Урология : учебник / Б. К. Комяков. - 2012. - 464 с.: ил.
|
|
|
|
Глава 2. АНАТОМИЯ И ФИЗИОЛОГИЯ МОЧЕПОЛОВЫХ ОРГАНОВ
Мочеполовой аппарат (apparatus urogenitalis) включает в себе мочевые (organa urinaria) и половые (organa genitalia) органы. Органы эти тесно связаны друг с другом по своему развитию и анатомо-функциональному состоянию, чем и обусловлено их объединение под названием «мочеполовой аппарат».
2.1. АНАТОМИЯ ПОЧЕК И МОЧЕВЫВОДЯЩИХ ПУТЕЙ
Почки (ren, греч. - nephros) - парный орган, располагающийся в забрюшин-ном пространстве, в поясничной области, по бокам от позвоночника. Правая почка находится на уровне Th XII-L III; левая - на уровне Th XI-L II. Правая почка лежит ниже левой: XII ребро пересекает ее на границе средней и верхней трети, левую почку - приблизительно посередине. Размеры почки - 10 12 х х 5 - 6 х 4 см, масса 180-200 г (рис. 2.1).
Почка делится на сегменты, что связано с особенностями ветвления почечной артерии. Следует выделить следующие сегменты:
■ верхний сегмент (segmentum superius);
■ верхний передний сегмент (segmen-tum superius anterius);
■ нижний сегмент (segmentum inferius);
■ нижний передний сегмент (segmentum inferius anterius);
■ задний сегмент (segmentum posterius).
Ворота почки (hilium renis) - место проникновения в почку элементов почечной ножки. Снаружи почка покрыта фиброзной капсулой (capsula fibrosa), которая рыхло связана с паренхимой. Далее ее окружает жировая капсула, пред- и позадипочечные фасции (Герота). К верхнему полюсу правой почки прилежит правый надпочечник, передняя поверхность соприкасается с печенью и правым изгибом ободочной кишки; вдоль медиального края проходит нисходящая
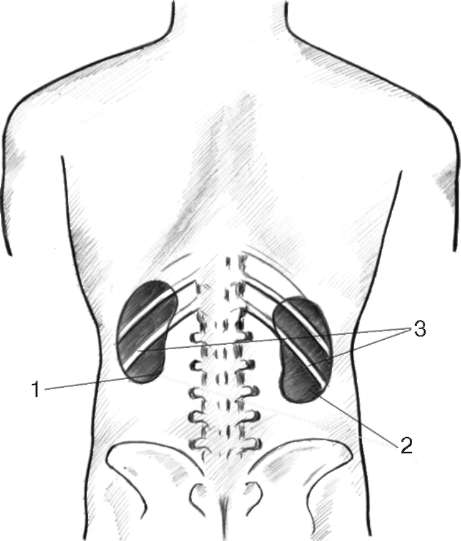
Рис. 2.1. Скелетотопия почек: 1 - левая почка; 2 - правая почка; 3 - XII ребро
часть двенадцатиперстной кишки. К верхнему полюсу левой почки прилежит левый надпочечник; передняя поверхность почки соприкасается с желудком, поджелудочной железой, левым изгибом ободочной кишки и начальной частью нисходящей ободочной кишки, ниже - с петлями тощей кишки; к латеральному краю прилежит селезенка.
Почка состоит из коркового (cortex renis) и мозгового (medulla renis) вещества. Корковое вещество расположено по периферии и между пирамидами (columnae renalis, c. Bertinii), мозговое вещество расположено в центре и представлено пирамидами (piramides renalis, p. Malpigii).
Кровоснабжение почки осуществляется за счет почечной артерии (a. renalis), которая делится на предлоханочную и позадилоханочную ветви; последняя питает задний сегмент почки.
Отток венозной крови происходит по одноименным венам в почечную (v. renalis) и нижнюю полую вену (v. cava inferior).
По ходу органа нервные волокна формируют почечное сплетение (plexsus renalis). Афферентная иннервация обеспечивается чувствительными волокнами передних ветвей нижних грудных и верхних поясничных спинномозговых
нервов, а также волокнами почечных ветвей блуждающего нерва (rr. renales n. vagi). Парасимпатическая иннервация происходит от волокон rr. renales n. vagi, а симпатическая образуется от ganglia aortorenalia из plexus coeliacus (plexsus aorticus abdominalis) по ходу почечных артерий.
Лимфа оттекает преимущественно в nodi lymphatici lumbales, aortici laterals, cavales laterals, coeliaci, iliaci interni, phrenici inferiors.
Почечные чашки и почечная лоханка . Главные собирательные структуры мочевых путей начинаются от почечных сосочков, из которых моча поступает в малые чашки. Число малых чашек составляет от 7 до 13. Каждая малая чашка охватывает от одного до трех сосочков. Малые чашки объединяются в две-три большие чашки, последние соединяются, образуя воронкообразную почечную лоханку.
Мочеточник (ureter) - парный трубчатый орган, обеспечивающий проведение мочи из почечной лоханки в мочевой пузырь, располагается в забрюшинном пространстве. Его длина составляет 25-30 см.
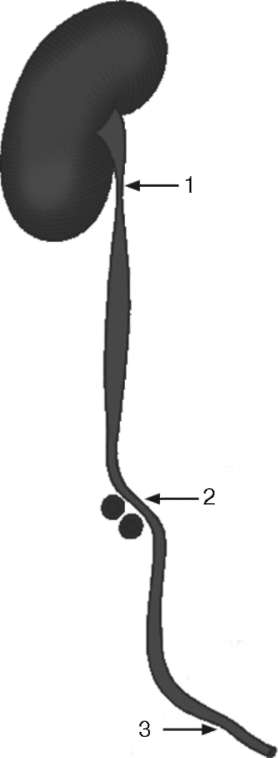
Рис. 2.2. Физиологические сужения мочеточника:
1 - пиелоуретральный сегмент; 2 - перекрест с подвздошными сосудами; 3 - пу-зырно-мочеточниковый сегмент
Мочеточник имеет три участка сужения: в лоханочно-мочеточниковом сегменте; в месте пересечения с подвздошными сосудами; в пузырно-мочеточниковом сегменте (рис. 2.2)
В воротах почки мочеточник располагается позади почечных сосудов, затем опускается по большой поясничной мышце, входит в малый таз, пересекая при этом спереди подвздошные сосуды (справа a. et. v. internae, слева a. et. v. iliacae communes). Затем мочеточник проходит по стенкам малого таза, направляясь к дну мочевого пузыря. У мужчин он перекрещивается с семявыносящими протоками, у женщин мочеточники проходят позади яичников, латеральнее шейки матки.
Различают следующие части мочеточников:
■ брюшная (pars abdominalis);
■ тазовая (pars pelvina);
■ внутристеночная (pars intramuralis), расположена в стенке мочевого пузыря. В клинической практике используется деление мочеточника по длине на
три части: верхняя, средняя и нижняя треть.
Стенка мочеточника состоит из трех слоев. Мочеточник окружен промежуточным слоем забрюшинной соединительной ткани (околомочеточниковой клетчаткой), которая, уплотняясь, образует для него футляр. Внутренняя часть стенки мочеточника - это слизистая оболочка, покрытая переходным многослойным эпителием. Основную толщину стенки мочеточника составляет мышечный слой, который, как принято считать, состоит из внутреннего продольного и внешнего циркулярного слоев. Четкой границы между ними нет, так как оба они идут под углом и проникают друг в друга. В терминальной части мочеточника мышечные волокна в основном имеют продольное направление. В пу-зырно-мочеточниковом сегменте мышечные волокна мочевого пузыря распространяются на мочеточник и отделяются от его мышц рыхлой соединительной тканью, которая известна как футляр Вальдейера.
Кровоснабжение мочеточника происходит из нескольких смежных сосудистых структур. Его верхняя часть, почечные чашечки и почечная лоханка снабжается кровью из почечной артерии. Средняя часть получает кровь от яичковых артерий. Дистальный отдел мочеточника кровоснабжается сосудами, отходящими от бифуркации аорты, а также от общей подвздошной артерии, внутренней подвздошной артерии и мочеточниковых ветвей верхней и нижней артерий мочевого пузыря. У женщин кровь к мочеточнику поступает из маточной артерии. Одноименные вены сопровождают артерии.
Мочеточник получает вегетативную иннервацию от нижнего брыжеечного, яичкового и тазового сплетений. Афферентные волокна, иннервирующие его, проходят в составе нервов Th XI - Th XII и L I. Нервы в основном идут по ходу кровеносных сосудов мочеточника. Лимфатические сосуды мочеточника обычно сопровождают артерии и впадают в лимфоузлы, прилежащие к верхней части почечной артерии. Из средней части мочеточника лимфа поступает в аортальные узлы, а из дистальной - во внутренние подвздошные.
Мочевой пузырь (vesica urinaria, греч. - cistis) - непарный полый мышечный орган, служащий для накопления и выведения мочи. Он имеет форму четырехгранника, но после заполнения становится сферическим. Мочевой пузырь
находится в полости малого таза, спереди располагается лобковый симфиз. Не-наполненный мочевой пузырь не выступает выше лонного сочленения, сильно наполненный - поднимается над ним. У мужчин к мочевому пузырю сзади прилежат прямая кишка, семенные пузырьки и ампулы семявыносящих протоков; сверху - петли тонкой кишки; дно соприкасается с предстательной железой. У женщин сзади к нему прилежат шейка матки и влагалище; сверху - тело и дно матки; дно мочевого пузыря расположено на мочеполовой диафрагме.
