Биология: учебник / Пехов А.П., -, 2010. - 664 с.
|
|
|
|
ГЛАВА II КЛЕТКА - ОСНОВА ЖИЗНИ
ИСТОРИЧЕСКИЕ СВЕДЕНИЯ
Клетки - это основная единица живого (биологической активности), ограниченная полупроницаемой мембраной и способная к самовоспроизведению в среде, не содержащей живых систем. Жизнь начинается с клетки. Вне клеток нет жизни.
Первые исследования клеток
восходят к XVII в., когда англичанин Роберт Гук (1635-1703),
рассматривая под примитивным микроскопом срезы пробки, в
В
В
1. Организмы состоят из клеток и продуктов их жизнедеятельности, причем клетки являются главной структурной единицей растений и животных.
2. Размножение клеток лежит в основе роста животных и растений.
Выдающийся вклад в дальнейшее развитие клеточной теории принадлежит Р. Вирхову, сформулировавшему в
положение «cellula e cellula» («каждая клетка из клетки»), означающее, что клетка может возникнуть лишь из предшествующей клетки и что других путей появления клеток не существует.
Современный этап клеточной теории характеризуется дальнейшим развитием установленных ранее основных положений о клетке и новыми обобщениями результатов, полученных при изучении синтеза нуклеиновых кислот, а также регуляции активности генов. Имеет новое обоснование положение теории о том, что организмы состоят из клеток и что клетка является основной структурнофункциональной единицей живого, вне которой нет жизни. Клетки - высокоорганизованные дифференцированные образования. Установлено, что активность организмов обусловлена активностью его клеток и рост, развитие и дифференцировка тканей зависят от образования новых клеток. Через клетки происходят поглощение, превращение, запасение и использование веществ и энергии. Структуры клеток являются «ареной», на которой осуществляются многочисленные биологические реакции.
МЕТОДЫ ИЗУЧЕНИЯ КЛЕТОК
Всеобъемлющим подходом к изучению клеток является системноструктурный подход.
Для изучения клеток используют микроскопическую технику в виде световой, фазово-контрастной, ультрафиолетовой, люминесцентной и электронной микроскопии. Последняя применяется в сочетании с техникой ультратонких срезов. Для получения трехмерных изображений клеток используют сканирующие электронные микроскопы. Для документации поведения живых клеток применяют замедленную киносъемку.
В цитологических исследованиях очень эффективны также цитохимические методы, основанные на том, что определенные реактивы (краски) избирательно окрашивают химические вещества цитоплазмы, и авторадиография - введение в клетки радиоактивных изотопов фосфора (32Р), углерода (14С) и водорода (3Н) с последующим обнаружением их клеточной локализации на фотоэмульсиях. Для выделения клеточных компонентов используют дифференциальное центрифугирование, а для разделения биологических молекул - хроматографию и электрофорез. Рентгеноструктурный анализ позволяет определять пространственное расположение молекул
и различных веществ, расстояние между отдельными молекулами, объем, форму и другие свойства молекул. Для изучения клеток используют также биохимические, генетические и иммунологические методы в сочетании с культивированием клеток на искусственных питательных средах.
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ КЛЕТОК
Различают прокариотические и эукариотические клетки. Первые типичны для одноклеточных организмов-прокариотов, которыми являются в основном бактерии, тогда как вторые характерны для организмов-эукариотов, большинство которых - многоклеточные организмы (табл. 1).
Таблица 1. Основные свойства прокариотических и эукариотических клеток
Свойство | Прокариоты | Эукариоты |
Капсула | Имеется у отдельных видов | Отсутствует |
Клеточная стенка | Имеется | Имеется в клетках растений, отсутствует в клетках животных |
Плазматическая мембрана | Имеется | Имеется |
Ядерная мембрана | Отсутствует | Имеется |
Количество хромосом | Одна | От нескольких до многих |
Количество групп сцепления | Одна или две | От нескольких до многих |
Химический состав хромосом | ДНК или две | Нуклеопротеиды |
Митохондрии | Отсутствуют | Имеются |
Деление | Простое | Митоз |
Клетки характеризуются значительным структурно-функциональным разнообразием. Самая простая организация характерна для прокариотических клеток, среди которых наиболее просто устроены микоплазмы. Микоплазменные клетки имеют овальную форму, а их размеры составляют 0,1-0,25 мкм в диаметре. Для них характерно наличие наружной плазматической мембраны (толщина около 8 нм), окружающей цитоплазму, в которой имеются молекула
ДНК, РНК, рибосомы и различные включения в виде гранул липидов и других веществ.
Более сложными прокариотическими клетками являются бактерии и синезеленые водоросли (цианобактерии). Бактерии имеют форму палочек или округлых образований микроскопических размеров. Например, размеры одиночной клетки E. coli (рис. 1) составляют 1 мкм в длину и 0,5-0,8 мкм в диаметре, объем - около 1 мкм3, а масса равна 10-12 г.
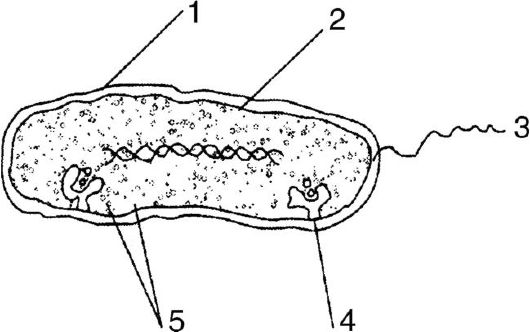
Рис. 1. Бактериальная клетка: 1 - клеточная стенка; 2 - плазматическая мембрана; 3 - жгутик; 4 - лизосома; 5 - митохондрия
Бактериальная клетка окружена трехслойной клеточной оболочкой толщиной около 40 нм, представляющей собой «мешок» или «конверт», в котором заключено клеточное содержимое в виде примерно 2?10-13 г белка, 610-15 г ДНК и 2?10-14 г РНК (в основном рибосомной РНК). В бактериальной клетке синтезируется около 2000 разных белков, большинство которых содержится в цитоплазме. Концентрация одних белков составляет 10-8 М, тогда как в других она достигает 2?10-4 М (от 10 до 200 000 молекул на клетку).
«Конверт» состоит из трех частей (рис. 2), из которых две части представлены наружным и внутренним слоями, являющимися мембранами и построенными преимущественно из липополисахаридов. Внешняя поверхность наружной мембраны в основном состоит из липополисахаридов, которые прикрепляются к располагающимся там же липидам. В состав наружной мембраны входят также белки. Внутренняя мембрана, называемая цитоплазматической, состоит из многих белков, включенных в фосфолипидный бислой (двойной слой).
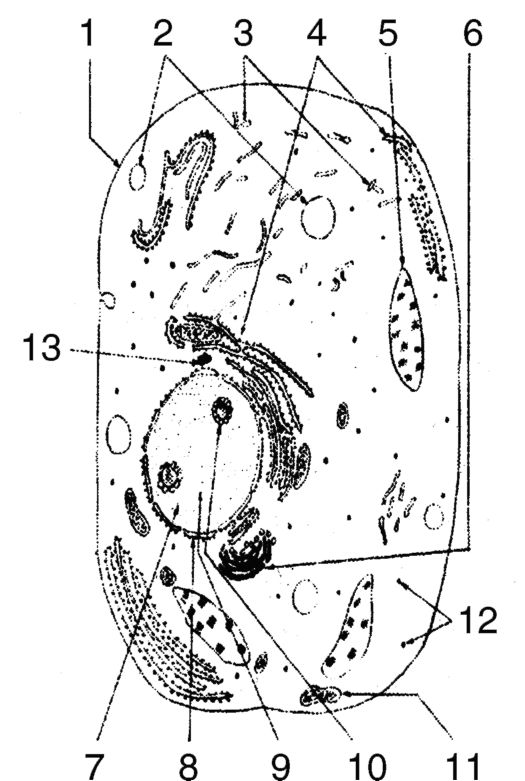
Рис. 2. Соматическая клетка: 1 - клеточная мембрана; 2 - вакуоль; 3 - эндоплазматическая сеть (гладкая);
4 - эндоплазматическая сеть (шероховатая);
5 - жюсома; 6 - комплекс Гольджи; 7 - ядро;
8 - ядерная оболочка; 9 - хромосома; 10 - ядрышко; 11 - митохондрия; 12 - рибосома; 13 - центриоль
Третью часть «конверта» составляет пептидогликановый слой, который непосредственно представляет собой клеточную стенку, лежащую между наружной и внутренней (цитоплазматической) мембраной. Определяя форму бактериальной клетки, пептидогликановый слой в химическом плане является одиночной молекулой, содержащей полисахаридные цепи, связанные с короткими пептидами. Внешняя мембрана прикреплена к пептидогликановому слою большим количеством (106) молекул липопротеида, протеиновый конец которых ковалентно прикреплен к диаминопимелиновой кислоте пептидогликана, тогда как их липидный конец «спрятан» во внешней мембране.
