися провизорными органами, представляющими собой зародышевые оболочки (желточный мешок, амнион, аллантоис).
Внутриутробное развитие. Это развитие также характерно для организмов, развивающихся прямым путем, например для млекопитающих, включая человека. Поскольку яйцеклетки этих организмов очень бедны питательными веществами, то все жизненные функции зародышей обеспечиваются материнским организмом посредством образования из тканей матери и зародыша провизорных органов, среди которых главным является плацента. Эволюционно внутриутробное развитие - самая поздняя форма, однако оно наиболее выгодно для зародышей, так как эффективно обеспечивает их выживание.
Онтогенез подразделяют на проэмбриональный, эмбриональный и постэмбриональный периоды. В случае человека период развития до рождения называют пренатальным или антенатальным, после рождения - постнатальным. Развивающийся зародыш до образования зачатков органов называют эмбрионом, после образования зачатков органов - плодом.
ПРОЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Этот период в индивидуальном развитии организмов связан с образованием гамет в процессе гаметогенеза. Мужские половые клетки не имеют существенных отличий от других клеток, тогда как яйцеклетки отличаются тем, что они содержат очень много желтка. Учитывая количество желтка и распространение его в яйцеклетках, последние классифицируют на три типа:
1) изолециталъные яйцеклетки, содержащие немного желтка, который локализован равномерно по всей клетке. Эти яйцеклетки продуцируются иглокожими (морскими ежами), низшими хордовыми (ланцетниками), млекопитающими;
2) телолецитальные яйцеклетки содержат большое количество желтка, который сосредоточен на одном из полюсов - вегетативном. Такие яйцеклетки продуцируются моллюсками, земноводными, реп- тилиями, птицами. Например, яйцеклетки лягушки состоят на 50% из желтка, яйцеклетки кур (в обиходе куриные яйца) - на 95%. На другом полюсе (анимальном) телолецитальных яйцеклеток сосредоточены цитоплазма и ядро;
3) центролецитальные яйцеклетки, в которых желтка немного и он занимает центральное положение. На периферии таких яйцеклеток располагается цитоплазма. Центролецитальные яйцеклетки проду- цируются членистоногими.
Для проэмбрионального периода характерно также то, что в этот период в гаметах происходят метаболические процессы, связанные с накоплением молекул ДНК.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Эмбриогенез (от греч. етbryоп - зародыш), или эмбриональный период, начинается со слияния мужских и женских половых клеток, которые представляют собой процесс оплодотворения яйцеклеток.
У организмов, для которых характерно внутриутробное развитие, эмбриональный период заканчивается рождением, а у организмов, которым свойственны личиночный и неличиночный типы рзвития, эмбриональный период завершается выходом организма из яйцевых или зародышевых оболочек соответственно. В пределах эмбрионального периода различают стадии зиготы, дробления, бластулы, образования зародышевых листков, гистогенеза и органогенеза.
Зигота. Оплодотворение заключается в серии процессов, в которых мужская половая клетка инициирует развитие яйцеклетки. В активированной мужской гаметой яйцеклетке происходит ряд физических и химических процессов, включая повышенный синтез белков. Перемещение протоплазмы ведет к установлению билатериальной симметрии яйцеклетки. Ядра сливаются, восстанавливается диплоидный набор хромосом. Таким образом создается одноклеточный организм.
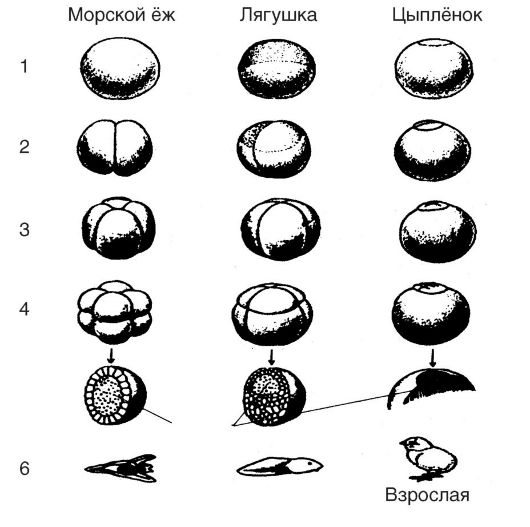
Дробление. Представляет собой начальный период развития зиготы (оплодотворенного яйца), которое заключается в делении зиготы путем митоза. Деление начинается с появления на поверхности яйцеклетки борозды. Первая борозда приводит к образованию двух клеток - двух бластомеров, вторая - четырех бластомеров, третья - восьми бластомеров (рис. 25). Группа клеток, образованная в результате последовательных дроблений, получила название морулы (от лат. morum - тутовая ягода).
Биологическое значение этой стадии заключается в том, что из крупной клетки, являющейся яйцеклеткой, образуются более мелкие клетки, в которых уменьшено отношение цитоплазмы к ядру.
Дробление зиготы завершается образованием многоклеточной структуры, получившей название бластулы (от греч. blastos - росток). Эта структура имеет форму пузырька, называемого бластодермой и состоящего из одного слоя клеток. Теперь эти клетки называют эмбриональными. По размерам бластула сходна с яйцеклеткой. В период дробления увеличивается количество ядер и общее число ДНК. Синтезируется также небольшое количество мРНК и тРНК, тогда как рибосомная РНК еще не обнаруживается.
Стадию бластулы проходят все животные, но в каждом случае есть особенности. У млекопитающих деление идет неравномерно, поэтому морулы состоят из разного количества клеток. Кроме того, из части клеток образуется структура, называемая трофобластом,
5
Бластоцель Личинка Головастик
форма
Рис. 25. Дробление зигот и образование бластул у разных организмов: 1 - исходная яйцеклетка; 2 - два бластомера; 3 - четыре бластомера; 4 - восемь бластомеров; 5 - бластула; 6 - взрослая форма
клетки которого питают зародыш и благодаря ферментам обеспечивают внедрение последнего в стенку матки. Позднее клетки трофобласта отслаиваются от зародыша и образуют пузырек, который заполняется жидкостью тканей матки.
Биологическое значение этой стадии заключается в том, что из крупной клетки, каковой является яйцеклетка, образуются более мелкие клетки, в которых уменьшено отношение цитоплазмы к ядру и ядро имеет новое цитоплазматическое окружение.
Гаструляция (от греч. gastre - полость сосуда). Это следующий за образованием бластулы процесс движения эмбриональных клеток, который сопровождается формированием двух или трех (в зависимости от вида животных) слоев зародыша, или так называемых зародышевых листков (рис. 26).
