Дерматовенерология : учебник для медицинских вузов / А. В. Самцов, В. В. Барбинов.-СПб. : СпецЛит, 2008.-352 с. : ил.
|
|
|
|
Глава 1. АНАТОМИЯ И ГИСТОЛОГИЯ КОЖИ
Кожа представляет собой крупнейший целостный многофункциональный орган, взаимосвязанный со всеми другими органами и системами организма. Непосредственно соприкасаясь с внешней средой, он выполняет барьерно-защитную функцию. Общая площадь поверхности кожи взрослого человека составляет 1,5-1,8 м2, а ее масса вместе с подкожной жировой клетчаткой достигает 17 % массы тела. В области носа, рта, мочеполовых органов и ануса кожа непосредственно переходит в слизистые оболочки. На поверхности кожи наблюдается сложный рисунок в виде треугольных и ромбических полей, сформированный многочисленными бороздками. Более грубые бороздки образуют складки в области ладоней, подошв, мошонки, а также морщины на лице. На ладонных и подошвенных поверхностях пальцев бороздки расположены параллельно и демонстрируют причудливые фигуры, индивидуальные для каждого человека. Эта их особенность используется в криминалистике для идентификации личности (дактилоскопия). Цвет кожи имеет матовый оттенок и обусловлен окраской основных ее тканей, просвечиванием кровеносных сосудов и содержанием пигмента меланина. В зависимости от соотношения этих компонентов у лиц различных рас он варьирует от светло-розового до темно-коричневого. Большая часть кожи покрыта волосами. Исключение составляют ладони, подошвы, боковые, ладонные и подошвенные поверхности пальцев, а также слизистые оболочки губ, ануса и мочеполовых органов. Также на поверхности кожи отмечаются едва заметные отверстия - поры сальных и потовых желез, расположенных в глубине органа. Сальные железы
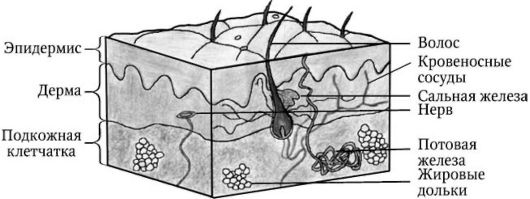
Рис. 1. Общее строение кожи
отсутствуют в области ладоней и подошв, а потовые - в области слизистой оболочки мочеполовых органов и кожи губ. На тыльных поверхностях дистальных отделов фаланг пальцев кистей и стоп расположены ногти.
Кожа состоит из трех компонентов, или отделов (рис. 1):
• эпидермиса (epidermis);
• дермы (dermis);
• подкожной жировой клетчатки (subcutis), или гиподермы (hypodermis).
Эпидемис является эпителиальной частью кожи, а дерма и гиподерма - соединительнотканной. Толщина кожи (без подкожной жировой клетчатки) колеблется от 0,8 мм в области век до 4-4,5 мм в области ладоней и подошв. Толщина гиподермы варьирует еще более - от полного ее отсутствия на отдельных участках, до 4-6 см в области живота у полных людей. Пограничная зона между эпидермисом и дермой имеет вид волнистой линии вследствие наличия в дерме выростов - сосочков, обусловливающих образование на поверхности кожи гребешков и борозд, формирующих кожный рисунок. В соединительнотканной части кожи (дерме и гиподерме) располагаются нервы, кровеносные и лимфатические сосуды, мышцы. Кроме того, кожа имеет собственные придаточные образования, к которым относятся волосы, сальные и потовые железы, а также ногти. Несмотря на внешнюю простоту, кожа отличается сложным строением. Эпителиальные структуры (эпидермис, волосы, сальные и потовые железы, ногти) развиваются из эктодермы, кожные нервы - из нейроэктодермы, а соединительнотканные волокни-
стые структуры, кровеносные и лимфатические сосуды, жировая клетчатка и мышцы имеют мезодермальное происхождение. Все компоненты кожи находятся в морфофункциональном единстве, обеспечивая множество ее физиологических функций.
1.1. ЭПИДЕРМИС
Эпидермис (epidermis) представляет собой многослойный плоский ороговевающий эпителий, обладающий выраженной регенераторной способностью. Толщина его варирует от 0,04 мм в области век до 1,6 мм в области ладоней и подошв. Он расположен на базальной мембране, отделяющей его от дермы, и состоит из клеток - эпидермоцитов. Выделяют пять его слоев:
1) базальный (stratum basale);
2) шиповатый (stratum spinosum);
3) зернистый (stratum granulosum);
4) блестящий (stratum lucidum);
5) роговой (stratum corneum).
Базальный слой располагается непосредственно на базаль-ной мембране и состоит из одного ряда призматических клеток, располагающихся перпендикулярно кожной поверхности. Над базальным слоем расположен шиповатый слой, состоящий из 3-8 рядов клеток многоугольной формы, отростки которых напоминают шипы растений. Мальпигиевым, или ростковым, слоем эпидермиса называется структурная единица, объединяющая базальный и шиповатый слои. Зернистый слой, расположенный над шиповатым, представляет собой 2 ряда (на ладонях и подошвах - 3-4) уплощенно-ромбических клеток. Блестящий слой наблюдается только на ладонях и подошвах. Он выглядит как бесцветная полоска, состоящая из вытянутых безъядерных клеток, расположенных над зернистым слоем. Поверхностным, соприкасающимся с внешней средой, является роговой слой, представляющий собой черепицеобразно расположенные безъядерные роговые чешуйки (корнеоциты). Толщина рогового слоя варьирует от 9 мкм в области век и мужских половых органов до 0,5 мм в области ладоней и подошв. В своей наружной части корнеоциты менее компактны и легко отделяются, обеспечивая процесс физиологического (нормального) шелушения.
В эпидермисе имеются множественные нервные окончания, а его питание осуществляется лимфой через расположенные между клетками межклеточные канальцы, ориентированные перпендикулярно поверхности кожи. Сосудов в эпидермисе нет.
Состав эпидермоцитов неоднороден и представляет собой следующие виды клеток:
1) кератиноциты;
2) меланоциты;
3) клетки Меркеля;
4) клетки Лангерганса;
5) клетки Гренстейна.
Кроме того, в эпидермисе могут встретиться отдельные лимфоциты, а иногда и тканевые базофилы.
1. Кератиноциты - эпителиальные клетки, расположенные во всех слоях эпидермиса, в каждом из которых они имеют различное строение.
Базальные кератиноциты имеют овально-цилиндрическую форму. Их ядра богаты хроматином, а контуры, равно как и плазмалемма, имеют ровные очертания. В цитоплазме базаль-ных клеток имеются фибриллярные образования в виде нежных тонофиламентов (толщиной 3-5 нм) и отдельных пучков - то-нофибрилл, состоящих из белка α-кератина. Их цитоплазма содержит большое количество рибосом и митохондрий, меланосо-мы (гранулы пигмента - меланина, поступающего из меланоци-тов) и лизосомы, осуществляющие фагоцитарную функцию. Между собой они соединяются с помощью десмосом, а к базаль-ной мембране прикреплены полудесмосомами. Десмосомы образованы двумя плотными белковыми образованиями, расположенными друг напротив друга за плазмалеммами соседних клеток.