Мочевой пузырь имеет четыре поверхности: верхнюю, две нижнебоковые и заднюю, или дно (fundus vesicae). Сверху он покрыт брюшиной, пустой мочевой пузырь лежит экстраперитонеально, в наполненном состоянии - мезоперитоне-ально. Пространство между передней поверхностью мочевого пузыря и лобком называется предпузырным пространством (spatium prevesicale), или пространством Ретциуса. У мочевого пузыря имеются верхушка (apex vesicae) - суженная передневерхняя часть, тело (corpus vesicae) - средняя часть, дно - нижняя, несколько расширенная часть, шейка мочевого пузыря (cervix vesicae) располагается у места перехода его в мочеиспускательный канал (здесь находится внутреннее отверстие мочеиспускательного канала). Мышечная оболочка мочевого пузыря, за исключением сфинктера, в целом формирует мышцу, выталкивающую мочу (m. detrusor vesicae), и состоит из тех мышечных слоев: наружного продольного, среднего циркулярного и внутреннего продольного. Изнутри мышечный слой мочевого пузыря покрыт хорошо развитой слизистой, состоящей из переходного эпителия (рис. 11, см. цв. вклейку). На дне мочевого пузыря расположен треугольник (треугольник Льето). Его вершины - это устья мочеточников (рис. 12, см. цв. вклейку), основание образует межмочеточниковую складку; в треугольнике Льето отсутствуют складки слизистой оболочки.
Основное кровоснабжение мочевой пузырь получает из внутренней подвздошной артерии, дополнительное - из нижней и верхней пузырных артерий. У женщин в кровоснабжении мочевого пузыря участвуют также маточные и влагалищные артерии. Вены не сопровождают артерии, а формируют сложное сплетение, сосредоточенное в основном на нижней поверхности и в области дна мочевого пузыря. Венозные стволы впадают во внутренние подвздошные вены.
Мочевой пузырь иннервируется мочепузырным сплетением (plexus vesikalis) - частью тазового сплетения, которое находится на боковых поверхностях прямой кишки. Симпатические волокна берут начало в сегментах Th X-L XII спинного мозга. Парасимпатические волокна идут от сегментов S II-S IV и в составе тазовых нервов достигают тазового сплетения. Иннервация детру-зора преимущественно парасимпатическая, тогда как шейку мочевого пузыря у мужчин иннервируют симпатические, а у женщин парасимпатические нервы. К сфинктеру мочеиспускательного канала подходят волокна тазовых внутренностных нервов.
Лимфа оттекает преимущественно в nodi limphatica paravesicales, pararectales, lumbales, iliaci interni.
Женский мочеиспускательный канал (urethra feminina) начинается от мочевого пузыря внутренним отверстием (ostium urethrae internum) и представляет собой трубку длиной 3-3,5 см, слегка изогнутую выпуклостью кзади и огибающую снизу и сзади нижний край лобкового симфиза. Вне периода прохожде-
ния мочи через канал передняя и задняя стенки его прилежат одна к другой, но стенки канала отличаются значительной растяжимостью, и просвет его может быть растянут до 7-8 мм. Задняя стенка канала тесно соединяется с передней стенкой влагалища. При выходе из таза канал прободает мочеполовую диафрагму (diaphragma urogenitale) с ее фасциями и окружен исчерченными произвольными мышечными волокнами сфинктера (m. sphincter urethrae).
Наружное отверстие женской уретры (ostium urethrae externum) открывается в преддверие влагалища впереди и выше отверстия влагалища и представляет собой узкое место канала. Стенка женского мочеиспускательного канала состоит из оболочек мышечной, подслизистой и слизистой. В рыхлом подслизистом слое (tela submucosa), проникая также в мышечную оболочку (tunica muscularis), находится сосудистое сплетение, придающее ткани на разрезе пещеристый вид. Слизистая оболочка (tunica mucosa) образует продольные складки. В канал открываются, особенно в нижних частях, многочисленные слизистые железки (glandulae urethrales).
Кровоснабжение женский мочеиспускательный канал получает из a. vesicalis inferior и a. pudenda interna. Вены вливаются через венозное сплетение (plexus venosus vesicalis) в v. iliaca interna. Лимфатические сосуды из верхних отделов канала направляются к Inn. iliaci, из нижних - к Inn. inguinales.
Иннервация: из plexus hypogastrics inferior, nn. splanchnici pelvini и n. pudendus.
Мужской мочеиспускательный канал проводит не только мочу, но и сперму, поэтому он будет рассмотрен вместе с мужской половой системой.
2.2. АНАТОМИЯ МУЖСКИХ ПОЛОВЫХ ОРГАНОВ
К мужским половым органам (рис. 2.3) (organa genitalia masculina) относятся яички с их оболочками, семявыносящие протоки с семенными пузырьками, предстательная железа, бульбоуретральные железы, половой член и мужской мочеиспускательный канал, носящий смешанный характер мочеполовой трубки.
Яички (testes) представляют собой два тела овальной формы, несколько сплющенных с боков, расположенных в мошонке. Длинник яичка равен в среднем 4 см, поперечник - 3 см, масса составляет от 15 до 25 г. Левое яичко обычно опущено несколько ниже, чем правое. К заднему краю яичка подходят семенной канатик (funiculus spermaticus) и придаток яичка (epididymis); последний располагается вдоль заднего края. Epididymis представляет собой узкое длинное образование, в котором различают верхнюю, несколько утолщенную часть - головку придатка (caput epididymidis) и нижний, более заостренный конец - хвост (cauda epididymidis); промежуточный участок составляет тело (corpus epididymidis).
Яичко окружено плотной фиброзной оболочкой беловатой окраски (tunica albuginea), лежащей непосредственно на его паренхиме. По заднему краю яичка фиброзная ткань оболочки вдается неглубоко внутрь железистой ткани в виде неполной вертикальной перегородки, или утолщения (mediastinum testis); от него лучеобразно отходят фиброзные перегородочки (septula testis), которые своими наружными концами прикрепляются к внутренней поверхности (tunicae alhugineae) и таким образом делят всю паренхиму яичка на дольки (lobuli testis).
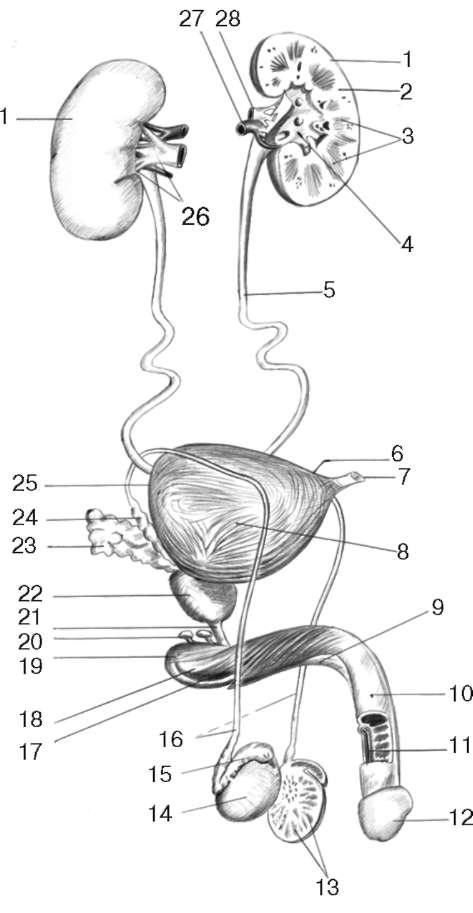
Рис. 2.3. Мочеполовые органы мужчины: 1 - почка; 2 - корковое вещество почки; 3 - почечные пирамиды; 4 - почечная лоханка; 5 - мочеточник; 6 - верхушка мочевого пузыря; 7 - срединная пупочная связка; 8 - тело мочевого пузыря; 9 - тело полового члена; 10 - спинка полового члена; 11 - губчатая часть мочеиспускательного канала; 12 - головка полового члена; 13 - дольки яичка; 14 - яичко; 15 - придаток яичка; 16 - семявыносящие протоки; 17 - луковично-пещеристая мышца; 18 - корень полового члена; 19 - луковично-губчатая мышца; 20 - бульбоуретральная железа; 21 - перепончатая часть мочеиспускательного канала; 22 - предстательная железа; 23 - семенной пузырек; 24 - ампула семявыносящего протока; 25 - дно мочевого пузыря; 26 - почечные ворота; 27 - почечная артерия; 28 - почечная вена
Число долек яичка доходит до 250- 300. Верхушки долек обращены к перегородке яичка (mediastinum testis), а основания - к белочной оболочке (tunica albuginea). Придаток яичка также имеет tunica albuginea, но более тонкую.
Паренхима яичка состоит из семенных канальцев. Различают извитые (tubuli seminiferi contorti) и прямые (tubuli seminiferi recti) семенные канальцы. В каждой дольке имеются 2-3 канальца и более. Имея извилистое направление в самой дольке (tubuli seminiferi contorti), приближаясь к перегородке яичка (mediastinum testis), извитые канальцы соединяются друг с другом и непосредственно у mediastinum суживаются в короткие прямые семенные канальцы. Прямые канальцы открываются в сеть яичка (rete testis), расположенную в толще mediastinum. Из сети яичка открываются 12-15 выносящих канальцев (ductuli efferentes testis), которые направляются к головке придатка. По выходе из яичка выносящие канальцы становятся извилистыми и образуют ряд конических долек придатка (lobulis coni epididymidis). Ductuli efferentes открываются в одиночный канал придатка (ductus epididymidis), который, образуя многочисленные изгибы, продолжается в семявыно-сящий проток (ductus deferens). Семя-выносящий проток (ductus defferentes), дольки придатков (lobuli epididymidis) и его начальный отдел образуют в совокупности головку придатка.
Местом образования сперматозоидов - основной части спермы (sperma) - являются лишь tubuli seminiferi contorti. Tubuli recti и канальцы сети яичка принадлежат уже к семевыносящим путям. Жидкая составная часть спермы только в не-
значительном количестве продуцируется яичками, так как представляет собой главным образом продукт выделения придаточных желез полового аппарата, открывающихся в выводящие пути.