Развитие (гаструляция) изолецитальных яиц происходит путем инвагинации (впячивания) вегетативного полюса вовнутрь бластулы, в результате чего противоположные полюса почти сливаются, а бластоцель (полость бластулы) почти либо полностью исчезает. Внешний
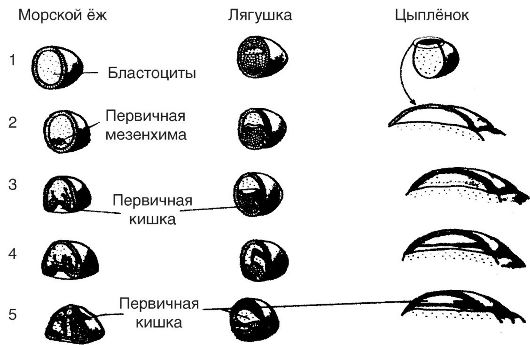
Рис. 26. Гаструляция у различных организмов: 1 - бластоцель; 2 - первичная мезенхима; 3, 4, 5 - первичная кишка
слой клеток зародыша получил название эктодермы (от греч. ectos - снаружи, derma - кожа), или наружного зародышевого листка, тогда как внутренний - энтодермы (от греч. entos - внутри), или внутреннего зародышевого листка. Образующаяся при этом полость получила название гастроцеля, или первичной кишки, вход в которую называют бластопором (первичным ртом).
Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако хордовым в период гаструляции присуще развитие третьего зародышевого листка - мезодермы (от греч. mesos - средний), образующегося между эктодермой и энтодермой.
Гаструляция является необходимым пререквизитом для последующих стадий развития, поскольку она приводит клетки в положение, открывающее возможность формировать органы. Дифференцированный на три эмбриональных закладки зародышевый материал дает начало всем тканям и органам развивающегося зародыша.
Развитие (дифференцировка) зародышевых листков сопровождается тем, что из них формируются различные ткани и органы. В частности, из эктодермы развиваются эпидермис кожи, ногти и волосы, сальные и потовые железы, нервная система (головной мозг, спинной мозг, ганглии, нервы), рецепторные клетки органов чувств, хрусталик глаза, эпителий рта, носовой полости и анального отверстия, зуб-
ная эмаль. Из энтодермы развиваются эпителий пищевода, желудка, кишок, желчного пузыря, трахеи, бронхов, легких, мочеиспускательного канала, а также печень, поджелудочная железа, щитовидная, паращитовидные и зобная железы. Из мезодермы развиваются гладкая мускулатура, скелетные и сердечные мышцы, дерма, соединительная ткань, кости и хрящи, дентин зубов, кровь и кровеносные сосуды, брыжейка, почки, семенники и яичники. У человека первыми обособляются головной и спинной мозг. Через 2 месяца появляются почти все структуры тела. Связь зародыша со средой осуществляется через провизорные органы. Органогенез заканчивается к концу эмбрионального периода. Если дефинитивное ротовое отверстие образуется на месте первичного рта (бластопора), то этих животных называют первичноротыми (черви, моллюски, членистоногие).
Если же дефинитивный рот образуется в противоположном месте, то этих животных называют вторичноротыми (иглокожие, хордовые).
Для обеспечения связи зародыша со средой служат так называемые провизорные органы, которые существуют временно. В зависимости от типа яйцеклеток провизорными органами являются разные структуры. У рыб, рептилий и птиц к провизорным органам относится желточный мешок. У млекопитающих желточный мешок закладывается в начале эмбриогенеза, но не развивается. Позднее он редуцируется. Наружная оболочка эмбриона называется хорионом. Она врастает в матку. Место наибольшего врастания в матку называют плацентой. Зародыш с плацентой связан через пуповину, или пупочный кантик, в котором имеются кровеносные сосуды, обеспечивающие плацентарное кровообращение. Метаболизм плода обеспечивается через плаценту.
В основе формообразующего взаимодействия частей эмбриона лежат определенным образом скоординированные процессы обмена веществ. Закономерностью развития является гетерохронность, под которой понимают разное во времени образование закладок органов и различную интенсивность их развития. Быстрее развиваются те органы и системы, которые должны раньше начать функционировать. Например, у человека зачатки верхних конечностей развиваются быстрее, чем нижних.
Зародыши чрезвычайно чувствительны к разным воздействиям. Поэтому различают критические периоды, т. е. периоды, когда зародыши наиболее чувствительны к повреждающим факторам. В случае человека критическими периодами эмбрионального онтогенеза
являются первые дни после оплодотворения, время образования плаценты и роды.
Ядра соматических клеток способны обеспечивать нормальное развитие яйцеклеток, что было выяснено в экспериментах по пересадке ядер соматических клеток в яйцеклетки, лишенные ядер.
Эксперименты также показали, что трансформация отдельного бластомера из 8- и 16-клеточных эмбрионов овец одной породы в безъядерную половину яйцеклетки (после рассечения последней пополам) другой породы сопровождалась развитием жизнеспособных эмбрионов и рождением ягнят.
Со времен Гиппократа (V в. до н. э.) обсуждается вопрос о причинах, которые инициируют рождение плода. В частности, сам Гиппократ предполагал, что развитие плода инициирует собственное рождение. Новейшие экспериментальные работы английских исследователей, выполненные на овцах, показали, что у овец инициация окотов контролируется комплексом гипоталамус + гипофиз + надпочечники плодов. Повреждение ядер гипоталамуса, удаление передней доли гипофиза или надпочечников пролонгирует беременности овец. Напротив, введение овцам аденокортикотропного гормона (секрета передней доли гипофиза) или кортизола (секрета надпочечников) сокращает сроки их беременностей.
Довольно частым нарушением развития является разделение зародыша очень ранней стадии развития, что сопровождается развитием однояйцевых (монозиготных) близнецов (рис. 27). Известны и так называемые сиамские близнецы, представляющие собой неразделенные организмы. Неразделенность встречается разной - от незначительного соединения до почти полного слияния двух организмов с разделенными головами или ногами. Иногда из двух сиамских близнецов один нормален, но другой чрезвычайно изменен, будучи прикрепленным к нормальному, являясь, по существу, паразитом.
