Биология: учебник / Пехов А.П., -, 2010. - 664 с.
|
|
|
|
ГЛАВА V НАСЛЕДСТВЕННОСТЬ И ИЗМЕНЧИВОСТЬ.
Генетический материал
Наследственность и изменчивость - это важнейшие свойства живого, которые не только отличают живое от неживого, но и опре- деляют совместно с размножением бесконечное продолжение (непрерывность) жизни.
НАСЛЕДСТВЕННОСТЬ И НЕПРЕРЫВНОСТЬ
ЖИЗНИ
Из всех органических молекул способностью к саморепродукции обладают только нуклеиновые кислоты. Между тем, находясь в клетках, они контролируют их структуру и свойства (активность). Поэтому уникальность жизни в генетическом смысле заключается в том, что нуклеиновые кислоты через половые клетки обеспечивают химическую связь между поколениями. Благодаря размножению, наследственности и изменчивости жизнь видов продолжается бесконечно долго как непрерывное чередование поколений с сохранением между ними химических связей.
Уникальность жизни определяется также постоянством видов. В процессе размножения исходные организмы всегда продуциру- ют самих себя, т. е. «подобное рождает подобное». Потомство пары мышей всегда является мышами, точно так же как две бактериальные клетки, появившиеся в результате деления одной бактериальной клетки, являются бактериями того же вида. Следовательно, постоянство видов определяется передачей сходства от родителей к потомству, т. е. наследованием свойств своих родителей, вследствие чего организмы всех поколений (генераций) в пределах вида характеризуются общим наследственным (генетическим) поведением.
Наследственность - это передача сходства от родителей к потомству или склонность организмов походить на своих родителей. Наследственность означает передачу анатомических, физиологических и других свойств и особенностей от организмов одних поколений (генераций) к организмам других. Поскольку связь между поко-
лениями обеспечивается половыми клетками, а оплодотворение представляет собой слияние ядер этих клеток и образование зиго- ты, то ядра половых клеток составляют физическую основу такой связи. Когда речь идет о наследственности организмов, то следует понимать, что единственным материалом, который наследуется потомством от своих родителей, является генетический материал, сосредоточенный в ядерных структурах (хромосомах) и представляющий собой гены (единицы наследственности). Следовательно, потомство наследует от своих родителей не признаки (свойства), а гены, которые контролируют эти признаки (свойства), причем показателем генетической детерминируемости признаков является наследуемость последних.
Различают наследование, которое не связано с полом, и наследование, контролируемое, ограничиваемое и сцепленное с полом. Под наследованием, не связанным с полом, понимают то наследование, которое не зависит от пола организмов-родителей или потомства. При наследовании, контролируемом полом, проявление генов отмечают у обоих полов, но по-разному. Наследование, ограничиваемое полом, заключается в том, что проявление генов происходит лишь у одного пола. Наконец, наследование, сцепленное с полом, обусловлено локализацией соответствующих генов в половых хромосомах. Кроме этих типов наследования, различают также полигенное наследование, когда наследуемость признака подвержена контролю со стороны нескольких генов.
Однако организмы, которые произошли от какой-либо пары родителей, не все являются совершенно одинаковыми. В одном и том же помете мышей или в одной и той же культуре бактерий (происходящей от одной бактериальной клетки) можно встретить организмы, которые по каким-то признакам будут отличаться от родителей. Иногда у потомства обнаруживаются признаки, которые были присущи лишь далеким предкам, или признаки, которые являются совершенно новыми не только для их родителей, но и для дальних предков. Следовательно, для индивидуальных организмов характерны различия, изменчивость признаков.
Изменчивость - это свойство, противоположное наследственности. Она заключается в изменениях генетического материала, сопровождаемых изменениями признаков организмов. Результатом изменчивости является образование новых вариантов организмов, разнообразие жизни.
НАСЛЕДСТВЕННОСТЬ И СРЕДА
Непрерывность жизни является генетической, ибо наследственность и изменчивость поддерживают стабильность свойств видов и способность организмов к изменчивости. Однако генетическая непрерывность жизни связана не только с наследственностью и ее изменчивостью, но и со средой, в которой живут и развиваются организмы.
Все организмы живут и размножаются в среде, условия которой не безразличны для них. До тех пор, пока организм живет, его наследственность взаимодействует со средой. Внешняя среда влияет на выражение наследственных признаков и определяет степень их проявления. Взаимодействие наследственности и среды определяет, каким организм является в данный момент и как он должен развиваться в будущем. Можно сказать, что наследственность предполагает, каким организм должен стать, но не каким он будет. То, каким организм станет в действительности, решается взаимодействием наследственности и среды.
Геном - совокупность генов или полное количество ДНК, харак- терное для клеток организмов.
Генотип - совокупность всех генов данного организма, его инди- видуальная генетическая конструкция, которую он получает от своих родителей. Генотип относительно стоек на протяжении всей жизни индивидуума. Для взрослого человека характерен тот же генотип, который был присущ ему в период внутриутробного роста и развития, в детстве, отрочестве, юности, старости.
Фенотип - совокупность всех признаков (свойств) данного орга- низма (размеры, форма, строение, масса тела, цвет, химический состав, поведение и т. д.). В противоположность генотипу фенотип изменяется в процессе роста и развития организма на протяжении всей его жизни, но является различным в эмбриональном периоде, после рождения, во время полового созревания и т. д.
Генотип организма определяют путем наблюдения его действия (реализации генетической информации) и влияния на фенотип в условиях определенной среды. Когда два или несколько организмов растут и развиваются в одинаковых условиях, но фенотипы их различны, то это означает, что такие организмы имеют разные генотипы. Строго говоря, фенотипы являются результатом взаимодействия различных генов (компонентов генотипа) между собой и генотипа со средой. Поэтому нельзя думать, что организм
или какие-либо признаки организма зависят исключительно только от генотипа или только от среды. Два одинаковых генотипа могут развиваться в разных условиях и дать разные фенотипы. Точно так же два разных генотипа могут развиваться в условиях одинаковой среды и дать разные фенотипы. Кроме того, если организмы, развивающиеся в условиях сходной или неодинаковой среды, имеют сходные фенотипы, это еще не означает, что их генотипы одинаковы. В таких случаях часто имеет значение доминантность признаков, которая может осложнить установление генотипа.
Фенотип организма с определенным генотипом формируется не только под влиянием факторов среды, действующих в данное время, но и тех факторов, которые действовали ранее на протяжении всей жизни организма. В случае человека любой индивидуум с его физическими, анатомическими, физиологическими и психическими свойствами является продуктом роста и развития, детерминируемого определенным генотипом и осуществляющегося в условиях среды с определенной последовательностью различных факторов этой среды, включая социальные. Другими словами, каждый индивидуум представляет собой продукт его генотипа и жизненного опыта.
Наиболее демонстративно взаимодействие наследственности и внешней среды проявляется у однояйцовых (идентичных) близнецов, которые действительно имеют одинаковые генотипы, вследствие чего одинаковы и их фенотипы. Многие наблюдения свидетельствуют о том, что жизнь и воспитание идентичных близнецов в различных семьях и в условиях разной среды всегда приводили к тому, что близнецы сохраняли физическое сходство, но различались между собой как личности. Примеры, касающиеся однояйцовых близнецов, свидетельствуют о том, что наследственные задатки проявляются лишь в условиях определенной среды. В частности, они служат одним из решающих доказательств того, что на развитие умственных способностей человека, которые сами по себе детерминированы генетически, может оказывать влияние и среда.
Итак, хотя фенотип нельзя свести только к генотипу или среде, различия в фенотипе могут определяться раздельными или совместными различиями в генотипе или среде. Наследственность и среда постоянно взаимодействуют, определяя свойства организмов. Это, однако, не означает абсолютного влияния среды на проявление всех признаков. Известны отдельные признаки, развитие которых
настолько сильно ограничено генотипом, что они не подвержены модификации ни одним из известных факторов среды.
Можно сказать, что эти признаки генетически очень узко детерминированы к существующему разнообразию среды. Примером таких признаков являются группа крови и цвет глаз у человека. Одновременно есть признаки, которые зависят только от внешней среды, например уродства новорожденных в результате приема ядов или алкоголя беременными женщинами, но такие признаки не имеют прямого отношения к наследственности.
В биологии вообще и генетике в частности большое значение имеют вопросы, связанные с природой изменчивости организмов и с отношением изменчивости к наследственности, ибо причины различий между индивидуальными организмами не всегда одинаковы и могут быть обусловлены как факторами среды, так и факторами наследственности (генами).
Никогда нельзя найти пару организмов, которые были бы совершенно одинаковы фенотипически. В лесу, степи или на возделываемой делянке даже рядом растущие растения различаются между собой, ибо они получают разное количество света, воды, минеральных веществ. Животные также различны между собой в пределах одного вида, так как никогда не получают точно одинакового количества корма в разных местах и в разное время. Следовательно, находясь в разных условиях по отношению к питательным веществам, свету, температуре и другим внешним факторам, организмы даже с одинаковым генотипом всегда различаются между собой фенотипически. Такие различия между сходными по генотипу организмами получили название фенотипической изменчивости, модификаций или ненаследственной изменчивости.