Артериями, питающими яичко и придаток, являются a. testicularis, a. ductus deferentis и отчасти a. cremasterica. Венозная кровь оттекает из testis и epididymis в лозовидное сплетение (plexus pampiniformis) и далее в яичковую вену (v. testicularis).
Лимфатические сосуды от яичка идут в составе семенного канатика и, минуя паховые узлы, оканчиваются в поясничных лимфатических узлах (nody lymphatici lumbales). Это, так же как высокое положение а. и v. testiculares, связано с закладкой яичка в поясничной области. Нервы яичка образуют симпатические сплетения plexus testicularis и plexus deferentialis вокруг одноименных артерий.
Семявыносящие протоки (ductus deferens) являются непосредственным продолжением канала придатка и отличаются от последнего большей толщиной стенок. Отделенный от яичка сосудами (а. и v. testiculares), семявыносящий проток поднимается кверху и входит в состав семенного канатика.
В составе семенного канатика ductus deferens поднимается вертикально вверх к поверхностному паховому кольцу. Пройдя в паховом канале косо вверх и ла-терально, он у глубокого пахового кольца оставляет vasa testiculares (последние направляются в поясничную область) и идет вниз и назад по боковой стенке таза, будучи прикрыт брюшиной. Достигнув мочевого пузыря, проток загибается к его дну и подходит к предстательной железе. В нижнем своем отделе он заметно расширяется в виде ампулы семявыносящего протока (ampulla ductus deferentis). Длина ductus deferens равняется 40-45 см, средний диаметр - 2,5 мм, ширина просвета - всего 0,2-0,5 мм. Стенка ductus deferens состоит из трех слоев: наружной фиброзной, средней мышечной и внутренней слизистой оболочки.
Семенные пузырьки (vesiculae seminales) лежат латерально от семявыносящих протоков, между дном мочевого пузыря и прямой кишкой. Каждый семенной пузырек имеет в расправленном виде длину до 12 см, в нерасправленном - 5 см. Нижний заостренный конец семенного пузырька переходит в узкий выделительный проток (ductus excretorius), который соединяется под острым углом с ductus deferens той же стороны, образуя вместе с ним семявыбрасывающий проток (ductus ejaculatorius). Последний представляет собой тоненький каналец длиной около 2 см, который, начавшись от места слияния ductus deferens и ductus excretorius, проходит через толщу предстательной железы и открывается в предстательную часть мочеиспускательного канала узким отверстием у основания семенного бугорка.
Стенки семенных пузырьков состоят из тех же слоев, что и ductus deferens. Семенные пузырьки представляют собой секреторные органы, которые вырабатывают жидкую часть спермы.
Сосуды и нервы: ductus deferens получает питание из a. ductus deferenis (ветвь a. iliaca interna), семенные пузырьки - из аа. vesicalis inferior, ductus deferentis, rectales. Венозный отток происходит по v. deferentialis, которая впадает в v. iliaca interna. Отток лимфы происходит в наружные, внутренние подвздошные и крестцовые лимфатические узлы. Ductus deferens и семенные пузырьки иннервиру-ются plexus deferentialis, образованным нервами из plexus hypogastricus inferior.
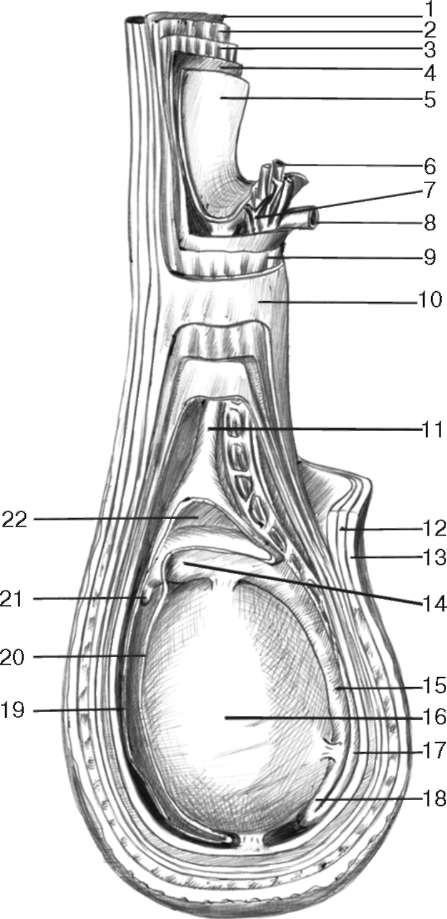
Рис. 2.4. Оболочки яичка и семенного канатика:
1 - наружная косая мышца живота; 2 - внутренняя косая мышца живота; 3 - поперечная мышца живота; 4 - поперечная фасция; 5 - брюшина; 6 - яичковая артерия; 7 - лозовидное сплетение (венозное); 8 - семявыносящий проток; 9 - внутренняя семенная фасция; 10 - наружная семенная фасция; 11 - влагалищный отросток брюшины; 12 - мясистая оболочка; 13 - кожа; 14 - головка придатка яичка; 15 - тело придатка яичка; 16 - яичко; 17 - семявыносящий проток; 18 - хвост придатка яичка; 19 - влагалищная оболочка яичка (пристеночный листок); 20 - влагалищная оболочка яичка (висцеральный листок); 21 - привесок придатка яичка; 22 - серозная полость яичка
Семенной канатик (funiculus spermatacus) - круглый тяж длиной 16-20 см, покрытый наружной семенной фасцией и расположенный между внутренним паховым кольцом и верхним полюсом яичка. В состав его входят семявыносящий проток, яичковая артерия, артерия семявыно-сящего протока, лазовидное венозное сплетение, лимфатические сосуды яичка и его придатка, нервы и влагалищный отток брюшины.
Мошонка (scrotum) является обособленным выпячиванием передней брюшной стенки. Она состоит из двух раздельных камер, в которых располагаются яички, их придатки и часть семенного канатика.
Оболочки яичка и семенного канатика, считая снаружи, следующие: кожа (cutis), мясистая оболочка (tunica dartos), наружная семенная фасция (fascia spermatica externa) фасция мышцы, поднимающей яичко (fascia cremasterica), мышца, поднимающая яичко (m. cremaster), внутренняя семенная фасция (fascia spermatica interna), влагалищная оболочка яичка (tunica vaginalis testis) (рис. 2.4). Такое большое число оболочек яичка соответствует определенным слоям передней брюшной стенки. Яичко при смещении из брюшной полости как бы увлекает за собой брюшину и фасции мышц живота и оказывается окутанным ими. В результате в соответствии со строением слоев передней брюшной стенки образуются следующие оболочки яичка.
1. Кожа мошонки тонкая и имеет более темную окраску по сравнению с другими участками тела. Она снабжена многочисленными крупными сальными железами, секрет которых имеет характерный запах.
2. Мясистая оболочка расположена тотчас под кожей. Она представляет собой продолжение подкожной соединительной ткани из паховой области и промежности, но лишена жира. В ней находится значительное количество гладкой мышечной ткани. Tunica dartos образует для каждого яичка по одному отдельному мешку, которые соединяются по средней линии так, что получается перегородка (septum scroti), прикрепляющаяся по линии raphe.
3. Наружная семенная фасция является продолжением поверхностной фасции живота.
4. Фасция мышцы, поднимающей яичко, представляет собой продолжение fasciae intercruralis, отходящей от краев поверхностного пахового кольца; она покрывает m. cremaster, поэтому и называется fascia cremasterica.
5. Мышца, поднимающая яичко, состоит из пучков исчерченных мышечных волокон, являющихся продолжением поперечной мышцы живота (т. transversus abdominis). При сокращении m. cremaster яичко подтягивается кверху.
6. Внутренняя семенная фасция расположена тотчас под m. cremaster. Она представляет собой продолжение fasciae transversalis, охватывает кругом все составные части семенного канатика и в области яичка прилежит к наружной поверхности его серозного покрова.
7. Влагалищная оболочка яичка образуется из влагалищного отростка брюшины (processus vaginalis) и формирует замкнутый серозный мешок, состоящий из двух пластинок - пристеночной (laminaparietalis) и висцеральной (lamina visceralis). Между ними в патологических случаях может скопиться большое количество серозной жидкости и образоваться водянка яичка (см. главу 14.3).
Половой член (penis) составляет вместе с мошонкой наружные половые органы. Он состоит из трех тел: парного пещеристого (corpus cavernosum penis) и непарного губчатого (corpus spongiosum penis). Название этих тел обусловлено тем, что они состоят из многочисленных перекладин, фиброзно-эластических тяжей с примесью неисчерченных мышечных волокон, среди густого сплетения которых есть промежутки - пещеры, выстланные эндотелием и заполненные кровью (рис. 2.5).
Corpora cavernosa penis представляет собой два длинных цилиндрических тела с заостренными концами, из которых задние расходятся и образуют ножки полового члена (crura penis), прикрепляющиеся к нижним ветвям лобковых костей. Corpus spongiosum penis, покрытое белочной оболочкой (tunica albuginea corporis spongiosi), лежит под пещеристыми телами члена и пронизано во всю длину мочеиспускательным каналом. Оно имеет меньший, чем два других пещеристых тела, диаметр (1 см), но в отличие от них утолщается на обоих концах, образуя спереди головку члена (glans penis), а сзади луковицу (bulbus penis).