Пространство между наружной и внутренней мембраной, содержащее пептидогликановый слой, носит название периплазматиче- ского пространства.
На долю мембран и клеточной стенки приходится около 20% всего клеточного белка. На поверхности клеточной стенки у бактерий многих видов могут быть жгутики или жгутики и пили, а снаружи от клеточной стенки может находиться также капсула, как, например, у пневмококков. Эти структуры обладают диагностическим значением.
Основное вещество бактерий - это цитоплазма, представляющая собой раствор белка, концентрация которого составляет 200 мг/мл.
В цитоплазме бактерий имеются ядерная область, рибосомы (около 10 000) и различные включения в виде гранул гликогена, липидов, серы. В ядерной области обнаруживаются волокна диаметром 3-5 нм, представляющие собой скрученные двойные цепи одиночной молекулы ДНК.
Клетки синезеленой водоросли сходны по своему строению с бактериями, но в отличие от последних содержат хлорофилл и каротин.
Наиболее сложная организация присуща эукариотическим клеткам животных и растений. Строение клеток животных и растений характеризуется принципиальным сходством, но форма, размеры и масса их чрезвычайно разнообразны и зависят от того, является ли организм одноклеточным или многоклеточным. Например, диатомовые водоросли, эвгленовые, дрожжи, миксомицеты и простейшие - одноклеточные эукариоты, тогда как организмы подавляющего большинства других типов это многоклеточные эука- риоты, количество клеток у которых составляет от нескольких (например, у некоторых гельминтов) до миллиардов (у млекопитающих) на организм. Организм человека состоит из около 1015 разных клеток, которые различаются между собой по осуществляемым ими функциям.
В случае человека насчитывают более 200 типов клеток. Наиболее многочисленными клетками в организме человека являются эпите- лиальные, среди которых различают ороговевающие клетки (волос и ногтей); клетки, обладающие всасывательной и барьерной функциями (в желудочно-кишечном тракте, мочеполовых путях, роговице, влагалище и других системах органов); клетки, выстилающие внутренние органы и полости (пневмоциты, серозные клетки и многие другие). Различают клетки, обеспечивающие метаболизм и накопление резервных веществ (гепатоциты, жировые клетки). Большую группу составляют клетки, секретирующие внеклеточный матрикс (амилобласты, фибробласты, остеобласты и др.) и гормоны, а также сократительные клетки (скелетных и сердечных мышц, радужной оболочки и других структур), клетки крови и иммунной системы (эритроциты, нейтрофилы, эозинофилы, базофилы, Т-лимфоциты и др.). Существуют также клетки, выполняющие роль сенсорных преобразователей (фоторецепторы, осязательные, слуховые, обонятельные, вкусовые и другие рецепторы). Значительное число клеток представлено нейронами и глиальными клетками ЦНС. Имеются также специализированные клетки хрусталика глаза, пигментные
и питающие клетки. Далее следует назвать половые клетки. Известны и многие другие типы клеток человека.
В природе не существует некоей типичной клетки, ибо все они характеризуются чрезвычайным разнообразием. Тем не менее все эукариотические клетки существенно отличаются от прокариотических клеток по ряду свойств, и прежде всего по объему, форме и размерам. Объем большинства эукариотических клеток превышает объем прокариотов в 1000-10 000 раз. Такой объем прокариотических клеток связан с содержанием в них различных органелл, осуществляющих всевозможные клеточные функции. Для эукариотических клеток характерно также наличие большого количества генетического материала, сосредоточенного в основном в относительно большом количестве хромосом, что обеспечивает им большие возможности в дифференцировке и специализации. Не менее важной особенностью эукариотических клеток является то, что им присуща компартментализация, обеспеченная наличием внутренних мембранных систем. В результате этого многие ферменты локализуются в определенных компартментах. Например, почти все ферменты, катализирующие синтез белков и животных клеток, локализованы в рибосомах, тогда как ферменты, катализирующие синтез фосфоли- пидов, сосредоточены в основном на клеточной цитоплазматической мембране. В отличие от прокариотических клеток в эукариотических клетках имеется ядрышко.
Эукариотические клетки по сравнению с прокариотическими обладают более сложной системой восприятия веществ из окру- жающей среды, без чего невозможна их жизнь. Существуют и другие различия между эукариотическими и прокариотическими клетками. Метаболиты могут изменять объем клеток.
Форма клеток бывает самой разнообразной и часто зависит также от выполняемых ими функций. Например, многие простейшие имеют овальную форму, тогда как эритроциты являются овальными дисками, а мышечные клетки млекопитающих вытянуты. Размеры эукариотических клеток микроскопические (табл. 2).
Некоторые виды
клеток характеризуются значительными размерами. Например, размеры
нервных клеток у крупных животных достигают нескольких метров в длину, а
у человека - до
Считают, что чем крупнее организм в пределах вида, тем крупнее его клетки. Однако для родственных видов животных, разли-
чающихся по размерам, характерны и сходные по размерам клетки. Например, у всех млекопитающих сходны по размерам эритроциты.
Таблица 2. Средние размеры клеток животных и растений
Происхождение клетки | Диаметр, мкм | Объем, мкм3 |
Клетка печени человека | 20 | 4000 |
Малая клетка тимуса | 6 | 120 |
Клетка меристемы (корешок лука) | 17 | 2600 |
Клетка паренхимы плода растения | 1000 | 1-108 |
Клетки различаются также и по массе. Например, одиночная клетка печени (гепатоцит) человека весит 19-9 г.
Соматическая клетка человека (типичная эукариотическая клетка) представляет собой образование, состоящее из множества структурных компонентов микроскопических и субмикроскопических размеров (см. рис. 2).
Использование электронной микроскопии и других методов позволило установить чрезвычайное разнообразие в структуре как оболочки и цитоплазмы, так и ядра. В частности, был установлен мембранный принцип строения внутриклеточных структур, исходя из которого различают ряд структурных компонентов клетки, а именно:
Мембранная система.
Цитоплазматический матрикс (основное вещество клеток). Клеточные органеллы (внутриклеточные компартменты). Клеточные включения.
Мембранная система. Эта система представлена клеточной плазматической (цитоплазматической) мембраной, цитоплазматической (эндоплазматической) сетью (ретикулум) и пластинчатым комплексом Гольджи.
Плазматическая (цитоплазматическая) мембрана имеет толщину 8-12 нм и состоит из трех слоев, два из которых являются белковыми слоями толщиной по 3 нм каждый, а третий (внутренний) - двойным фосфолипидным слоем толщиной 6 нм (рис. 3). Плазматическая мембрана - полупроницаемая структура. Через нее в клетку входят питательные вещества и выходят все «отходы» (продукты секреции). Она создает барьер проницаемости. В результате этого плазматическая мембрана регулирует обмен различными веществами между клеткой и внешней средой. В плазматической мембране содержатся многие важные ферменты и системы активного транспорта ионов натрия и калия.
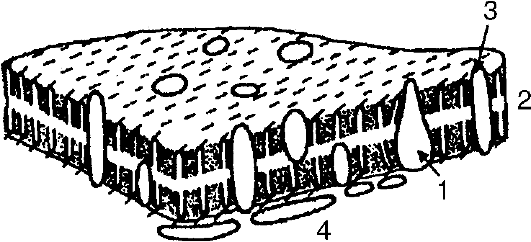
Рис. 3. Модель структуры мембраны: 1 - белки; 2 - двойной слой фосфолипидов; 3 - углеводы на наружной поверхности мембраны; 4 - углеводы на внутренней
поверхности мембраны
На поверхности плазматической мембраны обнаруживают ряд специальных образований в виде микроворсинок и ресничек. Микроворсинки очень часты в эпителии кишечника и почек. Реснички - это своеобразные выросты цитоплазмы. У эритроцитов мембрана является гладкой (элементарная мембрана). У некоторых одноклеточных организмов-эукариотов плазматическая мембрана также содержит реснички (микроворсинки), различные выпячивания и выросты, переходящие в пузырьки. На внешней поверхности клеток животных обнаружены гликопротеиды как компоненты плазматической мембраны. Предполагают, что поверхностные гликопротеиды обеспечивают адгезионную способность клеток в тканях и, следовательно, слипание однотипных клеток. В мембранах эритроцитов содержится гликопротеид, получивший название гликофорина (м.м. 30 000). Этот гликопротеид состоит из 130 аминокислотных остатков и большого количества (60% всей молекулы) остатков сахара. Кроме того, в эритроцитарной мембране содержится белок спектрин, молекулы которого формируют скелет мембраны.