Однако различия между организмами могут вызываться и другими причинами. При одних и тех же условиях кормления, климата и т. д. щенок всегда вырастает в собаку, а котенок - в кошку, ибо организмы этих видов имеют принципиально различные генетические основы. Но в случае разных генотипов индивидуальные организмы одного и того же вида также могут различаться между собой по отдельным признакам. Поэтому изменчивость, детерминируемую наследственными факторами, называют генотипической или наслед- ственной изменчивостью. Ее возникновение связано с изменениями (мутациями) генов. Сочетание мутантных генов с немутантными или другими мутантными генами создает генотипическое разнообразие организмов.
Принципиальное значение имеет определение степени раздельного влияния наследственности и среды на фенотипические различия индивидуальных организмов в пределах видов. Оценка этих влияний сопряжена со многими трудностями и в каждом отдельном случае нуждается в специальном анализе, тем не менее практика сельского хозяйства и экспериментальные исследования свидетельствуют, что такое определение вполне возможно.
Известно, что улучшение агротехники при культивировании растений или условий содержания и кормления при разведении домашних животных, генотип которых характеризуется не очень благоприятными возможностями, приводит лишь к некоторому повышению урожая или продуктивности, причем не воспроизводимому в потомстве этих организмов. В то же время среди культивируемых растений можно найти генотипические варианты, которые дают больший урожай, а среди животных - генотипические варианты, характеризующиеся большей продуктивностью по мясу, молоку, шерсти и т. д. Давно замечено, что даже незначительное генотипическое улучшение дает эффект, поскольку контролирующие его гены передаются по наследству, и это улучшение воспроизводится в потомстве. Более того, генетическое совершенствование продуктивности культурных растений и домашних животных привело к созданию огромного сортового и породного разнообразия этих организмов. Разумеется, сортовые достоинства растений зависят от качества почвы, климатических условий, удобрений и т. д., а породные достоинства животных - от условий их содержания и кормления.
Таким образом, практический опыт свидетельствует, что фенотипические различия между организмами определяются в основном генотипом.
Более точные данные о степени влияния генотипа и среды на фенотипические различия дали экспериментальные исследова- ния, выполненные в условиях контролируемой среды и использовании организмов со сходными генотипами или генотипами, различающимися между собой по определенным генам - клонов чистых и инбредных организмов.
Клоном называется потомство вегетативно размножающегося индивидуального организма, например культура бактерий, полученных в результате размножения одиночной бактериальной клетки; культур соматических клеток, полученных из одиночных соматических клеток; групп деревьев, развивавшихся из черенков, взятых
от одного растения. Чистая линия - это потомство, полученное от индивидуального организма в результате самоопыления или самооплодотворения. Размножение многих культивируемых растений (пшеница, горох и т.д.) происходит главным образом путем самоопыления. У некоторых животных (пресноводные улитки) имеет место самооплодотворение. Как и в случае клонов, все члены линии являются генетически однородными организмами, так как обладают одинаковыми наледственно-константными признаками. Их генетическое однообразие более совершенно, чем у потомства, получаемого после перекрестного опыления разных организмов.
Инбредные линии - это организмы, разводимые в мире раздельнополых животных путем неоднократных скрещиваний между собой близких родственников (братьев и сестер). Генотипическое однообразие инбредных животных становится более выраженным с каждым новым скрещиванием. Например, известен ряд инбредных линий белых мышей, крыс и морских свинок. Исследования клонов, чистых и инбредных линий организмов позволили не только измерить действие факторов внешней среды, но и более точно определить влияние генотипа на фенотипические различия. Результаты, полученные в этой области, совпадают с данными сельскохозяйственной практики.
Исследования взаимодействия генотипа и среды на примере организмов многих видов показали, что для реакции определенного генотипа в ответ на фактор внешней среды всегда характерен диапазон, измеряемый количеством фенотипов, продуцируемых этим генотипом. Разнообразие фенотипов, возникающих в результате взаимодействия определенного генотипа с разными факторами среды (разными средовыми условиями), генетики называют нормой реакции этого генотипа.
Многие генетические, детерминированные реакции организмов на внешние факторы имеют адаптивный характер, что обеспечивает жизнь и размножение организмов в колеблющихся условиях среды. Среди адаптивных реакций различают физиологический гомеостаз и гомеостаз развития. Физиологический гомеостаз - это генетически детерминированная способность организмов противостоять колеблющимся условиям внешней среды. У млекопитающих, в том числе у человека, типичным примером физиологического гомеостаза является константность осмотического давления в клетках и концентрации водородных ионов в крови вследствие функциони-
рования почек и наличия в крови буферных субстанций. Гомеостаз развития - это генетически детерминированная способность организмов так изменять отдельные реакции, что функции организмов при этом в целом сохраняются. Например, выход из строя одной почки сопровождается тем, что остающаяся почка выполняет двойную нагрузку. Примером гомеостаза развития может быть также приобретение переболевшим организмом иммунитета против соответствующей инфекции.
Часто между физиологическим гомеостазом и гомеостазом развития очень трудно выявить различия, поэтому многие адаптивные реакции носят промежуточный характер. Примером такой адаптивной реакции является изменение количества эритроцитов в крови у людей в зависимости от пребывания их на той или иной высоте над уровнем моря. Количество эритроцитов у людей, живущих в разных высотных условиях, повышается по мере удаления от уровня моря. Это связано с тем, что уменьшение содержания кислорода в атмосфере вызывает интенсификацию его транспорта эритроцитами в результате увеличения количества последних. Возвращение человека из высокогорного района в район, лежащий на уровне моря, сопровождается снижением количества эритроцитов.
Норма реакции у всех организмов имеет пределы, определяя их фенотипическое разнообразие лишь в условиях среды, кото- рая для организмов любого вида не имеет резких и необычных отклонений. Например, многие тропические растения выживают в условиях повышенных или пониженных температур, характерных для стран с жарким климатом. Однако они погибают от мороза, к которому устойчивы растения, являющиеся обычными обитателями северных широт. В случае человека потеря генотипом в результате мутации способности детерминировать адаптивные реакции на факторы обычной для него среды сопровождается наследственным заболеванием.
Оценка различных форм взаимодействия наследственности и среды позволяет считать, что наследуется генотип, но не фенотип. Свойства и признаки организмов не наследуются в готовом виде. Они формируются в процессе развития индивидуума, причем это развитие находится под контролем генов и факторов среды. Последние могут изменить проявление признаков, определяемых нормой реакции. Следовательно, каждый признак обусловлен как наследственностью, так и средой.
МЕТОДЫ И УРОВНИ ИЗУЧЕНИЯ НАСЛЕДСТВЕННОСТИ
Главным и единственным методом изучения наследственности является классический генетический (гибридологический) анализ, или, как его еще называют, формальный генетический анализ. Он заключается в последовательном разложении генома анализируемого организма на группы сцепленных генов, а групп сцепления - на генные локусы с дальнейшим установлением последовательности генных локусов вдоль хромосомных пар и выяснением тонкой структуры генов (секвенирование генов).
Генетический анализ в принципе подобен химическому анализу, задача которого заключается в разложении сложных химических соединений на более простые компоненты. Однако в отличие от хими- ческого анализа, например, нуклеопротеидов, расщепление которых на структурные части основано на гидролизе, классический генетический анализ основывается на расщеплении (сегрегации) и рекомбинации генов в мейозе и осуществляется путем скрещивания особей с разными признаками и учета результатов скрещиваний.
Схема генетического анализа организмов состоит из ряда последовательных этапов, а именно:
1. Идентификация генов.
2. Установление генных локусов на хромосомных парах.
3. Установление последовательности генных локусов вдоль хромосомных пар.
4. Выяснение тонкой структуры генов.
Результаты генетического анализа оформляют путем составления генетических карт (нуклеотидных карт).
Одним из важнейших показателей эффективности генетического анализа является его разрешающая способность, которая в общих чертах может быть аналогизирована с разрешающей способностью оптических методов исследования. Подобно тому как разрешающая способность оптических приборов (микроскопов) ограничена волновой природой света, разрешающая способность генетического анализа ограничивается количеством исследуемого потомства, получаемого в скрещиваниях, ибо чем большее количество потомства, тем шире возможность обнаружения среди них редких рекомбинантов и, следовательно, установления частоты кроссинговера.
Начиная с
На этом организме была показана «мощь» генетического анализа. Однако разрешающая способность генетического анализа всегда имеет ограничения, поскольку возможность получения большого количества потомства всегда ограничена до определенных пределов даже у тех видов, у которых оно составляет сотни организмов на пару, как, например, у D. melanogaster. Поэтому у организмов, размножающихся половым путем, в том числе и у плодовой мушки, возможно выполнение лишь трех первых этапов генетического анализа.
