Биология: учебник / Пехов А.П., -, 2010. - 664 с.
|
|
|
|
ГЛАВА XI ПРОИСХОЖДЕНИЕ ЧЕЛОВЕКА
Эволюция человека, или антропогенез (от греч. anthropos - человек, genesis - развитие), - это исторический процесс эволюционного становления человека. Науку, изучающую происхождение человека, называют антропологией, возникновение которой датируют рубежом XVIII-XIX вв.
Эволюция человека качественно отличается от эволюции организмов других видов, ибо в человеке действовали не только биологические, но и социальные факторы. Сложность проблемы антропогенеза углубляется тем, что человек сам по себе двулик. Одним лицом он обращен к животному миру, из которого вышел и с которым остается связанным анатомически и физиологически, а вторым - в мир создаваемых коллективным трудом научных и технических достижений, культуры и т.д. Человек, с одной стороны, является биологическим существом, с другой - социальным. Сущность человека не есть абстракт, присущий отдельному индивиду. В своей действительности она - совокупность всех общественных отношений.
Вокруг проблем происхождения человека всегда шла и идет острейшая идеологическая борьба между идеализмом и материализмом. Идеализм приписывал человеку сверхъестественное происхождение. Отрывая сознание от тела и признавая его первичным, независимым от тела, идеалисты признают независимость в создании тела и души, идеалисты признают отделяемость души от тела после смерти. Идеалистические концепции сливаются с религиозными пред- ставлениями, в рамках которых по библейской истории утверждается внезапное сотворение человека на шестой день творения, состоявшегося несколько тысяч лет назад. Напротив, материализм утверждает, что человек вышел из природы, оставаясь тесно связанным с ней, со всем живым миром. Диалектический материализм рассматривает человека в качестве единого целого. Поэтому в рамках материалистических представлений человеческое сознание не представляет собой какой-либо самостоятельной и независимой от тела сущности.
Как отмечали многие, сознание представляет собой функцию мозга человека.
Будучи связанным узами кровного родства с животным миром, человек имеет свою биологическую предысторию. Однако человек имеет качественные отличия от животных. Эти отличия возникли под влиянием социальных факторов в процессе исторического развития человеческого общества.
Наследственная конституция человека сложилась в результате постепенного и длительного процесса эволюции. У непосредственных предшественников человека в процессе эволюции на базе генетической программы непрерывно возникали противоречия между их морфо-физиологической организацией и формирующимися элементарными способами «инстинктивно-трудовой» деятельности. Разрешение этого противоречия путем естественного отбора вначале привело к изменениям передних конечностей, затем к развитию коры больших полушарий головного мозга и, наконец, к появлению сознания. Можно сказать, что это был первый, но решающий акт в завершении специализации генов на гены структуры и гены-регуляторы, причем сознание обеспечило не только становление, но и дальней- шее развитие человека.
В последующем темпы биологического развития человека стали снижаться. Поскольку появление сознания обеспечивало новые формы и возможности приспособления к среде, это вело к уклонениям от действия естественного отбора, в результате чего биологическое развитие уступило место социальному развитию и совершенствованию. Как заметил в свое время К.А. Тимирязев (1843-1920), биологическая эволюция человека осталась за порогом его истории. Таким образом, современный человек является продуктом его генов и среды.
В научном плане учение об антропогенезе тесно связано с учением о расогенезе, где также идет острейшая идеологическая борьба. Извращение научных представлений о происхождении рас ведет к расизму, в основе которого лежит реакционная идея о высших и низших расах.
ВЗГЛЯДЫ НА АНТРОПОГЕНЕЗ В ПРОШЛОМ
Удивительно, но история взглядов на антропогенез гораздо древнее самой антропологии, ибо еще ученые античного мира обсуждали вопросы о месте человека в природе и о появлении человека.
Например, Анаксимандр (610-546 гг. до н.э.) и Аристотель (384-322 гг. до н.э.), определяя место человека в природе, признавали его потомками животных. Разделяя животных на «кровяных» и бескровных, Аристотель отнес человека к группе «кровяных» животных, а между человеком и животными в «кровяной» группе поместил обезьян. То, что человек близок к животным, признавал Гален (130-200 гг. н.э.), который формулировал заключения об анатомии человека на основе результатов вскрытия низших обезьян.
К. Линней (1707-1778) по сравнению со своими предшественниками пошел намного дальше, выделив отряд приматов, отнеся к нему полуобезьян, обезьян и род людей с одним видом - Человек разумный и подчеркивая сходство человека и обезьян. Однако Линнею не удалось правильно очертить границы этого отряда, ибо он относил к нему одновременно то ленивца, то летучих мышей. Не все современники К. Линнея признавали его систему, в частности принадлежность человека к отряду приматов. Были предложены также варианты системы, в которых ранг человека был значительно завышен, поскольку за человеком признавалось отдельное царство природы. Этим, по существу, отрывали человека от животных.
Идею родства человека и обезьян поддерживали в XVIII в. И. Кант (1724-1804), Д. Дидро (1713-1384), Ж. Ламетри (1709-1751), А.А. Каверзнев (род. 1748), А.Н. Радищев (1749-1802), Ж.-Б. Ламарк (1744-1829). Более того, Ж. Ламетри даже считал, что таксономическое объединение человека и животных не унижает человека, что способность ощущения и мышления есть у всех животных и зависит от степени их организации.
Вопреки правильному решению проблемы о родстве человека с животными, вопрос о том, как произошел человек, в трудах древних философов остался открытым. Считают, что первая гипотеза антропогенеза была сформулирована, вероятно, Ламарком. Полагая, что человек имел обезьяноподобных предков, Ламарк впервые назвал очередность эволюционных достижений в превращении обезьяноподобного предка в человека, причем важнейшее значение он придал переходу древесных четвероногих к двуногой локомоции и к жизни на земле. Ламарк описал изменения скелета и мускулатуры предков человека в связи с переходом к прямохождению. Но, переоценив роль среды, он, как и в случае других организмов, все же неправильно представлял движущие силы эволюции человека.
Н.Г. Чернышевский (1828-1889) считал, что антропогенез является одним из проявлений исторического развития природы. А. Уоллес (1823-1913) высказал предположение, что в эволюции человека большое значение имели формы, передвигавшиеся на двух конечностях, и что вслед за прямохождением происходило увеличение мозга. Он предположил, что филогенез человека очень длителен во времени. Несомненно, что эти и другие сходные высказывания были шагом вперед в понимании вопроса о появлении человека, однако они не были исчерпывающими и не привели к образованию научной теории происхождения человека.
КОНЦЕПЦИЯ ЖИВОТНОГО ПРОИСХОЖДЕНИЯ ЧЕЛОВЕКА
В основе современных представлений о происхождении человека лежит концепция, в соответствии с которой человек вышел из мира животных, причем первые научные доказательства в пользу этой концепции были представлены Ч. Дарвиным в его труде «Происхождение человека и половой отбор» (1871). В последующем по мере развития анатомии и эмбриологии эти доказательства пополнились новыми данными, которые указывали на анатоми- ческое сходство эмбрионального развития человека и животных. В настоящее время в пользу концепции животного происхождения человека служит ряд доводов, наиболее важными из которых являются следующие:
1. Для человека характерны все черты, присущие типу Хордовые, подтипу Позвоночные, а именно:
а) билатеральная (двусторонняя) симметрия в строении тела;
б) наличие в зародышевом развитии хорды и жаберных щелей в полости глотки;
в) расположение нервной системы в форме дорсальной трубки;
г) наличие пяти отделов головного мозга, развитого позвоночного столба, двух пар конечностей. Сердце развивается на брюшной стороне тела.
2. Для человека характерны все черты класса Млекопитающие, а именно:
а) живорождение и вскармливание молоком, наличие молочных желез, волосяного покрова;
б) теплокровность и обилие потовых желез для обеспечения терморегуляции;
в) разделение полости тела диафрагмой на брюшной и грудной отделы;
г) наличие четырехкамерного сердца, левой дуги аорты, отсутствие в зрелых эритроцитах ядер;
д) дыхательная система представлена легкими, бронхами, альвеолами;
е) наличие всех костей, характерных для млекопитающих. У человека нет ни одной лишней кости, которая бы отсутствовала у млекопитающих. В скелете имеется 7 шейных позвонков, 2 мыщелка затылочной кости и 3 слуховых косточки, характерные для млекопитающих;
ж) наличие молочных и постоянных зубов трех групп;
з) проявление атавистических признаков, наличие рудиментарных органов (мышцы, приводящие в движение ушную раковину, отросток слепой кишки, третье веко глаза и др.).
3. Для человека характерны все черты подкласса Планцентарные, а именно:
а) наличие плаценты;
б) вынашивание плода внутри тела матери и питание его через плаценту.
4. Для человека характерны все черты отряда Приматы, а именно:
а) наличие одной пары грудных молочных желез;
б) концы пальцев (концевые фаланги) имеют ногти, а ладони покрыты узорами;
в) противопоставление большого пальца передней конечности остальным, что обеспечивает брахиацию (использование конечностей для хватательных движений);
г) наличие менструального периода и беременности длительностью в 9 месяцев;
д) антигены системы АВ0 человека и человекообразных обезьян сходны. Группы крови А и В обнаружены у всех человекообразных обезьян, группа 0 - лишь у шимпанзе. По существу, кровь шимпанзе и гориллы можно переливать человеку;
е) наличие сходства в количестве и строении хромосом. Для человека характерны 23 пары хромосом, для человекообразных обезьян - 24 пары, из которых 13 пар по своему строению одинаковы в обоих случаях;
ж) наличие значительной гомологии ДНК человека с ДНК обезьян. Например, гомология ДНК человека и шимпанзе составляет 91-92%, человека и гиббона - 76%, а человека и макаки-резус - всего лишь 6%;
з) одинаковая чувствительность человека и человекообразных обезьян к возбудителям одних и тех болезней и сходство клинического проявления последних;
и) сходство между генами, контролирующими синтез белков у приматов. Это сходство устанавливают на основе структуры белков, контролируемых этими генами. Например, в клетках всех организмов, дыхание которых связано с использованием кислорода, син- тезируется цитохром С. Этот белок представляет собой сложную молекулу, состоящую примерно из 100 аминокислот. Как следует из табл. 30, степень сходства структуры цитохрома С коррелирует с анатомическим, физиологическим и филогенетическим сходством приматов. Человек отличается от других всего лишь одной аминокислотной заменой.