В состав этих образований входят белки десмоплакин I и II типа, энвоплакин, периплакин и плакоглобин. С ними связываются подходящие к ним пучки внутриклеточных кератиновых тонофиламентов. Связывающие тонкие тонофиламенты, состоящие из белков десмоглеинов и десмоколинов, пронизывают плазмолемму и межклеточное пространство шириной 25-30 нм, и зигзагообразно соединяясь со встречными тонофиламентами соседней клетки, переходят в ее плазмолемму и десмосому. Полудесмосомы представляют собой образования, напоминающие половинки десмосом, прикрепленных к базальной мембране. Базальные кератиноциты осуществляют начало синтеза предшественника кератина - прекератина, который далее превращается в фибриллярный кератин, нити которого скручиваются, формируя кератиновые тонофиламенты. Тонофиламенты образуют каркас базальных ке-ратиноцитов, их десмосом и полудесмосом. Далее базальные филамен-ты постепенно трансформируются в тонофибриллы, уже различимые в световом микроскопе.
Базальные кератиноциты обладают митотической активностью и имеют две субпопуляции клеток. Одна из них постоянно
пролиферирует, в результате чего более молодые клетки остаются на некоторое время на базальной мембране, а более старые "выталкиваются" по направлению к поверхности кожи. В процессе движения они проходят все стадии дифференцировки (шиповатые клетки, зернистые клетки) и превращаются в кор-неоциты. Вторая субпопуляция клеток является резервной и находится в состоянии покоя.
Шиповатые кератиноциты имеют неправильную многоугольную форму. Ядра их округлые, с четко очерченной ядерной мембраной и хорошо просматривающимся ядрышком. В цитоплазме шиповатых клеток имеется большое количество фибриллярных образований, представляющих собой многочисленные пучки различных диаметров и конфигураций, рибосом, обеспечивающих усиление синтеза прекератина, а также сохраняются меланосомы. Десмосомы шиповатых клеток (узелки Биццоце-ро) более крупные - до 300-700 нм. Сцепление клеток шиповатого слоя осуществляется как за счет крупных десмосом, так и за счет проникновения цитоплазматических отростков одних клеток в углубления других (по типу "застегнутой молнии").
Зернистые кератиноциты имеют вытянутую параллельно поверхности кожи уплощенно-ромбическую форму. Ядра клеток обладают плеоморфизмом (округлые, овальные, вытянутые, лишенные ядрышка, подвергшиеся кариорексису и т. д.), а плаз-малемма на отдельных участках имеет зазубренные контуры. В цитоплазме встречается множество гранул кератогиалина, хорошо окрашенных и имеющих вид зерен с диаметром в несколько микрометров и, соответственно, хорошо различимых в световом микроскопе. Ни кератина, ни гиалина эти гранулы не содержат. Вместе с тонофибриллами они образуют тонофибриллярно-кера-тогиалиновые комплексы, обеспечивающие синтез филагрина - основного белка гранул.
Филагрин вместе с филаментами обеспечивает правильное расположение и сцепление корнеоцитов. Зернистые кератиноциты продолжают синтез прекератина, с уже большей молекулярной массой, чем у прекератина, синтезируемого базальными и шиповатыми клетками. Кроме того, в периферических отделах цитоплазмы зернистых клеток имеются кератиносомы (видоизмененные лизосомы или гранулы Од-ланда). Они содержат набор веществ (гидролитические ферменты, гли-колипиды, гликопротеиды, стерины), обеспечивающих образование межклеточного цементирующего гидрофобного вещества, выполняющего барьерную функцию рогового слоя эпидермиса.
Корнеоциты (ранее называвшиеся роговыми чешуйками) представляют собой безъядерные удлиненные клетки, располо-
женные черепицеобразно параллельно поверхности кожи. Они имеют длину до 10-12 мкм и толщину от 0,1 до 1 мкм. Сцепляются корнеоциты как за счет взаимопроникающих выростов плотной оболочки, так и остатков ороговевших десмосом. Кор-неоцит заполнен аморфным веществом, пронизанным керати-новыми фибриллами. В нижних рядах отдельных клеток сохраняется значительное количество гранул кератогиалина, что микроскопически соответствует блестящему слою эпидермиса. В небольших количествах роговые клетки содержат зрелый кератин.
2. Меланоциты - отросчатые клетки нейроглиального происхождения, расположенные в базальном слое эпидермиса и отвечающие за синтез меланина. Меланин представляет собой пигмент, защищающий кожу от вредного воздействия ультрафиолетовых лучей и придающий ей коричневый оттенок. Мела-ноциты имеют ядра с неровными очертаниями мембраны и многочисленные отростки, в цитоплазме которых содержатся множественные меланосомы - структуры, осуществляющие биосинтез меланина. Отростки клеток при помощи десмосом и полудесмосом соединяют меланоцит с несколькими соседними кератиноцитами, меланоцитами и базальной мембраной.
Меланин синтезируется из тирозина под воздействием каталитических ферментов - тирозиназы и ДОФА-оксидазы и посредством экзо-цитоза передается в межклеточное пространство, откуда и фагоцитируется кератиноцитами. Меланогенез с помощью меланоцитстимулирую-щего гормона контролируется гипофизом. В его процессе также участвуют ионы меди, витамин С, адреналин и норадреналин, гормоны щитовидных и половых желез. Цвет кожи и степень ее защиты от излучения зависят от количества меланина в кератиноцитах, а не от количества самих меланоцитов, синтез меланина в которых увеличивается под воздействием ультрафиолетового облучения.
3. Клетки Меркеля представляют собой осязательные нейро-эндокринные клетки, расположенные в базальном и шиповатом слоях эпидермиса, наружных слоях эпителия волосяного фолликула и некоторых слизистых оболочках. Они несколько крупнее кератиноцитов, имеют округлую или вытянутую форму, содержат удлиненное и слегка сегментированное ядро. В их цитоплазме располагаются филаменты цитокератинов и специфичные для этих клеток нейроэндокринные гранулы диаметром 100-200 нм.
В гранулах содержится комплекс нейропептидов (мет-инкефалин, фактор роста нервов, вазоинтестинальный полипептид, бомбезид, субстанция Р, эндорфины). С кератиноцитами клетки Меркеля соединя-
ются при помощи десмосом, а с нервными окончаниями в зоне своего нижнего полюса они формируют образования, напоминающие синапсы. В области этих образований в цитоплазме концентрируются нейро-эндокринные гранулы, а между плазмолеммой клеток и нервными окончаниями имеется синаптическая щель размером 50 нм. Некоторые клетки Меркеля свободны и не связаны с нервными волокнами.