Однако изучение других генетических систем, в частности микроорганизмов, показало, что половая репродукция не является единственным путем, при котором осуществляются объединение, расщепление и рекомбинация генетических структур, происходящих от исходных (родительских) организмов. Эти процессы могут проходить и при других формах генетического обмена. У микроорганизмов (Е. coli), бактериальных вирусов (фагов) и микроскопических грибов такими формами генетического обмена являются трансформация, конъюгация и трансдукция. Общим для них в сравнении с половой репродукцией высших организмов служит то, что они приводят к объединению в одной клетке родительских генов и обеспечивают их расщепление и рекомбинацию, т. е., являясь альтернативами половой репродукции, представляют собой системы рекомбинации. Поэтому генетический анализ основывается и на таких системах рекомбинации. Более того, использование этих систем рекомбинации привело к повышению разрешающей способности генетического анализа в гигантских размерах, ибо появилась возможность оперировать с огромным количеством организмов в потомстве, а также легко осуществлять тесты комплементации, что позволило не только создать генетические карты ряда организмов (Е. coli, В. subtilis, фаги, низшие грибы), но и изучить тонкое строение их генов.
В качестве экспериментальных моделей широко используют также дрожжи. Являясь простейшими эукариотами, эти организмы облада- ют всеми преимуществами бактерий. Но кроме того, они оказались доступными для изучения на них генетики митохондрий, сплайсинга РНК, гаплоидии и диплоидии.
Классический генетический анализ применяют в генетике растений и животных, а также их культивируемых клеток. Однако по отношению к высшим организмам тех видов, которым присуще длительное время между генерациями и малое количество потомства на пару, он либо невозможен, либо очень затруднен. Из-за невозможности классического генетического анализа организмов ряда видов изучение их наследственности проводят с помощью других методов. Например, для изучения наследственности человека применяют метод родословных (генеалогический анализ), цитогенетический, популяционный, близнецовый и другие современные методы (см. гл. VIII).
Развитие физико-химической биологии привело к разработке методов анализа генома организмов, включая полимеразную цепную реакцию, секвенирование ДНК, выделение новых кДНК, а также методологию генной инженерии (молекулярного клонирования, генетических манипуляций), под которой понимают совокупность экспериментальных методов, позволяющих конструировать и реконструировать молекулы ДНК, т. е. создавать генотипы с заданными свойствами. Она основана на данных о свойствах ДНК и некоторых ферментов. Установлено, что ферменты эндонуклеазы, выделяемые из микробов, способны разрезать (рестриктировать) замкнутые кольцевые молекулы ДНК на линейные сегменты, причем в строго определенных участках (сайтах узнавания), т. е. из одной молекулы ДНК можно получить линейные сегменты ДНК строго определенных размеров и в определенном количестве. Эти ферменты были названы рестриктазами. Напротив, другие ферменты обладают противоположной функцией применительно к ДНК, так как способны лигировать (сшивать) в единые структуры сегменты ДНК, образованные рестриктазами. Ферменты с лигирующей способностью получили название лигаз. Последние могут «сшивать» сегменты, происходящие из разных ДНК, формируя рекомбинантную молекулу ДНК. В качестве основы для создания рекомбинантных гибридных молекул ДНК используют фрагменты (рестрикты) исследуемой ДНК и плазмиды или вирусы, которые выполняют роль так называемого генетического вектора. На конечном этапе манипулирования исследуемые
фрагменты ДНК объединяют с вектором, а затем образованные таким путем рекомбинантные молекулы ДНК вводят в бактериальные клетки, в частности в Е. coli или другие микробы. В результате репликации вектора в бактериях происходит поддержание гибридных молекул ДНК. Следовательно, клонируя клетки, содержащие гибридные молекулы ДНК, клонируют, по существу, и эти молекулы (т. е. интересующие исследователя гены). Таким образом, генетические манипуляции осуществляются здесь на генном уровне.
Известна также методология клеточной инженерии, позволяющая вести генетические манипуляции на клеточном уровне (т. е. получе- ние моноклональных антител), гибридизацию соматических клеток, оплодотворение в пробирках, выращивание растений из одной клетки (см. гл. XIV).
НУКЛЕИНОВЫЕ КИСЛОТЫ КАК ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ
В соответствии с современными представлениями генетическим материалом являются нуклеиновые кислоты.
Нуклеиновые кислоты были обнаружены в ядрах клеток в
Материалом, из которого состоят гены, является дезоксирибонуклеиновая кислота (ДНК), а материалом, обеспечивающим декодирование генетической информации, является РНК. У отдельных
вирусов первичным генетическим материалом служит рибонуклеиновая кислота (РНК). Таким образом, нуклеиновые кислоты - это хранители (носители) и переносчики генетической информации.
Сложившееся
представление о том, что первичным генетическим материалом у
абсолютного большинства живых существ является ДНК, основано на ряде
прямых и косвенных доказательств, среди которых исторически самым
первым прямым доказательством генетической роли ДНК является
установленная еще в
Решающее прямое доказательство генетической роли ДНК было обеспечено разработкой методов генной инженерии, позволившей конструировать рекомбинантные молекулы ДНК с заданными свойствами. К настоящему времени возможности генной инженерии показаны на примере клонирования многих генов самых различных организмов.
Что касается косвенных доказательств, то они известны очень давно и их несколько. Для ДНК характерна специфичность лока- лизации в клетках, поскольку она обнаруживается только в ядрах клеток (хромосомах), митохондриях (у животных) и хлоропластах (у растений). У многих микроорганизмов ДНК локализована только в ядерной области (нуклеоиде) или в цитоплазме в виде плазмид. Для организмов каждого вида характерно определенное количество ДНК на клетку (табл. 6).
Данные табл. 6 показывают, что, начиная с вирусов, содержание ДНК прогрессивно повышается у бактерий, а затем и у позвоночных и, таким образом, находится в прямой связи со сложностью органи- зации и поведения организмов. Известны, однако, исключения.
Таблица 6. Количество нуклеотидов в геномах различных организмов*
Организмы | Гаплоидный ядерный геном | Митохондриальный геном | |
Гаплоидный набор хромосом | Нуклеотидные пары | Нуклеотидные пары | |
Аденовирус | - | 3,0 ? 10-5-3,3 ? 105 | - |
Фаг Т2 | - | 2,0 ? 104 | - |
Фаг Т5 | - | 1,3 ? 105 | - |
Е. coli | - | 4,5 ? 106 | - |
Дрожжи (Saccharomyces cerevisiae) | 18 | 1,8 ? 107 | 7,4 ? 104 |
A. tholicma | - | 7,0 ? 107 | - |
Дрозофила (D. melanogaster) | 4 | 2,7 ? 107 | 1,8 ? 104 |
Лягушка (Rana pipiens) | 13 | 6,0 ? 109 | 1,6 ? 104 |
Мышь (Mus muscuhis) | - | 3,0 ? 109 | 1,5 ? 104 |
Крыса (Rattus norwegius) | 21 | 6,0 ? 109 | 1,5 ? 104 |
Человек (Homo sapiens) | 23 | 5,8 ? 109 | 1,5 ? 104 |
* 1000 пар оснований = 617 500 дальтон.
Для организмов каждого вида характерно то, что количество ДНК в соматических (диплоидных) клетках вдвое больше, чем в половых (гаплоидных). Будучи постоянным на клетку у организмов всех видов, содержание ДНК не подвержено влиянию со стороны физиологических факторов, включая пол и старение, а также неблагоприятных воздействий, например голодания, повышения или понижения температуры. Косвенным доказательством генетической специфичности ДНК являются также данные о способности искусственно синтезированных аналогов азотистых оснований ДНК вызывать наследственные изменения клеток вследствие прямого включения их в ДНК клеток и данные о том, что мутагенный эффект УФ-излучения тесно связан со спектром поглощения его молекулами ДНК.
ХИМИЯ И СТРУКТУРА ДНК
Молекулы ДНК являются линейными макромолекулами, представляющими собой длинные двойные цепи (тяжи) полимеров, составленных из мономеров, получивших название нуклеотидов (малых органи- ческих молекул) и являющихся строительными блоками ДНК.
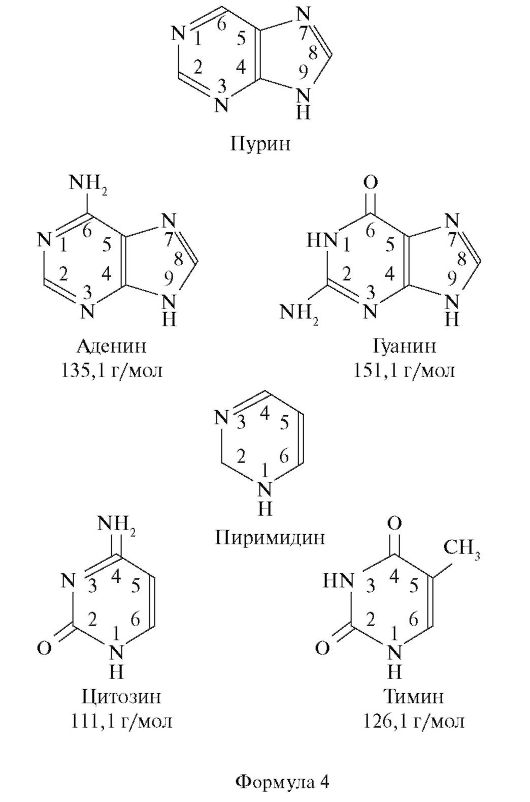
У всех живых существ макромолекулы ДНК построены по одному и тому же плану. Они слагаются в основном из одних и тех же нуклеотидов, каждый из которых содержит по одной молекуле фосфорной кислоты и сахара, а также одно из четырех азотистых оснований - аденин, гуанин, цитозин или тимин. Аденин и гуанин являются пуриновыми основаниями, тогда как тимин и цитозин - пиримидиновыми. Пурины и пиримидины называют основаниями по той причине, что в кислой среде они способны присоединять к себе ион Н+. Пиримидины являются производными шестичленного пиримидинового кольца, тогда как пурины представляют основания, у которых второе пятичленное кольцо слито с шестичленным кольцом (форм. 4).