У клеток растений наружной структурой служит жесткая клеточная стенка, построенная из молекул целлюлозы, создающих очень прочные волокна, погруженные в матрикс из других полисахаридов и полимерного вещества лигнина. Клетки наружных слоев растений иногда покрыты очень тонким слоем восковидного вещества. На поверхности плазматических мембран имеются электрически заряженные группы, из-за которых поддерживается разность элек- трических потенциалов на мембранах. На поверхности плазмати-
ческих мембран имеются также специфические рецепторы (участки распознавания) для гормонов и других соединений. Кроме того, здесь же локализированы особые рецепторы, ответственные за индивидуальную тканевую совместимость. Считают, что рецепторные участки формируются гликопротеидами и ганглиозидами.
Цитоплазматическая (эндоплазматическая) сеть (ретикулум) представлена
пронизывающими однослойными мембранными полостями (трубочками,
цистернами, вакуолями) разных размеров, заполненными белковыми
гранулами. Открыта К. Портером в
Различают гранулярный (шероховатый) эндоплазматический ретикулум, который выстлан множеством рибосом диаметром порядка 21-25 нм и молекулярной массой 4?106, служащих центрами синтеза молекул белков, и агранулярный (гладкий) эндоплазматический ретикулум, на котором нет рибосом, но на нем синтезируются липиды и углеводы. Степень насыщенности гранулярной эндоплазматической сети рибосомами определяет степень интенсивности синтеза белков. У человека и животных агранулярной сетью богаты клетки коры надпочечников, яичников и семенников, печени, скелетных мышц.
Цитоплазматическая сеть без перерыва соединена с цитоплазматической мембраной, ядерной мембраной и пластинчатым комплексом Гольджи. Это позволяет синтезируемым белкам проходить в комплекс Гольджи, откуда после специальной обработки они выводятся из клетки или идут на построение лизосом.
Плазматическая мембрана, мембраны эндоплазматической сети, а также ядер, митохондрий и хлоропластов (см. ниже) представляют собой чрезвычайно сложные структуры, обладающие рядом важнейших биологических свойств. Многие мембраны содержат ферменты, транспортные системы, с помощью которых осуществляется перенос молекул питательных веществ и неорганических ионов внутрь клеток и внутри клеток, а также вывод из клеток продуктов жизнедеятельности. Мембранные структуры способны к самовосстановлению, если в них по каким-то причинам возникают повреждения.
В процессе искусственного растирания или гомогенизации клеток с экспериментальными целями образуются мелкие частицы диа- метром 50-150 нм, состоящие из фрагментов эндоплазматического ретикулума и плазматической мембраны. Эти структуры получили название микросом, и их широко используют в лабораторной рабо-
те для решения тех или иных вопросов молекулярной организации клеток.
Комплекс Гольджи. Этот комплекс, называемый еще пластинчатым, был открыт итальянцем Камилло Гольджи еще в
Цитоплазматический матрикс. Этот
структурный компонент является основным веществом (цитоплазмой,
гиалоплазмой) клетки. Первые электронно-микроскопические изображения
цитоплазмы были получены шведским ученым Ф. Шестрандом еще в
В цитоплазме содержатся микрофиламенты (нити) толщиной 4-5 нм и микротрубочки, представляющие собой полые цилиндрические структуры диаметром 25 нм, а также фрагменты проме-
жуточных размеров. Эти структуры составляют жесткую конструкцию (каркас) в клетке, называемую цитоскелетом и определяющую внешний вид и форму клеток. Микрофиламенты состоят из белка, сходного с сократительным белком актином.
Объединяясь, микрофиламенты формируют пучки, в которые входят дополнительные белки (анкерин, спектрин и др.). Основная функция микрофиламентов заключается в обеспечении сократительных процессов клеток, в упрочении мембран. Микротрубочки построены из белков α- и β-тубулина, а также γ-тубулина. Для микротрубочек характерен ряд функций. Формирование микротрубочек происходит в интерфазе клеток в так называемых центрах организации микротрубочек (ЦОМ), которые «окружают» центриоли, в результате чего предполагают, что центриоли являются ЦОМ. В каждом ЦОМ содержится по 10-13 молекул γ-тубулина и примерно по 7 молекул других белков, включая димер α/β-тубулина. Эти белки формируют структуру, которая образует микротрубочный «ансамбль». Их значение до конца не выяснено, но предположительно заключается в том, что они обеспечивают перемещение клеточных органелл, включая хромосомы, внутри клеток.
В составе цитоплазмы обнаруживают ферменты, полисахариды, АТФ, тРНК, ионы Са, Na, К и других химических элементов.
Основная функция цитоплазматического матрикса заключается в том, что он является внутренней средой клетки, поддерживающей мембранные системы, органеллы и включения. Цитоскелет выполняет опорную функцию. Микрофиламенты способствуют упрочению мембранной системы, а микротрубочки, как отмечено выше, обеспечивают перемещение клеточных органелл и транспорт химических соединений из одних отсеков клетки в другие. Цитоскелет имеет значение также в делении клетки.
Клеточные органеллы. Эти структуры представлены ядром, хромосомами, ядрышком, центриолями, митохондриями, рибосомами, лизосомами. Они характерны, за некоторым исключением, как для клеток животных, так и клеток растений.
Ядро в клетке имеет форму и размеры, зависящие от формы, размеров и функций клетки. В клетках эукариотических организмов содержится, как правило, по одному ядру, реже по два и более. В зрелом состоянии эритроциты млекопитающих и клетки ситовидных трубочек покрытосеменных растений лишены ядер, тогда как клетки скелетных мышц позвоночных и млечных сосудов растений
являются многоядерными. Для инфузорий характерно наличие двух ядер - одно небольшое (микронуклеус) и одно крупное политенное (макронуклеус).
Ядро имеет округлую, палочковидную, четковидную вытянутую и другие формы. Размеры его колеблются от 2 до 100 мкм, а объем составляет около 65 мкм3. Особенно крупные ядра характерны для половых клеток (размером до 500 мкм). Отношение объема ядра к объему цитоплазмы называют ядерно-плазменным отношением, которое у клеток всех типов обычно постоянно.
Строение ядра характеризуется сложностью, хотя принципиально одинаково в клетках всех эукариотических организмов. В случае животных клеток ядро располагается в центре клетки. Напротив, в клетках растений оно имеет пристеночную локализацию. Однако положение, форма и размеры ядра могут меняться в зависимости от интенсивности метаболизма. В ядре содержатся хромосомы и ядрышко (см. ниже). Благодаря им ядро плотно заполнено ДНК, РНК и белками. Содержимое ядра иногда называют нуклеоплазмой или кариоплазмой. Она отделена от цитоплазмы ядерной мембраной, построенной из двух слоев (наружного и внутреннего) толщиной по 7 нм каждый и имеющей поры диаметром 40-100 нм. Поры занимают около 5% площади ядра. Через ядерные поры синтезируемая в ядре РНК выходит в цитоплазму, где она участвует в трансляции генетической информации (синтезе белков).
Ядерная мембрана и ядерные поры объединены с мембранной системой клетки, в результате чего клетка, по существу, канализиро- вана различными проходами, обеспечивающими движение веществ в клетке начиная с плазматической мембраны. В порах обнаружены глобулярные и фибриллярные белковые структуры. Количество пор зависит от вида клеток и обычно увеличивается в S-периоде. Между мембранными слоями имеется так называемое перинуклеарное про- странство шириной 20-60 нм. Наружная ядерная мембрана часто переходит в эндоплазматическую сеть. Когда клетки делятся, ядерная оболочка исчезает, а после деления восстанавливается. Ядерное вещество (кариоплазма) - плотный коллоид.
Важнейшая функция ядра заключается в том, что оно является центром управления в клетке.