Таблица 30. Сравнительные данные о структуре цитохрома С у разных организмов
Сравниваемые виды организмов | Кол-во аминокислотных различий |
Человек - обезьяна | 1 |
Человек - лошадь | 12 |
Человек - собака | 10 |
Человек - крупный рогатый скот | 3 |
Человек - куры | 10-15 |
Человек - тунец | 17-21 |
В случае α-гемоглобина человека и гориллы имеется лишь одно различие в последовательности аминокислот, тогда как человека и лошади - 18 различий, человека и карпа - 71 различие. Между человеком и шимпанзе имеется исключительное сходство по строению белков (различие по 44 функциональным белкам не превышает 1%).
Место человека в системе животного мира определяется тем, что он относится к типу Хордовые, подтипу Позвоночные, классу Млекопитающие, подклассу Плацентарные, отряду Приматы, семейству Гоминиды, роду Homo. Как считал Ч. Дарвин, «...животные - наши братья по боли, болезням, смерти, страданию и голоду, наши рабы в самой тяжкой работе, наши товарищи в наших удовольстви-
ях - все они ведут, может быть, свое происхождение от одного общего с нами предка - нас всех можно было бы слить вместе».
Однако человек имеет существенные отличия от животных, на что также обращали внимание еще древние. Например, Анаксагор (500-428 гг. до н.э.) и Сократ (469-399 гг. до н.э.) считали, что специфическим признаком человека является наличие руки, которая выделила его из всего мира. Называя человека «животным общественным», Аристотель ссылался на такие отличия, как двуногое хождение, больший по величине мозг, способность к речи и мышлению. Позднее К. Линней в качестве специфических отличий человека от обезьян называл речевую способность, а также способность накапливать и передавать в поколениях опыт, письменность, печать. По этой причине он и называл человека разумным. А.Н. Радищев обращал внимание на такие отличи- тельные свойства человека, как способность к прямохождению, наличие рук, речи, разума.
Современные
представления относительно отличий человека от животных основывается
прежде всего на данных о различиях в развитии мозга и на способности
человека к абстрактному мышлению. Средняя масса мозга человека
составляет 1350-1500 г, тогда как гориллы и шимпанзе - всего лишь
Среди других
отличительных признаков человека следует назвать особенности челюстей, а
также строение и расположение зубов, которые являются иными по
сравнению с животными. Важнейшими отличиями служат дифференцировка
верхних и нижних конечностей, характерные изгибы позвоночника, широкий
таз. Только человек способен к балансированию на двух конечностях (к
двуногой локомоции). У человека самой мощной является бедренная кость,
выдерживающая нагрузку до
Важнейшей отличительной способностью человека является то, что он обладает сознанием, способностью к абстрактному мышлению, общению с помощью речи (2-й сигнальной системы) и абстрактных
символов (письма), а также к передаче и воспроизведению информации.
Человек способен производить орудия труда для воздействия на природу, он развил технологию, создал города, литературу, музы- ку и т.д. Являясь социальным существом, человек способен думать о прошлом, планировать будущее. Этими свойствами животные не обладают.
ЭТАПЫ АНТРОПОГЕНЕЗА
Филогенез человека представляют исходя из дарвиновского положения, в соответствии с которым предками человека были обезьяноподобные существа, а развитие человека и человекообразных обезьян представляло собой не последовательные ступени, а параллельные ветви эволюции, характеризующиеся глубоким расхождением с эволюционной точки зрения. Ч. Дарвин называл современных человекообразных обезьян двоюродными братьями человека. Однако это сравнение не совсем верно, ибо в наше время гоминид и понгид считают дальними родственниками, у которых вымерли промежуточные ветви, т.е. человек произошел от каких-то животных, одна ветвь которых ведет к современным высшим обезьянам, вторая - к человеку.
Чтобы правильно понять самые ранние этапы эволюции человека, необходимо начать с эволюции отряда приматов, в состав которого входят семейства понгид и гоминид. Находки палеонтологов позволяют проследить развитие животных, приведшее к появлению первых приматов начиная с мезозойской эры, в частности с триаса, когда из тогдашних видов пресмыкающихся возникли

Рис. 113. Тупайя
первые млекопитающие, получившие название Protoinsectivara. Предполагают, что на основе именно этих примитивных млекопитающих в дальнейшем возникли более совершенные млекопитающие - насекомоядные, ставшие предками тупай - современных низших приматов (рис. 113), от которых через разные промежуточные формы одна из эволюционных ветвей повела к приматам, впервые появившимся в меле (рис. 114).
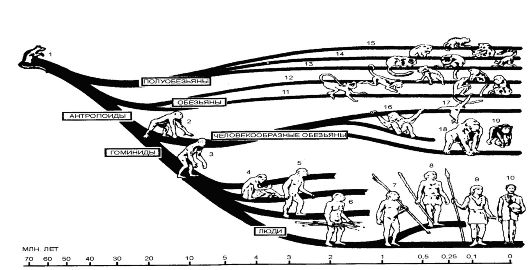
Рис. 114. Генеалогическое древо приматов: 1 - плезиадацис; 2 - дриопитек африканский; 3 - рамапитек; 4 - австралопитек африканский; 5 - стралопитек войсен; 6,7 - Homo erectus; 8 - неандерталец; 9 - Homo sapiens; 10 - современный человек; 11 - узконосые обезьяны Старого Света; 12 - широконосы обезьяны Нового Света; 13 - лемуры; 14 - лоры; 15 - долгопяты; 16 - орангутаны; 17 - гиббоны; 18 - гориллы; 19 - шимпанзе
Отряд приматов классифицируют на подотряды полуобезьян (Prosimii) и обезьян (Antropoidea). Наиболее ранние приматы из подотряда Antropoidea были многочисленны в начале палеоцена (65 млн лет назад). В олигоцене (40 млн лет назад) приматы этого подотряда в результате эволюции дали начало трем большим группам:
1. Надсемейство Ceboidea. Представители этого надсемейства в ходе эволюции в свою очередь дали начало обезьянам Нового Света (широконосым), которые эволюционировали в игрунков и капуцинов. Это надсемейство представляет собой приматов, не имеющих значения в эволюции человека.
2. Надсемейство Cercopithecoidea. Представители этого надсемейства дали начало обезьянам Старого Света (узконосым), которые эволюционировали в макак и павианов. Это надсемейство тоже не имеет значения для эволюции человека.
3. Надсемейство Hominoidea (гоминиды). Представители этого над- семейства дали начало человекообразным обезьянам и человеку. Эволюция этого надсемейства происходила независимо от других приматов. Первые гоминоидные формы появились еще в верхнем плиоцене. В пределах надсемейства Hominoidea выделяют семейства Pliopithecidae, представители которого полностью вымерли, Oreopithecidae, представители которого тоже полностью вымерли, Pongidae (человекообразные обезьяны), представители которого частично вымерли, и Hominidae (люди), представители которого тоже частично вымерли. Понгид делят на подсемейства гиббонов (Hilolatenae) и настоящих человекообразных обезьян (Pongidae), к которым относятся орангутаны, гориллы и шимпанзе.
Начало эволюции семейства понгид (Pongidae) и гоминид (Hominidae) датируется временем, когда одна из ветвей эволюции повела к дриопитековым (древесным человекообразным обезьянам), сходным по костям с гориллами, шимпанзе и человеком. Наибольшая концентрация ископаемых останков дриопитеков обнаружена в Восточной Африке, однако их формы были найдены и в Европе и Азии, на территории ФРГ, Франции, России, КНР, Турции. Возраст дриопитеков составляет около 25 млн лет. Считают, что дриопитековые представляют собой узел родословного древа приматов, от которого берут начало три линии развития - к орангутанам, человекообразным обезьянам и человеку.
У истоков расхождения путей эволюции понгид и гоминид стоит один из дриопитеков, названный в честь индийского бога Рамы
рамапитеком, челюсть которого была найдена Д. Льюисом в
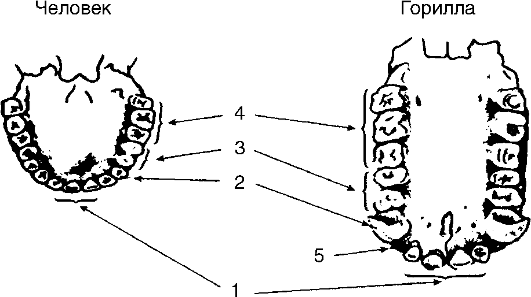
Рис. 115. Зубная аркада человека и человекообразной обезьяны: 1 - резцы; 2 - клыки; 3 - малые коренные; 4 - коренные; 5 - семиальное пространство
После рамапитека современная антропология не располагает какими-либо ископаемыми останками, в результате чего невозможно составить полный эволюционный ряд. Тем не менее условно филогенез человека можно представить в виде ряда последовательных, сменяющихся одна за другой стадий. В частности, различают несколько стадий:
1. Предшественник - предки рода Homo, представляющие собой ископаемых обезьянолюдей (австралопитеков).