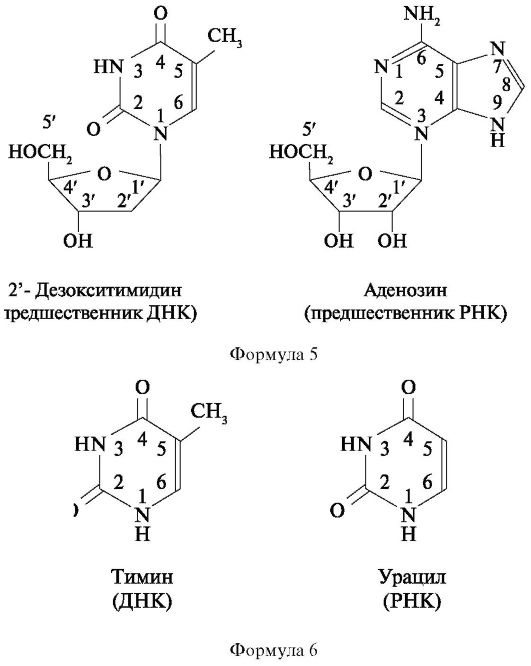
Сахаром в ДНК является 2-дезокси-Д-рибоза, отличающаяся от глюкозы тем, что в ее молекуле не 6, а 5 атомов углерода, т. е. является пятиуглеродным сахаром (пентозой). Особенность этого сахара состоит также в том, что он имеет атом водорода (Н), присоединенный к одному (специфическому) из атомов углерода, но не гидроксильную группу. Следовательно, этот сахар представляет собой дезоксирибозу, так как является рибозой, лишенной кислорода.
Сахарофосфат соединяется с азотистым основанием посредством β-гликозидной связи. Основание прикрепляется к I положению дезоксирибозы. Структура, образованная соединением азотистого осно- вания и сахара, носит название нуклеозида. Таким образом, химическими группами, которые образуют ДНК, являются пуриновые и пиримидиновые азотистые основания (аденин, гуанин, тимин и цитозин), сахар (2-дезокси-Д-рибоза) и фосфорная кислота.
РНК характеризуется такой же структурой, как и ДНК. Однако в отличие от ДНК в РНК сахаром является рибоза с кислородом, представляющая собой сахар с 5 атомами углерода, к одному из которых прикреплена 2'-гидроксильная группа (-ОН). Кроме того, в РНК тимин не имеет метильной группы и является урацилом, т. е. в РНК тимин заменен на урацил, также являющийся пиримидиновым основанием (форм. 5, 6).
Нуклеиновые кислоты называют кислотами по той причине, что их фосфатные группы освобождают в растворах ионы водорода.
Для состава ДНК характерны закономерности, известные в качестве правил А. Чаргаффа, а именно:
1. Сумма нуклеотидов, содержащих пуриновые азотистые основания, равна сумме нуклеотидов, содержащих пиримидиновые азотистые основания (А + Г = Т + Ц, или А + Г/Т + Ц = 1).
2. Содержание аденина равно содержанию тимина, а гуанина - содержанию цитозина (А = Т, или А/Т = 1 и Г = Ц, или Г/Ц = 1).
3. Содержание Г + Т = А + Ц, или Г + Т/А + Ц = 1.
4. ДНК из разных источников может иметь различия, обусловленные в одних случаях преобладанием аденина над гуанином и тимина над цитозином (А + Т > Г + Ц), в других - преобладанием гуанина и цитозина над аденином и тимином (Г + Ц > А + Т).
Данные о нуклеотидном составе оснований ДНК разных организмов приводятся в табл. 7.
Для ДНК характерна структура трех видов - первичная, вторичная и третичная.
Таблица 7. Нуклеотидный состав оснований ДНК
Организмы | Среднее значение содержания ГЦ, % |
Вирусы | 28-74 |
Бактерии | 26-74 |
Водоросли | 36-68 |
Грибы | 36-54 |
Высшие растения | 36-48 |
Простейшие | 22-62 |
Беспозвоночные | 34-44 |
Позвоночные | 40-44 |
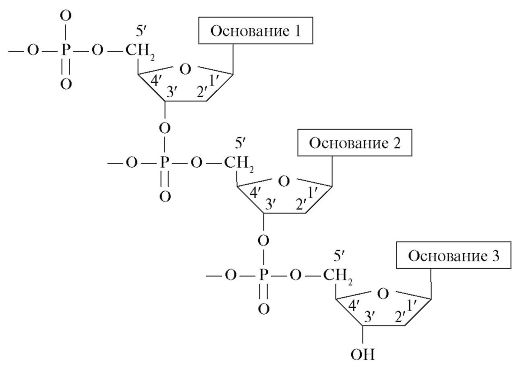
Первичная структура ДНК заключается в том, что ДНК состоит из нуклеотидных цепей, у которых скелетную основу составляют чередующиеся сахарные и фосфатные группы, объединенные ковалентными 31-, 51-фосфодиэфирными скелетными связями, а боковые группы представлены тем или иным основанием (одним из четырех) и присоединяются одна к другой молекулой сахара. Последовательно располагающиеся нуклеотиды ковалентно связаны фосфодиэфирными связями между сахарным остатком и фосфатом и в результате этого объединены в полинуклеотидную цепь. Таким образом, первичная структура ДНК (как и РНК) определяется последовательностью нуклеотидов и характером их связей между сахарным остатком и фосфатом (рис. 33, 34).
Представления о вторичной структуре ДНК (рис. 35, 36) были сформулированы Д. Уотсоном и Ф. Криком еще в
Молекула ДНК построена из двух скрученных направо спиралевидных полинуклеотидных цепей, причем каждый виток спирали соответствует 10 азотистым основаниям или расстоянию в 3,4 нм. Молекулы ДНК, цепи которых скручены направо, первоначально назвали В-формой.
Обе цепи объединены в результате закручивания одной цепи вокруг другой по общей оси. Из-за противоположной последователь- ности атомов в каждой цепи обе цепи инвертированы относительно одна другой, т. е. направление вдоль дуплекса есть 3'-- 5' для одной цепи и 5' - 3' для другой.
Формула 7
Сахарофосфатные группы располагаются на внешней стороне двойной спирали, тогда как основания находятся внутри спирали под прямым углом и вдоль ее оси. Диаметр молекулы составляет 2 нм, расстояния между отдальными азотистыми основаниями в молекуле равны 0,34 нм. Таким образом, ДНК представляет собой скрученную в правостороннем направлении двойную спираль, в которой пары азотистых оснований А-Т и Г-Ц в комплементарных полинуклеотидных цепях подобны перекладинам лестницы, а сахарофосфатные цепи являются каркасом этой лестницы.
4. Цепи в молекуле не идентичны, но комплементарны и удерживаются слабыми водородными связями между азотистыми основаниями, причем спаривание азотистых оснований для связывания цепей имеет специфический характер. Водородные связи устанавливаются не просто между азотистыми основаниями цепей, а специфически между пуриновым азотистым основанием одной цепи и пиримидиновым азотистым основанием другой. В результате этого аденин одной из цепей связывается с тимином другой цепи двумя водородными связями, тогда как гуанин одной из цепей связывается с цитозином, находящимся в другой цепи, посредством трех водородных связей.