4. Клетки Лангерганса - дендритические клетки, имеющие моноцитарное происхождение. Они относятся к системе фагоцитирующих мононуклеаров и являются тканевыми макрофагами. Эти клетки располагаются как в базальном, так и в нижних рядах шиповатого слоя эпидермиса. Их ядра имеют сегментированную форму и 1-2 ядрышка среднего размера. Цитоплазма клеток светлая и содержит множество органелл, среди которых встречаются особые структуры - так называемые гранулы Бирбека в форме теннисной ракетки, размерами от 30 нм в ширину до 200 нм в длину. Роль этих гранул полностью не установлена, однако известно, что в них накапливается эпидермальный кейлон, подавляющий пролиферацию кератиноцитов. Клетки Лангерганса имеют многочисленные отростки, с помощью которых соединяются друг с другом и с соседними кератиноцитами, образуя характерную ветвистую сеть. Отростки нижнего слоя клеток проникают в дерму.
Клетки Лангерганса не являются оседлыми и постоянно мигрируют в дерму и лимфатические узлы, выполняя свою макро-фагальную функцию. Они осуществляют процессинг и презентацию антигенов Т-хелперам (как проникающим в эпидермис, так и находящимся в дерме или лимфатических узлах), а также сек-ретируют ряд медиаторов (интерлейкины-1, -4, γ-интерферон, фактор некроза опухолей и др.), регулирующих пролиферацию и дифференцировку Т-лимфоцитов и других иммунокомпетент-ных клеток. Количество клеток Лангерганса составляет около 3-4 % всех эпидермоцитов.
5. Клетки Гренстейна представляют собой клетки, относящиеся к популяции дендритических клеток. Морфологически они весьма напоминают клетки Лангерганса, образуя похожую ветвистую сеть, но отличаются от них главным образом отсутствием гранул Бирбека и некоторых макрофагальных мембранных антигенов (HLA-DR). В то же время клетки Гренстейна экс-прессируют молекулы, весьма напоминающие антиген - маркер тимоцитов, таким образом свидетельствуя, что они иммуноло-гически отличаются от клеток Лангерганса. Функция этих клеток окончательно еще не установлена.
1.2. ДЕРМО-ЭПИДЕРМАЛЬНОЕ СОЕДИНЕНИЕ
Дермо-эпидермальное соединение (рис. 2) представляет собой сложную специализированную межклеточную структуру (базальную мембрану) толщиной 0,7-1,0 мкм, с помощью которой эпидермис прикрепляется к дерме. Эта структура состоит из следующих четырех компонентов:
1) клеточных мембран и полудесмосом базальных эпидер-моцитов;
2) светлой пластинки (lamina lucida);
3) плотной пластинки (lamina densa);
4) фиброретикулярной пластинки.
Полудесмосомы являются участками утолщения плазмолеммы ба-зальных кератиноцитов, соединенных с внутриклеточными кератино-выми тонофиламентами. Эти утолщения состоят из плакиновых белков, формирующих так называемый "антиген буллезного пемфигоида I типа" (BP 230). Кроме того, они содержат также плектин, от которого в сторону светлой пластинки отходят пронизывающие плазмолемму
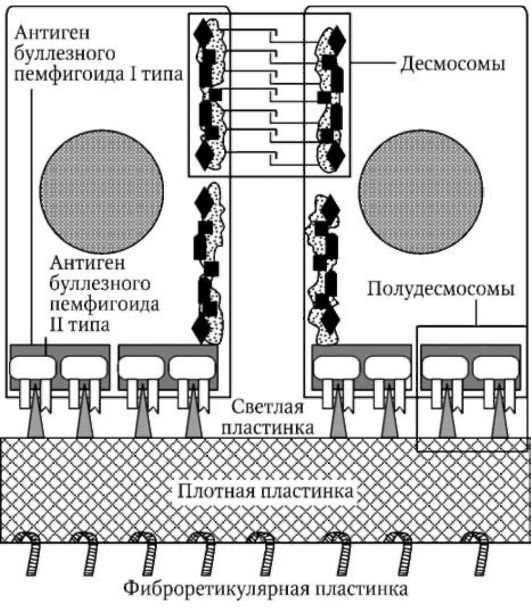
Рис. 2. Строение базального слоя и базальной мембраны эпидермиса
белковые тонофиламенты, состоящие из "антигена буллезного пемфи-гоида II типа" (BP 180) и интегрина.
Светлая пластинка (толщиной 30-40 нм) представляется преимущественно аморфной, однако в центральной части она содержит плотную линию, вертикально соединенную с внеклеточными тонофиламен-тами базальных полудесмосом. Светлая пластинка содержит ламинин, нидоген, энтактин, фибронектин и коллаген IV типа.
Плотная пластинка (толщиной 30-60 нм) состоит из волокнистого материала, скрепленного аморфным веществом. В ней содержится коллаген IV и V типа, протеогепарансульфат и ламинин.
Фиброретикулярная пластинка представляет собой субэпидермаль-ное сплетение соединительнотканных волокон, соединяющее дерму с базальной мембраной. Она образована пучкообразными крепящими (якорными) фибриллами, расположенными вертикально и соединенными верхними концами с плотной пластинкой, а также микрофибриллами эластических волокон и одиночными коллагеновыми волокнами.
Дермо-эпидермальное соединение выполняет как опорную функцию, так и функцию проникновения и транспортировки физиологических веществ и клеток из дермы в эпидермис и обратно.
1.3. ДЕРМА
Дерма (dermis) состоит из:
1) волокнистой соединительной ткани;
2) основного (аморфного) вещества;
3) клеточных элементов.
Волокнистая соединительная ткань и основное вещество объединяются в так называемое межклеточное вещество. Дерма в 15-40 раз толще эпидермиса и, помимо основных своих компонентов, содержит придатки кожи (волосы, сальные и потовые железы), кровеносные и лимфатические сосуды, мышцы и нервы.
Анатомически дерма разделяется на сосочковую и сетчатую (ретикулярную) части. Верхний ее отдел, представленный сосочками, вдающимися в эпидермис, относится к сосочковой дерме. Нижняя ее часть, сверху ограниченная концевыми отделами эпидермальных отростков, а снизу без видимой границы переходящая в гиподерму, - к ретикулярной. Кроме того, выделяют еще и периаднексальную дерму, подразумевая под этим понятием тот ее отдел, который непосредственно окружает придатки кожи. Сосочковая дерма вместе с периаднексальной объединяются в так называемую адвентициальную дерму, представляющую собой морфофункциональную единицу.
Волокнистая соединительная ткань состоит из коллаге-новых, эластических и ретикулиновых волокон.
1. Коллагеновые волокна сгруппированы в пучки, образованные фибриллярным белком - коллагеном, и обеспечивают функцию механического каркаса дермы. В сосочковом слое эти пучки мелкие и ориентированы хаотично. В сетчатом слое они более грубые и располагаются параллельно поверхности кожи.
Каждое коллагеновое волокно состоит из коллагеновых фибрилл (толщиной 40-50 нм), упакованных в определенном порядке и склеенных цементирующим веществом. По полипептидному строению в со-сочковом слое в основном располагается коллаген III типа, а в сетчатом - I типа. В состав их аминокислотных цепей входят пролин, окси-пролин, глицин, лизин и оксилизин. Часть лизина и пролина остается в коллагене в гидроксилированном виде, участвуя в образовании химических связей, обеспечивающих особую прочность коллагена.
