Медицинская и биологическая физика. Курс лекций с задачами : учеб. пособие / В.Н. Федорова, Е.В. Фаустов. - 2008. - 592 с.
|
|
|
|
ЛЕКЦИЯ 12 БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ
1. Ионные потоки в мембране.
2. Потенциал покоя. Уравнение Гольдмана-Ходжкина-Катца.
3. Потенциал действия и его распространение.
4. Основные понятия и формулы.
5. Задачи.
В живых организмах происходят разнообразные электрические процессы. Функционирование живых тканей сопровождается электрическими явлениями. Генерация и распространение электрических потенциалов - важнейшее физическое явление в живых клетках и тканях.
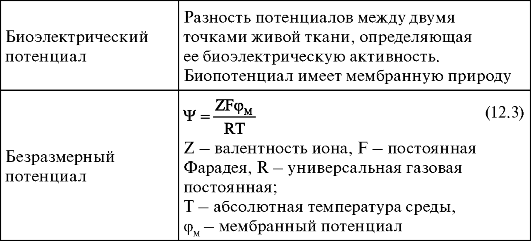
Биоэлектрический потенциал - разность потенциалов между двумя точками живой ткани, определяющая ее биоэлектрическую активность. Биопотенциал имеет мембранную природу.
12.1. Ионные потоки в мембране
Сквозь мембрану проходят потоки ионов, для которых справедливо уравнение Нернста-Планка (11.18):
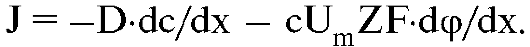 Эйнштейн показал, что подвижность ионов Um выражается через коэффициент диффузии D и абсолютную температуру Т: Um = D/(RT), тогда уравнение Нернста-Планка принимает вид
Эйнштейн показал, что подвижность ионов Um выражается через коэффициент диффузии D и абсолютную температуру Т: Um = D/(RT), тогда уравнение Нернста-Планка принимает вид
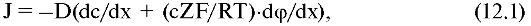 где с - концентрация ионов; R - универсальная газовая постоянная.
где с - концентрация ионов; R - универсальная газовая постоянная.
В первом приближении можно считать, что поле внутри мембраны однородно. В этом случае dp/dx = φм/l, где L - толщина мембраны, а φм - разность потенциалов между ее поверхностями, называемая мембранным потенциалом. Теперь уравнение Нернста-Планка будет иметь вид
 Для упрощения записи введем величину ψ, которую называют
Для упрощения записи введем величину ψ, которую называют
безразмерным потенциалом:
 12.2. Потенциал покоя. Стационарный потенциал Гольдмана-Ходжкина-Катца
12.2. Потенциал покоя. Стационарный потенциал Гольдмана-Ходжкина-Катца
Потенциал покоя
В нормально функционирующей клетке поддерживается наиболее благоприятный состав ионов. Различие в их концентрациях по разные стороны мембраны приводит к появлению разности потенциалов.
Потенциал покоя - разность потенциалов между цитоплазмой и окружающей средой в нормально функционирующей клетке.
В 1902 г. Бернштейн предположил, что потенциал покоя обусловлен проницаемостью мембраны для ионов К+. В состоянии покоя плотность потока ионов равна нулю, и уравнение (12.5) принимает следующий вид:
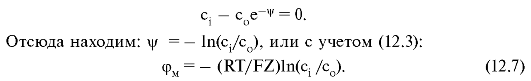 Потенциал, определяемый формулой (12.7), называют равновесным потенциалом. Расчеты,
выполненные по этой формуле, существенно расходятся с
экспериментальными данными при низких концентрациях К+. Это указывает на
то, что «калиевая» теория потенциала несовершенна.
Потенциал, определяемый формулой (12.7), называют равновесным потенциалом. Расчеты,
выполненные по этой формуле, существенно расходятся с
экспериментальными данными при низких концентрациях К+. Это указывает на
то, что «калиевая» теория потенциала несовершенна.