Хромосомы располагаются в ядре. Их название введено в литерату- ру В. Вальдейлом в
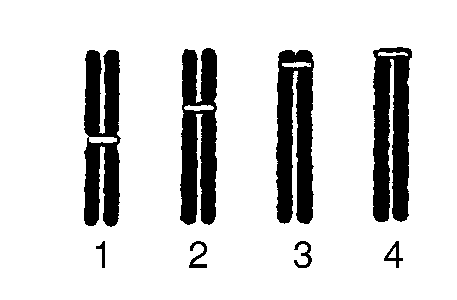
Рис. 4. Типы хромосом: 1 - метацентрические; 2 - субметацентрические; 3 - акроцентрические; 4 - телоцентрические
касающиеся длины и положения перетяжки (центромеры) по длине хромосомы. Каждая из митотических индивидуальных хромосом состоит из двух сестринских хроматид, удерживаемых центромерой. В зависимости от локализации центромер различают метацентрические, субметацентрические, акроцентрические и телоцентрические хромосомы (рис. 4). Количество хромосом постоянно в ядрах соматических клеток, где они находятся в парах. Диплоидный набор хромосом называется кариотипом (от греч. сагуоп - ядро, type - форма, тип). Для различных организмов характерны разные по количеству диплоидные наборы хромосом (табл. 3). В зависимости от строения хромосомы человека классифицируются на группы (рис. 5).
Таблица 3. Количество хромосом в соматических клетках отдельных организмов
Вид животных и растений | Количество хромосом (в диплоидном наборе) | |
Малярийный плазмодий | Plasmodmm malariae | 2 |
Плодовая мушка | Drosophila melanogaster | 8 |
Комнатная муха | Musca domestica | 12 |
Сазан | Cyrpinus carpio | 104 |
Лягушка зеленая | Rana esculenta | 26 |
Голубь | Columba livia | 80 |
Кролик | Lepus cunimlus | 44 |
Шимпанзе | Antropopitecus pan | 48 |
Человек | Homo sapiens | 46 |
Картофель | Solanum tuberosum | 48 |
Рожь | Secale cereale | 14 |
Пшеница (мягкая) | Tritium | 42 |
Кукуруза | Zea mays | 20 |
Когда ядра соматических метафазных клеток окрашивают основными красителями, то при микроскопии препаратов в ядерной зоне обнаруживают глыбки и гранулы окрашенного вещества, получившего название хроматина.
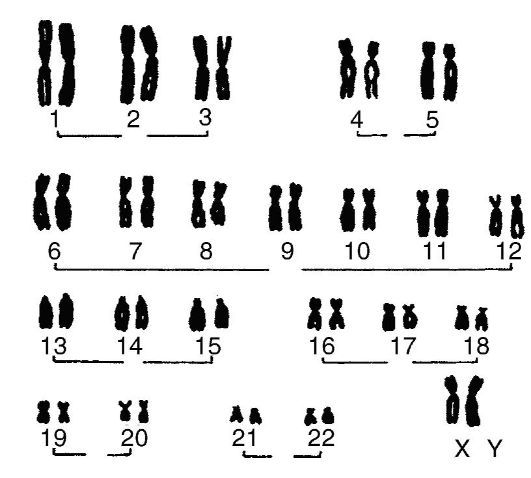
Рис. 5. Кариотип человека
Именно из этого вещества во время деления клеток организуются хромосомы. При изучении таких микроскопических препаратов отмечают, что одни районы хромосом окрашены очень интенсивно, другие слабее. Интенсивно окрашивающийся хроматин получил название гетерохроматина, менее окрашивающийся - эухроматина. Последний содержит области, на которых очень активно синтезируется РНК.
Исследование хромосом типа ламповых щеток, содержащихся в ооцитах многих животных, и политенных хромосом, обнаруживаемых в клетках насекомых, показало, что они имеют петельную структуру. Эта структура характерна, вероятно, для организации хроматина у всех организмов, включая человека.
В клетках многих видов организмов гетерохроматин концентрируется в основном в районах, фланкирующих центромеры.
Половые хромосомы часто почти полностью представлены гетерохроматином. Гетерохроматином богаты районы повторов последо- вательностей ДНК (см. гл. V).
Когда гетерохроматин исследуют под электронным микроскопом, то отмечают, что он построен из плотно упакованных хроматиновых нитей диаметром 25 нм. Эухроматин составлен из менее плотно упакованных нитей, но такого же диаметра. Гетерохроматин сохраняется
в высококонденсированной форме на протяжении всего клеточного деления, тогда как эухроматин менее конденсирован и невидим в интерфазе при исследовании хромосом в световом микроскопе. Имеющиеся данные показывают, что большинство исследованных генов эукариот локализовано в эухроматированных (менее конденсированных) районах хромосом, тогда как гетерохроматин (высококонденсированный) генетически не активен.
Химический состав хроматина довольно прост. Он состоит из ДНК (15%), белков (75%), а также некоторого количества РНК (10%). Различают хромосомные белки двух типов - основные белки (положительно заряженные при нейтральном рН), называемые гистонами, и гетерогенные белки, по большей части кислые (отрицательно заряженные при нейтральном рН), называемые негистоновыми. Молекулярная масса гистонов составляет 11 000-21 000. Гистоны являются основными белками по той причине, что содержат позитивно заряженные аминокислоты аргинин и лизин в количестве от 20 до 30% аминокислотных остатков белка. Группы R аргинина и лизина позволяют гистонам действовать в качестве поликатионов и взаимодействовать с ДНК, которая является полианионной благодаря ее отрицательно заряженным фосфатным группам. Другими словами, по причине протонирования боковых групп R-остатков аргинина и лизина гистоны, соединяясь с отрицательно заряженной двухцепочечной ДНК, образуют комплексы ДНК-гистоны.
Гистоны присутствуют в хроматине всех высших эукариотов в количествах, эквивалентных числу ДНК. У всех высших растений и животных они представлены пятью главными белками, обозначенными символами H1, Н2а, Н2Ь, Н3 и Н4. Количество ДНК и гистонов в хромосомах эквивалентно. Они присутствуют в клетках всех типов, кроме сперматозоидов некоторых организмов. В последних обнаруживают основные белки, называемые протаминами. Все пять гистонов присутствуют в молярных соотношениях 2Н1:2Н2а:2Н2Ь: 2Н3:2Н4:2. Для них характерны различия по молекулярной массе и аминокислотному составу. Гистон HI богат лизином (29%), гистоны Н2а и Н2Ь богаты как лизином, так и аргинином, а гистоны Н3 и Н4 - аргинином. Для гистонов Н3 и Н4 характерно сходство последовательностей аминокислот; для других гистонов заметное сходство отсутствует. Данные о константности гистонов H1, Н2а, Н2Ь, Н3 и Н4 в клетках организмов разных видов позволяют предположить, что гистоны имеют значение в упаковке ДНК. Кроме того,
они неспецифически вовлечены в регуляцию экспрессии генов (см. гл. VII).
Негистоновая белковая фракция хроматина состоит из различных очень гетерогенных белков. Состав этой фракции широко варьируется в клетках разных организмов даже одного и того же вида. Предполагают, что негистоновые белки также принимают участие в регуляции экспрессии специфических генов.
Каждая хроматида содержит
одиночную гигантскую двухцепочечную молекулу ДНК, вытянутую по всей
длине хромосомы. Это заключение основано на данных о размерах самых
больших молекул ДНК в ядрах клеток дрозофилы и других организмов, а
также на данных об общем количестве ДНК, присутствующей в самых больших
хромосомах дрозофилы. Наиболее крупные молекулы ДНК, выделенные из
хромосом, имеют контурную длину порядка
Количество ДНК и гистонов в хроматине эквивалентно. Будучи специфически связанными с ДНК, гистоны принимают участие в формировании основных структурных субъединиц хроматина (хромосом) - нуклеосом, которые являются элементарными единицами организации хроматина. Каждая нуклеосома представлена сегментом ДНК диаметром около 30 нм и длиной 146-240 пар оснований, намотанной 1,8 раза вокруг гистонового стержня, состоящего из пары гистонов. Нуклеосома закручена в сложную нить 2 раза при участии гистона HI в качестве кросслинкера, что дает структуру с упаковочным отношением 25:1, близким к интерфазному хроматину. Чтобы сформировалась митотическая хромосома из нити длиной 30 нм, необходима дальнейшая компактизация с помощью специфических негистоновых белков, приводящая к формированию скелета хромосомы. Установлено, что нуклеосомы являются репрессорами инициации транскрипции в эукариотических клетках. Нарушение структуры нуклеосом ведет к освобождению транскрипции от репрессии.