Задняя часть полового члена, прикрепленная к лобковым костям, носит название корня (radix penis). Кпереди половой член оканчивается головкой. Промежуточная между головкой и корнем часть называется телом (corpus penis). Верхняя поверхность тела шире нижней и носит название спинки (dorsum penis). К нижней поверхности прилежит corpus spongiosum penis. На головке члена имеется вертикальная щель - наружное отверстие мочеиспускательного канала (ostium urethra externum); головка с дорсальной и латеральной сторон несколько выдается
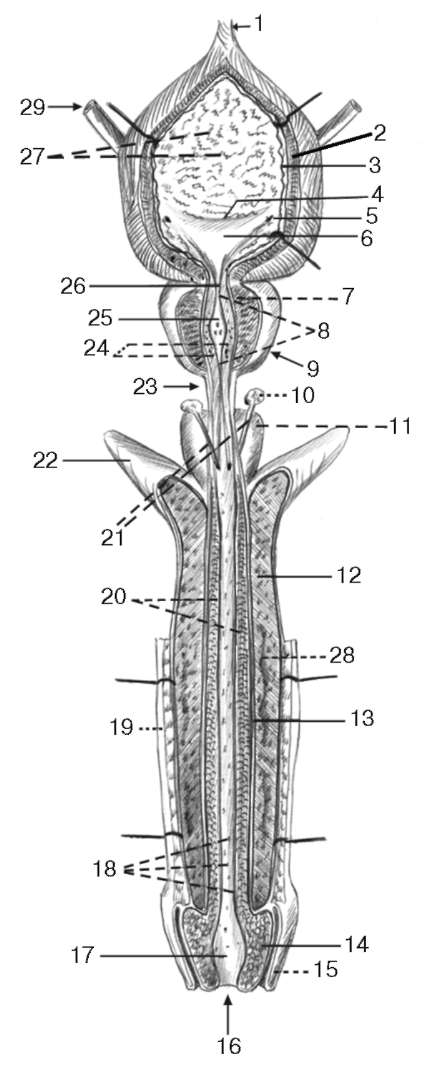
Рис. 2.5. Мочевой пузырь, мужской мочеиспускательный канал, предстательная железа, пещеристые и губчатое тела полового члена:
1 - срединная пупочная связка; 2 - мышечная оболочка мочевого пузыря; 3 - подслизистая основа; 4 - межмочеточниковая складка; 5 - отверстие мочеточника; 6 - треугольник мочевого пузыря; 7 - мышечное вещество предстательной железы; 8 - предстательная часть мочеиспускательного канала; 9 - предстательная железа; 10 - бульбоуретральные железы; 11 - луковица полового члена; 12 - пещеристые тела полового члена; 13 - белочная оболочка пещеристых тел; 14 - головка полового члена; 15 - крайняя плоть; 16 - наружное отверстие мужского мочеиспускательного канала; 17 - ладьевидная ямка мочеиспускательного канала; 18 - лакуны или крипты мочеиспускательного канала; 19 - кожа; 20 - губчатое тело полового члена; 21 - выводной проток бульбоуретральной железы; 22 - ножка полового члена; 23 - перепончатая часть мочеиспускательного канала; 24 - проток предстательной железы;
25 - семенной бугорок предстательной железы;
26 - внутреннее отверстие мочеиспускательного канала; 27 - складки слизистой; 28 - глубокая артерия полового члена; 29 - правый мочеточник
над уровнем пещеристых тел, этот край головки носит название corona glandis, а сужение позади него - collum glandis.
Кожа полового члена у основания головки образует свободную складку, которая носит название крайней плоти (preputium). На нижней стороне головки члена крайняя плоть соединена с кожей головки уздечкой (frenulum preputii). Вокруг corona glandis и на внутреннем листке крайней плоти расположены различной величины сальные железки (glandulae preputiales). Секрет этих желез входит в состав препуциальной смазки (smegma preputii), собирающейся в пространстве между головкой и крайней плотью - полости крайней плоти, открывающейся спереди отверстием, которое пропускает головку при отодвигании крайней плоти кзади.
Величина полового члена зависит от количества крови в камерах пещеристых и губчатого тел. Кровь приносится к половому члену через аа. profundae et
dorsalis penis. Артериальные ветви, проходя в соединительнотканных перегородках, распадаются на тонкие завитковые артерии, которые открываются прямо в кавернозные пространства. Отводящие от пещеристых тел кровь вены (venae cavernosae) вливаются в vv. profundae penis и в v. dorsalis penis. Благодаря особому устройству кровеносных сосудов члена кровь в пещеристых телах может задерживаться, что приводит к их уплотнению при эрекции.
Артерии полового члена являются ветвями a. femoralis и a. pudenda interna. Венозный отток происходит по vv. dorsales penis superficialis et profundae в v. femoralis и в plexus venosus vesicalis. Лимфоотток осуществляется в Inn. lymphoidi inguinales и узлы полости малого таза.
Афферентная иннервация осуществляется n. pudendus, эфферентная симпатическая - из plexus hypogastrics inferior, парасимпатическая - nn. erigentes.
Мужской мочеиспускательный канал (urethra masculina) представляет собой трубку длиной 16-22 см и диаметром 0,5-0,7 см, простирающуюся от мочевого пузыря до наружного отверстия мочеиспускательного канала на головке полового члена (см. рис. 2.5). Urethra служит не только для выведения мочи, но также для прохождения спермы, которая поступает в мочеиспускательный канал через ductus ejaculatorius. Мочеиспускательный канал проходит через различные образования, поэтому в нем различают три части: pars prostatica, pars membranacea и pars spongiosa.
Предстательная часть (pars prostatica), ближайшая к мочевому пузырю, проходит через предстательную железу (см. рис. 2.5). Длина этой части около 2,5 см. Предстательная часть, особенно ее средний отдел, является наиболее широким и растяжимым участком мочеиспускательного канала. На задней стенке находится небольшое срединное возвышение - семенной бугорок (colliculus seminalis) (рис. 2, см. цв. вклейку).
По окружности предстательной части мочеиспускательного канала имеется кольцо мышечных волокон, составляющих часть гладкой мышечной ткани предстательной железы, выполняющее функцию третьего (непроизвольного) сфинктера мочеиспускательного канала.
Перепончатая часть (pars membranacea) представляет собой участок мочеиспускательного канала на протяжении от верхушки предстательной железы до bulbus penis, длина ее около 1 см. Таким образом, этот отдел канала является наиболее коротким и в то же время наиболее узким из всех трех. Он лежит кзади и книзу от дугообразной связки лона (lig. arcuatum pubis), прободая на своем пути diaphragma urogenitale с ее верхней и нижней фасциями; нижний конец перепончатой части на месте прободения нижней фасции представляет собой самый узкий и наименее растяжимый участок канала, что необходимо учитывать при введении катетера, чтобы не повредить канал. Перепончатая часть мочеиспускательного канала окружена мышечными пучками произвольного сфинктера (m. sphincter urethrae).
Губчатая часть (pars spongiosa) длиной около 15 см окружена тканью corpus spongiosum penis. Часть мочеиспускательного канала соответственно bulbus penis несколько расширена; на остальном протяжении до головки диаметр канала равномерный, в головке на протяжении приблизительно 1 см он опять расширяется, образуя ладьевидную ямку уретры (fossa navicularis urethrae). Наружное
отверстие - малорастяжимая часть мочеиспускательного канала, что следует учитывать при катетеризации.
Кроме анатомического деления мочеиспускательного канала на три части, в урологической практике (соответственно течению воспалительных процессов) используется разделение его на два отдела: переднюю уретру (pars spongiosa) (рис. 1, см. цв. вклейку) и заднюю - остальные две части (рис. 2, см. цв. вклейку). Границей между ними служит сфинктер мочеиспускательного канала, который препятствует проникновению инфекции из передней уретры в заднюю.
На всем протяжении слизистой оболочки, за исключением ближайшего к наружному отверстию участка, в канал открываются многочисленные железки (glandulae urethrales, старое название - glandulae littrei, откуда название воспаления этих железок - литтреит). Кроме того, преимущественно на верхней стенке мочеиспускательного канала, в особенности кпереди от луковицы, находятся небольшие углубления - лакуны мочеиспускательного канала (lacunae urethrales); отверстия их обращены кпереди и прикрыты клапанооб-разными заслонками. Кнаружи от подслизистой основы располагается слой неисчерченных мышечных волокон (изнутри продольных, снаружи - циркулярных).
Артерии мочеиспускательного канала происходят из ветвей a. pudenda interna. Разные отделы канала питаются из различных источников: pars prostatica - из ветвей a. rectalis media и a. vesicalis inferior; pars membranacea - из a. rectalis inferior и a. perinealis; pars spongiosa - из a. pudenda interna. В васкуляризации стенок канала участвуют также a. dorsalis penis и a. profunda penis.
Венозная кровь оттекает к венам полового члена и мочевого пузыря. Лим-фоотток происходит из pars prostatica к лимфатическим сосудам предстательной железы, из pars membranacea и pars spongiosa - к паховым лимфатическим узлам. Иннервация осуществляется из nn. perinei и n. dorsalis penis (из n. pudendus), а также из вегетативного сплетения plexus prostaticus.
Бульбоуретральные железы (glandulae bulbourethral) представляют собой две железки, каждая диаметром 0,5-0,7 см, которые располагаются в толще diaphragma urogenitale над задним концом bulbus penis, кзади от pars membranacea urethrae. Выводной проток этих желез открывается в губчатую часть мочеиспускательного канала в области bulbus. Железы выделяют тягучую жидкость, которая защищает стенки мочеиспускательного канала от раздражения мочой.
Артерии к бульбоуретральным железам подходят из a. pudendae internae. Венозный отток происходит в вены bulbus и diaphragmae urogenitale. Лимфатические сосуды идут к Inn. lymphoidi iliaci interni. Иннервируются железы из n. pudendus, а также из вегетативного сплетения plexus prostaticus.
Предстательная железа (prostata) - непарный железисто-мышечный орган, напоминающий по форме усеченный конус. В ней выделяют верхушку (apex), основание (basis), переднюю и заднюю поверхности (facies anterior et posterior) (см. рис. 2.5). Масса ее около 25 г; вертикальный размер - приблизительно 3 см, горизонтальный - 4 см, сагиттальный - около 2,5 см. Она эксцентрично охватывает начальную часть мочеиспускательного канала и тесно прилежит своим основанием ко дну мочевого пузыря, а верхушкой - к мочеполовой диафрагме. Задняя поверхность предстательной железы граничит со стенкой прямой кишки и отделяется от
нее лишь тонкой пластинкой тазовой фасции (septum rectovesicale). Мочеиспускательный канал проходит через предстательную железу от ее основания к верхушке, располагаясь в срединной плоскости, ближе к ее передней поверхности.