Стационарный потенциал Гольдмана-Ходжкина-Катца
Причина отклонения равновесного потенциала от опытных данных заключается в проницаемости мембраны и для других ионов, которые вносят свой вклад в образование мембранного потенциала. Основной вклад в суммарный поток зарядов, а следовательно, в создание и поддержание потенциала покоя, помимо К+, вносят ионы Na+, Cl-. Суммарная плотность потока этих ионов с учетом их знаков равна
 Знак «-» перед JCl- указывает на отрицательный заряд.
Знак «-» перед JCl- указывает на отрицательный заряд.
В стационарном состоянии (когда параметры системы не изменяются) суммарная плотность потока равна нулю, т.е. число различных ионов, проходящих в единицу времени через мембрану внутрь клетки, равно числу ионов, выходящих из клетки через мембрану: J = 0
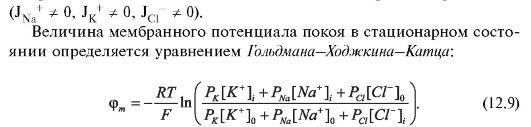 Здесь, во избежание сложностей с индексацией, для обозначения концентрации вместо буквы с использованы квадратные скобки: []i и[]0 - концентрации соответствующих ионов внутри и вне клетки.
Здесь, во избежание сложностей с индексацией, для обозначения концентрации вместо буквы с использованы квадратные скобки: []i и[]0 - концентрации соответствующих ионов внутри и вне клетки.
Очевидно, что формула для равновесного потенциала (12.7) получается из формулы стационарного потенциала (12.9) при РNa = 0 и РС1.= 0 Таким образом, уравнение Гольдмана-Ходжкина-Катца существенно уточняет теорию Бернштейна.
Большая часть сведений о нервных клетках получена при изучении аксона кальмара, достигающего почти миллиметровой толщины. Его изолированные нервные волокна довольно долго сохраняют способность передавать нервные импульсы. Рассчитаем стационарный мембранный потенциал для клеток аксона кальмара. При вычислении по формуле (12.9) вместо самих коэффициентов проницаемости можно использовать отношение между ними, которое для аксона выражается следующими числами:
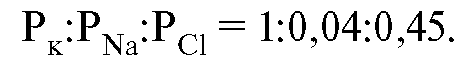 Значения концентраций приведены ниже.
Значения концентраций приведены ниже.
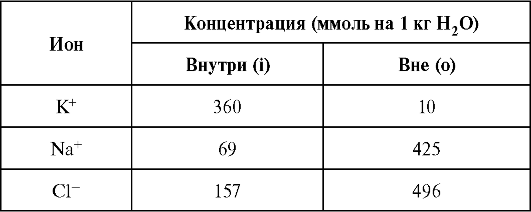 Подставив эти значения в формулу (12.9) при Т = 303 К (30 °С), получим:
Подставив эти значения в формулу (12.9) при Т = 303 К (30 °С), получим:
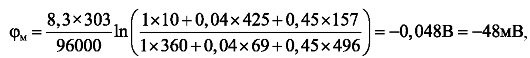 Что достаточно хорошо согласуется со значением, определенным опытным путем.
Что достаточно хорошо согласуется со значением, определенным опытным путем.
12.3. Потенциал действия и его распространение
Все живые клетки при действии различных раздражителей переходят в возбужденное состояние. При возбуждении разность потенциалов между клеткой и окружающей средой изменяется, появляется электрический импульс, возникает потенциал действия. Общее изменение разности потенциалов на мембране, происходящее при возбуждении клеток, определяет потенциал действия.
Потенциал действия - разность потенциалов между цитоплазмой и окружающей средой при возбуждении.
Механизм распространения потенциала действия по нервному волокну рассматривается в курсе нормальной физиологии. В данном курсе рассмотрим некоторые физические аспекты этого процесса, которые иллюстрирует рис. 12.1.