В метафазных хромосомах ДНК организуется в петли длиной 30 000-40 000 пар оснований, а каждая петля хроматина прикрепля- ется к гистоновому скелету хромосомы на его основании в результате взаимодействия ДНК + белок или ДНК + РНК.
Гистон HI связан с линкерной ДНК, принимая участие в стабилизации сверхскрученных ДНК между нуклеосомами, но способ этой связи полностью еще не выяснен. Длина сегментов ДНК (линкеров), которые соединяют нуклеосомы, составляет от 15 до 100 нуклеотидных пар в зависимости от типа клетки. Результаты Х-дифракции кристаллов изолированных нуклеосом указывают на то, что они имеют дисковидную структуру и состоят из двух симметричных половин. ДНК каждой половины намотана в форме суперспирали на поверхность гистоновых стержней.
Для хромосом характерно наличие отдельных сайтов, которые определяют их хрупкость, что создает условия для нарушений структуры хромосом, сопровождающихся хромосомными мутациями (см. гл. VII).
Ядрышко - это очень мелкая структура диаметром 1-5 мкм, кото- рая локализуется в ядре. Количество ядрышек бывает разным - от одного до нескольких. Центральная часть в ядрышке представлена фибриллярной частью (цепи ДНК, ядрышковые организаторы и рибонуклеопротеиды) и гранулярной (формирующиеся субъединицы рибосом). Ядрышко - это место синтеза мрРНК. Матрицей для синтеза рРНК является ДНК ядрышкового организатора. Синтезируемая рРНК объединяется с белком, и образующиеся рибонуклеопротеиды служат затем материалом для сборки из них субъединиц рибосом. Последние через ядерные поры направляются в цитоплазму, участвуя там в формировании рибосом, на которых происходит затем синтез белков.
Центриоли представляют собой гранулярные цилиндры диаметром около 0,15 мкм и длиной 0,5 мкм, локализирующиеся парами в клетке около ядра. Эти пары называют диплосомами (центросомами). Структурно они представляют систему мембранных трубочек. Диплосома окружена другими микротрубочками, называемыми центросферой. Обе эти структуры образуют клеточный центр.
Функцией клеточного центра является организация цитоскелета клеток. Кроме того, центриоли участвуют в делении клеток, расходясь к полюсам клетки. Микротрубочки, протягивающиеся взаимно, образуют митотическое веретено. В клетках растений этой органеллы нет.
Митохондрии присутствуют во всех клетках организмов, которые используют для дыхания кислород. В одной клетке может находиться 50-5000 митохондрий. Они имеют форму палочек, нитей или гранул, а их размеры достигают 7 мкм (рис. 6).
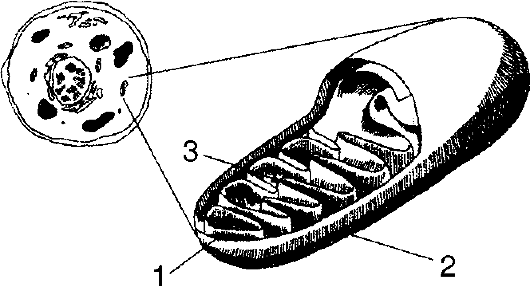
Рис. 6. Митохондрия: 1 - внутренняя мембрана; 2 - внешняя мембрана; 3 - криста
Митохондрии образованы двумя мембранами - наружной и внутренней, между которыми образуется пространство шириной 10-20 нм. Внутренняя мембрана формирует складки (кристы), которые погружены в матрикс, представляющий собой молекулярное содержимое митохондрии. Наружная мембрана проницаема для низкомолекулярных соединений. Проникновение веществ во внутреннее пространство (матрикс) митохондрий контролируется внутренней мембраной. Содержимое митохондрий составляют белки, фосфолипиды, ДНК, РНК и рибосомы, которых значительно меньше, чем у рибосом, локализованных в цитоплазме. Наличие этих структур обеспечивает собственную митохондриальную белоксинтезирующую систему. Митохондрии способны к самовоспроизведению путем деления или почкования в период S-фазы и других фаз клеточного цикла. Митохондрии имеются также у простейших. В частности, трипаносомы содержат по одной митохондрии.
Митохондрии представляют собой «силовые станции», в которых происходят основные процессы получения и накопления энергии путем окисления молекул пищи (окислительное фосфорилирование) для образования АТФ. В матриксе содержатся ферменты, а реакции окисления проходят на поверхности внутренней мембраны. Энергетические потребности клеток определяют уровень размножения митохондрий. Биохимические реакции в митохондриях пред- ставляют собой «митохондриальную дыхательную цепь», контролируемую более 80 генами.
В клетках растений вместо митохондрий имеются пластиды. Среди пластид различают хлоропласты, которые содержат хлорофилл, лей- копласты (бесцветные пластиды), в которых происходит накопление
крахмала, и хромопласты, в которых осуществляется синтез пигмента плодов. У пурпурных бактерий имеются хроматофоры.
Для хлоропластов характерно мембранное строение. Встречаясь в клетках в количестве около 40 экземпляров на клетку, они имеют своеобразную форму двояковыпуклых линз и размеры 5-10 мкм. Каждый хлоропласт окружен двойной мембраной. Наружная мембрана гладкая, состоит из белковых субъединиц, а внутренняя - складчатая. Внутренняя мембрана содержит хлорофилл, а также ферменты, синтезирующие АТФ и органические соединения с помощью АТФ. Кроме того, в хлоропластах имеется некоторое количество ДНК и небольших по размерам рибосом, что обеспечивает собственную хлоропластную белоксинтезирующую систему.
Для пластид характерны переходы из одного типа в другой. Например, зеленые хлоропласты листьев переходят в желтые хло- ропласты (осенью, когда изменяется окраска листьев), а лейкопласты зеленеющих клубней картофеля переходят в хлоропласты.
Рибосомы представляют собой гранулы, расположенные в цито- плазматическом матриксе и связанные с мембранами цитоплазматической сети. Их размеры составляют 15-35 нм в диаметре. Они построены на 1/3 из белка и на 2/3 из РНК (двух субъединиц). Количество рибосом в клетках очень большое. Например, в одной бактериальной клетке (кишечной палочке) их содержится около 6000 экземпляров. Рибосомы образуют группы, называемые полирибосомами.
Функции рибосом заключаются в том, что на них осуществляется синтез белков.
Лизосомы представляют собой мельчайшие пузырьковидные обра- зования, окруженные однослойной мембраной и содержащие ферменты. Их размеры составляют 0,2-0,8 мкм. Они обеспечивают изоляцию гидролитических ферментов.
В лизосомах обнаружено более 40 различных гидролитических ферментов (протеиназ, нуклеаз, липаз, кислой фосфатазы, гликоидаз и др.), осуществляющих внутриклеточное расщепление макромолекул, проникающих в клетки посредством фагоцитоза и пиноцитоза. Лизосомы чрезвычайно разнообразны по строению и частным функциям. Предполагают, что они образованы мембранами комплекса Гольджи и представляют собой систему удаления из клеток конечных продуктов обмена. Повреждение лизосом сопровождается растворением клеток. Следовательно, благодаря лизосомам происходит
защита клеток от собственных ферментов. В эволюционном плане они являются аналогами пищеварительных вакуолей одноклеточных организмов.
Пероксисомы (иногда их называют микротельцами) представляют собой образованные цистернами эндоплазматической сети пузырьки размером 0,3-1,5 мкм, ограниченные однослойной мембраной. Эти пузырьки содержат каталазу и некоторые оксидазы. Обильными по содержанию пероксисом являются эпителии печени и почек. Функция пероксисом заключается в том, что в них происходит раз- рушение клеточных перекисей и холестерина. Кроме того, они содержатся в зеленых листьях растений. Известны также глиоксисомы, содержащиеся в проростках семян масличных растений.
Клеточные включения. В клетках животных включения представлены жировыми каплями, гранулами гликогена, зимогеновыми гранулами (депо ферментов в клетках поджелудочной железы). Особенно богаты жиром у млекопитающих клетки соединительной ткани. Гликогена много в клетках поперечно-полосатых мышц, печени и в нейронах. Встречаются также белковые включения (в цитоплазме яйцеклеток, печени, в теле простейших).
Следует отметить также, что некоторые клетки животных содержат в качестве включений пигменты. Например, в клетках коркового вещества надпочечников накапливаются липохромы. Пигментами являются гемоглобин и меланин, а также ретинин (в зрительном пурпуре сетчатки глаза).