2. Архантропы - древнейшие люди, представленные ископаемыми останками.
3. Палеоантропы - древние люди, непосредственные предки Homo sapiens. Представлены ископаемыми останками.
4. Неоантропы - ископаемые формы, связанные с культурами позднего палеолита (Homo sapiens fossilis) и ныне живущие формы (Homo sapiens recens).
Предполагают, что эволюция предков человека была постепенной, с различной частотой в разные периоды.
Стадия предшественника. В
В последующем
ископаемые останки гоминид, которые характеризовались уменьшенными
клыками, увеличенным мозгом и спо- собностью к прямохождению, были
обнаружены в Восточной Африке. Их возраст составляет около 3,7-3,0 млн
лет. Эти останки были отнесены к виду Australopithecus afarensis, которого считают сейчас самым ранним австралопитеком, предшествующим позднейшим плиоплейстоценовым гоминидам. В
в Восточной Африке были обнаружены ископаемые останки «гиперсильного» австралопитека, названного A. boisei, возраст которого 2,0-1,3 млн лет.
На основании изучения тазовых и бедренных костей многие антропологи считают, что австралопитеки были двуногими существа- ми, способными стоять и передвигаться подобно человеку. Поскольку ископаемые останки австралопитеков находили в пещерах, где одновременно обнаруживали и зазубренные (подправленные) камни, то, вероятно, они жили в пещерах, которые использовали в качестве убежищ, и изготовляли самые простейшие орудия (рис. 116).

Рис. 116. Австралопитеки
Австралопитеки были охотниками на мелких животных, но использовали также в пищу зерна, плоды диких деревьев, корни. Вегетарианская пища, обилие растений и их семян, возможно, способствовали эволюции более мелких клыков.
В
его стопы уже не был отведен в сторону, что указывает на полное завершение прямохождения. Кости кисти этого существа сочетали как примитивные (обезьяньи), так и прогрессивные (человеческие) черты. Поскольку вместе с костными останками были найдены грубые галечные орудия и расколотые кости (для добывания мозга), то Лики заключил, что это существо было и творцом этих орудий. Чтобы подчеркнуть способность существа изготовлять простейшие орудия, Лики назвал его Н. habilis (человек умелый).
В
60-70-е гг. XX в. к востоку от озера Рудольф (Кения) Л. Лики нашел
остатки галечной культуры, возраст которых составляет 2,6 млн лет.
Здесь же было обнаружено множество скелетных останков австралопитеков и
более прогрессивных гоминид древностью тоже 2,6 млн лет. В
Итак,
палеонтологические данные в совокупности свидетельствуют о том, что
непосредственными животными предками человека служат австралопитеки -
высокоразвитые двуногие обезьяноподобные существа. Их рост составлял
120-130 см, объем головного мозга - около 500 мл, масса тела - около
передних конечностей для передвижения по деревьям и на земле к свободному передвижению в вертикальном положении при прямом торсе на сводчатой стопе, а также в использовании рук для изготовления простейших орудий, австралопитеки пересекли, по существу, границы звериной сущности. Можно сказать, что они уже не были обезьянами, но еще и не являлись людьми.
Принципиальный вопрос заключается в эволюционных отношениях между плиоплейстоценовыми гоминидами. В современной литературе опубликовано несколько филогений основных таксонов гоминид. Р. Лики считал, что Н. habilis является результатом эволюции Australopithecus afarensis. Ученый с сотрудниками опубликовал результаты изучения 22 особенностей черепа K. NM 17 000 и нижней челюсти черепа KNM-WT 16 005, принадлежащих A. boisei и найденных в плиоценовых отложениях (возраст около 2,5 млн лет) к западу от озера Туркана в Кении, в сравнении с ископаемыми останками A. afarensis, A. africanus, A. robustus и A. boisei. В соответствии с этими результатами предполагается, что A. africanus эволюционировал независимо от A. robustus и A. boisei, которые родственны между собой, и что эволюционная линия, ведущая от австралопитека к роду Homo, начинается с A. africanus.
В
Стадия архантропа (древнейшие люди). Архантропы занимают среднее положение между австралопитеками и современным человеком. С зоологической точки зрения они входят в один комплекс или вид Н. erectus (Человек выпрямленный) (рис. 117).
Открытие архантропов произошло в
также грубые каменные орудия, что свидетельствует о способности яванского питекантропа к трудовой деятельности.

В
Костные
останки ископаемых людей, сходных с питекантропами и синантропами,
были обнаружены на территории ФРГ (гей- дельбергский человек), Алжира
(атлантроп), ВНР, Чехии, Марокко и других стран. Абсолютный возраст
этих людей составляет 400 тыс. лет. Они умели изготовлять орудия из
камня и гальки. В
Древнейшие люди были более крупными, чем австралопитеки, могли ходить более долгое время прямо и без напряжения, полностью использовали руки. Их орудия были совершеннее орудий австра- лопитеков. Превосходя Н. habilis, они охотились на них и убивали их. Обитая в разных географических районах (рис. 118), древнейшие люди жили в пещерах, использовали огонь, но не могли его добы-
Рис. 117. Homo erectus
вать самостоятельно, а лишь поддерживали. Судя по увеличенному размеру мозга, в котором все еще оставались уплотненной теменная доля, узкими височные доли, они должны были обладать речью. Деятельность Н. erectus по добыванию пищи путем охоты изменила их коллективный характер. Архантропы уже планировали и координировали свои действия, в результате чего могли вести охоту на крупных животных, включая мамонтов. Уже на этом этапе возникло разделение труда между теми, кто охотился, и теми, кто оставался в лагере для наблюдения за детьми, стариками и выполнения мелких работ.
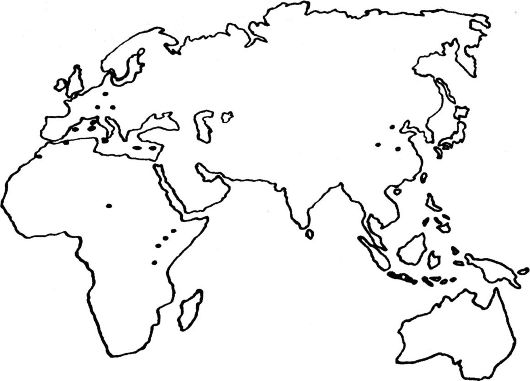
Рис. 118. Распространение Homo erectus. (Места обитания на карте показаны
точками.)
Предполагают, что представители Н. erectus берут начало от одной из ветвей австралопитеков, возможно Н. habilis, которые 2-1,5 млн лет назад начали расселяться по Африке, а затем проникли в Средиземноморье, Южную, Центральную и Юго-Восточную Азию. Имея долгую эволюционную историю, все представители Н. erectus тем не менее характеризуются недостаточным темпом прогрессивных морфологических изменений. Их морфологические преобразования завершились появлением лишь форм с менее угловатым черепом и более тонкими костными структурами, но эти изменения не сопровождались резким совершенствованием методов
изготовления каменных орудий. На этой стадии в эволюции человека в качестве фактора эволюции главенствовал естественный отбор. Предполагают, что популяции Н. erectus были прямыми предками ранних представителей палеоантропов, хотя точное время, способ трансформации в палеоантропов и география процесса остаются дискуссионными.
Стадия палеоантропа (древние люди). В
Череп этих древних людей
был значительно удлинен, с характерным низким сводом и сплошными
выступающими надбровными валиками, хорошо развитыми лобными пазухами.
Объем мозга составлял 1200-1600 мл (как у современного человека), рост -
около
Многие антропологи предполагают, что неандертальцы произошли от каких-то архантропов около 300-200 тыс. лет назад, причем их было две группы - ранние и поздние (классические). Ранние неандертальцы произошли от одной из популяций Н. erectus, жили 300-250 тыс. лет назад и являлись предками как поздних (классических) неандертальцев, так и современных людей. Широкое распространение поздних неандертальцев приходится на период 35-60 тыс. лет назад. Время жизни неандертальцев совпало с периодом очередного наступления ледников. Они строили простые жилища, защищались от холода с помощью одежды и шкур зверей, изготовляли более совершенные кремниевые орудия (остроконечные иглы из каменных пластинок), что позволяло им успешно вести борьбу с мамонтами.
Неандертальцы внезапно исчезли 30-40 тыс. лет назад. Предполагают, что они были либо поглощены, либо частично истреблены людьми современного типа. Более того, поскольку у них были недоразвитыми лобные доли головного мозга, которые обычно ответственны за поведе-
ние, то, вероятно, конфликты между неандертальцами способствовали распаду их групп (общин), вели к самоистреблению.
Стадия неоантропа. Под этим названием понимают как ископаемые формы человека современного физического типа, так и ныне живущих людей. В
Кроманьонцы не отличаются от людей современного физического типа. Если бы кроманьонец, одетый в современную одежду, шел по улице, его трудно было бы сейчас выделить в толпе. В их пещерах были найдены стрелы, гарпуны, иглы, что указывает на усовершенствование методов изготовления орудий. Средний возраст кроманьонцев составлял около 30 лет, хотя отдельные из них доживали до 50 лет и более. Новые данные показывают, что представители Н. sapiens современного физического типа появились 100 000-200 000 лет назад.
Палеонтологические находки указывают на то, что в период появления современных людей доживали свой век и последние неандертальцы. В частности, на территории Палестины найдены скелетные останки гибридов между неандертальцами и людьми современного типа. Как неандертальцы, так и кроманьонцы были искусными охотниками. Подсчитано, что они ответственны за истребление многих млекопитающих. В частности, за истребление мегатерий, мамонтов, саблезубых кошек, бизонов. Более того, примерно около трети мле- копитающих было уничтожено еще 50 тыс. лет назад. Предполагают, что резкое уменьшение диких животных способствовало переходу людей от охотничьего сообщества к сельскохозяйственному.