Итак, в процессе развития высших эукариотов одиночная оплодотворенная клетка-зигота в ходе дальнейшего развития в результате митоза дает начало клеткам разных типов - эпителиальным, нервным, костным, клеткам крови и другим, которые характеризуются разнообразием морфологии и макромолекулярного состава. Однако для клеток разных типов характерно и то, что они содержат одинаковые наборы генов, но являются высокоспециализированными, выполняя лишь одну или несколько специфических функций, т. е. одни гены «работают» в клетках, а другие неактивны. Например,
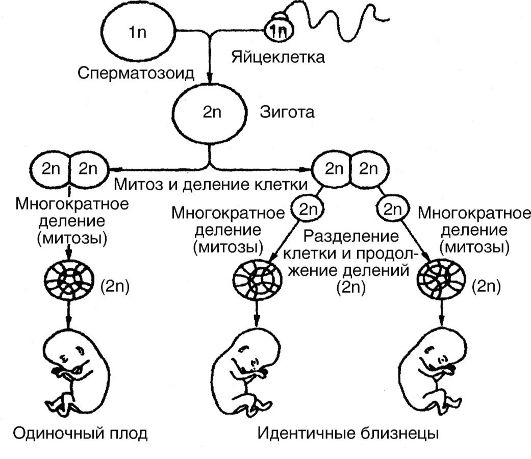
Рис. 27. Развитие монозиготных близнецов
только эритроциты специфичны в синтезе и хранении гемоглобина. Точно так лишь клетки эпидермиса синтезируют кератин. Поэтому давно возникли вопросы о генетической идентичности ядер соматических клеток и о контрольных механизмах развития оплодотворенных яйцеклеток как пререквизита в познании механизмов, лежащих в основе дифференцировки клеток. В дифференциации клеток имеют большое значение стволовые клетки (см. ниже). В экспериментах на мышах показано, что клетки одного типа способны к конверсии в клетки другого типа. В частности, показано, что дифференцирующиеся или дифференцированные клетки печени конвертируются в клетки поджелудочной железы.
Начиная с 50-х гг. XX в. во многих лабораториях были выполнены эксперименты по успешной пересадке ядер соматических клеток в яйцеклетки, искусственно лишенные собственных ядер. Исследование ДНК из ядер разных дифференцированных клеток показало, что почти во всех случаях геномы содержат одинаковые наборы последовательностей нуклеотидных пар. Известны исключения, когда эритроциты млекопитающих теряют свои ядра в течение последней стадии дифференцировки. Но к этому времени пулы стой-
ких гемоглобиновых мРНК уже синтезированы, так что ядра больше не нужны эритроцитам. Другими примерами служат гены иммуно- глобулинов и Т-клеток, модифицируемые в ходе развития.
Одним из крупных этапов в направлении познания контрольных механизмов эмбрионального онтогенеза стали результаты экспериментов, выполненных в 1960-1970 гг. английским исследователем Д. Гердоном с целью выяснить, обладают ли ядра соматических клеток способ- ностью обеспечивать дальнейшее развитие яйцеклеток, если эти ядра ввести в яйцеклетки, из которых предварительно удалены собственные ядра. На рис. 28 приведена схема одного из этих экспериментов, в котором ядра соматических клеток головастика пересаживали в яйцеклетки лягушки с предварительно удаленными ядрами. Эти эксперименты показали, что ядра соматических клеток действительно могут обеспечивать дальнейшее развитие яйцеклеток, так как они оказались способными оплодотворять яйцеклетки и «заставляли» их развиваться дальше. Этим была доказана возможность клонирования животных.
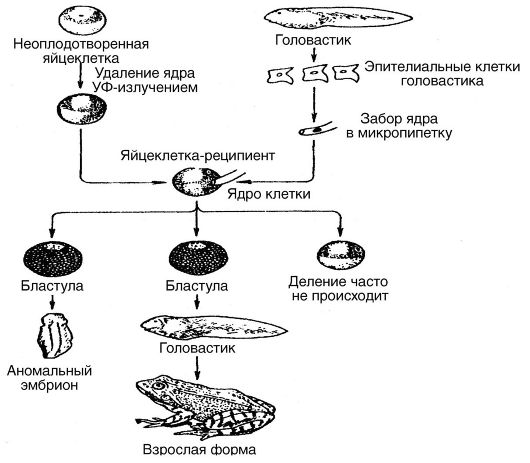
Рис. 28. Схема эксперимента по пересадке ядер соматических клеток в безъядерные яйцеклетки (Д. Гердон, 1968)
Позднее другими исследователями были выполнены эксперименты, в которых показано, что перенос отдельных бластомеров из 8- и 16-дневных эмбрионов овец одной породы в безъядерную половину яйцеклетки (после рассечения последней пополам) другой породы сопровождался формированием жизнеспособных эмбрионов с последующим рождением ягнят.
В начале
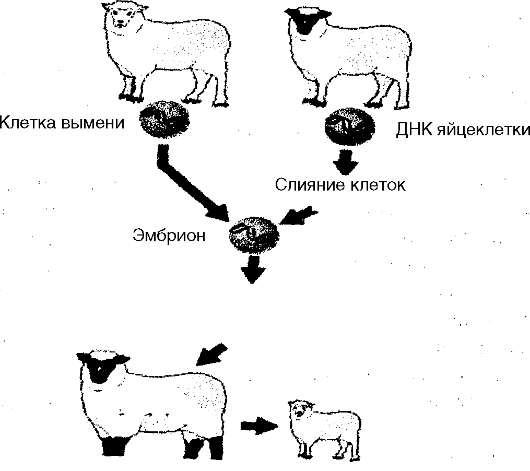
Введение культуры в организм матери
Рис. 29. Пересадка ядер соматических клеток в безъядерные яйцеклетки
Оценка этих результатов показывает, что млекопитающих можно размножать асексуальным путем, получая потомство животных, клетки которых содержат ядерный материал отцовского или материнского происхождения в зависимости от пола овцы-донора. В таких клетках
лишь цитоплазма и митохондрии имеют материнское происхождение. Это заключение имеет чрезвычайно важное общебиологическое значение, расширяет наши взгляды на потенциал размножения животных. Но важно также добавить, что речь идет о генетических манипуляциях, которые в природе отсутствуют. С другой стороны, в практическом плане эти генетические манипуляции представляют собой прямой путь клонирования высокоорганизованных животных с заданными свойствами, что имеет важное экономическое значение. В медицинском плане этот путь, возможно, будет использован в будущем для преодоления мужского бесплодия.
Итак, генетическая информация, необходимая для нормального развития эмбриона, не теряется в течение дифференцировки клеток за счет так называемых стволовых клеток, которые обладают потенцией развиваться в различные типы клеток тела. Когда стволовые клетки делятся, каждая новая клетка имеет потенцию или оставаться стволовой клеткой, или стать клеткой с более специализированной функцией (мышечные клетки, клетки крови или клетки головного мозга). Оплодотворенная яйцеклетка является тотипотентной, так как дает начало разным типам клеток тела. Тотипотентные стволовые клетки могут дать начало любому типу клеток, кроме тех, которые необходимы для развития плода. Стволовые клетки, которые могут дать начало самым различным клеточным типам, обычно называют мультитотипотентными клетками. Установлено, что взрослые стволовые клетки могут продуцировать дифференцированные клетки из эмбриональных несвязанных тканей. Соматические клетки тоже обладают свойством, получившим название тотипотентности, т. е. в их геноме содержится вся информация, полученная ими от оплодотворенной яйцеклетки, давшей им начало в результате дифференциации. Наличие этих данных с несомненностью означает, что дифференциация клеток подвержена генетическому контролю. Изучение стволовых клеток имеет значение для медицины.