В клетках растений, например, таких как картофель, злаковые, наиболее частым включением является крахмал. В качестве включений в клетках растений встречаются также жиры, содержание которых очень большое в семенах масличных культур. Очень широко распространены пигменты.
Участки (компартменты) цитоплазматического матрикса, лишенные мембран в клеточных органеллах, получили название цитозоля.
ХИМИЧЕСКИЙ СОСТАВ
Клеточное вещество - сложный полифазный коллоид, т.е. представляет собой систему из двух несмешивающихся фаз. Одна из этих фаз структурно является цитоплазматическим матриксом и выполняет роль водной фазы с переходами от жидкого до твердого состояния, тогда как другая является мембранной системой и выполняет
роль относительно жидкой фазы. Цитоплазма практически бесцветна, имеет характер раствора.
В элементном составе клетки насчитывают более 70 элементов, среди которых наиболее частыми являются кислород, углерод, водо- род, азот. На долю кислорода приходится 65% общей массы, на долю углерода - 18%, водорода - 10%, азота - 3%. После этих элементов идут кальций, фосфор, калий, сера, натрий, хлор. Поскольку все эти элементы встречаются в клетках в большом количестве, часто их называют макроэлементами. Марганец, медь, йод, кобальт и другие, обнаруживаемые в микроколичествах, называют микроэлемен- тами.
Химические элементы, входящие в состав клеток и обладающие биологическими функциями, называют биогенными.
Как правило, содержание катионов и анионов отличается от содержания их в той среде, в которой находятся клетки. Например, концентрация К+ в мышечных клетках в несколько десятков раз выше, чем в крови. Концентрация солей в клетках определяет буферность ее содержимого, под которой понимают уровень концентрации водородных ионов в клетках (рН).
Соединяясь химическими связями, группы атомов образуют молекулы химических соединений.
В клетках человека химические элементы имеют следующие значения (в % к сухой массе):
Кислород - 65 Марганец - 0,0003
Углерод - 18 Медь - 0,0002
Водород - 10 Йод - 0,0004
Азот - 3 Кобальт - следы
Фосфор - 1 Цинк - следы
Кальций - 1,5 Молибден - следы
Калий - 0,35 Никель - следы
Сера - 0,25 Алюминий - следы
Натрий - 0,15 Барий - следы
Хлор - 0,15 Стронций - следы
Магний - 0,05 Титан - следы
Железо - 0,004 Литий и др. - следы
Клетки построены из неорганических и органических соединений. Важнейшим неорганическим соединением клетки является вода, которая составляет около 70% ее массы. У отдельных организмов, например медуз, содержание воды превышает 95%. Вода имеет
исключительно важное значение для жизнедеятельности клеток, представляя собой среду, в которой осуществляются важнейшие реакции, лежащие в основе синтеза и распада веществ.
Кроме того, она является растворителем различных химических веществ, принимает участие в выведении из клеток продуктов обмена. Наконец, она поддерживает тепловой режим клетки.
Другие неорганические соединения, встречающиеся в клетках, представлены минеральными солями. Вместе с белками минеральные соли входят в состав протоплазмы. Встречаются калиевые, натриевые, магниевые соли, соли серной, соляной, фосфорной и других кислот. Важнейшая роль минеральных солей заключается в определении ими кислотно-щелочного состояния протоплазмы. Они необходимы также для размножения клеток.
Органические (углеродсодержащие) соединения клетки - углеводы, липиды, белки, нуклеиновые кислоты, АТФ и стероиды. Как было отмечено, молекулы этих соединений часто называют биологическими молекулами.
Углеводы являются соединениями углерода, водорода и кислорода. Различают полисахариды (СбН10О5), дисахариды (С12Н22О11) и простые сахара - моносахариды (СбН12Об). Две молекулы моносахарида продуцируют одну молекулу дисахарида и одну молекулу воды, тогда как из п молекул моносахарида продуцируют одну молекулу полисахарида и п-1 молекул воды. Следовательно,
2С6Н12О6 С12Н22О11 + Н2О
тогда как
^(C6H12U6) -> (С6Н10О5) + (п - 1) Н2О.
Основные
углеводы протоплазмы - глюкоза, гликоген (в клетках животных) и
крахмал (в клетках растений). Углеводы входят в состав мембранной
системы клеток. Они являются также важнейшим источником энергии в
организме, которая освобождается в результате
окислительно-восстановительных реакций. Установлено, что окисление
Липиды - соединения, состоящие из жирных кислот и спиртов. В частности, одна молекула глицерина (трехатомного спирта) и три молекулы жирной кислоты образуют одну молекулу липида и три молекулы воды. Например, стеарин образуется в результате реакции
между одной молекулой глицерола и тремя молекулами стеариновой кислоты:
С3Н5(ОН)3 + 3С17Н35СООН -> (С17Н35СОО)3С3Н5 + 3Н2О.
Глицерол Стеариновая кислота Стеарин Вода
Липиды
в соединении с белками входят в состав мембранной системы клеток,
обнаруживаются в нервной ткани, мужских половых клетках, в семенах
растений. Поскольку углеводы могут переводиться в липиды, то последние
выполняют роль накопителей энергии. Окисление липидов сопровождается
выделением энергии. Например, окисление
Белки - наиболее сложные химические соединения, характеризующиеся большой молекулярной массой. Белки построены из аминокислот, которые содержат аминогруппу (-NH2) и карбоксильную группу (-СООН). Молекула белка представляет собой, по существу, L-аминокислоты, соединенные пептидными связями в неопределенно длинные цепи:
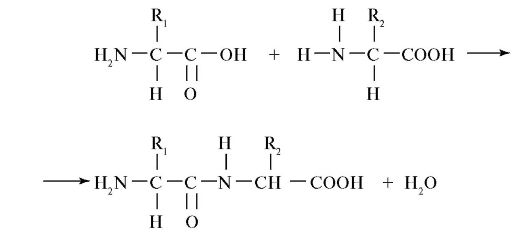
где R является радикалом, состоящим из гидроксильной (ОН), сульфгидрильной (SH) и других групп, и частью молекулы.
Известно 20 различных L-аминокислот. Они различаются между собой боковыми группами, прикрепленными к альфа-углероду.
На долю белков в протоплазме приходится 50-70% от общего количества органических соединений. Белки являются прежде всего строительным материалом. Они входят в состав практически всех клеточных структур. Белки являются также катализаторами
(ферментами) всех реакций и регуляторами экспрессии генов. Они являются гормонами, токсинами, антителами. Теряя аминогруппы (дезаминируясь), белки могут стать также источником энергии.
Нуклеиновые кислоты - органические соединения, содержащие углерод, кислород, водород, азот и фосфор. Различают дезоксирибонуклеиновую и рибонуклеиновую кислоты (ДНК и РНК). Важнейшая биологическая роль нуклеиновых кислот заключается в том, что они служат хранителями генетической информации.
Стероиды представляют собой органические соединения, содержащие углерод, водород и кислород. Их биологическая роль состоит в том, что они являются гормонами.
РАЗМНОЖЕНИЕ КЛЕТОК
Размножение, или пролиферация (от лат. proles - потомство, ferre - нести), клеток - это процесс, который приводит к росту и обновлению клеток. Данный процесс характерен как для одноклеточных, так и многоклеточных организмов.
Клетки-организмы (одноклеточные организмы) размножаются простым делением надвое (бактерии, саркодовые), множественным делением (споровики и др.) или другим путем. Поэтому у бактерий и одноклеточных животных удвоение клеток представляет собой размножение их как самостоятельных организмов, поскольку из исходной формы образуются две новые клетки, каждая из которых является организмом. Каждая дочерняя клетка или клетка-организм получает полную генетическую информацию, несомую исходной клеткойорганизмом.
Соматические клетки многоклеточных организмов размножаются путем сложного деления, которое получило название митотического деления (митоза) и представляет собой механизм, посредством которого одиночные клетки репродуцируют себя, в результате чего происходит рост организма. Образовавшиеся в результате деления дочерние клетки подобны исходной (материнской) клетке, отличаясь от последней лишь меньшими размерами. Однако вслед за делением дочерние клетки растут, быстро достигая размеров материнской клетки. Из-за точности митотического деления клеток митоз является скорее удвоением клеток, чем делением.
Биологический смысл митотического деления состоит в том, что он является ключевым событием в точной репликации всех хромосом
еще до того, как произойдет деление ядра и клетки. В результате митоза дочерние клетки после деления получают хромосомы в точно таком же количестве, какое имела их родительская клетка. Следовательно, митотическое деление есть особый способ упорядоченного деления клеток, при котором каждая из двух дочерних клеток получает в точности такое же число и те же типы хромосом, какие имела материнская клетка. При каждом митозе образуется копия каждой хромосомы, причем механизм их распределения между дочерними клетками очень точен.