2. Эластические волокна обеспечивают функцию возврата кожи в первоначальное состояние после ее растяжения. Они состоят из фибриллярной (гликопротеидной) части и аморфной, представляющей собой белок - эластин.
Соотношение этих компонентов в волокнах, расположенных в разных отделах дермы, неодинаково. В сосочковой части в волокнах преобладает гликопротеидный компонент, обеспечивая им необходимую прочность для участия в формировании базальной мембраны эпидермиса. В сетчатой части расположены "зрелые" эластические волокна, максимально насыщенные эластином (до 80-95 %), отвечающим за их эластические свойства. В состав эластина входят полипептидные цепи, содержащие преимущественно аланин, пролин, десмозин, глицин и валин.
3. Ретикулиновые волокна наиболее распостранены в периад-нексальной дерме и располагаются преимущественно вокруг сосудов, волосяных фолликулов, сальных и потовых желез. В связи с их свойствами окрашиваться нитратом серебра и не окрашиваться обычными методами, они также называются аргирофиль-ными. Ретикулиновые волокна состоят как из гликопротеидно-го, так и фибриллярного компонента, представляющего собой коллаген III типа. Напоминая по своему строению коллагено-вые, эти волокна выполняют функцию механического каркаса для сосудов и придатков кожи.
Основное вещество представляет собой комплекс гликоза-миногликанов, гликопротеидов и протеогликанов. Также оно содержит жиры, неорганические вещества и воду.
1. Гликозаминогликанами дермы являются хондроитин-(4,6)-сульфат, гепарин, гиалуроновая кислота и дерматансульфат.
Их молекулы представляют собой длинные цепи повторяющихся сахаров. Благодаря неплотной пространственной упаковке полисахаридов, гликозаминогликаны обладают способностью поглощать и удерживать большое количество воды, создавая необходимые условия как для клеточных реакций, так и для биохимических процессов. Гиалуроновая кислота не образует комплексов с белками, быстро повреждается гиалуронидазой, осуществляя еще большую проницаемость основного вещества.
2. Гликопротеиды представляют собой соединения длинных полипептидных цепей с короткими разветвленными цепями са-харидов. К наиболее важным из них относится фибронектин, играющий значительную роль в адгезии и кооперации клеток соединительной ткани, образовании коллагеновых волокон и развитии иммунных реакций.
3. Молекулы протеогликанов состоят из длинных белковых цепей, по бокам соединенных с большим количеством нераз-ветвленных цепей гликозаминогликанов. Они обеспечивают гидратацию, прочность основного вещества, его способность деформироваться и восстанавливаться.
Клеточные элементы дермы представлены фибробластами, тучными клетками, гистиоцитами, а также, в незначительном количестве, лимфоцитами и эозинофильными лейкоцитами.
1. Фибробласты - неоднородная популяция клеток, обеспечивающая синтез волокнистой соединительной ткани и основного вещества. К фибробластам относятся слабодифференциро-ванные фибробласты, дифференцированные фибробласты, фиброциты и фиброкласты. Они приступают к дифференцировке, образуясь из предшественников - перицитов - клеток, входящих в состав стенок кровеносных сосудов.
Слабодифференцированные фибробласты имеют звездчатую форму, округлое ядро и базофильную цитоплазму со слаборазвитой эндоплаз-матической сетью. Дифференцированные фибробласты имеют 12-20 мкм в диаметре, большие светлые ядра с крупными ядрышками, слегка ба-зофильную цитоплазму, в которой хорошо развиты клеточные орга-неллы. Среди последних особенно заметны характерные фибриллы. Дифференцированные фибробласты постепенно переходят в фиброциты - функционально неактивную субпопуляцию фибробластов, имеющих веретенообразную форму, вытянутое гиперхромное ядро и частично деградированные органеллы. На рибосомах фибробластов осуществляется синтез начальных полипептидных цепей коллагена, которые далее, после гидроксилирования и гликозилирования, сшиваются ди-сульфидными мостиками в трехцепочечную спираль тропоколлагена. После завершения процесса создания молекулы тропоколлагена фиб-
робласты его секретируют и дальнейший синтез зрелого коллагена осуществляется уже внеклеточно. Менее изучен процесс синтеза эластических и аргирофильных волокон, осуществляемый другими субпопуляциями фибробластов. Обновление межклеточного вещества происходит за счет его разрушения другой субпопуляцией клеток - фиброкластами, выполняющими фагоцитарную функцию и гидролиз соединительной ткани с помощью своих лизосом.
2. Тучные клетки (лаброциты, мастоциты) - клетки, являющиеся потомками стволовой клетки крови и тканевыми аналогами базофильных лейкоцитов крови. Лаброциты характеризуются плеоморфизмом, отличаясь друг от друга как размерами, так и очертаниями. Ядра клеток крупные, округлые или овальные. Характерная особенность этих клеток - наличие в их цитоплазме специфических гранул, содержащих биологически активные вещества: гистамин, гепарин, серотонин, хондроитин-сульфат, гиалуроновую кислоту и др. Содержимое этих гранул оказывает регулирующее влияние на проницаемость сосудов кожи, количество воды в межклеточном веществе дермы, пролиферацию и дифференцировку различных клеток, синтез гликоз-аминогликанов.
3. Гистиоциты представляют собой дермальные макрофаги и относятся к системе фагоцитирующих мононуклеаров. Они имеют очертания неправильной формы, образованные многочисленными отростками. Ядра клеток крупные, темные, бобовидной формы с отдельными инвагинациями. Цитоплазма ва-куолизирована и имеет хорошо развитые многочисленные лизо-сомы, которые с помощью своих лизосомальных ферментов осуществляют главную функцию макрофагов - фагоцитоз. Если же захваченные чужеродные вещества не могут быть переработаны окончательно, макрофаги осуществляют их процессинг, переводя антигены веществ в высокоиммунную форму. Далее макрофаги, вместе с собственным антигеном, экспрессируют переработанный продукт на своей поверхности для последующей кооперации его с лимфоцитами. Кроме того, макрофаги секре-тируют различные медиаторы межклеточного взаимодействия (интерлейкин-1, γ-интерферон, фактор некроза опухолей и др.), осуществляющие регуляцию пролиферации и дифференцировки иммунокомпетентных клеток и фибробластов. В цитоплазме некоторых гистиоцитов обнаруживаются гранулы Бирбека, свидетельствующие о том, что эти макрофаги являются мигрирующими из эпидермиса и обратно в эпидермис клетками Лангерганса.