Установлено, что интенсивный белковый синтез, следующий за оплодотворением, у большинства эукариотов не сопровождается синтезом мРНК. Изучение овогенеза у позвоночных, в частности у амфибий, показало, что интенсивная транскрипция происходит еще в течение профазы I (особенно диплонемы) мейоза. Поэтому генные транскрипты в форме молекул мРНК или про-мРНК сохраняются в яйцеклетках в бездействующем состоянии. Установлено, что у эмбриональных клеток имеет место так называемое асимме-
тричное деление, заключающееся в том, что деление эмбриональной клетки дает начало двум клеткам, из которых лишь одна наследует белки, принимающие участие в транскрипции. Таким образом, неравное распределение транскрипционных факторов между дочерними клетками ведет к экспрессии в них разных наборов генов после деления, т. е. к дифференциации клеток.
У амфибий и, возможно, у большинства позвоночных генетические программы, контролирующие раннее развитие (до стадии бластулы), устанавливаются еще в течение овогенеза. Более поздние стадии развития, когда начинается клеточная дифференциация (примерно, со стадии гаструлы), нуждаются в новых программах для экспрессии генов. Таким образом, дифференцировка клеток связана с перепрограммированием генетической информации в том или ином направлении.
Характерная особенность дифференцировки клеток заключается в том, что она необратимо ведет к тому или иному типу клеток. Этот процесс носит название детерминации и также находится под генетическим контролем, а как сейчас предполагают, дифференциация и детерминация клеток регулируются взаимодействием клеток на основе сигналов, осуществляемых пептидными ростовыми факторами через тирозинкиназные рецепторы. Вероятно, существует много таких систем. Одна из них заключается в том, что дифференциация мышечных и нервных клеток регулируется нейрорегулина- ми, представляющими собой мембранные белки, действующие через один или более тирозинкиназный рецептор.
Генетический контроль детерминации демонстрируется также существованием так называемых гомейотропных или гомеозисных мутаций, которые, как показано у насекомых, вызывают изменения при детерминации в специфических имагинальных дисках. В результате некоторые части тела развиваются не на своих местах. Например, у дрозофил мутации трансформируют детерминацию антенного диска в диск, который развивается в аппендикс конечности, протянутой от головы. У насекомых из рода Ophthalmoptera структуры крыльев могут развиваться из диска для глаз. У мышей показано существование генного кластера (комплекса) Нох, который состоит из 38 генов и контролирует развитие конечностей.
Самостоятельное значение имеет вопрос о регуляции активности генов в период эмбрионального развития. Считают, что в ходе дифференцировки гены действуют в разное время, что выражается
в транскрипции в различных дифференцированных клетках разных мРНК, т. е. имеют место репрессия и дерепрессия генов. Например, количество генов, транскрибируемых в РНК в бластоцитах морского ежа, равно 10%, в клетках печени крыс - тоже 10%, а в клетках тимуса крупного рогатого скота - 15%. Предполагают, что в контроле транскрипционного статуса генов принимают участие негистоновые белки. В пользу этого предположения свидетельствуют следующие данные. Когда хроматин клеток в фазе S транскрибируется в системе in vitro, то синтезируется только гистоновая мРНК, а в след за нею и гистоны. Напротив, когда используют хроматин клеток из (^-фазы, то никакой гистоновой мРНК не синтезируется. Когда же непистоновые белки удаляются из хроматина (1-фазы и замещаются негистоновыми хромосомными белками, синтезированными в фазе S, то после транскрипции такого хроматина in vitro синтезируется гистоновая мРНК. Более того, когда негистоновые белки происходят из (1-фазы клеток, а ДНК и гистоны - из S-фазы клеток, никакой гистоновой мРНК не синтезируется. Эти результаты показывают, что содержащиеся в хроматине негистоновые белки определяют возможность транскрипции генов, кодирующих гистоны. Поэтому считают, что негистоновые хромосомные белки могут играть важную роль в регуляции и экспрессии генов у эукариот.
Имеющиеся данные позволяют считать, что в регуляции транскрипции у животных принимают участие белковые и стероидные гормоны. Белковый (инсулин) и стероидные (эстрогон и тестостерон) гормоны представляют собой две сигнальные системы, используемые в межклеточных коммуникациях. У высших животных гормоны синтезируются в специализированных секреторных клетках. Освобождаясь в кровяное русло, они поступают в ткани. Поскольку молекулы белковых гормонов имеют относительно крупные размеры, то они не проникают в клетки, поэтому их эффекты обеспечиваются белками-рецепторами, локализованными в мембранах клеток-мишеней, и внутриклеточными уровнями циклического АМФ (цАМФ). Напротив, стероидные гормоны являются малыми молекулами, вследствие чего легко проникают в клетки через мембраны. Оказавшись внутри клеток, они связываются со специфическими рецепторными белками, которые имеются в цитоплазме только клеток-мишеней. Как считают, комплексы гормон - белковый рецептор, концентрируясь в ядрах клеток-мишеней, активируют транскрипцию специфических генов через взаимодействие с определенны-
ми негистоновыми белками, которые связываются с промоторными районами специфических генов. Следовательно, связывание ком- плекса гормон + белок (белковый рецептор) с негистоновыми белками освобождает промоторные районы для движения РНК-полимеразы. Обобщая данные о генетическом контроле эмбрионального периода в онтогенезе организмов, можно заключить, что его ход контролируется дифференциальным включением и выключением действия генов в разных клетках (тканях) путем их дерепрессии и репрессии.
ПОСТЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
После появления организма на свет начинается его постэмбриональное развитие (постнатальное для человека), которое у разных организмов протекает от нескольких дней до сотен лет в зависимости от их видовой принадлежности. Следовательно, продолжительность жизни - это видовой признак организмов, не зависящий от уровня их организации (см. ниже).
В постэмбриональном онтогенезе различают ювенильный и пубертатный периоды, а также период старости, заканчивающийся смертью.
Ювенильный период. Этот период (от лат. juvenilis - юный) опреде- ляется временем от рождения организма до полового созревания. Он протекает по-разному и зависит от типа онтогенеза организмов. Для этого периода характерно либо прямое, либо непрямое развитие.
В случае организмов, для которых характерно прямое развитие (многие беспозвоночные, рыбы, пресмыкающиеся, птицы, млекопи- тающие, человек), вылупившиеся из яйцевых оболочек или новорожденные сходны со взрослыми формами, отличаясь от последних лишь меньшими размерами, а также недоразвитием отдельных органов и несовершенными пропорциями тела (рис. 30).