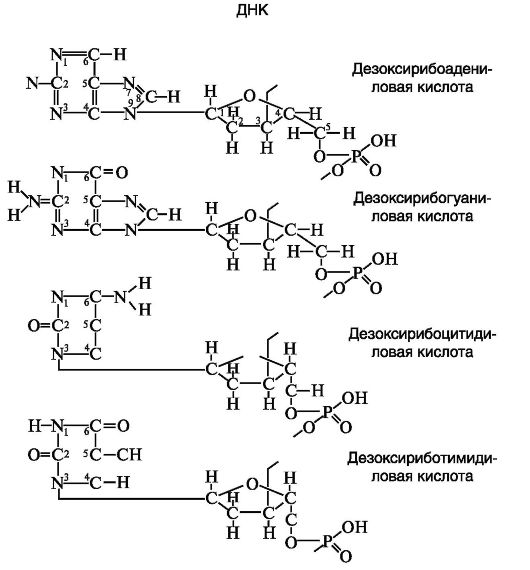
Рис. 33. Структурная формула ДНК
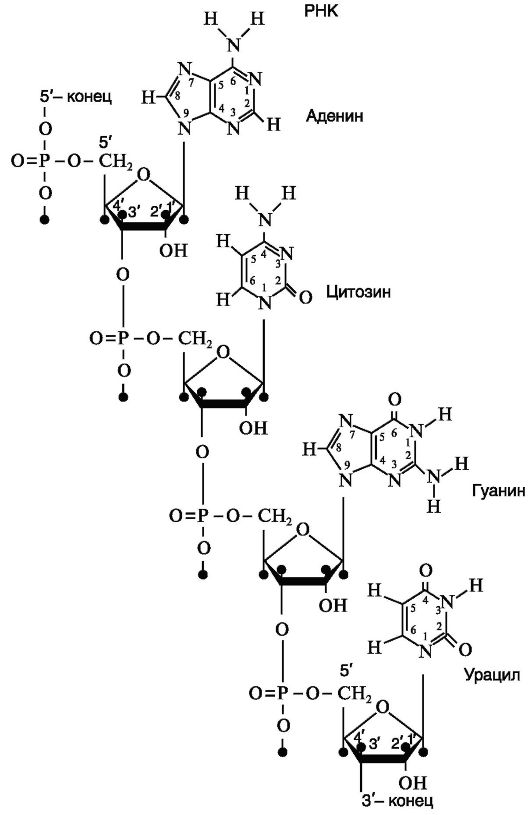
Рис. 34. Структурная формула РНК
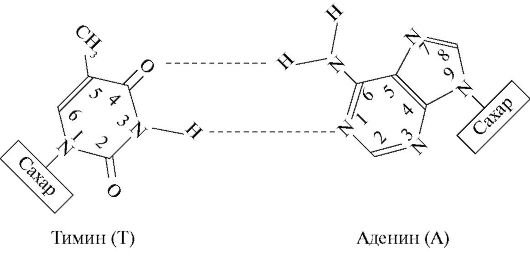
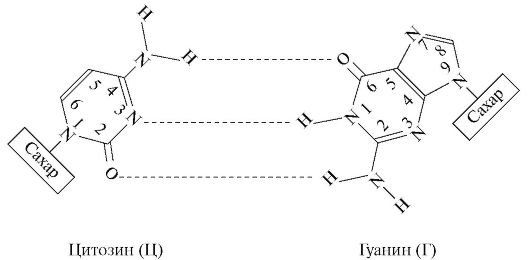
Формула 8
Дезоксирибозные остатки пар А-Т и Г-Ц разделены одинаковыми расстояниями. Для сахарофосфатных скелетных связей характерна полярность, поскольку фосфат связывает группу 3'-ОН одной дезоксирибозы с группой 5'-ОН другой, тогда как комплементарные цепи имеют противоположную полярность.
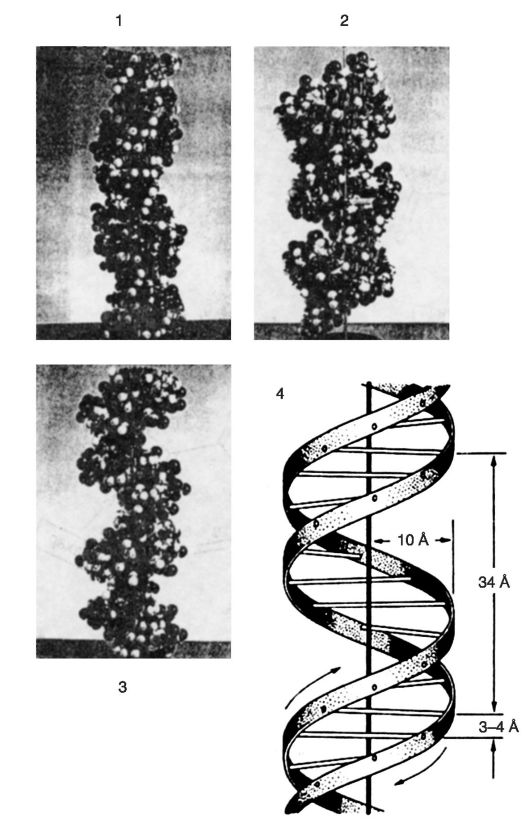
Рис. 35. Вторичная структура ДНК: 1 - В-форма; 2 - А-форма; 3 - Z-форма; 4 - размеры молекул
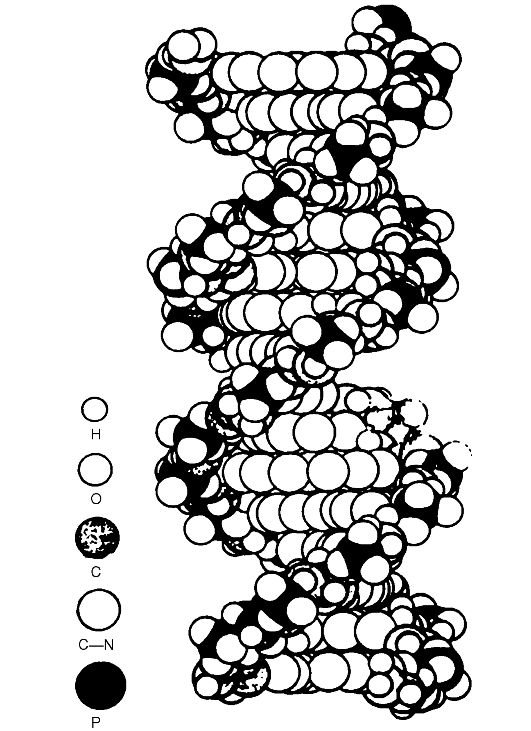
Рис. 36. Пространственная атомная модель ДНК
Двойная спираль имеет упорядоченный характер, поскольку каждая связь основание-сахар имеет одинаковое расстояние от оси спирали и перевернута на 36?. Как видно, вторичная структура отражает собой форму нуклеиновой кислоты.
Исследования рентгеновской дифракции молекул ДНК показали, что количество оснований в витках закрученной направо спирали может составлять не только 10, как у В-формы, но и 11, а то и 9,3 основания. Эти формы спиралей получили название А- и С-форм. Установлено также, что в молекулах ДНК встречаются районы, цепи в которых закручены налево. Эти районы получили название Z-форм. Различия между А-, В-, С- и Z-формами приведены в табл. 8, однако степень регулярности и конформации Z-формы еще не выяснена.
Таблица 8. Свойства разных конформационных форм ДНК
Свойство | Формы спиралей | |||
А | В | С | Z | |
Направление скрученности | вправо | вправо | вправо | влево |
Диаметр молекулы | 23 А | 19 А | 19 А | 18 А |
Количество оснований в витке | 11 | 10 | 9,3 | 12 |
Степень суперскручивания ДНК зависит от ферментов, в частности от динамического баланса между взаимоантагонистическими ферментами ДНК-гиразой, которая ответственна за суперскручи- вание, и ДНК-топоизомеразой I, которая устраняет суперскручивание.
Третичная структура ДНК связана с трехмерной пространственной конфигурацией молекул и зависит от внутримолекулярных усло- вий. Однако эта структура достаточно еще не изучена.
Размеры
молекул ДНК обычно устанавливают определением молекулярной массы в
дальтонах и длины в количестве пар оснований. Молекулярная масса пары
А-Т составляет 617 дальтон, пары Г-Ц - 618 дальтон. Молекулярная масса
1000 пар азотистых оснований (1 килобаса) составляет 617 500 дальтон,
или 6,175-105/6,02-
Препараты ДНК, выделяемой из клеток с помощью обычных методов, имеют молекулярную массу порядка 1,0-107. Длина витка по оси ДНК В-формы равна 3,4 нм. Расстояние между парами оснований в ДНК В-формы Е. coli составляет 0,34 нм.
Для
характеристики строения ДНК используют также такие физические
константы, как плотность ее при центрифугировании в градиенте тяжелых
металлов и температура плавления. Первая константа отражает
полидисперсность препаратов ДНК, тогда как вторая - их гетерогенность.
Нагревание ДНК в растворах разрывает водородные связи между основаниями в
парах и разрушает вторичную структуру ДНК, т. е. вызывает плавление
ДНК. В
Плавление ДНК есть ее денатурация. Однако замечательное свойство денатурированной ДНК заключается в том, что она способна к ренатурации in vitro, т. е. может восстанавливать двухцепочечную структуру, причем ренатурация является очень точной. Две цепи денатурированной ДНК могут ренатурировать в природную двухцепочечную спиральную форму, если их последовательности комплементарны или, другими словами, если последовательности цепей позволяют формирование пар оснований, соединенных водородными связями. Ренатурацию можно оценить и в качестве гибридизации
ДНК.
Между тем способность самокомплементарных последовательностей к гибридизации и формированию двухцепочечной спирали присуща не только ДНК, но и РНК. В результате этого in vitro можно конструировать двухцепочечные гибридные структуры РНК-РНК или РНК-ДНК. Способность нуклеиновых кислот к ренатурации имеет значение в изучении специфики отдельных последовательностей, а также в таксономии.
В зависимости от локализации ДНК в клетке различают ядерные (хромосомные) и экстраядерные (экстрахромосомные) детерминанты наследственности. Кроме того, известны транспозируемые генетические элементы (инсерционные последовательности, транспозоны и др.).
ЯДЕРНЫЕ (ХРОМОСОМНЫЕ) ДЕТЕРМИНАНТЫ НАСЛЕДСТВЕННОСТИ
С учетом эволюционного уровня организмов существует несколько форм организации ядерных генетических детерминантов.