Сзади и сверху к предстательной железе прилежат семенные пузырьки, а ме-диальнее их - семявыносящие протоки. Выводной проток семенного пузырька сливается под острым углом с расширенной частью семявыводящего протока. Образовавшиеся после этого семяизвергающие протоки (ductus ejaculatorius) проникают через заднюю поверхность предстательной железы, направляясь в толще ее вниз, медиально и кпереди, открываясь в pars prostatica urethrae двумя отверстиями на семенном бугорке. Щелевидное отверстие на верхушке семенного бугорка ведет в небольшой слепой кармашек, расположенный в толще предстательной железы, который носит название предстательная маточка (utriculus prostaticus). Название указывает на происхождение этого образования из слившихся нижних концов ductus paramesonephricus, из которых у женщины развиваются матка и влагалище.
Снаружи предстательная железа покрыта капсулой, богатой эластическими волокнами и содержащей мощные пучки гладких мышц, составляющих кольцевую предстательную мышцу. Вверху она сливается с круговым мышечным слоем мочевого пузыря; внизу - с мышцами, образующими произвольный сфинктер перепончатой части уретры.
Макроскопически в предстательной железе различают три доли: две боковые - правую и левую, разделенные между собой бороздкой (определяемой при пальпаторном исследовании), и среднюю долю (перешеек), которая располагается между задней поверхностью мочеиспускательного канала, дном мочевого пузыря и обоими семявыносящими протоками.
Микроскопическое (морфологическое) исследование предстательной железы не подтверждает деление ее на самостоятельные доли. Предстательная железа состоит из 30-50 трубчато-альвеолярных желез, между которыми находится соединительная ткань, богатая гладкомышечными волокнами. Железки предстательной железы открываются в предстательную часть мочеиспускательного канала вокруг семенного бугорка 20-30 выводными протоками. В подсли-зистом слое предстательной части мочеиспускательного канала располагаются также периуретральные железы, каждая из которых открывается в просвет мочеиспускательного канала.
Кровоснабжение предстательной железы осуществляется из aa. vesicalis inferiores и aa. rectalis mediae. Они проникают в нее многочисленными веточками вдоль семявыбрасывающих протоков, образуя богатую сеть капилляров. Большое количество вен предстательной железы, анастомозируя между собой, образуют вокруг нее сплетение, являющееся частью мочеполового венозного сплетения, имеющего связь с венозным сплетением прямой кишки.
Лимфатические сосуды начинаются в паренхиме предстательной железы и образуют вокруг нее, особенно на нижней поверхности, богатую лимфатическую сеть. Оттуда лимфоотток осуществляется в предпузырные лимфатические узлы, в лимфатические сосуды, проходящие вблизи мочеточников и семявы-носяших протоков по боковым стенкам таза к наружным и внутренним подвздошным лимфатическим узлам.
Иннервация предстательной железы осуществляется чувствительными и постганглионарными симпатическими и парасимпатическими нервными волокнами из нижнего подчревного сплетения (plexus hypogastricus inferior).
Семенной бугорок (colliculus serninalis), или семенной холмик, - возвышение продолговатой формы, расположенное на задней стенке предстательной части мочеиспускательного канала. Возвышение имеет длину около 2 см, ширину 3-4 мм и высоту 3-4 мм. Основу его составляет продольно-осевой тяж из эластических волокон, имеющих связь с продольными мышечными пучками пузырного треугольника и мембранозным отделом уретры.
Между эластическими волокнами заключено большое количество нервных волокон и окончаний. Поверхность семенного бугорка покрыта эпителием переходного типа.
Центральную часть тела семенного бугорка занимает предстательная, или мужская, маточка (utriculus prostaticus). Она является рудиментом сращенных концевых отделов мюллеровых ходов (длина ее 5-10 мм, глубина 3-5 мм, ширина 2-4 мм). В центре ее на семенном бугорке открывается отверстие диаметром 1-2 мм, ведущее в полость мужской маточки на глубину до 3-5 мм. По бокам от входа в полость мужской маточки, а иногда и на дне ее полости открываются отверстия семявыбрасывающих протоков (по одному справа и слева). С обеих сторон от семенного бугорка имеются углубления, куда открываются устья выводных протоков предстательной железы.
2.3. ФИЗИОЛОГИЯ ПОЧЕК И МОЧЕВЫВОДЯЩИХ ПУТЕЙ
Почка - паренхиматозный, наиболее сложно устроенный орган в мочевой системе. Структурно-функциональными единицами ее являются нефроны, которые, в процессе образования мочи, обеспечивают все основные функции органа. К ним относятся: регуляция водно-электролитного баланса организма; задержка жизненно важных веществ, таких как белок и глюкоза; поддержание кислотно-основного баланса; экскреция продуктов обмена, водорастворимых токсинов, лекарств; регуляция осмотического и кровяного давления, эритро-поэза; эндокринная функция.
Регуляция водно-электролитного баланса организма. Почки позволяют человеку есть и пить в соответствии с его привычками без изменения состава жидкостных и электролитных параметров организма.
Кровоснабжение почек в норме составляет 20 % от сердечного выброса. Примерно 99 % почечного кровотока приходится на корковый и 1 % - на мозговой слой почек. Большинство нефронов расположено в корковом, наружном слое органа. Мозговой, внутренний слой почки содержит специализированные нефроны в юкстамедуллярной области, лежащей на границе мозгового слоя. Эти нефроны обладают высокой концентрационной способностью, механизм которой будет рассмотрен ниже.
Почка - поистине уникальный орган, имеющий два капиллярных бассейна, состоящих из двух типов капилляров: клубочковых, находящихся под высоким
давлением и осуществляющих фильтрацию, и околоканальцевых (паратубуляр-ных), с низким давлением. Все это позволяет фильтровать и реабсорбировать большие объемы жидкости.
Нефрон - структурно-функциональная единица почки. Каждая почка содержит около миллиона нефронов. Нефрон состоит из клубочка и канальцев (рис. 2.6). Канальцы разделены на следующие отделы: проксимальный каналец, петля мозгового слоя (петля Генле) и дистальный каналец, впадающий в собирательную трубку. Моча образуется в результате трехфазного процесса: 1) простой фильтрации; 2) избирательной реабсорбции; 3) пассивной реабсорбции и экскреции.
Фильтрация происходит через полупроницаемую стенку капилляров клубочка, которая в основном непроницаема для белков и крупных молекул. Таким образом, фильтрат не содержит белка и клеточных элементов. Клубоч-ковый фильтрат формируется путем продавливания крови через капилляры клубочков. Движущей силой фильтрации является гидростатическое давление, которое регулируется приносящей и выносящей артериолами и обеспечивается артериальным давлением. Каждую минуту фильтруется около 20 % почечного
плазмотока (125 мл/мин), что равняется скорости клубочковой фильтрации.
Для сохранения относительно постоянных величин почечного кровотока и скорости клубочковой фильтрации в клубочках поддерживается достаточно постоянное гидростатическое давление. При изменении артериального давления происходит сокращение или расширение афферентной и эфферентной артериол - сосудов мышечного типа, входящих и выходящих из каждого клубочка. Этот процесс называется ауторе-гуляцией.
Ауторегуляция скорости клубочко-вой фильтрации достигается посредством саморегуляции почечного кровотока и механизма обратной связи, известного как клубочково-канальцевое равновесие.
Клубочково-канальцевое равновесие. При уменьшении скорости клубочко-вой фильтрации происходит снижение тока жидкости в канальцах и увеличивается время реабсорбции ионов натрия и хлора. Уменьшение количества ионов натрия и хлора, достигающих дисталь-ного канальца, ведет к снижению сопротивления афферентных артериол
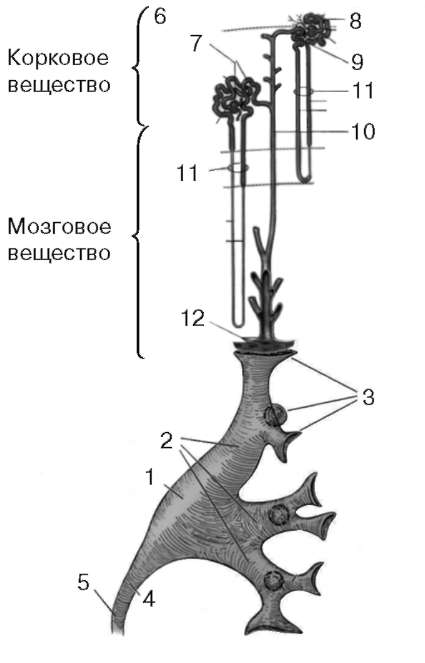
Рис. 2.6. Образование и выведение мочи:
1 - почечная лоханка; 2 - большие чашки; 3 - малые чашки; 4 - пиелоуретеральный сегмент; 5 - мочеточник; 6 - капсула; 7 - тельце почки; 8 - проксимальный извитой каналец; 9 - дистальный извитой каналец; 10 - собирательная трубочка;
11 - петля нефрона; 12 - решетчатое поле почечного сосочка
и сопровождается повышением почечного кровотока. При этом усиливается секреция ренина из юкстагломерулярного аппарата, что стимулирует выброс ангиотензина II, вызывающего сокращение эфферентных артериол. Повышение гидростатического давления в капиллярах клубочков возвращает скорость клубочковой фильтрации к ее нормальным значениям.