Распространение импульса определяется изменением состояния мембраны. В состоянии покоя (рис. 12.1, а) в результате активного транспорта концентрация ионов К+ в мембране значительно выше
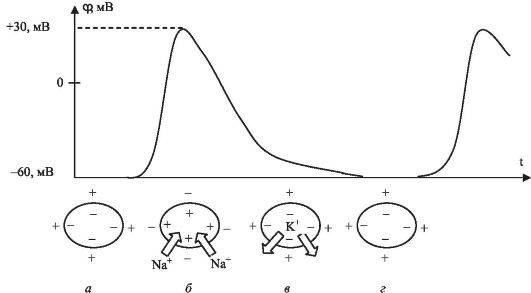 Рис. 12.1. Возникновение потенциала действия и деполяризация мембраны
Рис. 12.1. Возникновение потенциала действия и деполяризация мембраны
(в 36 раз), чем во внешней среде. Для ионов Na+ все наоборот (концентрация ионов Na+ снаружи мембраны в 6 раз выше, чем внутри). При этом на внутренней поверхности мембраны находится отрицательный заряд. При возбуждении будет происходить следующее.
1) В начале увеличивается проницаемость мембраны для ионов Na+.
Натриевые каналы открываются лишь при возбуждении. Ионы Na+ входят через мембрану внутрь клетки, в результате чего внутренняя поверхность мембраны изменяет свой заряд с «-» на «+», т.е. происходит деполяризация мембраны (рис. 12.1, б). Натриевый канал открыт малое время (0,5-1 мс). В течение этого времени происходит изменение мембранного потенциала от -60 до +30 мВ (генерация импульса величиной 90 мВ).
2) Во время генерации импульса натриевый канал закрывается и открывается калиевый канал. Ионы К+ частично выходят наружу (покидают клетку), что приводит к восстановлению отрицательного заряда на внутренней стороне мембраны (рис. 12.1, в). Во время импульса проводимость мембраны увеличивается в 1000 раз. Всего за время генерации одного импульса через квадратный микрон поверхности волокна проходит по 20 000 ионов натрия и калия.
3) Наступает рефрактерный период. Мембрана не воспринимает импульс, а возвращается в основное физиологическое состояние (рис. 12.1, г).
Таким образом, в мембране для ионов каждого вида (прежде всего для натрия и калия) имеется свой канал, пребывающий в открытом или в закрытом состоянии в зависимости от электрического потенциала мембраны. Благодаря такому устройству удается обеспечить быстрые потоки ионов через мембраны и, как следствие, быстрое изменение разности потенциалов между клеткой и окружающей средой.
Ионный канал впервые описал Р. Мак-Киннон. Канал состоит из внешней и внутренней полостей, разделенных фильтром (рис. 12.2).
Между внутренней полостью и телом клетки расположены ворота. Фильтр устроен хитро. Обычно ион калия или натрия движется по окружающей клетку среде в гидратированном виде: к нему присоединена «шуба» из четырех молекул воды. Фильтр же содержит
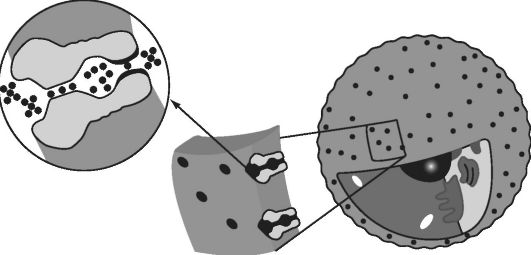 Рис. 12.2. Схема ионного канала (из нобелевской лекции Р. Мак-Киннона)
Рис. 12.2. Схема ионного канала (из нобелевской лекции Р. Мак-Киннона)
четыре атома кислорода, которые в случае, например, калиевого канала расположены на точно таком же расстоянии друг от друга, на каком расположены молекулы воды вокруг иона калия. Попав в фильтр, такой ион сменит шубу и даже этого не заметит. Если же ион будет другого размера, например маленький ион натрия, то пройти ему не удастся: водяная шуба не отцепится и будет мешать. Что же касается ворот, то они открываются в результате изменения конформации белка. Именно это изменение и происходит в ответ на изменение электрического потенциала мембраны.