В митотическом делении клетки различают две стороны - разделение ядра на два дочерних ядра (равное деление хромосом), называемое кариокинезом (митозом), и следующее затем разделение цитоплазмы с образованием двух дочерних клеток, называемое цитокинезом. Каждая из дочерних клеток содержит одно дочернее ядро. Кариокинез и цитокинез синхронны.
Существенной особенностью митотического деления является то, что оно в значительной мере сходно у всех организмов. Совокупность процессов, происходящих в клетке от одного деления до другого, получило название митотического цикла. Последний состоит из двух стадий - стадии покоя, или интерфазы, и стадии деления, или митоза (от греч. mitos - нить).
Интерфаза предшествует митозу, и в ней происходит синтез ДНК (рис. 7). Различают три периода интерфазы - пресинтетический, синтетический и постсинтетический. Пресинтетический период (G1), который называют еще первым интервалом (от англ. gap - интервал), является начальным периодом интерфазы. В этот период ДНК еще не синтезируется, однако происходит накопление РНК и белка. Обычно этот период длится 12-24 ч. Синтетический период (S) следует за 01-периодом и характеризуется тем, что в этот период в клетке происходит синтез ДНК, в результате чего количество последней удваивается. В этот период продолжается также синтез РНК и белков. Следовательно, наиболее фундаментальной особенностью S-периода является репликация генов.
К концу этого периода каждая из хромосом состоит из двух хроматид, т. е. удваивается набор генов каждой дуплицированной хромосомы. Длительность S-периода обычно составляет 5 ч. Постсинтетический период (G2) характеризуется остановкой синтеза ДНК и накоплением энергии. Продолжается также синтез РНК и белков.
Митоз сопровождается на протяжении четырех последовательных фаз - профазы, метафазы, анафазы и телофазы (рис. 8).
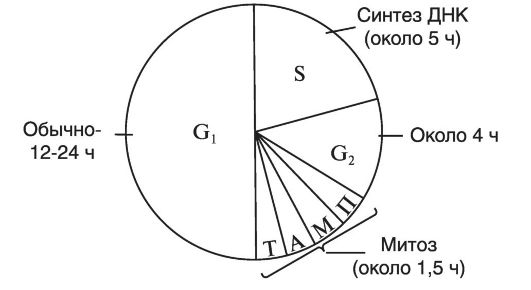
Рис. 7. Митотический цикл: П - профаза; М - метафаза; А - анафаза; Т - телофаза; (G1 + S +G2) - интерфаза
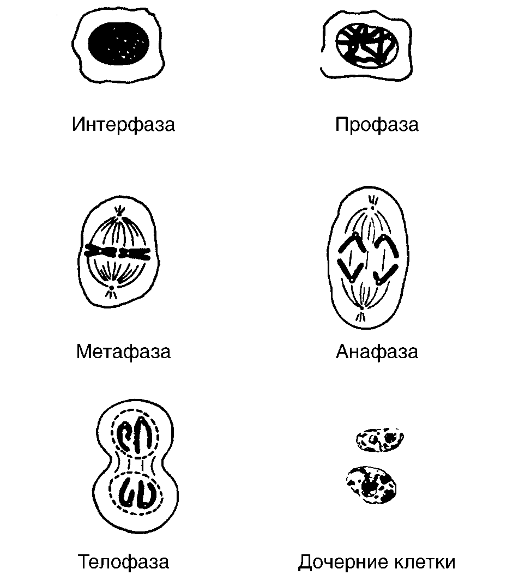
Рис. 8. Митоз
В профазе вначале происходит конденсация и спирализация (скручивание) хромосом, в результате чего они становятся видимыми при микроскопии окрашенных препаратов. Увеличивается диаметр каждого завитка. Ядерная мембрана растворяется под действием ферментов, ядрышко исчезает. Центросома делится на две центриоли, после чего последние расходятся к полюсам клетки. Затем между полюсами начинает формироваться ахроматиновая фигура, похожая на веретено, которое к концу этой фазы вытягивается вдоль клетки. Оно состоит из белка и РНК. К концу профазы каждая хромосома удваивается, в результате чего образуются две хроматиды (сестринские хроматиды), которые удерживаются вместе центромерой.
В метафазе хромосомы располагаются на экваторе веретена. Они имеют вид толстых образований, плотно свернутых спиралью, что облегчает подсчет и изучение их структуры. Будучи прикреплен- ными к нитям веретена центромерой, хроматиды пока удерживаются вместе, но плечи их уже разъединены.
В анафазе наступает продольное разделение хромосом, в результате чего каждая сестринская хроматида имеет собственную центромеру и становится дочерней хромосомой. Хромосомы удлиняются и двигаются к соответствующим полюсам веретена.
В телофазе дочерние хромосомы достигают полюсов, вытягиваются и деспирализуются. Образуется ядерная оболочка, вновь появляется ядрышко. Длительность телофазы составляет 20-30 мин.
На заключительном этапе клеточного деления происходит цитокинез. Этот процесс заканчивается образованием в экваториальной зоне клетки перетяжки, которая разделяет делящуюся клетку на две дочерние клетки. В отличие от соматических клеток животных, в клетках растений из-за ригидности их стенок формируется пластинка между будущими дочерними клетками. На каждой из сторон этой пластинки откладывается целлюлоза. Каждое клеточное деление является непрерывным процессом, поскольку ядерные и цито- плазматические фазы, вопреки различиям в содержании и по значению, координированы во времени.
Упорядоченность клеточных делений у эукариотов зависит от координации событий в клеточном цикле. У эукариот эта координация осуществляется путем регуляции трех переходных периодов в клеточном цикле, а именно: вступление в митоз, выход из митоза и прохождение через пункт S, называемый «Старт», который вводит инициацию синтеза ДНК (S-фазу) в клетке.
Временной порядок в клеточном цикле эукариотических клеток определяется семейством циклинзависимых киназ, действующих во взаимодействии с их регуляторными субъединицами - циклинами.
Продолжительность митотических циклов клеток различна и составляет от нескольких часов до нескольких дней. Однако она зависит от типа тканей, физиологического состояния и внешних факторов (температура, свет).
Существуют и другие регуляторы клеточного цикла. В частности, известны ингибиторы семейства циклинзависимых киназ. Такими ингибиторами являются белки р21, р16 и р27. Они ингибируют функции киназ путем связывания с ними.
Разные ткани характеризуются различной митотической активностью. Поэтому в зависимости от митотической активности различают стабильные, растущие и обновляющиеся ткани. Стабильные ткани - в них клетки не делятся, а количество клеточной ДНК постоянно. Например, клетки центральной и периферической нервной системы не делятся. В этих клетках происходят лишь возрастные изменения. Растущие ткани - в них клетки живут всю жизнь, но среди последних имеются такие, которые делятся посредством митоза. В результате этого наступает увеличение размеров органов. Примером растущих тканей являются ткани почек, желез внутренней секреции, скелетная и сердечная мускулатура. Обновляющиеся ткани - в них многие клетки подвержены митозам, в результате чего погибающие клетки компенсируются вновь образующимися. Примерами обновляющихся тканей являются клетки желудочно-кишечного, дыхательного и мочеполового трактов, эпидермиса, костного мозга, семенников и др. Для митозов характерны суточные колебания, волны.
У высших организмов митотическое деление клеток обеспечивает их рост с последующим увеличением массы тела и дифференциацией клеток. По мере индивидуального развития человека количество его клеток увеличивается, достигая у взрослого более чем 1015 клеток и оставаясь затем константным.
Большинство клеток в организме со временем стареет, в результате чего в них накапливаются липиды, кальций, пигмент «изнашивания» и они гибнут. Подсчитано, что организм взрослого человека ежедневно теряет около 1-2% своих клеток в результате их гибели. Например, клетки печени живут около 18 месяцев, эритроциты - 4 месяца. Однако погибающие клетки замещаются новыми. Считают,
что клеточное содержание организма человека обновляется примерно каждые 7 лет. Особенно сильно замещение клеток идет в ткани крови за счет интенсивного образования их в кроветворных тканях. Применительно к другим видам клеток процесс обновления происходит с очень быстрой скоростью. Например, эпителий желудка и кишечника крыс обновляется каждые 72 и38 ч соответственно, эпителий тонкого кишечника человека - каждые 7-8 дней. Однако нервные клетки функционируют на протяжении всей жизни орга- низма.