На стадии неоантропа на развитие человека стали оказывать все большее влияние культура, социальные закономерности.
ПРАРОДИНА ЧЕЛОВЕКА
Вопрос о прародине человека, под которой понимают место, где протекал процесс самой ранней стадии антропогенеза, возник еще в XIX в. Палеонтологи, антропологи и археологи в качестве воз-
можного центра зарождения человечества называли самые различные части земного шара (почти все континенты). Однако Ч. Дарвин в качестве прародины человека назвал Африку, отмечая, что именно в Африке обитают обезьяны, стоящие по морфологии и развитию наиболее близко к современному человеку. Действительно, наибольшая концентрация и разнообразие форм высокоорганизованных обезьян были достигнуты вначале в Африке и Южной Азии. На этих континентах были обнаружены и рамапитеки, имеющие прогрессивные признаки. Однако судьба процесса гоминидизации оказалась разной на этих территориях. В плиоцене в Южной Азии резко изменились климатические условия, стали часты засухи, поднятие Гималаев сопровождалось образованием пустынь. Это привело или к вымиранию, или переселению гоминид, что остановило процесс их эволюции. Напротив, в Африке сложились другие климатические условия. Местом, где продолжилось развитие гоминид, стала приэкваториальная часть Восточной Африки с ее постоянным благоприятным климатом, богатством ландшафта. Таким образом, Восточная Африка оказалась прародиной человека, ибо здесь развитие гоминид достигло такого уровня, когда они отделились от животных.
Как свидетельствуют результаты исследований мтДНК индивидов, принадлежащих к разным европейским, американским, афри- канским, азиатским популяциям, трансформация архаичных форм в Н. sapiens произошла в Африке в период между 100-150 тыс. лет назад. Современные люди являются потомками «митохондриальной Евы», жившей 140 000-280 000 лет назад в Африке. Альтернативный взгляд заключается в том, что представители Н. erectus обитали одновременно около 1 млн лет назад как в Африке, так и в Азии и что трансформация архантропов) в в современных людей шла во многих местах Старого Света. Однако палеонтологические данные не подтверждают мысль о том, что азиатские формы Н. erectus являются предками Н. sapiens. Скорее можно предполагать, что популяции неафриканских форм Н. erectus замещались сапиентами, мигрировавшими из Африки. Люди современного типа появились в Восточной Азии 75 000-150 000 лет назад. В Америку они пришли из Северного Китая через Берингию и Аляску. Современные люди были в Бразилии уже 32 000 лет назад.
ФАКТОРЫ АНТРОПОГЕНЕЗА
Осмысливая происхождение человека, Ч. Дарвин придавал большое значение таким факторам антропогенеза, как изменчивость телесных и психических свойств предков человека, естественный половой отбор.
Современные представления сводятся к пониманию того, что люди являются продуктом действия тех же факторов эволюции, которые создали других животных, однако с учетом специфики человека должны были действовать и специфические факторы антропогенеза. Основы учения о специфических факторах антропогенеза были заложены Ф. Энгельсом в его работе «Роль труда в процессе превращения обезьяны в человека» (1871-1878). Считая, что появление человека не служит лишь ступенью в эволюции природы, ибо с человеком возникает новая сила в виде труда, который, воздействуя на природу, не только изменяет ее, но изменяет и человека, Ф. Энгельс назвал важнейшим специфическим фактором антропогенеза труд. По Ф. Энгельсу, труд отделил человека от животных, а люди благодаря совместной деятельности руки, органов речи и мозга стали обладать способностью выполнять усложняющиеся операции, ставить перед собой каждый раз все более высокие цели и достигать их.
Новая аргументация в объяснении причин, заставивших австралопитеков прибегнуть к труду, подтверждает заключения Ф. Энгельса о роли труда. В частности, сейчас предполагают, что имевшие в плейоцене место бурные тектонические процессы, активный вулканизм, горообразование, землетрясения и другие факторы привели в Восточной Африке к обнажению урановых руд и повышению уровня ионизирующей радиации. Являясь мутагенным фактором, ионизирующая радиация индуцировала мутации австралопите- ков, которые в совокупности сделали предков настолько слабыми, что они оказались неспособными добывать пищу животными методами. Единственным в этих условиях путем выжить был переход части австралопитеков от использования камней и палок к изготовлению более совершенных орудий нападения и защиты. В после- дующем изготовление орудий обеспечило древнейшим людям выход на господствующее положение в природе. Поскольку для общения необходим был язык, то в головном мозге развивались теменные и лобные доли (центры речи), совершенствовались анализаторы.
Однако изготовление эффективных орудий возможно было только на основе навыков, наследуемых от предыдущих поколений, что вело
к повышению роли и значения в первобытных сообществах людей старшего поколения, хранящих опыт изготовления орудий, охоты и т.д. Чем лучше охранялись слабые, тем больше оставалось шансов на выживание. Видимо, в этот период стала прекращаться внутривидовая борьба, падать значение естественного отбора. Напротив, важным фактором антропогенеза становились возникавшие социальные закономерности. Таким образом, создав социальные отношения в группах древнейших и древних людей, естественный отбор уступил место социальным факторам. В борьбе за существование стали побеждать не только те, что были самыми сильными, но и те, которые охраняли слабых (стариков, женщин, детей). В результате этого в борьбе за существование стали лидировать те группы или племена, в которых больше заботились о поддержании стариков, ибо последние сохраняли опыт в изготовлении орудий, поиск мест охоты и съедобных растений, ориентировку на местности и т.д. Следовательно, если у ранних австралопитеков в основном действовал естественный отбор особей, лучше владеющих руками, а также отбор на повышение роли орудийной деятельности, то в возникновении Н. habilis большое значение имел альтруизм, ставший фактором эволюции и дававший многие преимущества в условиях коллективной жизни. Возникновение социальной среды стало необратимым. Популяции, лучше изготовлявшие и использовавшие орудия, вытесняли отстающие популяции в райо- ны, менее благоприятные для жизни, что вело к их уничтожению.
Определенное значение в антропогенезе имела мясная пища, которая стала, по существу, самостоятельным фактором антропоге- неза. Как предполагают, австралопитеки были хищными существами. Н. habilis со своими орудиями оказался более опасным хищником и для самих австралопитеков. Возможно, что уже на этой стадии возник каннибализм. Современные представления о роли этого фактора сводятся к тому, что употребление мяса обеспечивало предков современного человека незаменимыми аминокислотами, которых нет в пище из растений.
РАСЫ И ИХ ПРОИСХОЖДЕНИЕ
Расы - это исторически сложившиеся группировки (группы популяций) людей разной численности, характеризующиеся сходством морфологических и физиологических свойств, а также общностью занимаемых ими территорий. Развиваясь под влиянием исто-
рических факторов и принадлежа к одному виду (Н. sapiens), раса отличается от народа, или этноса, который, обладая определенной территорией расселения, может состоять из нескольких расовых ком- плексов, или, наоборот, к одной и той же расе может принадлежать ряд народов и носителей многих языков.
О существовании рас люди знали еще до нашей эры. Тогда же были предприняты и первые попытки объяснить их происхождение. Например, в мифах древних греков происхождение людей с черной кожей объяснялось неосторожностью Фаэтона (сына бога Гелиоса), который на солнечной колеснице так приблизился к Земле, что обжег стоявших на ней белых людей. Греческие философы в объяснениях причин возникновения рас большое значение придавали климату. В соответствии с библейской историей родоначальниками белой, черной и желтой рас были сыновья Ноя - любимый богом Яфет, Сим и проклятый богом Хам (соответственно).
Стремления
систематизировать представления о физических типах народов, населяющих
земной шар, датируются XVII в., когда, опираясь на различия людей в
строении лица, цвета кожи, волос, глаз, а также особенности языка и
культурных традиций, французский врач Ф. Бернье впервые в
Француз Ж. Блюменбах в
1. Европейская группа, включая лопарей, южных европейцев и северных африканцев.
2. Африканская группа.
3. Азиатская группа, включая жителей индийского субконтинента.
4. Американская группа, включая всех аборигенов.
5. Тихоокеанская группа (меланезийцы, полинезийцы, австралоиды).
На основе понимания расогенеза в качестве процесса, продолжающегося и в настоящее время, Ф. Добржанский (1962) классифицировал человечество на 34 расы (рис. 119), а именно:
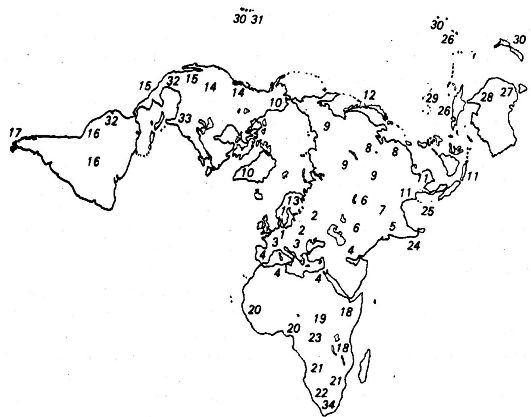
Рис. 119. Географическое распространение рас человека. Обозначение их в тексте цифрами
1. Северо-западная европейская - жители Скандинавии, северной части ФРГ, Северной Франции, Великобритании и Ирландии.
2. Северо-восточная европейская - жители Польши, Европейской части России, Украины, Белоруссии, большинство существующих популяций в Сибири.
3. Альпийская - жители территорий, простирающихся от Центральной Франции, южной части ФРГ, Швейцарии, Северной Италии до побережья Черного моря.
4. Средиземноморская - популяции по обе стороны Средиземноморья, от Танжера до Дарданелл, Аравии, Турции, Ирака.
5. Хинду - жители Индии, Пакистана.