Характерной особенностью роста в ювенильный период организмов, подверженных прямому развитию, является то, что проис- ходит увеличение количества и размеров клеток, изменяются пропорции тела. Рост человека в разные периоды его онтогенеза показан на рис. 31. Рост разных органов человека неравномерен. Например, рост головы заканчивается в детстве, ноги достигают пропорциональной величины примерно к 10 годам. Наружные половые органы очень быстро растут в возрасте 12-14 лет. Различают определенный и неопределенный рост. Определенный рост характерен для организмов, которые к определенному возрасту прекращают свой рост,
например насекомые, млекопитающие, человек. Неопределенный рост характерен для организмов, которые растут всю жизнь, напри- мер моллюски, рыбы, земноводные, рептилии, многие виды растений.
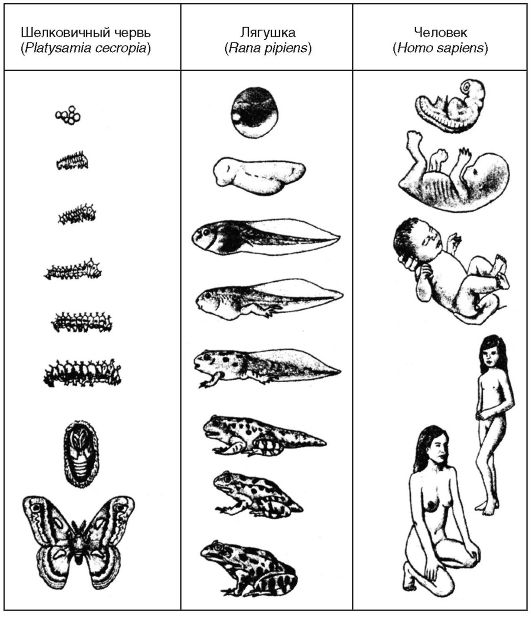
Рис. 30. Прямое и непрямое развитие организмов разных видов
В случае непрямого развития организмы претерпевают превращения, называемые метаморфозами (от лат. metamorphosis - превращение).
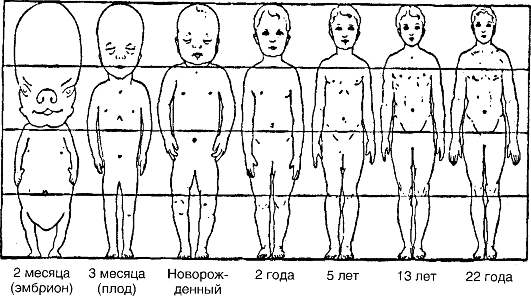
Рис. 31. Рост и развитие в разные периоды онтогенеза человека
Они представляют собой видоизменения организмов в процессе развития. Метаморфозы широко встречаются у кишечнополостных (гидры, медузы, коралловые полипы), плоских червей (фасциолы), круглых червей (аскариды), моллюсков (устрицы, мидии, осьминоги), членистоногих (раки, речные крабы, омары, креветки, скорпионы, пауки, клещи, насекомые) и даже у некоторых хордовых (оболочечники и земноводные). При этом различают полные и неполные метаморфозы. Наиболее выразительные формы метаморфоз наблюдаются у насекомых, которые подвергаются как неполным, так и полным метаморфозам.
Неполное превращение - это такое развитие, при котором из яйцевых оболочек выходит организм, строение которого сходно со строением взрослого организма, но размеры намного меньше. Такой организм называют личинкой. В процессе роста и развития размеры личинок увеличиваются, но имеющийся хитинизированный покров мешает дальнейшему увеличению размеров тела, что приводит к линьке, т. е. сбрасыванию хитинизированного покрова, под которым находится мягкая кутикула. Последняя расправляется, и это сопровождается увеличением размеров животного. После нескольких линек животное достигает зрелости. Неполное превращение характерно, например, в случае развития клопов.
Полное превращение - это такое развитие, при котором из яйцевых оболочек освобождается личинка, существенно отличающаяся
по строению от взрослых особей. Например, у бабочек и многих насекомых личинками являются гусеницы. Гусеницы подвержены линьке, причем могут линять по нескольку раз, превращаясь затем в куколки. Из последних развиваются взрослые формы (имаго), которые не отличаются от исходных.
У позвоночных метаморфозы встречаются среди земноводных и костных рыб. Для личиночной стадии характерно наличие про- визорных органов, которые либо повторяют признаки предков, либо имеют явно приспособительное значение. Например, для головастика, являющегося личиночной формой лягушки и повторяющего признаки исходной формы, характерны рыбообразная форма, наличие жаберного дыхания и одного круга кровообращения. Приспособительными признаками головастиков являются их присоски и длинный кишечник. Для личиночных форм характерно также и то, что по сравнению со взрослыми формами они оказываются приспособленными к жизни в совершенно иных условиях, занимая другую экологическую нишу и другое место в цепи питания. Например, личинки лягушек имеют жаберное дыхание, тогда как взрослые формы - легочное. В отличие от взрослых форм, которые являются плотоядными существами, личинки лягушек питаются растительной пищей.
Последовательность событий в развитии организмов часто называют жизненными циклами, которые могут быть простыми и сложными. Наиболее простые циклы развития характерны, например, для млекопитающих, когда из оплодотворенной яйцеклетки развивается организм, который снова продуцирует яйцеклетки, и т. д. Сложными биологическими циклами являются циклы у животных, для которых характерно развитие с метаморфозами. Знания о биологических циклах имеют практическое значение, особенно в случаях, когда организмы являются возбудителями или переносчиками воз- будителей болезни животных и растений.
Развитие и дифференциация, связанные с метаморфозами, являются результатом естественного отбора, благодаря которому многие личиночные формы, например гусеницы насекомых и головастики лягушек, адаптированы к среде лучше, чем взрослые половозрелые формы.
Пубертатный период. Этот период называют еще зрелым, и он связан с половой зрелостью организмов. Развитие организмов в этот период достигает максимума.
На рост и развитие в постэмбриональный период большое влияние оказывают факторы среды. Для растений решающими факторами являются свет, влажность, температура, количество и качество питательных веществ в почве. Для животных первостепенное значение имеет полноценное кормление (наличие в корме белков, углеводов, липидов, минеральных солей, витаминов, микроэлементов). Важны также кислород, температура, свет (синтез витамина D).
Рост и индивидуальное развитие животных организмов подвержены нейрогуморальной регуляции со стороны гуморальных и нервных механизмов регуляции. У растений обнаружены гормоноподобные активные вещества, получившие название фитогормонов. Последние влияют на жизненно важные отправления растительных организмов.