После смерти клетки в ней происходят коагуляция протоплазмы, распад митохондрий и других органелл в результате аутолиза (активации внутриклеточных ферментов).
Наряду с делением клеток путем митоза известен амитоз (от греч. а - отрицание, mites - деление ядра), под которым понимают прямое деление ядра клетки. При амитозе сохраняются интерфазное состояние ядра, ядрышко, ядерная мембрана. Ядро клетки делится на две части без формирования веретена, в результате чего образуется двуядерная клетка. Амитоз встречается в клетках скелетной мускулатуры, кожного эпителия, соединительной ткани. Однако считают, что амитоз является редким и аномальным механизмом в размножении клеток.
ЭВОЛЮЦИЯ КЛЕТОК
Предполагают, что предковой клеточной формой была примитивная прокариотическая клетка, возникшая около 3,5-109 лет назад. Клетки этого типа для обеспечения своего существования и раз- множения вначале использовали органические молекулы небиологического происхождения. Однако в последующем у примитивных прокариотических клеток стали развиваться механизмы синтеза и энергетического обеспечения. Первые прокариотические клетки обладали наиболее простыми каталитическими системами, в результате чего обеспечение их энергией основывалось на брожении. В последующем отдельные виды прокариотических клеток переключились с брожения на дыхание, что способствовало более эффективному получению энергии. Таким образом, эволюционные изменения прокариотических клеток шли по линии развития у них различных метаболических путей. Их геном развивался в направлении формирования «голых» молекул ДНК.
Эволюционные изменения эукариотических клеток шли в направлении увеличивающегося разнообразия в форме, размерах, структуре и функциях с одновременной компартментализацией биохимических систем и сохранением общего для всех клеток аэробного метаболизма. Считают, что эукариотические клетки возникли менее 1 млрд лет назад из прокариотических клеток, причем для объяснения их происхождения выдвинуты четыре гипотезы.
В соответствии с одной из этих гипотез (гипотезой клеточного симбиоза), которая сейчас является наиболее распространенной, предполагают, что эукариотическая клетка - это симбиотическая структура, состоящая из нескольких клеток разных типов, объединенных общей клеточной мембраной. В частности, предполагают, что пластиды клеток современных зеленых растений происходят из водорослей, бывших предками современных синезеленых водорослей и способных к аэробному фотосинтезу, а митохондрии эукариотических клеток ведут начало от аэробных бактерий, которые вступили в симбиоз с анаэробными клетками, способными к фотосинтезу, что привело к образованию клетки, которая могла существовать в атмосфере кислорода, и использованию кислорода путем дыхания. Относительно ядра есть мнение, что оно является рудиментом также какого-то древнего внутриклеточного симбионта, утратившего после включения в исходную клетку свою цитоплазму. В пользу этой гипотезы свидетельствуют данные о симбиотических взаимоотношениях некоторых современных организмов. Например, одноклеточная зеленая водоросль хлорелла (Chlorella) обитает в цитоплазме зеленой парамеции (Paramechim bursaria). Из-за способности к фотосинтезу она снабжает парамеции питательными веществами. Далее, пластиды и митохондрии содержат собственную систему генетической информации о синтезе белков в виде ДНК, мРНК, рРНК, тРНК и соответствующих ферментов. Для хлоропластов, митохондрий и клеток-прокариот характерно сходство способов репродукции (все они одинаково репродуцируются путем простого деления надвое). Наконец, мутации митохондриальных генов независимы от мутаций ядерных генов.
В соответствии с другой гипотезой считают, что эукариотическая клетка произошла от прокариотической клетки, содержавшей несколько генов, прикрепленных к клеточной мембране. В результате инвагинаций клеточной мембраны образовались мезосомы, способные первоначально к фотосинтезу. Однако в дальнейшем произошла
специализация этих органелл, в результате чего одна из них, утратив дыхательную и фотосинтетическую функции, развилась в ядро, другие, наоборот, развив эти функции, стали митохондриями у животных и пластидами у растений. В пользу этой гипотезы свидетельствуют данные о двойном строении мембран ядра, митохондрий и пластид.
В соответствии с третьей гипотезой, основанной на мысли о том, что все живые формы произошли от предковых анаэробных ферментативных гетеротрофов, эукариоты представляют собой сублинию безстеночных (анаэробных) прокариотов, которые развили способность к эндоцитозу. Посредством «заглатывания» других прокариотов, которые дали им дополнительные метаболические способности и которые в конце концов дегенерировали в органеллы, примитивная клетка (эукариот) стала эукариотической клеткой. Таким образом, прокариоты древнее, проще и примитивнее клеток-эукариот.
В соответствии с четвертой гипотезой предполагают, что эукариотические клетки возникли из прокариотической клетки, содержавшей много геномов, которые распадались на части, давшие начало структурам с разными функциями. В последующем шло клонирование структур со сходными функциями, после чего они покрывались двойными мембранами, что привело к образованию ядра, митохондрий, а позднее и мембранной сети. В пользу этой гипотезы свидетельствуют данные о сходстве генетического кода, содержащегося в ядерной и митохондриальной ДНК, а также о сходстве в регуляции дыхательной функции ядром и митохондриями.
Геном эукариотических клеток впоследствии развивался в направлении объединения молекул ДНК с белками и формирования хроматина и хромосом разной формы и в разном количестве. Специализация хроматина проявилась в формировании эухроматина и гетерохроматина и в формировании аутосом и половых хромосом. Что касается количества хромосом, то объяснить их эволюционную тенденцию пока трудно, поскольку многие примитивные организмы содержат в своих клетках большее число хромосом, чем организмы, занимающие высшие эволюционные ступени. Однако несомненно, что количественные и структурные изменения кариотипов в течение эволюции играли важную роль в видообразовании. Параллельно с этим происходило усложнение структур и функций клеточных компонентов, развитие регуляторных механизмов.
Несомненно эволюционное значение митоза. Считают, что точность разделения и распределения хромосом в результате мито-
за является условием, обеспечивающим многоклеточность. Однако происхождение самого митоза не имеет достаточных объяснений. Предполагают лишь, что он развился из примитивного митоза, представлявшего собой механизм, при котором расхождение реплицировавшихся хромосом происходило после вытягивания и разрыва веретена без разрушения ядерной мембраны.
У многих организмов характерен апоптоз, под которым понимают генетическую программу, по которой клетки совершают суицид. У бактерий апоптоз контролируется плазмидами и ядром, у многоклеточных организмов - белками ряда генов. Апоптоз - это эволюционно сохраняемый процесс.
ВОПРОСЫ ДЛЯ ОБСУЖДЕНИЯ
1. Сформулируйте основные положения клеточной теории. Как вы считаете, какова роль этой теории в биологии и медицине?
2. В чем заключаются доказательства того, что клетка является элементарной единицей жизни?
3. Назовите два процесса, которые являются общими для всех живых организмов.
4. Назовите и охарактеризуйте компоненты мембранной системы клетки.
5. Охарактеризуйте цитоплазматический матрикс и клеточные органеллы.
6. В чем заключается роль клеточного ядра?
7. В чем заключается роль митохондрий?
8. Сформулируйте определения клеточного цикла и митоза.
9. Что собой представляют лизосомы и какова их роль?
10. Каково значение ферментов в жизни клеток?
11. Каковы фазы митоза и сущность процессов, происходящих в этих фазах?
12. Какова форма хромосом в метафазе и чем она определяется?
13. В какой фазе происходит разделение центромеры и освобождение сестринских хроматид?
14. Определите, какая весовая часть ядра приходится на хроматин клетки (примерно), допуская, что диаметр ядра составляет 5 мкм, а плотность 1,1 г/см-3.
15. Считая, что хромосомы человека состоят на 15% из ДНК, определите массу всех хромосом его диплоидных клеток.
16. Что вы можете сказать о происхождении митоза?
17. Назовите принципиальные различия между клетками-прокариотами и клетками-эукариотами.
18. Что вы знаете об элементном составе клеток?
19. Что понимают под биологическими молекулами?
20. Какой представляется вам структура белков и что вы знаете о функциях белков?
21. Как вы понимаете происхождение клеток-прокариот?
22. Как вы понимаете происхождение клеток-эукариот?
23. Каково ваше мнение относительно развития генома эукариотических клеток?
24. Что вы знаете об амитозе?
25. Каковы причины гибели клеток? Что такое апоптоз?
26. Какие вы знаете методы изучения клеток?