1.4. ПОДКОЖНАЯ ЖИРОВАЯ КЛЕТЧАТКА (ГИПОДЕРМА)
Подкожная жировая клетчатка анатомически представляет собой волокнистую соединительную ткань, соединенную в морфо-функциональном единстве со скоплениями жировых клеток - липоцитов. Основная функция гиподермы - защита кожи от механических травм и разрывов, обеспечивается ее мягкостью и подвижным прикреплениям к подлежащим тканям организма. Поскольку волокнистая ткань гиподермы является продолжением соединительнотканных волокон сетчатого слоя дермы, эти два отдела кожи (дерма и подкожная жировая клетчатка) не имеют между собой четкой границы. Липоциты в гиподерме собраны в так называемые жировые дольки (лобули), которые разделены междольковыми перегородками - пучками соединительнотканных волокон (трабекулами или септами). Соответственно этим структурам подкожная жировая клетчатка разделяется на лобу-лярную и септальную (трабекулярную) части.
Септальная часть гиподермы по составу соединительнотканных волокон, основного вещества и клеточных элементов ничем не отличается от ретикулярной дермы. Исключение составляет только пространственная ориентация ее коллагеновых пучков, которые, спускаясь из дермы, изменяют свое горизонтальное расположение.
Лобулярная часть подкожной жировой клетчатки состоит из скоплений жировых клеток двух видов: белых и бурых липо-цитов.
1. Белые липоциты составляют подавляющее большинство жировых клеток. Они содержат вытянутое, смещенное к периферии клетки ядро, которое сдавливается крупной каплей жира, расположенной в их цитоплазме. Цитоплазма клеток, окружающая в виде узкого ободка каплю триглицеридов, содержит свободные рибосомы и митохондрии, слаборазвитые эндоплазма-тическую сеть и комплекс Гольджи. Скопления белых липоцитов обеспечивают, главным образом, энергетическую и защитно-механическую функцию.
2. Бурые липоциты, в отличие от животных, встречаются у человека в небольшом количестве, прежде всего в области шеи и межлопаточной области. У новорожденных бурая жировая ткань развита достаточно хорошо, но с возрастом она постепенно заменяется белой. Бурый липоцит значительно меньше белого, его ядро расположено в центре клетки, а цитоплазма содержит хорошо развитые митохондрии, окружающие многочисленные мелкие капельки жира. Митохондрии бурых липоцитов
содержат белок - термогенин, отсутствующий в митохондриях белых липоцитов. Термогенин разделяет дыхание и окислительное фосфорилирование, вызывая выделение значительного количества энергии, расходующейся на согревание организма.
Подкожная жировая клетчатка содержит луковицы волос, секреторные отделы эккринных и апокринных потовых желез, нервы, лимфатические и кровеносные сосуды. Мышцы в гиподерме отсутствуют.
1.5. МЫШЦЫ КОЖИ
Мышечными элементами кожи являются пучки гладких мышечных волокон, располагающиеся в дерме, а также поперечно-полосатые (мимические мышцы), которые обнаруживаются в коже лица. Наибольшее количество пучков гладких мышечных волокон располагается в области мошонки, крайней плоти, сосков молочных желез, ануса и подмышечных впадин. Основная их масса прикрепляется к центральным участкам волосяных фолликулов. Сокращаясь, она вызывает движение волос с образованием так называемой "гусиной кожи", а также обеспечивает выдавливание секрета сальных желез. Эти пучки носят название "мышц, поднимающих волосы" (mm. arrectores pilorum). На отдельных анатомических участках (лицо, тыл кистей и стоп, волосистая часть головы) располагаются гладкие мышцы, не соединенные с волосяными фолликулами. Поперечно-полосатые мышцы имеются только в коже лица, обеспечивая при сокращении его мимическую функцию.
1.6. ИННЕРВАЦИЯ КОЖИ
Кожа, являясь пограничным органом, осуществляющим контакт с внешней средой, снабжена нервно-рецепторным аппаратом, который связывает ее с центральной нервной системой с помощью нервных проводников. Иннервацию кожи осуществляют черепно-мозговые и спинно-мозговые нервы, а также вегетативные нервные волокна. Эти волокна подразделяются на моторные, чувствительные, секреторные и сосудодвигательные. Нервы кожи по ходу кровеносных и лимфатических сосудов попадают в гиподерму, где образуют крупное сплетение. От него отходят более тонкие ветви, которые разветвляются и образуют глубокое дермальное сплетение. Поднимаясь к эпидермису, мелкие веточки формируют поверхностное сплетение, располо-
женное в сосочковом слое дермы. Каждое из нервных сплетений обеспечивает иннервацию близлежащих придатков и сосудов. Аксоны нейронов ядер боковых рогов спинного мозга формируют начальные преганглионарные волокна симпатической нервной системы, заканчивающиеся в паравертебральных ганглиях. Постганглионарные нервные волокна симпатической нервной системы осуществляют эфферентную вегетативную инервацию сальных и потовых желез, мышц, поднимающих волосы, а также сосудов, вызывая их сужение. Наибольшее количество элементов нервной ткани обнаруживается в коже кончиков пальцев, красной каймы губ и половых органов. Чувствительная иннервация кожи осуществляется дендритами нейронов спи-нальных ганглиев, заканчивающихся специальными нервными окончаниями. Все рецепторные окончания разделяются на свободные и несвободные.
Свободные нервные окончания представляют собой концевые ветви дендрита, лишенные вспомагательных глиальных клеток. Такие окончания (в наибольшем количестве в коже век и гениталий) иннервируют эпидермис, подойдя к которому, нервное волокно теряет миелиновую оболочку и в виде заостренного осевого цилиндра заканчивается в межклеточном канальце базального или шиповатого слоя эпидермиса. Подобные свободные окончания, кроме того, имеются в поверхностном нервном сплетении дермы, а также в виде корзинчатых структур располагаются вокруг волосяных фолликулов.
Несвободные нервные окончания, в свою очередь, классифицируются на неинкапсулированные и инкапсулированные. Инкапсулированные от неинкапсулированных отличаются тем, что помимо нейроглиальных клеток, участвующих в рецепции, они имеют наружную соединительнотканную оболочку.
1. К неинкапсулированным несвободным рецепторным окончаниям относятся диски Меркеля, представляющие собой расширенные в виде дисков концевые отделы нейронов, формирующих синапсы с клетками Меркеля.
2. Инкапсулированные нервные окончания представляют собой пластинчатые тельца Фатера-Пачини, тельца Руффини, концевые колбы Краузе, осязательные тельца Мейсснера и ге-нитальные тельца Догеля.
Пластинчатые тельца Фатера-Пачини расположены глубже других рецепторных окончаний, как правило, в нижнем отделе дермы и в подкожной жировой клетчатке. Они имеют размеры, позволяющие иногда увидеть их невооруженным глазом. Располагаются тельца преимущест-
венно в области пальцев кистей, половых органов и сосков молочных желез. В центральной части их капсулы имеется полость в форме цилиндра, в которую входит теряющий миелиновую оболочку голый осевой стержень нервного волокна. Внутри капсулы дендрит покрывается несколькими слоями нейролеммоцитов, вместе с которыми он образует внутреннюю капсулу. Снаружи это образование покрыто соединительнотканной капсулой, состоящей из многослойных коллагеновых пластинок, содержащих фиброциты.