В клетках животных в процессе жизнедеятельности синтезируются химически активные вещества, влияющие на процессы жизнедеятельности. Нервные клетки беспозвоночных и позвоночных выра- батывают вещества, получившие название нейросекретов. Железы эндокринной, или внутренней, секреции также выделяют вещества, которые получили название гормонов. Эндокринные железы, в частности те, что имеют отношение к росту и развитию, регулируются нейросекретами. У членистоногих регуляция роста и развития очень хорошо показана на примере влияния гормонов на линьку. Синтез личиночного секрета клетками регулируются гормонами, накапливающимися в мозге. В особой железе у ракообразных вырабатывается гормон, тормозящий линьку. Уровни этих гормонов определяют периодичность линек. У насекомых установлены гормональная регуляция созревания яиц, протекающие диапаузы.
У позвоночных железами внутренней секреции являются гипофиз, эпифиз, щитовидная, паращитовидная, поджелудочная, над- почечники и половые железы, которые тесно связаны одна с другой. Гипофиз у позвоночных вырабатывает гонадотропный гормон, стимулирующий деятельность половых желез. У человека гормон гипофиза влияет на рост. При недостатке развивается карликовость, при избытке - гигантизм. Эпифиз продуцирует гормон, который влияет на сезонные колебания в половой активности животных. Гормон щитовидной железы влияет на метаморфоз насекомых и земноводных. У млекопитающих недоразвитие щитовидной железы ведет к задержке роста, недоразвитию половых органов. У человека из-за дефекта щитовидной железы задерживаются окостенение, рост
(карликовость), не наступает полового созревания, останавливается психическое развитие (кретинизм). Надпочечники продуцируют гормоны, оказывающие влияние на метаболизм, рост и дифференци- ровку клеток. Половые железы продуцируют половые гормоны, которые определяют вторичные половые признаки. Удаление половых желез ведет к необратимым изменениям ряда признаков. Например, у кастрированных петухов прекращается рост гребня, теряется половой инстинкт. Кастрированный мужчина приобретает внешнее сходство с женщиной (не растут борода и волосы на коже, отлагается жир на груди и в области таза, сохраняется тембр голоса и т. д.).
Фитогормонами растений являются ауксины, цитокинины и гиббереллины. Они регулирую рост и деление клеток, образование новых корней, развитие цветков и другие свойства растений.
На всех периодах онтогенеза организмы способны к восстановлению утраченных или поврежденных частей тела. Это свойство организмов носит название регенерации, которая бывает физиологической и репаративной.
Физиологическая регенерация - это замена утерянных частей тела в процессе жизнедеятельности организма. Регенерации этого типа очень распространены в животном мире. Например, у членистоногих она представлена линькой, которая связана с ростом. У рептилий регенерация выражается в замещении хвоста и чешуи, у птиц - перьев, когтей и шпор. У млекопитающих примером физиологической регенерации может быть ежегодное сбрасывание оленями рогов.
Репаративная регенерация - это восстановление части тела организма, отторгнутой насильственным путем. Регенерация этого типа возможна у многих животных, но ее проявления различны. Например, она часта у гидр и связана с размножением последних, поскольку из части регенерирует весь организм. У других организмов регенерации проявляются в виде способности отдельных органов к восста- новлению после утраты ими какой-либо части. У человека достаточно высокой регенеративной способностью обладают эпителиальная, соединительная, мышечная и костная ткани.
Растения многих видов также способны к регенерации.
Данные о регенерации имеют большое значение не только в биологии. Их широко используют в сельском хозяйстве, в медицине, в частности в хирургии.
Старость как этап онтогенеза. Старость является предпоследним этапом онтогенеза животных, причем ее длительность определяется
общей продолжительностью жизни, которая служит видовым признаком и у разных животных неодинаковая. Наиболее точно старость изучена у человека.
Известны самые различные определения старости человека. В частности, одно из наиболее популярных определений заключается в том, что старость есть накопление последовательных изменений, сопровождающих повышение возраста организма и увеличивающих вероятность его болезней или смерти. Науку о старости человека называют геронтологией (от греч. geron - старик, старец, logos - наука). Ее задачей является изучение закономерностей возрастного перехода между зрелостью и смертью.
Научные исследования в геронтологии распространяются на разные области, начиная с исследований изменений активности клеточных ферментов и заканчивая выяснением влияния психологических и социологических смягчений в стрессах среды на поведение старых людей.
В случае человека различают физиологическую старость; старость, связанную с календарным возрастом; и преждевременное старение, обусловленное социальными факторами и болезнями. В соответствии с рекомендациями ВОЗ пожилым возрастом человека следует считать возраст порядка 60-75 лет, а старым - 75 лет и более.
Старость человека характеризуется рядом внешних и внутренних признаков.
Среди
внешних признаков старости наиболее заметными являются снижение
плавности движений, изменение осанки, снижение эластичности кожи, массы
тела, упругости и эластичности мышц, появление на лице и других
участках тела морщин, выпадение зубов. Так, например, по обобщенным
данным, человек в возрасте 30 лет теряет 2 зуба (в результате
выпадения), в 40 лет - 4 зуба, в 50 лет - 8 зубов, а в 60 лет - уже 11
зубов. Заметным изменениям подвергается первая сигнальная система
(притупляется острота органов чувств). Например, максимальное
расстояние, при котором здоровые люди различают те или иные одинаковые
звуки, в 20-30 лет составляет
Среди внутренних признаков в первую очередь следует назвать такие признаки, как обратное развитие (инволюция) органов. Отмечается уменьшение размеров печени и почек, а также количе-
ства нефронов в почках (к 80 годам почти наполовину), что снижает функциональные возможности почек и отражается на водноэлектролитном обмене. Снижается эластичность кровеносных сосудов, уменьшается перфузия кровью тканей и органов, повышается периферическая сопротивляемость сосудов. В костях накапливаются неорганические соли, изменяются (обызвествляются) хрящи, снижается способность органов к регенерации. Происходят существенные изменения в клетках, замедляется деление и восстанов- ление их функционального тонуса, уменьшается содержание воды, снижается активность клеточных ферментов, нарушается координированность между ассимиляцией и диссимиляцией. В головном мозге нарушается синтез белков, в результате чего образуются аномальные белки. Повышается вязкость клеточных мембран, нарушаются синтез и утилизация половых гормонов, происходят изменения в структуре нейронов. Наступают структурные изменения белков соединительной ткани и эластичности этой ткани. Ослабляются иммунологические реакции, увеличивается возможность аутоиммунных реакций. Снижаются функции эндокринных систем, в частности половых желез. Поведение других признаков в старости показано на рис. 32.