6. Тюркская - жители Туркестана, Западного Китая.
8. Северо-китайская - жители Северного и Центрального Китая, Маньчжурии.
9. Классическая монголоидная - жители Сибири, Монголии, Кореи, Японии.
10. Эскимосская - жители арктической Азии и Америки.
11. Юго-восточная азиатская - жители Южного Китая до Таиланда, Бирмы, Малайи и Индонезии.
12. Айну - популяция аборигенов в Северной Японии.
13. Лопари (саами) - аборигены арктической Скандинавии и Финляндии.
14. Северо-американская индейская - туземные популяции Канады
и США.
15. Центрально-американская индейская - популяции, обитающие на территориях, простирающихся от юго-западных районов Северной Америки через Центральную Америку до Боливии.
16. Южно-американская индейская - популяции Перу, Боливии и Чили, занимающиеся главным образом сельским хозяйством.
17. Фужиан - аборигены юга Южной Америки, не занимающиеся сельским хозяйством.
18. Восточно-африканская - популяции Восточной Африки, Эфиопии, части Судана.
19. Суданская - популяции большей части Судана.
20. Лесная негроидная - популяции, обитающие в лесах Западной Африки и вдоль большей части реки Конго.
21. Банту - аборигены Южной Африки и части Восточной Африки.
22. Бушмены и готтентоты - аборигены, населяющие Южную Африку.
23. Африканские пигмеи - популяции малорослых людей, обитающих в лесах Экваториальной Африки.
24. Дравидианы - аборигены Южной Индии и Цейлона.
25. Негрито - популяция малорослых людей с вьющимися волосами на территории от Филиппин до Андаманса, Малайи и Новой Гвинеи.
26. Меланезийская папуасская - аборигены Новой Гвинеи до Фиджи.
27. Мураджиан - популяция аборигенов Юго-Восточной Австралии.
28. Карпентариан - популяция аборигенов Северной и Центральной Австралии.
29. Микронезийская - популяции островов западной части Тихого океана.
30. Полинезийская - популяции островов центральной и восточной части Тихого океана.
31. Неогавайская - популяция, недавно возникшая на Гавайских островах.
32. Ладино - популяция, недавно возникшая в Цетральной и Южной Америке.
33. Северо-американская цветная - негритянская популяция
в США.
34. Южно-африканская цветная - негритянская популяция Южной Африки.
В практическом плане очень популярна классификация рода человеческого на европеоидов, негроидов, монголоидов и австралоидов.
Европеоиды светлокожи, для них характерны прямые или волнистые светло-русые или темно-русые мягкие или средней жесткости волосы, серые, серо-зеленые и каре-зеленые широко открытые глаза, умеренно развитый подбородок, широкий таз, узкий и сильно выступающий нос, нетолстые губы и довольно обильный волосяной покров тела и лица. Для женщин этой расы характерны полусферические груди и выпуклые ягодицы. Люди, принадлежащие к этой расе, занимают большую часть Европы, а также прилегающие к ней области.
Негроиды темнокожи, для них характерны курчавые или шерстистые темные волосы, толстые губы, очень широкий и плоский нос, очень крупные зубы, карие или черные глаза, длинная голова, редкая растительность на лице и теле, узкий таз, большие ступни. Для женщин характерны груди конической формы и мало выпуклые ягодицы. Люди, принадлежащие к этой расе, занимают практически весь экваториальный пояс от Африки до островов Тихого океана. К этой расе принадлежит население Африки, а также негрито (пиг- меи), океанические негроиды (меланезийцы), южно-африканские бушмены и готтентоты.
Монголоиды смуглокожи, обладают желтой или желто-коричневой кожей. Для них характерны прямые жесткие иссиня-черные воло- сы, плоское скуластое лицо, узкие и слегка раскосые карие глаза со складкой верхнего века (третье веко, или эпикантус) во внутреннем углу глаза, плоский и довольно широкий нос, редкая растительность на лице и теле. Люди, принадлежащие к этой расе, занимают Восточную Сибирь и Монголию, Дальний Восток, Центральную и Юго-Восточную Азию. Смешанную монголоидную расу представ- ляют индонезийцы и американские индейцы.
Часто в виде отдельной расы выделяют австралоидов, которые почти так же темнокожи (их кожа имеет шоколадный цвет), как и негроиды, но для них характерны темные волнистые волосы, крупная голова и массивное лицо с очень широким и плоским носом, выступающим подбородком, значительный рост волос на лице и теле. Австралоиды являются аборигенами Австралии. Однако австралоидов часто считают негроидами. Иногда выделяют американоидов, для которых характерны смуглая кожа, скуластое лицо, достаточно выступающий нос и эпикантус, иссиня-черные волосы. Однако часто американоидов относят к монголоидам.
Данные о распределении групп крови и типов отпечатков пальцев в популяциях, принадлежащих к разным расам, представлены в табл. 31 и 32.
Таблица 31. Распределение групп крови в разных популяциях, %
Популяция | А | АВ | В | 0 | Rh |
Европеоиды | 5-40 | 1-37 | 4-18 | 45-75 | 25-46 |
Негроиды | 8-30 | 1-8 | 10-20 | 52-70 | 4-29 |
Монголоиды | 0-45 | 0-5 | 16-25 | 39-68 | 0-5 |
Американские индейцы | 0-20 | около 0 | 0-4 | 68-100 | около 0 |
Таблица 32. Распределение типов отпечатков пальцев в разных популяциях, %
Популяция | Типы отпечатков | ||
арковидные | петлевидные | закругленные | |
Европеоиды | 0-9 | 63-76 | 20-42 |
Негроиды | 3-12 | 53-73 | 20-40 |
Бушмены | 13-16 | 66-68 | 15-21 |
Китайцы, японцы, и родственные популяции | 1-5 | 43-56 | 44-54 I |
Американские индейцы | 46-61 | 35-57 | |
Австралийские аборигены | 0-1 | 28-46 | 52-73 |
Микронезийцы | 2 | 49 | 49-50 |
Правильное понимание процессов расогенеза зависит от подходов к определению сущности и классификации рас. Длительное время господствовал так называемый типологический подход, в соответствии с которым определение расы проводили на основе стереотипов, якобы отражающих все признаки расы. Поэтому, руководствуясь
характеристиками отдельных индивидов, считали, что между расами существуют абсолютные различия. Между тем развитие популяционной генетики показало, что типологический подход к пониманию природы рас не имеет достаточной аргументации.
Научно обоснованный подход к классификациям рас является популяционным подходом. Эффективность классификаций рас зависит от количества используемых признаков, ибо чем больше признаков для проведения различий между расами, тем более эффективной оказывается классификация. Поскольку расы являются популяциями, то, изучая какую-либо расу, описывают фактически большую группу людей, но не отдельного индивида. Поскольку в пределах каждой расы имеются громадные вариации наследственных характеристик, то невозможно найти критерии, на основе которых можно было бы подтвердить или исключить принадлежность всех индивидуумов к разным расам. Точно так же, как невозможно найти индивидуумов со всеми признаками, присущими одной расе. Расы должны рассматриваться как группы, но не как индивиды. Например, сравнение черепных костей шведов и русских показало, что между этими национальными группами имеются существенные различия, но они являются средними различиями, а между индивидуумами в каждой группе имеется перекрываемость. Таким образом, определение расовых различий по популярным стереотипам (типологически) является неправильным, поскольку абсолютных различий между расовыми группами не существует.
Представления о происхождении рас и первичных очагов расообразования отражены в нескольких гипотезах. В соответствии с гипотезой полицентризма, или полифилии (Ф. Вайденрайх, 1947), существовало четыре очага расообразования - в Юго-Восточной Азии и на Больших Зондских островах, в Восточной Азии, в Африке южнее Сахары и в Европе, или Передней Азии. В Европе, или Передней Азии, сложился очаг расообразования, где на основе европейских и переднеазиатских неандертальцев возникли европеоиды. В Африке на основе африканских неандертальцев появились негроиды, в Восточной Азии на основе синантропов возникли монголоиды, а в Юго-Восточной Азии и на Больших Зондских островах развитие на основе питекантропов и яванских неандертальцев привело к формированию австралоидов. Следовательно, европеоиды и негроиды, монголоиды и австралоиды имеют свои собственные очаги расообразования. Предполагается, что главным в расогенезе
были мутации и естественный отбор. Однако эта гипотеза вызывает возражения, поскольку не известны случаи, когда бы идентичные эволюционные результаты воспроизводились несколько раз (эволюционные изменения всегда являются новыми). Отсутствуют также научные данные, подтверждающие то, что каждая раса имеет свой собственный очаг расообразования.
В соответствии с гипотезой дицентризма Т. Гексли, Н. Кинса, Р. Биасутти, А. Тома, Г.Ф. Дебеца (1960) предполагается два объяснения происхождения рас. В соответствии с первым очаг расообразования европеоидов и африканских негроидов существовал в Передней Азии, тогда как очаг расообразования монголоидов и австралоидов был приурочен в Восточной и Юго-Восточной Азии. Европеоиды передвигались в пределах европейского материка и прилегающих к нему районов Передней Азии. Напротив, негроиды расселялись в основном вдоль тропического пояса. Монголоиды вначале заселяли Юго-Восточную Азию, затем часть их перешла на американский континент.
На основании второго объяснения европеоиды, африканские негроиды и австралоиды составляют один ствол расообразования, тогда как азиатские монголоиды и американоиды - другой ствол. В обоснование гипотезы дицентризма приводят данные о сходстве строения зубов монголоидов и австралоидов. Однако эта гипотеза имеет те же возражения, что и гипотеза полицентризма.