Вирусный геном. Наиболее простой формой ядерной организации генома вообще является вирусный геном, который, как уже отмечено, очень условно часто называют вирусной хромосомой.
Геном самых малых РНК-овых вирусов представлен последовательностями, состоящими из нескольких тысяч нуклеотидов, что соответствует нескольким генам. Например, полная нуклеотидная последовательность РНК-содержащего бактериального вируса MS2 составляет 3569 нуклеотидов (3 гена). Наименьшие по размерам ДНК-содержащие вирусы состоят из большого количества пар нуклеотидов. Например, полная нуклеотидная последовательность фага ФХ174 составляет 5375 пар нуклеотидов (8 генов). Геном более крупных ДНК-овых вирусов, например фага Т2 и оспенного вируса, состоит из около 150 генов. Геном цитомегаловируса составляет 229 000 азотистых оснований.
Геном прокариот. У микоплазм, являющихся примером, возможно, самых мелких прокариот, геном составляет всего лишь несколько сот тысяч пар оснований. Например, у Mycoplasma genitalium геном составляет 580 000 пар оснований, или 470 кодирующих генетических районов. Однако лишь один генетический район участвует в кодировании белков, тогда как в контроле адгезии (прилипания) к соматическим клеткам поражаемого организма участвует 5% генома. Возможно, это связано с тем, что М. genitalium существует в ассоциации с соматическими клетками человека.
ДНК в клетках Е. coli представлена одиночной двухцепочечной кольцевой молекулой, м. м. около 2-109, что составляет примерно 30-106
пар азотистых оснований. Она выполняет роль функционально активной
хромосомы, получившей название нуклеотида. Последний является
гаплоидной структурой. Поскольку расстояние между парами азотистых
оснований в ДНК Е. coli около 0,34 нм, то контурная длина молекулы ДНК бактерий этого вида составляет около
Хромосома Е. coli содержит около 3000-4000 генов, которые организованы на основе принципа колинеарности, означающего,
что существует линейное соответствие первичной структуры гена структуре полипептидной цепи, т. е. непрерывность последовательности нуклеотидов сопровождается непрерывностью последователь- ности аминокислот в полипептидах.
Геном Haemophilus influenzae составляет 1,8-106 пар оснований, которые организованы в 1743 кодирующих района.
Молекулы ДНК прокариотов содержат последовательности азотистых оснований, которые все являются кодирующими. В геноме прокариотов каждый ген размером в несколько тысяч пар оснований повторяется лишь один раз. Исключение составляют гены для рибосомной РНК. Если прокариотическая хромосома разрывается на несколько сегментов, то каждый из них будет содержать различные последовательности.
Геном эукариотов. Геном эукариотов по своим размерам больше генома Е. coli лишь в 2-10 раз, однако ДНК в нем значительно больше. Установлено, что помимо «работающей» ДНК, т. е. помимо эксонов и интронов, в хромосомах клеток эукариот, в частности в гетерохроматиновых районах хромосом, содержится много ДНК, которая является излишней, так как не транскрибируется. Такую ДНК называют сателлитной, или «эгоистической» (от англ. selfish - эгоистичный). Количество некодирующей ДНК у разных организмов различное. Например, у дрожжей оно составляет 30% от общего количества ДНК, у насекомых - 67%, цветковых растений (арабидопсис) - 69%, нематод - 75%, у человека - 73-91%.
Примечательной особенностью некодирующей ДНК является то, что определенные последовательности оснований в ней неоднократно повторяются. Такие последовательности называют повторяющимися или повторами. Напротив, последовательности ДНК, которые не повторяются, т. е. существуют в одиночных копиях, называют уникальными. Их фракция составляет обычно 40-80%.
Длина повторов составляет обычно от нескольких до десятков или сот пар оснований, причем у эукариотов повторы, как правило, локализованы в гетерохроматиновых районах хромосом и их содержание может быть как простым, например 5'-АТАТАТ-3' в одной цепи и 3'-ТАТАТА-5' в другой, так и более сложным, например 5 -ГАААААТГА-3' в одной цепи и 3'-ЦГТТТТАЦТ-5' в другой.
Среди повторяющихся некодирующих последовательностей ДНК различают микросателлитные, мини-сателлитные и сателлитные последовательности.
Микросателлитные (простые) повторяющиеся последовательности представляют собой короткие повторы (2-5 пар оснований). Они найдены в геномах насекомых, позвоночных и растений. В геноме человека такие последовательности встречаются в эухроматиновых районах в количестве примерно 100 копий на геном.
Мини-сателлитные повторяющиеся последовательности представлены повторами, состоящими примерно из 15 пар азотистых оснований. Эти повторы обнаружены в геномах многих позвоночных и растений, включая микроскопические грибы.
Сателлитные повторяющиеся последовательности состоят из 5-10 пар азотистых оснований, а иногда даже из 100 пар. Эти последовательности обнаружены в гетерохроматиновых районах хромосом человека ближе к центромерам или в Y-хромосоме.
Биологическое значение «эгоистической» ДНК остается неясным, так как точно не выявлены необходимость в этих сегментах ДНК и причины, поддерживающие их в составе кодирующей ДНК.
Биологическое значение повторов в «эгоистической» ДНК тоже не выяснено. Однако предполагают, что они вовлечены в регуляцию экспрессии и рекомбинацию генов, а также в «защиту» некоторых структурных генов, в частности генов, детерминирующих синтез гистонов, рРНК или рибосомных белков.
Для ДНК эукариотических клеток характерно и наличие палиндромов, т. е. обращенных повторов. Они встречаются в огромном количестве копий (возможно, в тысячах копий), причем их длина различна. Функции длинных палиндромов неизвестны. Что касается коротких палиндромов, то они служат сайтами узнавания ферментами (рестриктазами).
Повторяющиеся последовательности реплицируются в геномах благодаря совместной репликации с хромосомами. Реплицируются и хромосомы, и повторяющиеся последовательности.
Большинство структурных генов представлено однокопийными последовательностями ДНК, т. е. существуют в одной копии. Некоторые гены присутствуют в 1-3 копиях, но известны гены, которые представлены множественными копиями.
Поскольку
у эукариот ДНК содержится в каждой хромосоме, а каждая хромосома
представлена в двух (диплоидных) или более (полиплоидных) копиях, то
количество ДНК в хромосомах зависит от их плоидности. Например,
гаплоидный набор хромосом половых клеток человека содержит ДНК длиной
этой ДНК
разделяется между 23 хромосомами, для которых характерны разные размеры
и формы. Следовательно, в каждой хро- мосоме содержится ДНК длиной от
15 до
Гетерогенность длины нуклеосомной ДНК определяется вариабельностью в длине ДНК, сцепливающей одну нуклеосому с другой. Какова роль негистоновых белков, обычно обнаруживаемых в хроматине, - это вопрос, который подлежит выяснению.
В случае прокариотов между генами последовательности азотистых оснований ДНК и полипептидной цепью существует колинеарность. Однако у эукариотов гены вдоль хромосом располагаются не непре- рывно, поскольку они разделены другими последовательностями ДНК. Поэтому колинеарности в случае эукариотов не существует. Кроме того, в геноме эукариотов различают два типа последовательностей - транскрибируемые и транслируемые последовательности, которые определяют первичную структуру белков и называются эксонами, и транскрибируемые, но не транслируемые («молчащие») последовательности, называемые нитронами. Следовательно, гены содержат как эксоны, так и интроны (см. гл. VII). Можно сказать, что для генов эукариотов характерна мозаичность, которая имеет место практически на протяжении всего генома.
ЭКСТРАЯДЕРНЫЕ (ЭКСТРАХРОМОСОМНЫЕ)
ДЕТЕРМИНАНТЫ НАСЛЕДСТВЕННОСТИ
Длительное время считали, что ДНК содержится только в ядрах клеток, и вся наследственность понималась в качестве ядерной. Между тем с развитием молекулярно-генетических методов исследований стали обнаруживать ДНК, находящуюся за пределами ядра как у прокариотов, так и в клетках эукариотов. Эта ДНК получила название экстраядерной (экстрахромосомной) ДНК, а контролируемая такой ДНК наследственность - экстраядерной, или экстрахромосомной.
Перечень форм экстраядерных (экстрахромосомных) ДНК прокариотов и эукариотов приведен ниже.
1. ДНК плазмид: бактерии, низшие грибы и другие организмы.
2. ДНК органелл: митохондрии, хлоропласты, кинетопласты.
3. ДНК амплифицированных генов: гены, контролирующие синтез отдельных белков.
4. Малые полидисперсные кольцевые и линейные ДНК: экстрахромосомные копии повторяющихся (часто транспозируемых) последовательностей ДНК.
Плазмиды. Плазмиды встречаются в цитоплазме как прокариотов, так и эукариотов, причем у бактерий они являются обычными обитателями. В частности, они идентифицированы почти у всех видов бактерий, имеющих медицинское (являющихся возбудителями болезней) или сельскохозяйственное и промышленное значение.