Тельца Руффини обнаруживаются в нижнем отделе дермы и верхней части гиподермы и состоят из тонкой капсулы, к которой подходит нервное волокно, разветвляющееся на множественные, лишенные мие-линовой оболочки, концевые веточки.
Концевые колбы Краузе располагаются под сосочками в центральной части дермы. Они представляют собой механорецепторы, расположенные преимущественно в области кожи верхних и нижних конечностей. По своему строению их капсула напоминает колбу, в которой располагаются безмиелиновые терминальные веточки, являющиеся продолжением толстого афферентного нервного волокна.
Осязательные тельца Мейсснера расположены в сосочках. Они широко распространены и в большом количестве обнаруживаются в коже и слизистых оболочках пальцев стоп и кистей, ладоней, век, губ, половых органов, сосков молочных желез, языка. Структурно они имеют тонкую соединительнотканную капсулу, внутри которой расположен потерявший свои оболочки спиралевидный осевой цилиндр. Перпендикулярно цилиндру располагаются нейролеммоциты.
Генитальные тельца Догеля встречаются преимущественно в коже половых органов. Именно они обеспечивают повышенную чувствительность этой зоны. Это объясняется тем, что колба тельца Догеля окружена тончайшей соединительнотканной капсулой, в которую входят осевые цилиндры сразу нескольких нервных клеток. Раздражение через одно тельце одновременно нескольких дендритов разных нервов приводит к иррадиации возбуждения.
1.7. КРОВЕНОСНАЯ И ЛИМФАТИЧЕСКАЯ СИСТЕМЫ КОЖИ
Кровоснабжение кожи осуществляется из кожных ветвей артерий межмышечных фасциальных перегородок, а также мы-шечно-кожных и надкостно-кожных артерий. Входя в трабеку-лярную часть подкожной жировой клетчатки, они создают ответвления, питающие жировые дольки. В нижней части дермы артерии разделяются на многочисленные веточки, которые в горизонтальном направлении образуют анастомозы с пучками веточек соседних сосудов, формируя на границе дермы с гиподермой глубокую сосудистую сеть. Эта сосудистая сеть
осуществляет питание поверхностного слоя гиподермы и потовые железы. Часть сосудистых веточек основных артерий поднимается вверх и питает волосяные фолликулы и сальные железы. Другая их часть, истончаясь, поднимается до границы между сосочковым и ретикулярным слоями дермы, где образует многочисленные горизонтальные анастомозы поверхностной сосудистой сети. От нее берут начало капилляры, заканчивающиеся в сосочках дермы петельками, стенки которых состоят всего из 2-3 эндотелиоцитов. Из капилляров кровь попадает в венулы, которые образуют извилистую сеть непосредственно под сосочками. В центральной части дермы расположена вторая венозная сеть, в нижней части дермы - третья, а в подкожно-жировой клетчатке - четвертая сеть, содержащая уже крупные вены. В коже лица подсосочковая венозная сеть имеет более крупные, но укороченные сосуды, расположенные горизонтально поверхности кожи и соединяющиеся с глубжележа-щей венозной сетью под прямым углом.
Лимфатическая система кожи образует поверхностную сеть, начинающуюся с сосочковых синусов (лимфатических капилляров, расположенных в дермальных сосочках). Глубокая сеть лимфатических капилляров имеет отводящие сосуды, проникающие в гиподерму и снабженные клапанами. Анастомозируя между собой в подкожной жировой клетчатке, лимфатические сосуды формируют глубокое лимфатическое сплетение (plexus lymphaticus cutaneus).
1.8. ПРИДАТКИ КОЖИ
Придаточные образования кожи имеют эпителиальное происхождение и включают в себя волосы, эккринные и апокрин-ные потовые железы, сальные железы и ногти.
Волосы
Волос (pilus) анатомически включает в себя наружную часть, находящуюся выше уровня поверхности кожи - стержень и корень, расположенный в дерме, а иногда достигающий гиподермы. Корень, окруженный эпителием, внешне напоминающим инвагинированное продолжение эпидермиса, называется волосяным фолликулом. Снаружи волосяной фолликул окружает соединительнотканная волосяная сумка.
По толщине и длине стержней волосы делятся на:
1) длинные, расположенные в области волосистой части головы, лобка, половых органов, подмышечных впадин, бороды и усов;
2) щетинистые, локализованные в области бровей, ресниц, наружного слухового прохода и ноздрей;
3) пушковые, обнаруживаемые на всей остальной, большей части поверхности тела.
Корни и стержни волос сформированы тремя слоями: мозговым веществом, корковым веществом и кутикулой волоса, однако в пушковых волосах мозговое вещество отсутствует, поэтому длинные и щетинистые волосы толще пушковых.
Мозговое вещество состоит из одного или нескольких слоев клеток, дифференцирующихся из стволовых клеток, расположенных в нижнем полюсе корня. По мере дифференцировки эти клетки приобретают комплекс Гольджи, эндоплазматическую сеть, рибосомы и митохондрии. В средних слоях они уже имеют кератиновые филаменты и трихо-гиалиновые гранулы. В верхних отделах в клетках появляются пузырьки, постепенно сливающиеся в крупные капли, заполненные воздухом. Далее клетки теряют воду, постепенно высыхают и превращаются в "тени", разделяемые воздухом и гранулами меланина.
Корковое вещество также состоит из 1-3 слоев клеток, дифференцирующихся из клеток нижней части корня. Эти клетки несколько удлинены и располагаются вертикально. В них первоначально образуются все органеллы, которые постепенно теряются в процессе дифферен-цировки. В клетках коркового вещества образуются тонофиламенты, которые, формируя кератин, постепенно ориентируются вдоль длинного диаметра клетки, в них имеется также меланин и накапливаются пузырьки воздуха. На последней стадии дифференцировки образуются роговые пластинки, скрепленные цементирующим веществом.
Клетки кутикулы волоса происходят из камбиальных клеток корня и образуют 7-10 слоев. В этих клетках в процессе дифференцировки постепенно формируются органеллы, появляются темные гранулы, содержащие большое количество цистеина, и кератиновые тонофиламен-ты. Постепенно гранулы концентрируются возле мембраны клетки и сливаются, образуя так называемую экзокутикулу. В конечной стадии они резко уплощаются и располагаются черепицеобразно. Между собой клетки скреплены десмосомами.
Волосяной фолликул (рис. 3) состоит их трех сегментов:
1) нижнего сегмента (inferior), расположенного между основанием фолликула и местом прикрепления мышцы, поднимающей волос;
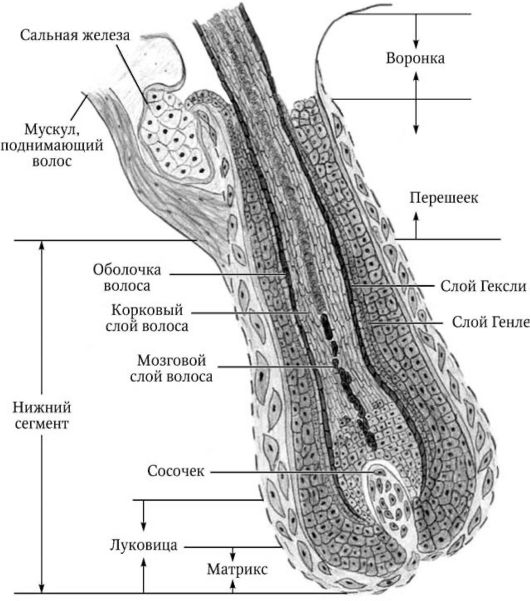
Рис. 3. Строение волоса
2) перешейка (istmus), или средней части, локализованной между участком прикрепления мышцы, поднимающей волос, и местом впадения выводного протока сальной железы;
3) воронки (infundibulum), соединяющей место впадения протока сальной железы с устьем фолликула.