В соответствии с гипотезой моноцентризма, или монофилии (Я.Я. Рогинский, 1949), которая основана на признании общности происхождения, социально-психического развития, а также одинакового уровня физического и умственного развития всех рас, расы возникли от одного предка, в одном месте, но протяженность территории этого места была очень большой, измеряемой многими тысячами километров. Предполагается, что формирование рас произошло на территориях Восточного Средиземноморья, Передней и, возможно, Южной Азии. По этой гипотезе одна ветвь архаичных палеоантропов дала начало монголоидам, которые как раса сформировались в несколько этапов. Архаичные палеоантропы, придя в Восточную Азию, встретили там архантропов, которых истребили или поглотили, унаследовав от них совкообразные резцы. Затем в Азию пришли люди современного типа. В результате метисации сложились протомонголоиды, часть которых 20 000-30 000 лет назад двинулась из Восточной Азии через Берингию в Северную Америку,
дав начало американским индейцам. Сохранив свой основной тип, они приобрели новые особенности. Из части протомонголоидов, оставшихся на азиатском континенте, несколько позже сформировались настоящие монголоиды. Другая ветвь архаичных палеоантропов дала начало европеоидным и негроидно-австралоидным группам. Предки европеоидов и негроидов занимали территории Восточного Средиземноморья и Южной Европы от Испании до города Воронежа в России. Формирование европеоидов и негроидов произошло в пале- олите. Позднее в результате изоляции, миграции и расселения возникли более мелкие расовые группировки.
Выделяют четыре этапа расообразования (В.П. Алексеев, 1985). На первом этапе расообразования происходило выделение первич- ных очагов расообразования (территорий, на которых происходит этот процесс) и формирование основных расовых стволов - западного (европеоиды, негроиды и австралоиды) и восточного (азиатские монголоиды и американоиды). Хронологически этот этап приходится на эпохи низшего или среднего палеолита (около 200 000 лет назад), т.е. совпадает с этапом формирования человека современного типа. Следовательно, оформление основных расовых сочетаний в западных и восточных районах Старого Света происходило одновременно с формированием признаков, присущих современному человеку, а также с переселением части человечества в Новый Свет. На втором этапе происходило выделение вторичных очагов расообразования и формирование эволюционных ветвей в пределах основных расовых стволов. Хронологически этот этап приходится на верхний палеолит и частично мезолит (около 15 000-20 000 лет назад). На третьем этапе выделялись третичные очаги расообразования, в пределах которых формировались локальные расы. По времени этот этап приходится на канун мезолита и неолита (около 10 000-12 000 лет назад). На четвертом этапе возникали четвертичные очаги расообразования, в которых формировались популяции с углубленной расовой дифференциацией, сходной с современной. Этот этап осуществлялся в эпоху бронзы и раннего железа (IV-III тыс. до н.э.).
Факторами расогенеза являются естественный отбор, дрейф генов, изоляция и смешение популяций, причем наибольшее значение имел естественный отбор, который выполнял формообразующую роль на ранних стадиях формирования рас.
Действие естественного отбора проявлялось в разных направлениях, а его эффективность зависела от селективного значения признака.
Например, за цвет кожи ответственны кожные клетки, содержащие меланин, синтез которого детерминируется генетически. Наличие пигмента определяется присутствием гена, контролирующего тирозиназу, обеспечивающую конверсию тирозина в меланин. Однако только этого гена еще недостаточно для объяснения природы цвета кожи. На пигментацию кожи оказывает влияние еще один ген, который детерминирует другой фермент, содержащийся в клетках людей с белой кожей и влияющий на количество меланина. Когда этот фермент синтезируется, то меланин образуется в малых количествах и цвет кожи становится белым. Напротив, когда он отсутствует (не синтезируется), то меланин образуется в больших количествах и цвет кожи становится черным. Имеются данные, позволяющие считать, что в определении цвета кожи принимает участие также и меланин - стимулирующий гормон. Таким образом, в контроле цвета кожи принимают участие как минимум три гена. Действие естественного отбора в случае такого расового признака, как цвет кожи, объясняется связью между солнечным освещением и продукцией витамина D, который необходим для поддержания в организме баланса кальция. Избыток этого витамина сопровождается отложением кальция в костях и способствует их хрупкости, тогда как недостаток кальция ведет к рахиту. Между тем количество витамина D, синтезируемого нормально в организме человека, контролируется количеством солнечного облучения, которое проникает в клетки глубже слоя меланина. Чем больше меланина в коже, тем меньше света, проникающего в клетки глубже меланина. Поэтому люди зависимы в продукции их витамина D от солнечного света. Чтобы витамин D продуцировался в количествах, достаточных для поддержания нормального баланса кальция, люди со светлой кожей должны находиться подальше от экватора. Напротив, люди с черной кожей должны быть ближе к экватору. Как видно, территориальное распределение людей с разной пигментацией кожи является функцией географической широты. Посветление кожи у европеоидов способствует более глубокому прохождению солнечного света в ткани человека, что ускоряет синтез противорахитического витамина D, который обычно медленно синтезируется в условиях недостатка солнечной радиации. Миграция людей с интенсивно пигментированной кожей к отдаленным от экватора широтам, а людей с недостаточно пигментированной кожей к тропическим широтам могла вести к дефициту витамина D у первых и избытку его у вторых с вытекающими
отсюда последствиями. Следовательно, цвет кожи имел селективное значение, являясь фактором контрселекции. Естественный отбор шел на цвет кожи, который возникал в качестве приспособительного признака к условиям географической среды обитания.
Другим расовым признаком, детерминирующим действие отбора, является тип телосложения. Например, эскимосы более низкорос- лы и плотны, чем зулусы. У эскимосов площадь поверхности тела меньше по сравнению с ее массой. Напротив, у зулусов, для которых характерно тонкое тело с длинными руками, площадь поверхности тела превалирует над его массой. В результате этого зулусы обладают преимуществом в выделении избытка тепла из тела, что очень важно в условиях жаркого климата. Поскольку отношение длины конечностей к массе тела изменяется у индивидов в направлении к экватору, то считают, что длинные конечности имеют преимущество по сравнению с массой тела в условиях жаркого климата. Длинные конечности обеспечивали индивидов дополнительной площадью тела, способствующей более быстрой потере тепла. Следовательно, большее значение этого отношения имело преимущество в жарком климате. Напротив, меньшее значение этого отношения имело преимущество в более прохладном климате, поскольку при меньшей площади тела в нем лучше сохраняется тепло. Таким образом, подобно цвету кожи тип телосложения коррелирует с географической широтой, на которой расселена раса, и в прошлом имел селективное значение, т.е. отбор шел на телосложение (табл. 33).
Выступающий узкий нос у европеоидов удлиняет носоглоточный путь, что способствует нагреванию холодного воздуха и защищает от переохлаждения гортань и легкие. Напротив, очень широкий и плоский нос (большее развитие слизистых) у негроидов способствует большей теплоотдаче. Их вытянутая высокая голова слабее нагревается, а курчавые волосы лучше предохраняют голову от перегрева, так как создают воздухоносную прослойку. Естественный отбор тоже шел на эти признаки, ибо они обеспечивали приспособление к условиям географической среды обитания.
Дрейф генов способен изменять генетическую структуру популяций. Подсчитано, что в результате дрейфа генов облик популя- ций может изменяться на протяжении 50 поколений, т. е. около
1250 лет.
Изоляция проявлялась в различных формах и объемах. В частности, географическая изоляция первобытных коллективов в эпоху
Таблица 33. Изменчивость отношений между длиной конечностей и массой тела в отдельных группах и популяциях
Популяция | Количество исследованных индивидов | Среднее отношение длины конечностей к массе тела | Географическая широта (примерная) |
Парижские рабочие | 73 | 4,88 | 50? с. ш. |
Французские солдаты | 50 | 4,91 | 50? с. ш. |
Франц. студенты | 8 | 4,94 | 50? с. ш. |
Финны | 47 | 4,89 | 60? с. ш. |
Британские солдаты | 300 | 5,0 | 55? с. ш. |
Украинцы | 504 | 5,08 | 50? с. ш. |
Сицилийцы | 120 | 5,09 | 40? с. ш. |
Арабы (йеменцы) | 31 | 5,63 | 20? с. ш. |
Нунгусы (Мозамбик) | 119 | 5,66 | 15? ю. ш. |
Дароды (Сомали) | 123 | 5,74 | 5? с. ш. |
Гобавины (Сомали) | 51 | 5,90 | 5? с. ш. |
Антумбы (Мозамбик) | 26 | 6,06 | 15? ю. ш. |
Кораны (Ю. Африка) | 18 | 6,21 | 25? ю. ш. |
Индийцы (Мадрас) | 35 | 6,41 | 10? ю. ш. |
Пигмеи Ака (Конго) | 95 | 6,98 | 5? ю. ш. |
Пигмеи Басуа (Конго) | 115 | 7,03 | 5? с. ш. |
палеолита сопровождалась дифференцировкой их генетического состава, прерыванием контактов с другими коллективами. Изоляция с помощью искусственных барьеров тоже оказывала влияние на генетическую дифференциацию популяций. Однако она влияла и на концентрацию культурных традиций. Например, изоляция в США индейцев от белых, а ирландцев от евреев вела и ведет к форми- рованию обособленных расовых свойств. В популяциях индейцев племени хопи, проживающих на юго-западе США, всегда изолировали мужчин-альбиносов, оставляя их в жилищах на период полевых работ. Однако это предоставляло им большую свободу в связях с женщинами племени, когда другие мужчины работали в поле, что приводило к повышению частоты рождения детей-альбиносов, к изменению генетической конституции племени. Следовательно, результатом изоляции была селекция генов альбинизма.