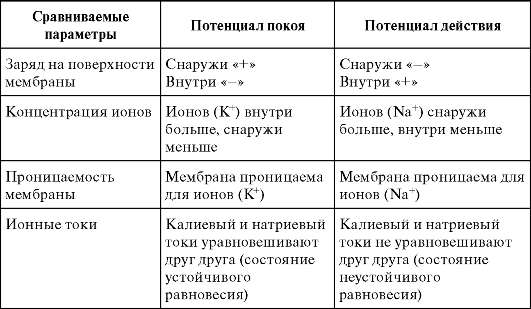
Сравнение потенциала покоя и потенциала действия
При возбуждении мембрана меняет избирательную проницаемость: из проницаемой главным образом для К+ мембрана становится проницаемой главным образом для А так как натрия снаружи больше, то он стремится внутрь и перезаряжает мембрану.
В таблице 12.1 приведено сравнение обоих потенциалов по некоторым факторам.
Таблица 12.1. Сравнение потенциала покоя и потенциала действия
 Распространение потенциала действия
Распространение потенциала действия
При деполяризации мембраны возникают токи, замыкающиеся через наружную проводящую среду. Между возбужденным и невозбужденным участками нервного волокна потечет электрический ток, так как у возбужденного участка внутренняя поверхность имеет положительный заряд, а у невозбужденного - отрицательный, и между ними возникнет разность потенциалов. Этот локальный ток служит раздражителем для невозбужденных участков нервного волокна, непосредственно примыкающих к месту деполяризации. В них также возникает возбуждение, т.е. потенциал действия (или деполяризация), и так далее. По поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении. Локальный ток, как и любой электрический ток, раздражает соседние невозбужденные участки и вызывает увеличение проницаемости мембраны. Это приводит к возникновению потенциалов действия в соседних участках. В то же время в ранее возбужденном участке происходят восстановительные процессы реполяризации. Вновь возбужденный участок, в свою очередь, становится электроотрицательным, и возникающий локальный ток раздражает следующий за ним участок. Этот процесс многократно повторяется
и обусловливает распространение импульсов возбуждения по всей длине клетки в обоих направлениях.
Процесс распространения потенциала действия происходит гораздо медленнее, чем течет локальный электрический ток.
У позвоночных животных повышение скорости распространения возбуждения достигается миелинизацией волокон (мякотные волокна). Волокно покрыто миелиновой оболочкой. Толщина оболочки составляет единицы мкм. Миелин - изолятор. Диффузия ионов через миелиновую оболочку невозможна. Поэтому в волокнах генерация потенциала действия сосредоточена только там, где миелиновая оболочка отсутствует. Эти места в мембране называются перехватами Ранвье (или активными узлами). В области перехвата мембрана контактирует с внеклеточным раствором. Длина перехвата составляет 2 мкм. От перехвата к перехвату (скачкообразно) нервные импульсы передаются через движение локальных токов. На долю перехватов приходится 0,02 % от общей длины нервного волокна.
На рисунке 12.3 показана схема распространения возбуждения по нервному волокну с перехватами Ранвье.
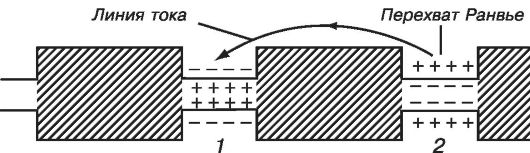 Рис. 12.3. Схема
скачкообразного проведения импульса. Ток идет между невозбужденным
перехватом 2 и возбужденным перехватом 1, но практически не течет через
миелин
Рис. 12.3. Схема
скачкообразного проведения импульса. Ток идет между невозбужденным
перехватом 2 и возбужденным перехватом 1, но практически не течет через
миелин
Миелиновая изоляция имеет большое сопротивление - в сотни раз выше, чем сопротивление мембраны аксона кальмара, а емкость - в сотни раз меньшую. В результате получается довольно хороший «кабель», а перехваты с каналами и насосами играют роль источника тока. Если возбудить один перехват, то генерируемый им ток почти без потерь достигнет следующего перехвата. Ток, подошедший к другому перехвату, возбуждает его, вызывает появление в этом месте потенциала действия, и процесс распространяется по всему волокну. Такое проведение возбуждения называют «прыгающим». Импульс
быстро перепрыгивает от одного перехвата к другому, затрачивая на распространение между перехватами только несколько сотых долей миллисекунды (50-70 мкс). Затраты энергии при таком распространении сигнала значительно меньше, чем по немиелинизированному волокну, так как общее количество ионов натрия, проходящих через мембрану в области перехватов, значительно меньше, чем если бы они проходили через всю поверхность мембраны.