Плазмиды бактерий - это генетические структуры, находящиеся в цитоплазме и представляющие собой молекулы ДНК размером от 2250 до 400 000 пар азотистых оснований. Они существуют обо- собленно от хромосом в количестве от одной до нескольких десятков копий на одну бактериальную клетку. Различают три типа бактериальных плазмид: факторы генетического переноса, коинтегративные и неконъюгативные плазмиды.
Факторы переноса обладают лишь генами репликации и переноса. Благодаря генам репликации такие плазмиды способны к бесконечно долгому поддержанию и воспроизводству в автономном (экстрахромосомном) состоянии, а благодаря генам переноса - к передаче от одних клеток к другим, часто преодолевая в скрещиваниях видовые и родовые барьеры. Бактерии, содержащие плазмиды этого типа, служат генетическими донорами. Они способны вступать в скрещивание с клетками, не содержащими плазмиды.
Коинтегративные плазмиды представляют собой фактор генетического переноса, сцепленный с генами, контролирующими синтез тех или иных белков, имеющих значение для бактерий. Например, плазмиды R контролируют синтез ферментов, придающих бактериям устойчивость к антибиотикам, сульфаниламидам и другим лекарственным веществам, плазмиды Ent - синтез энтеротоксинов, Col - колицинов, Шу - гемолизинов. Известны также плазмиды, контролирующие разрушение многих органических соединений и другие свойства. Благодаря фактору переноса эти плазмиды конъюгативны.
Неконъюгативные плазмиды - это плазмиды, которые не передаются от одних клеток к другим, так как они не обладают фактором
переноса. Они тоже детерминируют лекарственную устойчивость и другие свойства бактерий. Передача неконъюгативных плазмид от одних бактерий к другим обеспечивается содержащимися в клетках факторами переноса или коинтегративными плазмидами, которые мобилизуют их на перенос.
Среди эукариотов плазмиды идентифицированы у низших грибов. Одна из таких плазмид у дрожжей S. cerevisiae представляет собой кольцевые молекулы ДНК размером 6318 пар оснований, существующие в количестве 80 копий на гаплоидный геном и кодирующие белки, необходимые для собственной репликации и рекомбинации. У нейроспоры (Neurospora) плазмиды обнаружены в виде малых кольцевых молекул ДНК размером 4200-5200 пар оснований, встречающихся в количестве около 100 копий на гаплоидный геном, а у плесени Aspergilus niger - в виде кольцевых молекул ДНК размером около 13 500 пар оснований в количестве около 100 копий на клетку.
ДНК органелл. ДНК этого класса обнаружены в случае как низших, так и высших эукариотов.
Молекулы ДНК, выделяемые из митохондрий соматических клеток животных и хлоропластов клеток растений, характеризуются небольшими размерами. Например, размеры молекул ДНК (геномов) митохондрий (мтДНК) разных животных (включая плоских червей, насекомых и млекопитающих) составляют 15 700-20 000 пар азотистых оснований, человека - 16 569 пар азотистых оснований. У простейших, например у трипаносом и парамеций, мигохондриальный геном равен 22 000 и 40 000 пар оснований соответственно. Геном хлоропластов у высших растений составляет 12 000-200 000 пар оснований, у дрожжей - 78 000 пар оснований, у зеленых водорослей - 180 000 пар азотистых оснований. Во многих случаях показано, что ДНК митохондрий и хлоропластов сплошь состоит из нуклеотидных последовательностей, гомологичных последовательностям хромосомной ДНК.
Митохондриальный геном человека состоит из 13 генов, нуклеотидная последовательность которых определена и для которой характерно полное или почти полное отсутствие некодирующих участков. Эти гены кодируют собственные рибосомные рРНК (12 S и 16 S) и 22 разные транспортные РНК, а также разные полипептиды, включая субъединичные компоненты I, II, III, IV и V оксидазы цитохрома С, субъединицы 6 АТФ-фазы, цитохрома В и девяти дру- гих белков, функции которых неизвестны (см. гл. VIII).
Геном хлоропластов ряда высших растений состоит из 120 генов. Они кодируют 4 рибосомных РНК, 30 рибосомных белков, часть субъединиц хлоропластной РНК-полимеразы, часть белков, содержащихся в фотосистемах I и II, белковые субъединицы АТФ-синтетазы и отдельных ферментов цепи транспорта электронов, а также белковую субъединицу рибулозобисфосфаткарбоксилазы и очень многих тРНК. Хлоропластный геном очень сходен с бактериальным геномом как по организации, так и функциям.
В митохондриальном геноме человека, вероятно, отсутствуют интроны, но в ДНК хлоропластов некоторых высших растений, а также в ДНК митохондрий грибов интроны обнаружены.
Считают, что хлоропластные геномы высших растений остаются без изменений примерно несколько миллионов лет. Возможно, что такая древность характерна и для митохондриальных геномов млекопитающих, включая человека.
Характер передачи мтДНК по наследству у организмов различен. Например, у дрожжей в результате одинакового вклада мтДНК сливающимися гаплоидными клетками в зиготу митохондриальный геном наследуется потомством от обоих родителей. Между тем показано, что у D. melanogaster и мышей мтДНК передается по материнской линии. По данным посемейного распределения ДНК в больших семьях, предполагают, что мтДНК у человека также наследуется по материнской линии. У человека при оплодотворении в яйцеклетку передается только ядро сперматозоида. Следовательно, оплодотво- ренная яйцеклетка содержит лишь женские митохондрии (митохондриальные гены).
ДНК, обнаруживаемая в кинетопластах трипаносом, представлена малыми (2500 п. о.) и крупными (3700 п. о.) кольцевыми молекулами.
ДНК амплифицированных генов. Эта ДНК встречается в форме экстрахромосомных кольцевых молекул. Например, когда эукариотические клетки культивируют в средах с лекарственными веществами, то происходит селекция резистентных клеток с повышенным количеством копий гена, контролирующего резистентность. Клетки многих опухолей содержат также экстрахромосомные амплифицированные гены (наряду с хромосомными).
Малые полидисперсные кольцевые и линейные ДНК. Молекулы ДНК этого типа (мпкДНК) имеют размеры от нескольких сот до десятков тысяч нуклеотидных пар и встречаются как в цитозоле, так и в ядре и митохондриях клеток многих организмов-эукариотов. Эти моле-
кулы ДНК происходят или связаны с ДНК хромосом и органелл. Многие из этих молекул ДНК способны к транспозиции.
ТРАНСПОЗИРУЕМЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ
Транспозируемые (подвижные, мигрирующие, транслоцируемые) генетические элементы - это сегменты ДНК, способные к переме- щению в пределах одного генома или с одного генома на другой.
У прокариотов транспозируемые генетические элементы представлены сегментами ДНК двух типов - инсерционными последо- вательностями (IS) и транспозонами (Тп).
Инсерционные последовательности ДНК представляют собой последовательности, состоящие из 768-5000 пар азотистых оснований. Они обнаружены в плазмидах, фагах, бактериальных хромосомах, причем встречаются так часто, что многие исследователи считают их нормальными компонентами бактерий. Инсерционные после- довательности (ISI, IS2, IS4, IS5, IS102 и др.) в большинстве своем многокопийны. Они перемещаются с высокой частотой. Их миграция происходит на основе генетической рекомбинации.
Транспозоны организованы значительно сложнее, нежели инсерцион- ные последовательности. В упрощенном виде можно сказать, что транспозон представляет собой сегмент ДНК, середина которого представлена геном или генами устойчивости, а фланги - инсерционными последовательностями, обеспечивающими его передвижение. Размеры транспозонов - 2000-20 500 пар азотистых оснований. Для транспозонов характерны значительные инвертированные повторы.
Транспозируемые элементы клеток-прокариотов перемещаются по маршруту хромосома -- плазмида -- другая плазмида -- хромо- сома. Перемещение транспозонов обеспечивается специализированным репликативным процессом, который не связан с генерацией экстрахромосомных форм. В экспериментальных условиях любой транспозон можно включить практически в любую плазмиду.
Генетические элементы, сходные с транспозируемыми, существуют также в клетках эукариотов, где они представлены разными повторяющимися последовательностями ДНК. Одни из этих элементов транспозируются в результате повторного включения (реинсерции) в геном продукта реверсивной транскрипции (копии РНК). Такие элементы получили название ретроэлементов. Напротив, другие элементы транспозируются прямо через копии ДНК.
Наиболее известными ретроэлементами являются ретротранспозоны с короткими терминальными повторами, такие как ретротранспозоны I или R2 у дрозофил, Line у млекопитающих, Ingi у трипаносом. Копии этих ретротранспозонов кодируют белки, необходимые для обратной транскрипции, т. е. их транспозиция осуществляется с использованием РНК в качестве интермедиата. К этой категории ретроэлементов принадлежат также последовательности с длинными терминальными повторами, в частности последовательности copia и gypsi у дрозофил, 7у~фактор дрожжей и LI-элементы у млекопитающих. У этих последовательностей повторы достигают 500 пар оснований. Наконец, ретротранспозонами многие считают также последовательности, которые, помимо инсерционной способности, обладают инфицирующими свойствами, например отдельные ретровирусы (лейкоза птиц, лейкемии млекопитающих, иммунодефицита млекопитающих).