Рис. 32. Изменение некоторых признаков человека с возрастом: 1 - скорость проведения нервных импульсов; 2 - уровень основного обмена; 3 - сердечный индекс; 4 - уровень почечной фильтрации по инсулину; 5 - дыхательные объемы легких; 6 - уровень плазмотока в почках
Стремления понять природу старения организма возникли давно. В Древней Греции Гиппократ считал, что старение связано с неумеренностью в пище, недостаточным пребыванием на свежем воздухе. По мнению Аристотеля, старение обусловлено расходом тепловой энергии организмом. Значение пищи как фактора старения отмечал также Гален. Но долгое время для объективного понимания этой проблемы не хватало научных данных. Лишь в XIX в. в изучении старения наметился некоторый прогресс, стали возникать теории старения.
Одной из первых наиболее известных теорий старения организма человека является теория немецкого врача X. Гуфеланда (1762-1836), который придавал в долголетии значение трудовой деятельности. До нас дошло его высказывание о том, что ни один лентяй не дожил до преклонного возраста. Еще более известна эндокринная теория старения, которая берет начало из опытов, выполненных еше в середине позапрошлого века Бертольдом (1849), который показал, что пересадка семенников от одних животных к другим сопровождается развитием вторичных половых признаков. Позднее французский физиолог Ш. Броун-Секар (1818-1894) на основе результатов впрыскиваний себе экстрактов из семенников утверждал, что эти инъекции производят благотворное и омолаживающее действие. В начале XX в. уже сложилось убеждение в том, что наступление старости связано с угасанием деятельности желез внутренней секреции, в частности половых желез. В 20-30-е гг. XX в. на основе этого убеждения в разных странах было проделано много операций по омолаживанию пожилых или старых людей. Например, Г. Штейнах в Австрии перевязывал у мужчин семенные канатики, что вело к прекращению внешней секреции половых желез и якобы к некоторому омоложению. С.А. Воронов во Франции пересаживал семенники от молодых животных к старым и от обезьян к мужчинам, а Тушнов в СССР омолаживал петухов, вводя им гистолизаты половых желез. Все эти операции приводили к некоторым эффектам, но лишь временным. После названных воздействий процессы старения продолжались, причем еще интенсивнее.
В начале XX в. возникла микробиологическая теория старения, творцом которой был И.И. Мечников, различавший физиологиче- скую старость и патологическую. Он считал, что старость человека является патологической, т.е. преждевременной. Основу представлений И.И. Мечникова составляло учение об ортобиозе (orthos -
правильный, bios - жизнь), в соответствии с которым основной причиной старения является повреждение нервных клеток продуктами интоксикации, образующимися в результате гниения в толстом кишечнике. Развивая учение о нормальном образе жизни (соблюдение правил гигиены, регулярный труд, воздержание от вредных привычек), И.И. Мечников предлагал также способ подавления гнилостных бактерий кишечника путем употребления кисломолочных продуктов.
В 30-е гг. XX в. широкое распространение получила теория о роли центральной нервной системы (ЦНС) в старении. Творцом этой теории является И.П. Павлов, который установил интегрирующую роль ЦНС в нормальном функционировании организмов. Последователи И.П. Павлова в экспериментах на животных показали, что преждевременное старение вызывается нервными потрясениями и продолжительным нервным перенапряжением.
Заслуживает упоминания теория возрастных изменений соединительной ткани, сформулированная в 30-е гг. XX в. А.А. Богомольцем (1881-1946). Он считал, что физиологическую активность организма обеспечивает соединительная ткань (костная ткань, хрящи, сухожилия, связки и волокнистая соединительная ткань) и что изменения коллоидного состояния клеток, потеря их тургора и т. д. определяют возрастные изменения организмов. Современные данные указывают на значение накопления кальция в соединительных тканях, так как он способствует потере ее упругости, а также уплотнению сосудов.
Для современных подходов к познанию сущности и механизмов старения характерно широкое использование данных физико- химической биологии, и в частности достижений молекулярной генетики. Наиболее распространенные современные представления о механизме старения сводятся к тому, что в процессе жизни в клетках организма накапливаются соматические мутации, в результате которых происходит синтез дефектных белков или нерепарируемые сшивки ДНК с белком. Поскольку дефектные белки играют дозинте- грирующую роль в клеточном метаболизме, то это ведет к старению. В случае культивируемых фибробластов показано, что связанные со старыми клетками белки и мРНК подавляют синтез ДНК в молодых фибробластах.
Известна также гипотеза, в соответствии с которой старение считается результатом изменения митохондриальных метаболитов с последующим нарушением функций ферментов.
У человека показано существование генов, определяющих сроки развития наследственных дегенеративных процессов, связанных со старением. Ряд исследователей считают, что причиной старения являются изменения в системе иммунологической защиты организма, в частности аутоиммунные реакции на структуры организма, имеющие жизненное значение. Наконец, в объяснениях механизмов старения специалисты большое внимание уделяют повреждениям белков, связанным с образованием свободных радикалов. Наконец, иногда придают значение освобождающимся после распада лизосом гидролазам, которые разрушают клетки.
Однако исчерпывающей теории старения все же не создано, поскольку ясно, что ни одна из этих теорий самостоятельно объяс- нить механизмы старения не может.
Смерть. Смерть является завершающим этапом онтогенеза. Вопрос о смерти в биологии занимает особое место, ибо чувство смерти «...совершенно инстинктивно присуще человеческой природе и всегда составляло одну из величайших забот человека» (И.И. Мечников, 1913). Более того, вопрос о смерти стоял и стоит в центре внимания всех философских и религиозных учений, хотя философия смерти в разные исторические времена представлялась неодинаково. В античном мире Сократ и Платон доказывали бессмертие души, тогда как Аристотель отрицал платоновскую идею бессмертия души и верил в бессмертность человеческого духа, продолжающего жить после смерти человека. Цицерон и Сенека также признавали будущую жизнь, но Марк Аврелий считал смерть естественным явлением, которое следует принимать безропотно. В XVIII в. И. Кант и И. Фихте (1762-1814) тоже верили в будущую жизнь, а Г. Гегель придерживался убеждений, по которым душа поглощается «абсолютным существом», хотя природа этого «существа» не раскрывалась.
В соответствии со всеми известными религиозными учениями земная жизнь человека продолжается и после его смерти, и человек должен неустанно готовиться к этой будущей смерти. Однако естество- испытатели и философы, не признающие бессмертия, считали и считают, что смерть представляет собой, как неоднократно подчеркивал И.И. Мечников, естественный исход жизни организма. Более образное определение смерти заключается в том, что она «...есть явная победа бессмыслия над смыслом, хаоса над космосом» (В. Соловьев, 1894).
Научные данные свидетельствуют о том, что у одноклеточных организмов (растений и животных) следует отличать смерть от прекраще-
ния их существования. Смертью является их гибель, тогда как прекращение существования связано с их делением. Следовательно, недолговечность одноклеточных организмов компенсируется их раз- множением. У многоклеточных растений и животных смерть является в полном смысле слова завершением жизни организма.
У человека вероятность смерти повышается в пубертатный период. В частности, в развитых странах вероятность смерти возрастает почти экспоненциально после 28 лет.