Нарушение миелиновой оболочки ведет к нарушению распространения потенциала действия по нервному волокну (тяжелые нервные заболевания). Изучение особенностей распространения биопотенциала безусловно важно для многих направлений медицины.
Скорость распространения возбуждения по гладким немиелинизированным нервным волокнам пропорциональна квадратному корню из их радиуса v ~ √R . Поэтому головоногие моллюски пошли по пути увеличения радиуса нервного волокна, создав гигантские аксоны.
У беспозвоночных скорость распространения потенциала действия составляет 20-30 м/с.
Распространение потенциала действия по нервному волокну называется волной возбуждения. Эта волна не затухает, так как получает энергию из среды - от заряженной мембраны. Волна возбуждения является автоволной в активной среде возбудимых клеток.
12.4. Основные понятия и формулы
 Окончание таблицы
Окончание таблицы
 12.5. Задачи
12.5. Задачи
1. Рассчитать для равновесного состояния мембранный потенциал, создаваемый ионами K+, пренебречь проницаемостью ионов Na+ и С1-.
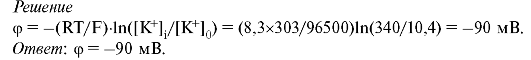 2. За какое время потенциал действия распространится на расстояние, равное S = 10 см, если принять скорость его распространения v = 20 м/с?
2. За какое время потенциал действия распространится на расстояние, равное S = 10 см, если принять скорость его распространения v = 20 м/с?
Решение
t = S/v = 0,1/20 = 5х10-3 с. Ответ: t = 5х10-3 с.
3. Принимая Сi/Со = 100, температуру Т = 300 К, найти потенциал покоя для одновалентного иона (|Z| = 1).
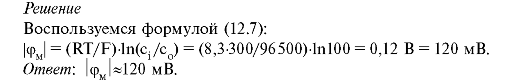 4. Определить,
какое количество одновалентных ионов должно перейти из цитоплазмы в
неклеточную среду для создания потенциала покоя φм ≈ 120 мВ? Принять площадь поверхности клетки S = 10-9 м2; удельную электроемкость мембраны (на единицу площади) Суд = 10-2Ф/м2.
4. Определить,
какое количество одновалентных ионов должно перейти из цитоплазмы в
неклеточную среду для создания потенциала покоя φм ≈ 120 мВ? Принять площадь поверхности клетки S = 10-9 м2; удельную электроемкость мембраны (на единицу площади) Суд = 10-2Ф/м2.
 5. Биологические
мембраны находятся под действием электрического поля, созданного за
счет различных концентраций заряженных ионов (К+ и Na+) по разные ее стороны. Разность потенциалов между цитоплазмой и внеклеточной средой достигает Δφ = 100 мВ = 10-1 В, а толщина мембраны h = 10 нм = 10-8 м. Определить: а) напряженность электрического поля в мембране; б) к чему ведет уменьшение толщины мембраны? Решение
5. Биологические
мембраны находятся под действием электрического поля, созданного за
счет различных концентраций заряженных ионов (К+ и Na+) по разные ее стороны. Разность потенциалов между цитоплазмой и внеклеточной средой достигает Δφ = 100 мВ = 10-1 В, а толщина мембраны h = 10 нм = 10-8 м. Определить: а) напряженность электрического поля в мембране; б) к чему ведет уменьшение толщины мембраны? Решение
а) Е = Δφ/h = 107 В/м.
б) даже небольшое уменьшение толщины мембраны приводит к увеличению напряженности Е и может вызвать электрический пробой, т.е. повреждение мембраны и гибель клетки. Таков механизм развития различных патологий, затрагивающих мембраны (при перикисном окислении липидов под действием радиации, УФ-облучения, вследствие канцерогенеза и т.д.).
Ответ: а) Е = 107 В/м.