Смешение популяций вело к смешению генотипов их членов, к формированию новых рас. Например, североамериканская цветная раса (негритянское население США) возникла в результате смешения лесной негроидной расы с расами банту, а также с северо-западной европейской, альпийской, средиземноморской и, возможно, другими расами. Южноафриканская цветная раса возникла в результате смешения банту, бушменов и готтентотов. В настоящее время на Гавайских островах формируется новая расовая группа на основе метисации европеоидов, монголоидов и полинезийцев.
Будущее рас определяется рядом действующих в наше время факторов. Продолжается рост численности населения земного шара, увеличивается миграция, повышается частота межрасовых браков. Вероятно, в результате подвижности популяций человека и межрасовых браков в будущем начнет формироваться единая раса человечества. В то же время предполагают и другое возможное последствие межрасовых браков, связанное с формированием новых популяций с собственными специфическими сочетаниями генов.
РАСИЗМ
Как в прошлом, так и в настоящее время основу расизма составляют извращенные представления о природе человека в результате преувеличения роли биологических факторов в его индивидуальном и историческом развитии.
Расизм зародился еще в рабовладельческом строе, однако основные расистские теории, которые положили начало расизму в современном понимании, были сформулированы в XIX в. В этих теориях обосновывались либо преимущества одних рас над другими в пределах европейских рас, либо превосходство белых над черными, либо пропагандировался сионизм. Без каких-либо научных обоснований расисты считают, что расы являются самостоятельными видами, происходящими от разных обезьяноподобных предков, либо признают общность происхождения рас, но утверждают, что расы различаются по скорости и уровню развития. Расисты делят расы на «низшие» и «высшие», полагая, что высокие цивилизации якобы созданы «высшими» расами.
В XIX в. в Европе возник культ арийской расы, который берет начало от санскритской легенды о светлых покорителях темнокожих индусов и персов. В качестве «высшей» расы французский социолог
Ж. Гобино (1854) назвал «германскую» расу, выступив одновременно против браков представителей «высших» и «низших» рас.
В последующем расизм оказался связанным с евгеникой и социалдарвинизмом. Евгеника оказалась привлекательной для расистов в качестве орудия улучшения расовых «достоинств». Что касается социал-дарвинизма, то он оказался выгодным для расистов по той причине, что открывал возможность рассматривать общественные явления на биологической основе. В начале XX в. для обоснования расового превосходства начали использовать данные генетики. Расисты стали утверждать о превосходстве в США белых над неграми и о биологической неполноценности негров, о вредности межрасовых браков.
В 20-е гг. XX в. в расизме появилось стремление найти автономные генетические факторы, детерминирующие признаки «высшей» расы, а также связать расизм с необходимостью геноцида. Это стремление привело к объединению расизма с фашизмом. «Расоведение» было возведено в фашистской Германии в ранг преступной государственной политики с геноцидом на оккупированных территориях.
Современные обоснования расизма - это смесь старых расистских и евгенических идей и их «доказательств» с современными биологизаторскими концепциями человека. Характерная особенность современного расизма заключается в том, что из расистского словаря совершенно исчезли старые доводы в пользу большей чувствительности «низших» рас к болезням. Напротив, появились доводы в пользу превосходящих умственных способностей «высших» рас. Современный расизм - это форма буржуазной идеологии, сущность которой составляет воинствующий шовинизм.
В научном плане расизм не выдерживает критики. Используемое для обоснования существования «низших» рас понятие о «чистых» расах - это расистское понятие, по которому представители «чистых» рас гомозиготны по всем локусам (имеют одинаковые аллели на каждом генетическом локусе). Генетической идентичности нет не только в случае рас, но даже в отдельных семьях. Полная гомозиготность существует только в случае однояйцовых близнецов. Нет также никаких оснований считать, что чистые расы существовали в прошлом, ибо в пользу этого допущения нет ни одного доказательства. Напротив, вся эволюционная история человечества - это непрерывное смешение рас. Таким образом, утверждение о том, что раса является хорошо дифференцированной генетической группой, неверно. Между тем
наука не располагает данными о связи между признаками, на основе которых идентифицируют расу, и психическими свойствами индивидуумов. Ни одна из рас не обладает монопольно «хорошими» генами и не является полностью свободной от «плохих» генов.
Расисты утверждают, что «чистые» расы (которых, фактически нет) обладают лишь «хорошими» генами, в том числе «хорошо» детерминирующими психические свойства. Гены, которые не желательны в одних географических и культурных условиях, могут быть желательны в других условиях. Наконец, многие из так называемых «превосходящих» и «отсталых» характеристик, иногда связанных с определенными расовыми группами, могут быть в основном или полностью результатом воздействия среды. Никто и никогда не показывал существования особых генов, контролирующих психическую деятельность людей, принадлежащих к той или иной расе. В формировании интеллекта, видимо, участвуют многие факторы. Каждый индивидуум должен оцениваться на основе его собственных характеристик, а не характеристик его расы.
Утверждения о различиях между расами по умственным способностям основаны на извращениях понятий равенства и неравенства, идентичности и разнообразия, недооценке роли среды. Равенство и неравенство - это понятия социальные и этические, тогда как идентичность и разнообразие - это понятия, применяемые к характеристикам генотипов людей. Любой индивидуум после рождения оказывается в определенной среде. Поскольку гены проявляются в одной среде и не проявляются в другой, то с учетом нормы реакции это означает, что не все гены, контролирующие физические свойства чело- века, могут проявляться в условиях той или иной среды и что сходные гены проявляются в разных средовых условиях. Но в отличие от среды животных среда человека является средой социальной. Благодаря сознанию человек действует в сфере общественной формы движения материи, подчиняет себе среду обитания и в конечном итоге создает надбиологическую среду. Гены интеллектуальности, если они есть, не обладают особенностями, позволяющими им определять верхние или нижние границы в развитии духовного облика индивидуума, группы индивидуумов, расы. Разные особенности в проявлении духовного потенциала расы определяются не генетическими недостатками или дефицитами, а генетическим разнообразием, которое складывается на основе генетических механизмов и проявляется в зависимости от разнообразия социальной среды.
Любой индивид независимо от расы развивается по генетической программе в результате взаимодействия с социальной средой. Следовательно, в онтогенезе любой индивид любой расы есть продукт собственной наследственности и социальной среды, тогда как в филогенезе человек является продуктом общественного развития, а расы являются результатом действия генетических механизмов лишь на основе полиморфизма человека (цвет кожи, группа крови, строение наружного уха, форма носа, структура волос и т.д.). Следовательно, расовые различия касаются чисто физических признаков, не имеющих отношения к чисто человеческим признакам, т.е. расы не имеют различий по чисто человеческим признакам. Для каждой расы характерно генетическое разнообразие, которое создает потенциал расы. Между расами есть различия лишь в общественноэкономическом развитии, однако эти различия не основываются на биологических закономерностях. Они являются результатом исторического и социального развития. Русский ученый Н.Н. МиклухоМаклай (1848-1888) еще в конце XIX в. доказал, что аборигены разной расовой принадлежности (из Новой Гвинеи, Микронезии и других районов мира) ничем не отличаются по умственным способностям от европеоидов.
ЭКОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ И ИЗМЕНЧИВОСТЬ СОВРЕМЕННОГО ЧЕЛОВЕКА
Человечество чрезвычайно расселено по земному шару, занимая районы, различающиеся по климату, ландшафту, геохимическим и другим особенностям. Между тем действие климатических особенностей, а также влияние гравитации, электромагнитного поля, радиации, патогенных организмов и других факторов сопровождается географической изменчивостью морфологических и физиологических свойств людей. Приуроченность этих свойств к определенным территориям свидетельствует об экологической изменчивости современного человека.
Среди населения арктических групп увеличено количество индивидов мускульного типа. Все члены этих групп характеризуются повышенным уровнем основного обмена, поглощения кислорода, энергетических процессов. У индивидов большинства арктических групп отмечается высокое содержание холестерина в крови. Жители
континентальных районов Сибири по сравнению с аборигенами Арктики чаще принадлежат к астеническому и пикническому типам телосложения. Они характеризуются относительной коротконогостью и длиннорукостью, более плоской грудной клеткой, увеличением жирового компонента тела. Для них характерна более высокая по сравнению с жителями умеренных районов теплопродукция, но одинаковый уровень холестерина в крови.
Люди высокогорных групп (горцы Кавказа, Памира и ТяньШаня, коренные жители Эфиопии и Индии, индейцы Перу и др.) характеризуются увеличенной емкостью грудной клетки и костномускульной массы тела. Для них характерен повышенный уровень эритроцитов (гемоглобина). Жители тропических широт Африки, Австралии, Океании, Индии и Америки характеризуются приспособленностью (защитой) их организмов к температуре, наличием защиты от перегрева. Для них характерна удлиненная форма тела, недостаточное развитие мускулатуры, повышенное количество потовых желез, усиленная теплоотдача и пониженный уровень энергетических процессов. Кроме того, им свойствен повышенный уровень иммуноглобулинов и пониженный уровень холестерина в крови. Для коренного населения пустынь характерны высокорослый тип тела, более низкое кровяное давление, повышенное содержание эритроцитов в крови.
Население зон умеренного климата по морфологическим и функциональным свойствам занимает среднее положение между жителями арктических и тропических групп. Жители умеренных зон подвержены влиянию геохимических свойств среды и высоты над уровнем моря. Например, минерализация их скелета зависит от содержания макро- и микроэлементов в почве и воде.
На основе зональной зависимости морфофункциональной изменчивости разных популяций человека предполагают существование адаптивных типов, которые не зависимы ни от расовой, ни от этни- ческой принадлежности и определяются нормой реакции, обеспечивающей равновесие популяций со средой. Адаптация человека к среде связана с изменением его морфологических и физиологических свойств. Поэтому одинаковые черты приспособленности к условиям тропических зон характерны как для коренных жителей Африки (негроидов), так и для европеоидов Индии, австралоидов. Единые черты приспособленности характерны также для жителей Крайнего Севера (ненцы, чукчи, эскимосы, саамы).