Юкстагломерулярный комплекс состоит из клеток плотного пятна (macula densa) - юкстагломерулярных клеток, являющихся специализированным эпителием дистального канальца, чувствительного к концентрации ионов натрия и способного влиять на клетки гладкой мускулатуры стенок афферентной и эфферентной артериол. Клетки macula densa также секретируют ренин - фермент, конвертирующий сывороточный белок ангиотензиноген в ангиотензин I. Впоследствии ангиотензин-превращающий фермент, который образуется в небольших количествах в легких, проксимальных канальцах и других тканях, превращает ангиотензин I в ангиотензин II, вызывающий вазоконстрикцию и повышающий артериальное давление. Ангиотензин II также стимулирует кору надпочечников, повышая секрецию альдостерона, который в свою очередь вызывает задержку воды и натрия, увеличивая объем циркулирующей крови.
Приведенная схема поддержания клубочково-канальцевого равновесия представляет собой систему отрицательной обратной связи. Другими словами, начальным стимулом системы является падение объема циркулирующей крови, приводящее к снижению перфузионного давления почек. Когда объем циркулирующей крови, перфузия почек и скорость клубочковой фильтрации восстановились, система отвечает снижением или выключением ответа на исходный стимул.
Селективная, или пассивная, реабсорбция. Функция почечных канальцев - селективная реабсорбция 99 % клубочкового фильтрата. Проксимальный каналец абсорбирует 60 % всех растворенных веществ, в том числе 100 % глюкозы и аминокислот, 90 % бикарбоната и 80-90 % неорганического фосфора и воды.
Реабсорбция происходит посредством активного и пассивного транспорта. Активный транспорт требует энергии для перемещения веществ против электрохимического или концентрационного градиента. Это основная детерминанта потребления кислорода почками. Посредством пассивного транспорта происходит реабсорбция веществ по электрохимическому и концентрационному градиентам или по градиенту давлений.
В основном реабсорбция осуществляется посредством активного транспорта веществ и свободного перемещения воды по принципу осмоса. При активной реабсорбции веществ происходит снижение их концентраций и, следовательно, падение осмотической активности в просвете канальца. Затем из-за присутствия осмотических сил вода перемещается из канальца в интерстиций, где концентрация осмотически активных веществ выше.
Петля Генле - это часть канальца, погружающаяся, или «изгибающаяся», из коркового слоя в мозговой (нисходящее колено) и затем возвращающаяся в кору почек (восходящая колено). Именно в этой части канальца моча при необходимости концентрируется. Это возможно благодаря высокой концентрации веществ в интерстиции мозгового слоя, которая поддерживается за счет наличия «противоточно-поворотной системы». Противоточно-поворотная система
поддерживает высокий осмотический градиент интерстиция мозгового слоя, что позволяет почкам концентрировать мочу. Петля Генле - это противоточ-но-поворотный множитель, а vasa recta (отдел перикапиллярной системы, входящий в мозговой слой в области высокой концентрации абсорбированных из первичной мочи веществ) - это противоточно-поворотный обменник, механизм которого описан ниже.
Функции различных частей петли Генле:
Нисходящее колено петли Генле относительно непроницаемо для растворенных веществ и хорошо проницаемо для воды, перемещаемой из канальца по осмотическому градиенту: жидкость в канальце становится гиперосмолярной.
Тонкий сегмент восходящего колена петли Генле практически непроницаем для воды, но в то же время проницаем для растворенных веществ, особенно ионов натрия и хлора, которые перемещаются по концентрационному градиенту из просвета канальца, жидкость в котором вначале становится изотонич-ной, а затем гипотоничной по мере выхода из нее ионов. Мочевина, абсорбировавшаяся в интерстиций мозгового слоя почки из собирательной трубки, диффундирует в восходящее колено. Это поддерживает концентрацию мочевины в интерстиции мозгового слоя, играя важную роль в процессе концентрации мочи.
Толстый сегмент восходящего колена петли Генле и начальный отдел дис-тального канальца непроницаемы для воды. Однако здесь происходит активный транспорт ионов натрия и хлора из просвета канальца, вследствие чего жидкость этого отдела канальца становится крайне гипотоничной.
Дистальный каналец и собирательная трубка: конечная концентрация мочи зависит от количества антидиуретического гормона, секретируемого задней долей гипофиза. В присутствии антидиуретического гормона дистальный каналец и собирательная трубка становятся проницаемыми для воды. При прохождении собирательной трубки через мозговой слой с высокой интерстициаль-ной концентрацией веществ вода выходит из просвета трубки и формируется концентрированная моча. При отсутствии антидиуретического гормона стенки дистального канальца становятся непроницаемыми для воды; таким образом формируется большое количество разведенной мочи.
Имеется тесная связь между гипоталамусом и задним гипофизом. В гипоталамусе присутствуют клетки-осморецепторы, чувствительные к изменению осмотического давления крови. При высоком потреблении воды наблюдается снижение осмотического давления крови, а при ее дефиците, соответственно, идет обратный процесс. При повышении осмотического давления крови нервные импульсы из гипоталамуса стимулируют задний гипофиз и усиливают секрецию антидиуретического гормона. В результате выработки антидиуретического гормона снижается потеря воды почками, так как она реабсорбируется в собирательных трубках.
Vasa Recta взаимодействует с петлей Генле посредством сложного механизма, направленного на концентрацию мочи путем противоточно-поворотного обмена. При отсутствии vasa recta высокая концентрация веществ в мозговом слое вымывалась бы током крови. Вещества диффундируют из сосудов, несущих кровь прямо в корковый слой и в сосуды, спускающиеся в мозговой слой,
в то время как вода делает противоположное: движется из нисходящих сосудов в восходящие. Эта система посредством подобного шунта позволяет веществам и воде рециркулировать внутри мозгового слоя.
Поддержание кислотно-основного состояния. Легкие и почки в совокупности поддерживают рН крови и внеклеточной жидкости в пределах 7,35-7,45 (34- 46 нмоль/л - концентрация Н+). Углекислый газ (СО2), растворенный в крови, является кислотой и элиминируется легкими. Почки же удаляют связанную кислоту посредством трех процессов: канальцевой секреции кислоты, клубоч-ковой фильтрации буферов, связанных с Н+, и образования аммиака.
1. Канальцевая секреция кислоты: бикарбонат натрия фильтруется в клубочке, реабсорбируясь затем в проксимальном канальце. Натрий абсорбируется посредством Na+/Н+-ионной помпы, обменивая Na+ на Н+-ионы на мембранах эпителия проксимального канальца.Na+/К+-помпа продвигает натрий через клетку из первичной мочи в обмен на калий.
2. Клубочковая фильтрация буферов, связанных с Н+:
А. Основная часть фильтруемого бикарбоната реабсорбируется (90 % в проксимальном канальце). Н+, высвобожденный при канальцевой секреции кислоты (см. выше), соединяясь с бикарбонатом (HCO3), формирует углекислоту:
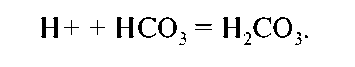
Карбоангидраза, присутствующая в клетках проксимального канальца, катализирует реакцию расщепления угольной кислоты на СО2 и Н2О. СО2 диффундирует в эпителиальную клетку и в присутствии карбоангидразы образует углекислоту. Последняя ионизируется до Н+ и НСО3. Н+ затем выкачивается из клетки в просвет канальца Na+/Н+-помпой, а натрий возвращается в плазму Na+/К+-помпой (см. выше); вода же абсорбируется пассивно.
B. Другие буферы, включая неорганический фосфат (НРО3), ураты и ионы креатинина, в дистальном отделе нефрона экскретируются в мочу как кислоты, если они связаны с Н+.
3. Аммиак (NH3) образуется ферментативно из глутамина и других аминокислот и секретируется в канальцы нефрона. Аммиак в сочетании с секретирован-ным в мочу Н+-ионом формирует недиффундирующий аммиак-ион (NH4-), выводимый с мочой.
Экскреция продуктов метаболизма. Фильтрация их происходит во время продвижения крови по клубочку. Некоторые ненужные организму вещества и чужеродные субстанции, например лекарственные препараты, не могут быть выведены из организма путем фильтрации. Такие вещества секретируются в канальцы нефрона и выделяются из организма с мочой.
Гормоны и почки. Ренин повышает продукцию ангиотензина II, высвобождаемого при снижении внутрисосудистого объема, например при кровопотере или дегидратации. Это ведет к:
■ констрикции эфферентных артериол для поддержания скорости клубоч-ковой фильтрации за счет повышения фильтрационного давления в клубочке;
■ выбросу альдостерона из коркового вещества надпочечников;
■ повышению секреции антидиуретического гормона задней долей гипофиза;
■ положительному инотропному действию на сердце и артериальной вазо-констрикции.
Альдостерон усиливает реабсорбцию ионов натрия и воды в дистальном канальце и собирательной трубочке, где Na+ обменивается на К+ и ионы водорода специфическими клеточными помпами. Секреция альдостерона усиливается при снижении концентрации Na+ в сыворотке крови. Это может произойти, например, при потере большого объема желудочного сока, так как желудочный сок содержит значительное число ионов натрия, хлора, водорода и калия. Следовательно, невозможно корригировать возникающий алкалоз и гипокалиемию без предварительного возмещения ионов натрия физиологическим раствором.
Предсердный натрийуретический пептид секретируется при повышении давления в предсердиях, например при сердечной недостаточности или жидкостной перегрузке. Предсердный натрийуретический пептид приводит к повышению потерь натрия, хлоридов и воды преимущественно за счет повышения скорости клубочковой фильтрации.
Антидиуретический гормон повышает проницаемость стенок дистального канальца и собирательной трубки для воды и таким образом концентрирует мочу. С другой стороны, при сниженной секреции антидиуретического гормона формируется значительное количество «разведенной» мочи. Подобная ситуация возникает преимущественно при падении концентрации натрия в плазме крови после потребления больших объемов воды. Снижение уровня натрия контролируется осморецепторами. При кровопотере или дегидратации гормоны взаимодействуют между собой, что играет роль в поддержании нормального внутрисосудистого объема.