1. Нижний сегмент представляет собой волосяную луковицу, с которой сливаются внутреннее и наружное корневые влагалища.
Нижняя часть волосяной луковицы называется матриксом и состоит из недифференцированных клеток фолликулярного эпителия, среди
которых встречаются меланоциты и клетки Лангерганса. В нижнюю часть луковицы также вдается соединительнотканный (волосяной) сосочек, богато снабженный сосудами. В луковице происходит активное митотическое деление фолликулярных клеток, обеспечивающее рост волоса со скоростью 1 см в месяц и длящееся от 2 до 7 лет (фаза анаге-на). В эту фазу идет активное образование пигмента - меланина, обусловливающее окраску волосяного стержня. После фазы анагена наступает промежуточная фаза катагена, длящаяся всего несколько недель, во время которой меланин не образуется, фолликул несколько сокращается и его основание перемещается вверх по направлению к поверхности кожи. После этого волос переходит в стадию отдыха (фаза тело-гена), продолжающуюся 3 месяца, во время которой он легко выдергивается или спонтанно выпадает. С момента самопроизвольного выпадения начинается рост нового волоса, который вырастает из того же фолликула, что и старый. Из каждого волосяного фолликула может вырасти от 20 до 30 волос. Выпадение волос является нормальным процессом обновления.
Внутреннее корневое влагалище простирается от матрикса до места впадения протока сальной железы, формируется эпителиальными клетками луковицы и состоит из кутикулы внутреннего влагалища, гра-нулосодержащего слоя Гексли и бледно-клеточного слоя Генле. Первоначально в клетках внутреннего корневого влагалища образуются то-нофиламенты и трихогиалиновые гранулы. Между собой и с клетками кутикулы волоса клетки внутреннего влагалища соединены десмосома-ми. В процессе дифференцировки клетки теряют органеллы, а также трихогиалиновые гранулы, десмосомы постепенно разрушаются, межклеточное пространство заполняется аморфным цементирующим веществом. В конечном итоге ороговевшие клетки кутикулы образуют че-репицеобразно расположенные безъядерные чешуйки.
Наружное корневое влагалище образовано неороговевающими фолликулярными клетками, соединяющимися в области впадения протока сальной железы с клетками эпидермиса, который в области воронки своими эпидермоцитами образует верхнюю часть влагалища.
2. Перешеек представлен стержнем волоса, покрытым внутренним и наружным корневыми влагалищами.
3. Воронка представляет собой стержень, окруженный наружным корневым влагалищем, являющимся в этой части продолжением эпидермиса.
Потовые железы
Потовые железы (glandulae sudoriferae) относятся к железистому аппарату кожи. По своему строению и функциональному значению они разделяются на эккринные и апокринные.
Эккринные (мерокриновые) железы - наиболее распространены и встречаются по всему телу, за исключением кожи губ и слизистых оболочек мочеполовых органов. На 1 см2 кожи располагается от 85 до 250 желез. Наибольшее их количество сосредоточено в области кожи ладоней, подошв, лба, живота, груди и подмышечных впадин. По своему строению они являются простыми трубчатыми, их секреция не сопровождается гибелью секреторных клеток. Железы состоят из секреторного отдела и выводного протока.
1. Секреторный отдел расположен на границе дермы и гиподермы или в верхней части гиподермы. Он закручен в клубочек и состоит из двух рядов клеток цилиндрического эпителия двух типов:
1) миоэпителиальных (миоэпителиоцитов), расположенных на базальной мембране;
2) секреторных (экзокриноцитов, судорифероцитов), лежащих проксимально.
Миоэпителиальные клетки располагаются между базальной мембраной железы и судорифероцитами. Между ними обнаруживаются малочисленные десмосомы, а иногда - отростки секреторных клеток. Ядра этих клеток имеют удлиненную форму с неровными очертаниями и располагаются в апикальном отделе клетки. Органеллы клеток очень немногочисленны и лежат в промежутках между множественными миофиламентами, занимающими большую часть цитоплазмы. Кроме того, в ней встречаются кератиновые фибриллы и немногочисленные гранулы гликогена.
Секреторные клетки делятся на светлые (слегка оксифильные) и темные (базофильные) клетки. Светлые клетки имеют округлые или овальные равномерно хроматизированные ядра, хорошо развитую эн-доплазматическую сеть, лизосомы, гранулы глигогена и капли липи-дов. Они продуцируют электролитную (жидкую) часть пота. Темные клетки чаще выстилают поверхность просвета железы, которая здесь представлена многочисленными ворсинками и выростами их цитолем-мы. В этих клетках имеются неправильной формы ядра с концентрацией хроматина по мембране, хорошо развитая гранулярная эндоплазма-тическая сеть и рибосомы, содержатся крупные гранулы (350-500 нм) протеогликанов и сиаломуцин, а также встречаются тонофиламенты, ориентированные в апикальной части параллельно цитолемме. Темные клетки формируют макромолекулярные углеводные и белковые компоненты пота.
2. Выводной проток эккринной потовой железы поднимается вверх из секреторного отдела и, слабо извиваясь, входит в основание эпидермального отростка, где приобретает спиральную
форму. Открывается выводной проток свободно на поверхности кожи. Диаметр его просвета - 12-15 мкм. Анатомически проток подразделяется на дермальный и эпидермальный отделы.
Дермальный отдел протока состоит из двух слоев базофильного кубического эпителия, клетки которого соединены между собой десмосо-мами. Ядра этих клеток имеют неправильные очертания, маленькое ядрышко и конденсацию хроматина по периферии. В базальной части цитоплазмы располагаются слаборазвитые комплекс Гольджи, эндо-плазматическая сеть, рибосомы и другие органеллы, а в апикальной ее части - множественные тонофиламенты, а также гранулы гликогена. Цитолемма имеет неправильные контуры и со стороны просвета образует многочисленные ворсинки.
Эпидермальный отдел протока выстлан одним слоем внутренних (люминальных) клеток, окруженных несколькими слоями наружных клеток, которые соединены между собой десмосомами. Строение этих клеток подобно стрению клеток дермального отдела, однако по мере продвижения к поверхности кожи в их цитоплазме увеличивается число тонофиламентов, появляются гранулы кератогиалина, свидетельствующие об ороговении клеток протока аналогично кератиноцитам эпидермиса.