Адаптивность человека имеет исторический характер. Предполагают, что у австралопитеков приспособительные реакции заключались в адаптации их к климату тропической зоны, а у архантропов эти реакции развивались в направлении формирования приспособленности к влажному тропическому и высокогорному климату. Расселение палеоантропов в Европе сопровождалось формированием адаптивного типа умеренного пояса (эпоха среднего палеолита). Арктические адаптивные типы возникли, вероятно, в эпоху верхнего палеолита.
Считают, что физический тип человека не изменился за последние 35 000-40 000 лет. Почти не изменился и интеллект человека. Однако экологические факторы сейчас воздействуют на человека больше, чем даже в прошлом веке. Поэтому современной тенденцией физического облика человека сейчас стали акселерация и секулярный тренд.
Акселерация (от лат. acceleration) - это ускорение роста людей и проявления их физиологических функций. Например, в начале XX в. длина тела у мужчин достигала своих обычных размеров к 25-26 годам, в настоящее время - к 18-19 годам. Будущее человека нельзя рассматривать без учета судеб этноса.
Секулярный тренд (от лат. secular trend - вековая
тенденция) - это увеличение длины тела, репродуктивного периода,
продолжитель- ности жизни и других важнейших свойств человека в
определенные (длительные) интервалы времени. Например, в России было
отмечено увеличение длины тела на
КУЛЬТУРНОЕ РАЗВИТИЕ ЧЕЛОВЕКА
В археологии четвертичный период подразделяют по-другому в сравнении со шкалой геологического времени. Археология в преде- лах четвертичного периода вместо эпох выделяют века, в частности древнекаменный (палеолит), среднекаменный (мезолит) и новокаменный (неолит) века, а также еще более поздние другие века, приуроченные к современному человеку и его культуре. Между геологическим и археологическим временем имеется некоторое совпадение. В целом палеолит соответствует плейстоцену, а мезолит и неолит - периоду после плейстоцена.
Данные археологии свидетельствуют о том, что появление предков современного человека сопровождалось формированием начал культуры. Биологическая эволюция человека заняла около 14 млн лет, тогда как начало его культурного развития датируется временем порядка 2-3 млн лет от наших дней, т.е. четвертичным периодом. Тем не менее биологическое и культурное развитие человека взаимосвязаны. Биологическое развитие оказывало влияние на культурное развитие, и наоборот. Культурное развитие влияло на физическое и психическое развитие человека и было плодотворным с самых первых шагов культуры, но на последних этапах эволюции человека оно приобрело решающее значение.
Начало культурного развития человека связано с его орудийной деятельностью. Рассматривая первые результаты орудийной деятель- ности человека (каменные и костяные орудия), нельзя не заметить, что в различных географических районах она характеризовалась разным уровнем обработки. Это объясняют обычно географическим местом обитания популяций, влиянием среды их обитания и степенью изолированности. Умение в изготовлении орудий и качество изготовляемых орудий послужили основанием для квалификации их в качестве культур. Характерная особенность этих культур заключается в их сменяемости. Но нельзя не отметить, что сменяемость отдельных из них задерживалась на очень долгое время по ходу истории человечества. Например, уровень культур некоторых аборигенов на африканском и австралийском континентах остается и сейчас почти на уровне каменного века.
Считают, что первые самые простые орудия из камня и кости были изготовлены Н. habilis, которому были известны орудия как из каменных отщепов, так и из крупных галек (олдованская культура). Для Н. erectus культура отщепов и культура ручных рубил также были хорошо известны в раннем палеолите, когда шло непрерывное совершенствование орудий, возникали стоянки.
В нижнем палеолите каменные орудия (ручные рубила) характеризовались лучшей обработкой, о чем свидетельствуют находки в пещере Азых (Азербайджан). Этот тип культуры известен под названием ашельской и шельской, которые значительно выше олдованской культуры.
Развитие культур нарастало в среднем палеолите. Тогда неандертальцами была создана так называемая мусьерская культура (на территории современной Франции), которая характеризовалась более
высоким уровнем изготовления каменных орудий. Одновременно появляются орудия и из кости (остроконечники, скребки и др.). Мусьерская культура длилась примерно 100 тыс. лет.
Особенно бурное развитие культуры приходится на поздний палеолит (35-40 тыс. лет назад). Увеличение числа стоянок и жителей в них вело к совершенствованию методов охоты, к дифференциации каменного инвентаря. Появляются инструменты для изготовления орудий. В конце последнего ледникового периода неандертальцами были разработаны довольно совершенные методы выделки кожи, возникли начала искусства. В верхнем палеолите искусство поднимается на новый уровень, о чем свидетельствуют образцы пещерной живописи (изображения животных и различных сцен на стенах пещер), найденные в Африке и Европе. К тому времени относятся первые погребения и начало формирования ритуальных церемоний.
Для древнейших и древних людей характерен каннибализм и ритуальный характер каннибализма. Считают, что неандертальцы имели представления о посмертной жизни. Главным оружием человека в мезолите была рогатина (пика) из тиса с острым каменным наконечником. Достигли значительного совершенствования способы охоты, рыболовства. Неандертальцы и ранние кроманьонцы уничтожали мамонтов, бизонов, мегатериев. Появляются лук и стрелы. Стали строится жилища.
В неолитическом веке человек стал уходить от стадного образа жизни. Этим временем датируется изобретение керамики, переход к оседлому образу жизни, образование дифференцированных общественных ячеек. От собирания и охоты он стал переходить к сельскому хозяйству (выращивание растений и разведение животных). На европейском континенте начала сельского хозяйства, сопровождаемые доместикацией растений и животных, а также неолитической технологией, восходят к IX-VI тыс. до н. э. Если 15 тыс. лет назад появился хлеб в виде поджаренных кусков зерновой массы, то уже 5-6 тыс. лет назад люди стали использовать «кислый» хлеб.
Каждая популяция людей вносила свой вклад в создание культур на ранних этапах цивилизации. Последующие поколения людей формировали более совершенные цивилизации. Благодаря распределению работы между разными социальными группами, конструированию новых инструментов и технологий, доместификации животных и растений, а также развитию экономических, социальных и политических институтов люди современного физического
типа создали чрезвычайно высокую цивилизацию, передаваемую от одних поколений к другим посредством языка (речи), письма, печати, а в новое время - посредством телефона, радио, телевидения и компьютерной техники. Каждая социальная группа стала действовать как часть экологического сообщества людей. В результате объединения и взаимодействия люди стали производить, хранить и транспортировать пищу в больших количествах, строить железные и шоссейные дороги, выпускать самолеты и космическую технику, запускать искусственные спутники Земли, обеспечивать общественный порядок, санитарное благополучие и т.д. Адаптация, одежда и жилища способствовали развитию жизни человека в самых различных климатических условиях.
В заключение важно еще раз подчеркнуть, что биологическое и культурное развитие человека шло синхронно. Тем не менее в раз- витии региональных сообществ имеются различия. Одни сообщества людей ушли далеко вперед, другие отстали в своем развитии. Эти различия еще не получили достаточного объяснения, хотя в числе причин, объясняющих отсталость отдельных народов, часто называют сверхиспользование природных ресурсов или исходный недостаток природных ресурсов, тяжелые климатические условия, войны, а также недостаточность регулирующих механизмов развития. Возможны и другие причины, но их обсуждение выходит за рамки данной темы.
ВОПРОСЫ ДЛЯ ОБСУЖДЕНИЯ
1. Когда стали интересоваться происхождением человека? Почему проблемы антропогенеза и расогенеза привлекают такое большое внимание сейчас?
2. В чем заключается концепция животного происхождения человека? Имеет ли она доказательства? Известны ли другие (альтернативные) концепции?
3. Назовите свойства, по которым человека отличают от животных. Могут ли современные человекообразные обезьяны эволюционировать в сторону человека?
4. Каково значение ископаемых останков в изучении антропогенеза? Какова роль других биологических наук в изучении антропогенеза?
5. Назовите основные этапы антропогенеза. Можно ли построить эволюционные ряды в применении к человеку?
6. Какое значение в эволюции человека имело появление прямохождения и развитие головного мозга?
7. В чем заключаются особенности Н. kahilis, каков его возраст и можно ли считать его эволюционной линией на пути к Н. sapiens?
8. Перечислите факторы антропогенеза, обратив внимание на специфические.
9. Когда и как естественный отбор действовал в эволюции человека? Действует ли сейчас естественный отбор в популяциях человека?
10. Какова роль труда в антропогенезе?
11. Как вы понимаете биологическую и социальную сущность человека?
12. В чем заключается ошибочность типологического подхода к определению рас? Приведите научное определение расы. Какова роль популяционного подхода к научному определению рас?
13. Есть ли разница между терминами «вид» и «раса»? Если имеется, то покажите ее на двух-трех примерах.
14. Приведите существующие классификации рас. Назовите основные расы рода человеческого. Что вкладывается в понятия «старые» и «новые» расы? С какой скоростью происходит расогенез?
15. Перечислите факторы расогенеза. В чем заключается различие в действии естественного отбора при видообразовании и расогенезе?
16. Существуют ли «чистые» расы? В чем заключается научная несостоятельность расизма?
17. Что понимают под конституциональными типами людей? Какие вы знаете классификации конституциональных типов и насколько они совершенны?
18. Что вы понимаете под географической изменчивостью человека? Есть ли разница между географической и экологической изменчивостью человека?
19. Какова связь между биологическим и культурным развитием человека?
20. Какое значение для студента-медика имеет изучение данных о происхождении человека?