К другим веществам, синтезируемым почками, относятся 1,25-дигидрок-си-витамин D (наиболее активная форма витамина D), обеспечивающий абсорбцию кальция из кишечника, и эритропоэтин, стимулирующий продукцию эритроцитов. Продукция этих веществ снижается при почечной недостаточности.
Физиология мочевыделения. Образующаяся в почечных канальцах моча выделяется в почечную чашечку, а затем в фазе ее систолы попадает в почечную лоханку. Последняя постепенно заполняется мочой, и по достижении порога раздражения возникают импульсы от барорецепторов, сокращается мускулатура почечной лоханки, раскрывается просвет мочеточника и моча благодаря сокращениям его стенки продвигается в мочевой пузырь.
Функция нижних мочевыводящих путей состоит из двух фаз - накопления мочи (резервуарная функция) и ее эвакуации. При этом отмечается определенный, но равномерный режим работы мочевого пузыря и запирательного аппарата, то есть чередование между его заполнением и опорожнением. При физиологических условиях заполнение мочевого пузыря происходит подсознательно и очень медленно (у взрослого человека в течение 2-4 часов), а эвакуация осуществляется по желанию и заканчивается в течение 20-30 секунд. У ребенка до 1,5-2 лет мочеиспускание рефлекторное. Во время роста, с развитием проводящих путей между центрами спинного мозга и головным мозгом, мочеиспускание становится контролируемым.
Процессы накопления и эвакуации мочи обеспечиваются определенными анатомическими образованиями и составляют единую функциональную систему. Удержание мочи обеспечивают следующие образования: шейка мочевого пузыря и проксимальная уретра, часто рассматриваемая как внутренний сфинктер (или гладкомышечный сфинктер мочевого пузыря), наружный сфинктер (или поперечно-полосатый сфинктер уретры), мышцы тазового дна. Накопление мочи обеспечивается мышцами мочевого пузыря. Функционально процесс удержания мочи связан со сложным комплексом физиологических механизмов как запирательного аппарата, так и мочевого пузыря, что обеспечивается рефлекторными актами и чисто механическими компонентами.
Для лучшего понимания нормальной функции и дисфункции нижних мочевых путей необходимо изложить особенности их нервной регуляции. Афферентная иннервация мочевого пузыря и уретры осуществляется за счет рецепторов, чувствительных к болевому, температурному и прессорному воздействию. Чувствительные рецепторы имеются во всех слоях мочевого пузыря, но наибольшее их количество расположено в области треугольника Льето. Выделяются резко специализированные рецепторы мочевого пузыря, реагирующие на быстрые изменения его объема, и окончания, воспринимающие медленные изменения внутреннего давления. По степени адаптации к наполнению различают фазные и тонические рецепторы мочевого пузыря. Важную роль в акте мочеиспускания играют рецепторы, заложенные в стенке уретры, особенно в ее проксимальном отделе, а также чувствительные рецепторы поперечно-полосатых мышц уретры и промежности.
Все нервные импульсы, генерируемые в нижних мочевыводящих путях, поступают в центральные отделы нервной системы, за счет чего осуществляется координированный акт мочеиспускания. К нервным центрам относятся:
■ интрамедиолатеральные клетки стволов и клетки вентральных рогов серого вещества сакрального отдела спинного мозга;
■ ретикулярная формация ствола мозга;
■ мозжечок, который получает нервные импульсы от детрузора и мышц тазового дна через спиноцеребеллярный тракт;
■ передняя группа ядер гипоталамуса;
■ базальные ганглии, образования стволовой клетки мозга: хвостатое ядро, чечевицеобразное ядро, черная субстанция, красное ядро;
■ зрительные бугры (неспецифические таламические ядра, которые расположены во внутреннем слое);
■ кора головного мозга - является центром координации акта мочеиспускания. Эфферентная система регуляции акта мочеиспускания начинается от коры
головного мозга, причем корковые центры регуляции акта мочеиспускания - понятие функционально-динамическое, имеющее, помимо постоянного анатомического представления, многочисленные условно-рефлекторные связи. Из клеток 5-го слоя коры больших полушарий возникают нисходящие волокна, идущие к субкортикальным образованиям и дальше к спинному мозгу. Однако считается, что нет убедительных доказательств наличия прямого кортикоспи-нального пути, а существуют многочисленные короткие пути и промежуточные пункты.
В медиальной ретикулярной формации начинаются аксоны, проходящие через вентральный ретикулоспинальный путь к спинным центрам мочеиспускания. Спинномозговые центры мочеиспускания располагаются в тора-колюмбальном и сакральном отделах спинного мозга, что соответствует парасимпатическим и симпатическим участкам вегетативной нервной системы. Моторные преганглионарные симпатические нейроны начинаются в клетках, которые локализуются в интермедиолатеральных ядрах сегментов от Th 12 до L 2, и участвуют в формировании чревных нервов и ганглиев чревного сплетения. Преганглионарные волокна проходят через ганглии паравертебрального ствола и в составе чревного нерва заканчиваются в узлах пузырных сплетений. Постганглионарные волокна направляются к детрузору, шейке мочевого пузыря и треугольнику Льето. Адренергические рецепторы размещаются в нижних мочевыводящих путях неравномерно: α-адренорецепторы преобладают в области шейки мочевого пузыря, проксимальной уретры, предстательной железе; Р-адренергические рецепторы располагаются в области тела мочевого пузыря. Рецепторы высвобождают адренергические нейромедиаторы (норад-реналин и адреналин). В мочевой системе преобладаютα1А- и α1D -адрено-рецепторы. На рис. 2.7 представлена локализация адренорецепторов в мочевом пузыре, простате и уретре.
Парасимпатические эфферентные группы, идущие к нижним мочевым путям, начинаются клеточными телами в сакральных парасимпатических ядрах (интермедиалатеральном сером веществе) от S2-S4 сегментов. Моторные волокна через вентральный корешок и далее в составе тазовых и подчревных нервов направляются к пузырному сплетению. Холинергиче-ские рецепторы преобладают в области тела мочевого пузыря и почти не встречаются в проксимальной уретре.
Возбуждение парасимпатической нервной системы ведет к сокращению детрузора. Возбуждение симпатической нервной системы вызывает неоднозначные функциональные изменения. Так, моторный β-адренер-гический эффект вызывает расслабление детрузора, раскрытие устьев мочеточников; α -адренергическое влияние вызывает сокращение три-гональной мышцы и интрамуральной мускулатуры мочеточников, повышает тонус области внутреннего сфинктера и проксимальной уретры.
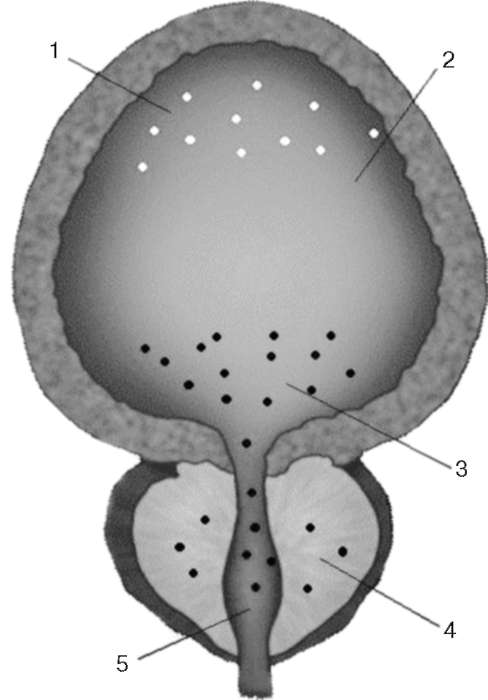
Рис. 2.7. Локализация адренорецепторов в мочевом пузыре, простате и уретре: 1 - β-адренорецепторы; 2 - мочевой пузырь; 3 -α -адренорецепторы; 4 - предстательная железа; 5 - проксимальная уретра
Таким образом, суммарное действие симпатической иннервации на нижние мочевыводящие пути заключается в постоянном поддержании тонуса внутреннего сфинктера и проксимальной уретры, раскрытии устьев мочеточников, расслаблении детрузора по мере наполнения мочевого пузыря. Прекращение симпатического влияния совпадает с активацией рефлекса мочеиспускания.
Помимо симпатической и парасимпатической иннервации акт мочеиспускания регулирует и соматическая нервная система. Эфферентные нейроны, идущие от серого вещества переднего рога S2-S4 сегментов спинного мозга через срамное сплетение и срамной нерв, заканчиваются в области поперечнополосатого наружного сфинктера и в мускулатуре тазового дна.
Акт мочеиспускания обеспечивается за счет сложной рефлекторной регуляции центральной и периферической нервной системы. Рефлекторные механизмы скоординированы во времени. Активность рефлекса отражает сумму всех возбуждающих и тормозящих нервных импульсов, воздействующих на аппарат нижних мочевыводящих путей.
Акт мочеиспускания осуществляется следующим образом. Сокращающийся m. detrusor urinae выжимает мочу из мочевого пузыря в мочеиспускательный канал, открывающийся благодаря расслаблению своих сфинктеров: непроизвольного (m. sphincter vesicae) и произвольного (m. sphincter urethrae). При этом у мужчин происходит также расслабление мышечной части предстательной железы, выполняющей функции третьего (непроизвольного) сфинктера. Закрытие мочевого пузыря происходит при расслаблении m. detrusor и сокращении названных сфинктеров.
Возбуждение центра мочеиспускания вызывает импульсацию в парасимпатических волокнах тазовых внутренностных нервов (nn. splanchnici pelvici), а мышца наружного сфинктера иннервируется соматическим нервом - ветвью полового нерва (n. pudendus).
Движение мочи по мочеиспускательному каналу играет важную роль в акте мочеиспускания: оно рефлекторно по афферентным волокнам полового нерва стимулирует сокращение мочевого пузыря. Поступление мочи в задние отделы мочеиспускательного канала и его растяжение способствуют сокращению мышцы мочевого пузыря. Передача афферентных и эфферентных импульсов этого рефлекса осуществляется по подчревному нерву (n. hypogastricus).