Секрет мерокриновых потовых желез (пот) представляет собой гипотонический раствор солей натрия, калия, железа, кальция, марганца и других металлов, содержащий также аминокислоты, мочевину, аммиак и молочную кислоту. Основным компонентом его является вода (99 %). Значения pH находятся между 4и7.
Апокринные железы распространены на ограниченных участках тела - в коже подмышечных впадин, лобка, паховых складок, половых органов, окружности ануса и соска молочных желез. Процесс секреции этих желез сопровождается разрушением апикальных отделов секреторных клеток. Секрет содержит вещества, разлагающиеся бактериями, что приводит к появлению запаха. Предполагается, что эти железы являются рудиментарными образованиями, так как они более развиты у животных, которые используют запах секрета в качестве коммуникационного средства. Железы состоят из секреторного (концевого) отдела и протока.
1. Секреторный отдел расположен в нижней части дермы или в гиподерме, глубже чем у эккринных потовых желез. Строение концевого отдела также имеет отличия. Он состоит из секреторных клеток только одного типа, окруженных снаружи миоэпителиоцитами.
Секреторные клетки (судорифероциты) имеют цилиндрическую форму, измененную в апикальной части ворсинками, вдающимися в просвет железы. Они содержат удлиненные ядра с уплотнением хроматина по периферии, эндоплазматическую сеть, комплекс Гольджи, митохондрии, гранулы пигмента, а также секреторные гранулы и многочисленные тонофиламенты. Секреторные гранулы неоднородны и имеют размеры от 100 нм до 1 мкм.
Миоэпителиоциты имеют такое же строение, как и миоэпителиаль-ные клетки эккринных потовых желез.
2. Проток апокринной железы открывается в воронку волосяного фолликула сразу же над местом впадения протока сальной железы. Отдельные протоки открываются непосредственно в эпидермис. Их строение аналогично строению дермального отдела протока эккринной потовой железы.
Пот апокринных желез более щелочной, чем пот эккринных, а также дополнительно содержит липиды, которые в секрете ме-рокриновых желез отсутствуют.
Сальные железы
Сальные железы (glandulae sebaceae) являются альвеолярными железами голокринового типа, так как процесс их секреции сопровождается разрушением сецернирующих клеток. Эти железы широко распространены, непосредственно связаны с волосяными фолликулами и отсутствуют только в коже ладоней и подошв. В один фолликул впадает проток одной или нескольких желез. На отдельных участках - в области малых половых губ, головки полового члена, внутреннего листка крайней плоти и венечной борозды (тизониевые железы), края век (мейбомиевы железы), а также соска и околососкового поля они не связаны с волосяными фолликулами (свободные железы). Размеры, количество и глубина залегания сальных желез сильно варьируют. Наиболее крупные железы обнаруживаются в возрасте 16-27 лет в области груди, кожи лица и спины.
Сальная железа состоит из секреторного (концевого) отдела и протока.
1. Секреторный отдел объединяет 1-2 дольки желез, которые состоят из альвеол, открывающихся в единый проток. Альвеола построена из себоцитов, которые формируют несколько слоев. Наружный (камбиальный) слой состоит из слабодиффе-ренцированных клеток. Эти клетки имеют крупные ядра, занимающие большую часть клетки. Цитоплазма содержит много-
численные митохондрии, рибосомы, гранулы гликогена. Эндо-плазматическая сеть развита слабо. Внутри от росткового слоя обнаруживаются частично дифференцированные крупные клетки, цитоплазма которых содержит капли липидов и полноценную эндоплазматическую сеть. Максимально дифференцированные клетки значительно увеличены в размерах, содержат большое количество капель жира, занимающих большую часть цитоплазмы, а также гиперхромные пикнотизированные ядра и немногочисленные органеллы. По мере накопления жира формируется полный некроз клетки, ведущий к образованию секрета железы.
2. Проток сальной железы выполнен многослойным эпителием кератинизированных клеток, переходящим в эпителий наружного волосяного влагалища. В некоторых клетках обнаруживаются ядра. Цитолемма клеток со стороны просвета имеет множество микроворсинок, а клетки с наружной стороны протока окружены базальной мембраной коллагеновых фибрилл.
Секрет сальных желез обеспечивает смазку волос и поверхности кожи, обладает водоотталкивающим эффектом, а также участвует в образовании кислотной мантии.
Секрет сальных желез (кожное сало) более чем на 50 % составляют триглицериды в виде свободных жирных кислот, а также восковые эфиры, сквален, холестероловые эфиры и хо-лестерол.
Ногти
Ноготь (unguis) представлен роговой пластинкой, покрывающей эпителий тыльной части дистального отдела фаланги пальца кисти или стопы. Этот эпителий в комплексе с подлежащей соединительной тканью образует ногтевое ложе. Ноготь разделяется на скрытую в ногтевом желобке часть - корень и видимую его часть - тело. Тело ногтя имеет розовый цвет, придаваемый ему кровью гемокапилляров. Проксимальная, элипсо-видной формы, прилегающая часть тела окрашена в белый цвет и называется луночкой ногтя. С боковой и задней части тело окружено кожными складками - ногтевыми валиками. Дисталь-ная, выступающая часть ногтевой пластинки называется свободным краем ногтя.
1. Ногтевая пластинка снаружи имеет гладкую поверхность, а ее внутренняя, прилегающая к ногтевому ложу сторона, образует множество гребешков и бороздок. Она состоит из роговых
чешуек полноценного кератина высокой прочности, сформировавшихся в результате "твердой" кератинизации клеток ногтевого ложа. Такая кератинизация, опускающая стадию образования кератогиалина, обеспечивает присутствие в ногтевой пластинке до 90 % прочного α-кератина. Проксимальная часть ногтевого ложа, обеспечивающая рост ногтевой пластинки, называется матрицей ногтя.
2. Матрица ногтя в основном состоит из эпителиальных клеток - онихобластов, помимо которых в ней встречаются ме-ланоциты, клетки Меркеля и клетки Лангерганса. Нижняя часть матрицы является ростковым слоем, а ее верхняя часть отвечает за дифференцировку клеток в роговое вещество пластинки. Это обеспечивает рост ногтя в длину. Роговая пластинка, продвигаясь по ногтевому ложу к дистальному отделу фаланги, одновременно остается плотно к нему прикрепленной.
3. Ногтевое ложе в дистальной части представляет собой эпидермис с неровной поверхностью, исчерченной бороздками и покрытой гребешками, входящими в соответствующие бороздки и гребешки ногтевой пластинки. Дерма под ногтевым ложем имеет множество сосочков, богато снабженных кровеносными сосудами. В области самой проксимальной части корня роговой слой эпидермиса фаланги заворачивается и обхватывает край пластинки, плотно к ней прикрепляясь и образуя желобок. Эта зона инверсии рогового слоя эпидермиса называется надногтевой кожицей, или эпонихием.
Ногти растут со средней скоростью около 1 мм в неделю, причем на кистях быстрее, чем на стопах. Ногтевая пластинка полностью обновляется за 180-230 дней. Наиболее медленно растут ногти больших пальцев стоп.