Различают клиническую и биологическую смерть человека. Клиническая смерть выражается в потере сознания, прекращении сердцебиения и дыхания, однако большинство клеток и органов все же остаются живыми. Происходит самообновление клеток, продолжается перистальтика кишечника. Клиническая смерть не «доходит» до биологической смерти, ибо она обратима, так как в состоянии клинической смерти можно «возвращать» к жизни. Например, собак «возвращают» к жизни через 5-6 мин, человека - через 6-7 мин от начала клинической смерти. Биологическая смерть характеризуется тем, что она необратима. Остановка сердцебиения и дыхания сопровождается прекращением процессов самообновления, гибелью и разложением клеток. Однако гибель клеток начинается не во всех органах одновременно. Вначале гибнет кора головного мозга, затем гибнут эпителиальные клетки кишечника, легких, печени, клетки мышц, сердца.
На представлениях о клинической смерти основаны мероприятия по реанимации (оживлении) организмов, что имеет исключительно важное значение в современной медицине.
ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
Сравнение данных о продолжительности жизни представителей флоры и фауны показывает, что среди растений и животных различ- ные организмы живут разное время. Например, травянистые растения (дикие и культурные) живут в течение одного сезона. Напротив, их древесные растения характеризуются большей продолжительностью жизни. Например, вишня живет 100 лет, ель - 1000 лет, дуб -
2000 лет, сосна - до 2000-4000 лет.
Рыбы многих видов живут 55-80 лет, лягушки - 16 лет, крокодилы - 50-60 лет, птицы некоторых видов - до 100 лет. Продолжительность жизни млекопитающих является меньшей.
Например, мелкий рогатый скот живет 20-25 лет, крупный рогатый скот - 30 лет и более, лошади - до 30 лет, собаки - 20 лет и более, волки - 15 лет, медведи - 50 лет, слоны - 100 лет. Среди млеко- питающих долгожителем является человек. Многие люди доживали до 115-120 лет и более.
В соответствии с существующими представлениями продолжительность жизни является видовым количественным признаком, подверженным контролю со стороны генотипа. Считают, что гене- тически детерминированная продолжительность жизни коррелирует с периодами онтогенеза (табл. 5).
Таблица 5. Зависимость уровня фибробластов при культивировании от возраста донора
Возраст донора фибробластов, лет | Количество удвоения |
Эмбрион | 50?10 |
0-20 | 30?10 |
20 | 20?10 |
Предполагают, что видовая продолжительность жизни является эволюционным приобретением вида. Что касается долгожительства отдельных индивидуумов, то его объяснение возможно допущением наличия в их генотипах сочетаний определенных генов, либо небольшого количества или полного отсутствия мутаций в их клетках.
А.А. Богомолец и И.И. Шмальгаузен подсчитали, что естественная продолжительность жизни человека должна составлять 120- 150 лет. Однако до этого возраста доживают лишь единицы. Поэтому фактическая продолжительность жизни не совпадает с естественной продолжительностью.
На среднюю продолжительность жизни оказывает влияние ряд факторов (снижение детской смертности, эффективность борьбы с инфекциями, успехи хирургии, улучшение питания и общих условий жизни). Главнейшие причины снижения продолжительности жизни заключаются в смертности от голода, болезней, недостаточной медицинской помощи. В России в последние годы отмечается отрицательная численность населения во всех русских городах и сельской местности за счет снижения рождаемости и высокой смертности.
В медицинском плане продолжительность жизни - это показатель здоровья нации. СССР по числу старых людей занимал первое место в мире. Например, на 1 млн жителей приходится 104 человека в возрасте свыше 90 лет, тогда как в Англии - 6, Франции - 7
и
США - 15 человек. В настоящее время отмечается изменение границ
трудоспособного населения по сравнению, например, с 30-ми гг. XX в.
Отличается также разрыв между пенсионным возрастом и активно- стью
людей. В
Гериатрия - наука, задачей которой является разработка способов нормализации изменяющихся функций стареющего организма. Однако современная медицина еще не располагает методами и средствами воздействия на нормальные физиологические процессы. Поэтому роль гериатрии ограничивается лечением возникающих в пожилом и старческом возрасте заболеваний и исключением (при наличии возможностей) факторов риска, вызывающих прежде- временное старение.
ВОПРОСЫ ДЛЯ ОБСУЖДЕНИЯ
1. Что вы понимаете под половым размножением организмов и какова его биологическая роль?
2. Опишите бесполое размножение и назовите его формы.
3. Опишите особенности полового процесса у одноклеточных и многоклеточных организмов.
4. Что такое гаметогенез?
5. Какова функция гамет каждого типа?
6. Какие вы знаете стадии в развитии гамет?
7. В чем заключаются сходства и различия между сперматогенезом и овогенезом?
8. Что такое мейоз и каково его биологическое значение?
9. Опишите фазы мейоза.
10. Влияет ли кроссинговер на результаты мейоза и как?
11. Опишите сущность оплодотворения.
12. В чем заключается разница между зигогенезом и партеногенезом?
13. Какова биологическая роль чередования гаплоидии и диплоидии?
14. Что представляет собой половой диморфизм? Что вы понимаете под гермафродитизмом? Наблюдаются ли случаи гермафродитизма у человека и как часто?
15. Как вы представляете эволюцию способов размножения?
16. Что вы понимаете под ростом и развитием организмов? Какова связь между ростом и дифференциацией клеток?
17. В чем заключаются молекулярные основы дифференцировки клеток? Что вы знаете о стволовых клетках?
18. Сформулируйте понятие об онтогенезе и назовите периоды онтогенеза.
19. Каковы различия между прямым и непрямым развитием?
20. В чем заключается влияние оплодотворения на яйцеклетки?
21. На каком этапе реализации генетической информации осуществляется контроль действия генов?
22. Каким образом оплодотворенная яйцеклетка развивается в многоклеточную структуру?
23. Каким образом развивающиеся клетки и ткани становятся отличными одни от других в процессе развития?
24. Могут ли восстанавливаться утерянные или поврежденные части тела?
25. Какова связь в онтогенезе между старостью и продолжительностью жизни?
26. Сформулируйте и определите разницу между естественной (вероятной) и фактической продолжительностью жизни.
27. Какие вам известны теории старения организмов?
28. Можно ли считать продолжительность жизни человека видовым признаком?
29. Какие факторы оказывают влияние на продолжительность жизни?
30. На чем основано клонирование организмов? Приведите примеры клонирования организмов.
31. В клетку проникает сперматозоид или его содержимое?
32. О чем свидетельствуют данные по клонированию?
33. Преформизм и эпигенез в истории биологии. Имеют ли они научное значение?
34. Каковы различия между клинической и биологической смертью?