2.4. ФИЗИОЛОГИЯ МУЖСКИХ ПОЛОВЫХ ОРГАНОВ
Физиология полового акта
Половой акт (синоним: коитус, совокупление, копуляция) является фрагментом сложной картины полового поведения человека. Несмотря на то что половой акт - парный физиологический процесс, изменения в организме мужчины и женщины существенно различаются. Поскольку, как правило, половой акт протекает в интимной обстановке, физиологические изменения организма до, во время и после полового акта описывались весьма умозрительно. В настоящее время во многом благодаря исследованиям, проведенным на добровольцах
с помощью специальной техники, фиксирующей изменения в организме мужчин и женщин во время полового акта, стала понятна его физиология.
Различают несколько стадий полового акта, переходящих друг в друга и объединенных общим понятием «половой цикл»:
■ возбуждение;
■ «плато»;
■ оргазм;
■ обратное развитие (детумесценция) .
Половому акту обычно предшествует период взаимных ласк. Для нормального осуществления полового акта у мужчин необходимо участие следующих друг за другом структурно-функциональных составляющих:
1) нейрогуморальной, обусловленной работой центральной нервной и эндокринной систем, которые обеспечивают силу полового влечения и возбудимость соответствующих отделов центральной нервной системы, регулирующих половое поведение;
2) психической, обусловленной работой коры больших полушарий, обеспечивающей направленность полового влечения и эрекцию до начала полового акта;
3) эрекционной, обусловленной преимущественно работой спинальных центров, во время которой происходят введение полового члена во влагалище и фрикции (движения полового члена во влагалище);
4) эякуляционно-оргастической, обусловленной также преимущественно работой спинальных центров, во время которой происходит эякуляция и возникает оргазм.
В стадии возбуждения у мужчины при половой стимуляции происходит усиление притока крови к половым органам при одновременном некотором затруднении оттока крови по венам. Это приводит к переполнению кровью пещеристых тел полового члена и увеличению его размеров. Полагают, что парасимпатический контроль просвета сосудов является ведущим в возникновении эрекции.
Введение полового члена, фрикции приводят у мужчин к увеличению полового возбуждения, учащению сердцебиений и дыхания, повышению артериального давления, гиперемии лица. Максимальных значений повышение артериального давления и частота сердцебиений у мужчины достигает в период оргазма, который переживается как сладострастное ощущение. Оргазм у мужчин начинается с ритмичных сокращений семявыносящих, семявыбрасывающих протоков и семенных пузырьков. При этом происходит выделение наружу под большим давлением эякулята. Оргазм у мужчин длится несколько секунд, после чего нормальная эрекция быстро ослабляется и возникает детумесценция - уменьшение кровенаполнения половых органов. За ним следует период половой рефрактерности. Повторная эрекция возможна спустя некоторое время.
Четкое определение понятий «норма», «нормальный» в физиологии полового акта весьма затруднено вследствие чрезвычайного переплетения биологических, социальных, индивидуальных особенностей личности. Полагают, что если половая жизнь не вызывает чувства усталости, недовольства, если в течение дня партнеры остаются веселыми и бодрыми, то очевидно, что их половая жизнь является оптимальной.
Гормональная регуляция физиологических функций
Мужские половые железы (яички). В них происходят процессы сперматогенеза и образование мужских половых гормонов - андрогенов.
Сперматогенез (от греч. sperma, родительный падеж spermatos - семя и genesis - образование) - процесс превращения диплоидных мужских половых клеток в гаплоидные, свободные и дифференцированные клетки - сперматозоиды.
Различают четыре периода сперматогенеза: 1) размножение; 2) рост; 3) деление и созревание; 4) формирование, или спермиогенез (спермиотелиозис). В первом периоде диплоидные исходные мужские половые клетки (сперматогонии) несколько раз делятся путем митоза (число делений у каждого вида постоянно). Во втором периоде половые клетки (сперматоциты 1-го порядка) увеличиваются в размерах, а их ядро проходит длительную профазу, во время которой совершается конъюгация гомологичных хромосом и кроссинговер, сопровождающийся обменом участками между гомологичными хромосомами, и образуются тетрады. В третьем периоде происходят два деления созревания (мейоз), осуществляется редукция, или уменьшение, числа хромосом вдвое (при этом в одних тетрадах при первом делении к полюсам веретена расходятся гомологичные хромосомы, при втором - хроматиды, а в других, наоборот, - сначала хроматиды, затем гомологичные хромосомы).
Таким образом, каждый сперматоцит 1-го порядка дает 2 сперматоцита 2-го порядка, которые после второго деления образуют четыре одинаковые по размерам гаплоидные клетки - сперматиды. Последние не делятся, вступают в четвертый период сперматогенеза, или спермиогенез, и превращаются в сперматозоиды: сперматида из округлой становится вытянутой, происходит новообразование одних структур (акросома, побочное ядро, жгутик и т. д.), исчезновение других (рибосомы, эндоплазматический ретикулум и т. д.). Большая часть цитоплазмы исчезает из клетки. Вытянутое ядро с конденсированным хроматином и акросомой (производное аппарата Гольджи) размещаются на апикальном полюсе клетки и образуют головку сперматозоида; центриоль ложится обычно у базального полюса ядра, от нее берет начало жгутик; митохондрии окружают центриоль или формируют так называемое побочное ядро, расположенное в промежуточном отделе сперматозоида. Зрелые сперматозоиды накапливаются в придатке семенника. Сперматогенез продолжается у мужчин до преклонного возраста.
Продолжительность полного сперматогенеза, состоящего из четырех циклов, составляет от 64 до 75 дней. Но созревают все сперматозоиды не одновременно: в любой момент в стенке канальца можно обнаружить сотни и сотни клеток на разных стадиях сперматогенеза - начальных, промежуточных и завершающих. Один цикл зародышевого эпителия составляет приблизительно 16 дней.
Образование андрогенов происходит в интерстициальных клетках - гланду-лоцитах (клетки Лейдига), локализующихся в интерстиции между семенными канальцами и составляющих примерно 20 % от общей массы яичек. Небольшое количество мужских половых гормонов вырабатывается также в сетчатой зоне коркового вещества надпочечников.
К андрогенам относится несколько стероидных гормонов, наиболее важным из которых является тестостерон. Продукция этого гормона определяет адекватное развитие мужских первичных и вторичных половых признаков (маскулинизирующий эффект). Под влиянием тестостерона в период полового созревания увеличиваются размеры полового члена и яичек, появляется мужской тип оволосения, меняется тональность голоса. Кроме того, тестостерон усиливает синтез белка (анаболический эффект), что приводит к ускорению процессов роста, физического развития, увеличению мышечной массы. Тестостерон влияет на процессы формирования костного скелета - он ускоряет образование белковой матрицы кости, усиливает отложение в ней солей кальция. В результате увеличиваются рост, толщина и прочность кости. При гиперпродукции тестостерона ускоряется обмен веществ, в крови возрастает количество эритроцитов.
Механизм действия тестостерона обусловлен его проникновением внутрь клетки, превращением в более активную форму (дигидротестостерон) и дальнейшим связыванием с рецепторами ядра и органелл, что приводит к изменению процессов синтеза белка и нуклеиновых кислот. Секреция тестостерона регулируется лютеинизирующим гормоном аденогипофиза, продукция которого возрастает в период полового созревания. При увеличении содержания в крови тестостерона по механизму отрицательной обратной связи тормозится выработка лютеинизирующего гормона. Уменьшение продукции обоих гона-дотропных гормонов - фолликулостимулирующего и лютеинизирующего - происходит также при ускорении процессов сперматогенеза.
У мальчиков в возрасте до 10-11 лет в яичках обычно отсутствуют активные гландулоциты (клетки Лейдига), в которых вырабатываются андрогены. Однако секреция тестостерона в этих клетках происходит во время внутриутробного развития и сохраняется у ребенка в течение первых недель жизни. Это связано со стимулирующим действием хорионического гонадотропина, который продуцируется плацентой.
Недостаточная секреция мужских половых гормонов приводит к развитию евнухоидизма, основными проявлениями которого являются задержка развития первичных и вторичных половых признаков, диспропорциональность костного скелета (несоразмерно длинные конечности при относительно небольших размерах туловища), увеличение отложения жира на груди, в нижней части живота и на бедрах. Нередко отмечается увеличение молочных желез (гинекомастия). Недостаток мужских половых гормонов приводит также к определенным нервно-психическим изменениям, в частности к отсутствию влечения к противоположному полу и утрате других типичных психофизиологических черт мужчины.
Добавочные половые железы постоянно испытывают на себе влияние андро-генов, которые способствуют их правильному формированию и нормальному функционированию. Тестостерон стимулирует образование фруктозы в семенных пузырьках, лимонной кислоты и фосфатазы в предстательной железе, кор-нитина в придатках яичка и пр.
Уменьшение содержания в семенной жидкости фруктозы, лимонной кислоты, кислой фосфатазы, корнитина может указывать на снижение внутрисекреторной
функции яичек. Обнаружено, что приблизительно через 7-10 дней после двусторонней орхэктомии мужские добавочные половые железы у грызунов атрофируются до минимума.
Нормальный уровень тестостерона в плазме взрослого мужчины составляет 12-35 нмоль/л или 345-1010 нг/дл.
Контрольные вопросы
1. Каково строение почки?
2. Что является структурно-функциональной единицей почки?
3. Расскажите о механизме образования мочи.
4. В чем заключается эндокринная функция почки?
5. Какие имеются физиологические сужения мочеточника?
6. Каково строение стенки мочевого пузыря? Что такое треугольник Льето?
7. Перечислите отделы мужского мочеиспускательного канала.
8. Каково количество и строение оболочек яичка?
9. Расскажите о строении и функции предстательной железы.
10. Опишите физиологию полового акта.
11. Какие существуют этапы сперматогенеза?