Элементы, которые сходны по транспозиции с транспозоном Tnl, транспозируются прямо через копии ДНК, характеризуются инвер- тированными терминальными повторами и открытыми рамками чтения, кодирующими фермент транспозазу. К таким элементам принадлежат транспозоны Р и hobo у дрозофил, Ас, ДЗ и Ми у растений кукурузы, Тс I у нематод, TU у морских ежей.
В геноме человека найдена последовательность AIu длиной порядка 300 пар оснований и повторяющаяся в 100 000-300 000 копиях на гаплоидный набор хромосом, что составляет около 5% генома человека. Alu-последовательности сходны с прямыми копиями ДНК на молекулах мРНК, ибо они содержат «отрезок» полидезоксиаденозина на их 3'-концах, а сходство Alu-последовательностей с транспозонами определяется тем, что они фланкированы прямыми повторами 7-20 пар оснований.
В геноме человека открыты тандемно расположенные повторяющиеся последовательности Hinf, составленные из субъединиц длиной 172 и 147 пар оснований, а также транспозоны Mariner, тоже представленные повторами оснований. Транспозоны Mariner обнаружены, кроме того, в геномах дрозофилы, отдельных членистоногих, нематод и планарий.
Биологическое значение транспозонов заключается прежде всего в том, что они являются мутагенами.
РЕПЛИКАЦИИ ДНК И ХРОМОСОМ
Все, что известно в настоящее время о репликации ДНК, выяснено в результате многолетнего экспериментального обоснования основных положений модели структуры и репликации ДНК по Д. Уотсону и Ф. Крику (1953).
Формулируя свою модель, Д. Уотсон и Ф. Крик предположили, что репликация ДНК происходит в несколько последовательных этапов, а именно: а) разрыв водородных связей между двумя полинуклеотидными цепями и разделение последних; б) разматывание полинуклеотидных цепей; в) синтез вдоль каждой из полинуклеотидных цепей новой цепи с комплементарной последовательностью азотистых оснований (рис. 37). Они предположили далее, что разделение и разматывание полинуклеотидных цепей начинается с одного конца молекулы, продолжается по направлению к другому ее концу и сопровождается одновременно идущим с того же конца молекулы синтезом новых полинуклеотидных цепей. Таким образом, в репликации ДНК каждая полинуклеотидная цепь действует в качестве шаблона для вновь синтезируемой полинуклеотидной цепи, причем шаблон обеспечивает выбор определенных нуклеотидных последовательностей из всех возможных последовательностей. В результате этого каждая новая молекула ДНК состоит из одной старой цепи и одной новой (дочерней), комплементарной старой. Этот способ репликации ДНК получил название полуконсервативной репликации.
Полуконсервативный характер репликации ДНК был доказан М. Месельсоном и Ф. Сталем в
У прокариотов репликация ДНК начинается с 0-пункта репликации, составленного примерно 300 нуклеотидами, и продолжается в двух направлениях, образуя репликационную «вилку». Скорость движения «вилки» (т. е. скорость полимеризации) составляет 500 нуклеотидов в секунду. Удвоение молекулы ДНК происходит за 40 мин. Кроме того, у прокариотов действует механизм «вращающееся колесо», по которому репликационная «вилка» двигается
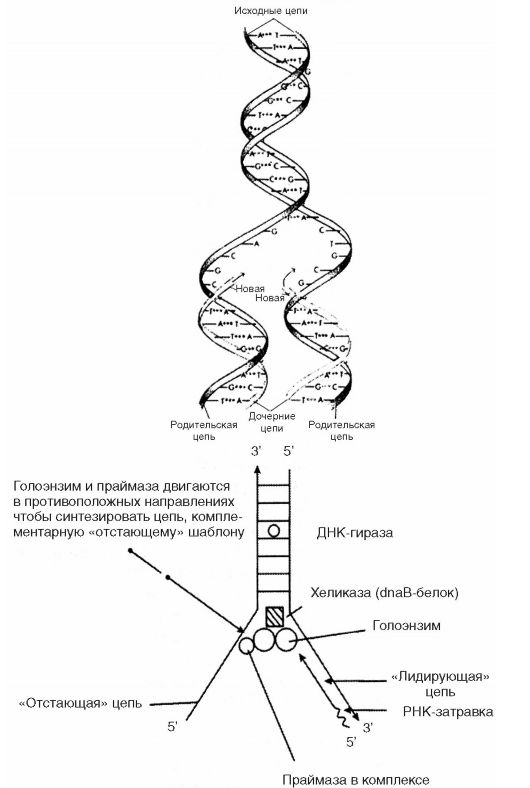
Рис. 37. Репликация ДНК по Д. Уотсону и Ф. Крику
вокруг кольца, генерируя цепи, на которых синтезируются комплементарные цепи.
Изучение ферментативного синтеза ДНК in vitro, компонентами которого являются ДНК-полимераза, дезоксирибонуклеозид 5'-три- фосфаты всех четырех азотистых оснований, ионы магния и ДНК- «затравка», показало, что присутствие всех этих компонентов в смеси сопровождается добавлением мононуклеотидов к растущему концу цепи ДНК, причем они добавляются к 3' -гидроксильному концу «затравочной» последовательности, и цепь растет в направлении от 5' - к 3' -концу. Реакция катализируется ДНК-полимеразой III. После добавления в смесь ДНК-«затравки» синтез ДНК не прекращается даже тогда, когда количество вновь синтезированной ДНК достигает количества ДНК-«затравки». Если же один из компонентов в смеси отсутствует, частота полимеризации снижается во много раз. Отсутствие ДНК-«затравки» полностью исключает реакцию.
Установлено, что для репликации ДНК Е. coli in vitro необходимы белки, детерминируемые генами dna A, dna В, dna С, dna G (белок dna А, белок хеликаза, белок dna С, белок праймаза соответственно, ДНК-гираза, а также белок, связывающийся с одиночными цепями ДНК и АТФ). Комплекс репликативных ферментов и белков получил название ДНК-репликазной системы (реплисомы).
Изучение ферментативного синтеза ДНК in vitro показало также, что копируются обе цепи, но так как цепи ДНК в спирали антипараллельны, то синтез (полимеризация) одной цепи происходит в направлении от 5'- к 3'-концу, тогда как другой - от 3'- к 5'-концу. Синтез цепи в направлении от 5'- к 3'-концу является непрерывным, тогда как синтез в направлении от 3' - к 5' - - прерывен, поскольку синтезируются короткие сегменты в направлении от 5' - к 3' -концу, которые затем воссоединяются ДНК-лигазой. Короткие сегменты по 1000-2000 нуклеотидов получили название фрагментов Р. Оказаки (рис. 38). Следовательно, рост обеих цепей обеспечивается одной и той же полимеразой (голоэнзим ДНК-полимеразой III). Репликационная «вилка» асимметрична. Цепь, синтезируемая непрерывно, называют лидирующей, а цепь, син- тезируемую непрерывно, - запаздывающей. «Запаздывание» второй цепи связано с тем, что синтез каждого фрагмента Оказаки осуществляется только тогда, когда в результате продвижения лидирующей цепи откроется необходимый участок цепи-шаблона.
У бактерий открыты ДНК-полимеразы I, II, III. Главной является ДНК-полимераза III, которая отвечает за элонгацию цепей ДНК.
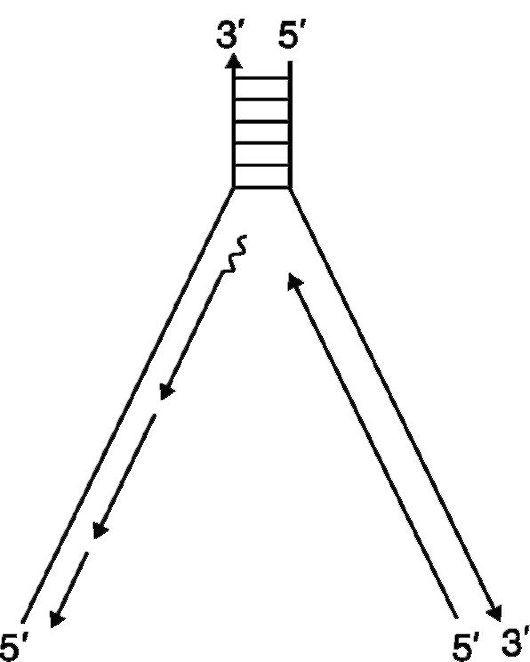
Рис. 38. Прерывистость репликации ДНК (синтез фрагментов Оказаки)
Что касается других ферментов, то ДНК-полимераза I заполняет бреши в запаздывающей цепи, тогда как функция ДНК-полимеразы II не совсем понятна.
В случае синтезлидирующей цепи у ДНК-полимеразы имеется спаренный 3' -конец, что позволяет начать полимеризацию следующей (новой) цепи. Однако для ДНК-полимеразы, синтезирующей «запаздывающую» цепь, необходима «затравка», обладающая спаренным 3'-концом (3'-гидроксильной группой). Эту затравку в виде коротких сегментов РНК синтезирует из рибонуклеозидтрифосфатов ДНК-праймаза на ДНК-шаблоне запаздывающей цепи. Данный процесс характерен тем, что предшествующий синтез коротких сегментов РНК «затравливает» каждую новую инициацию синтеза ДНК. Затем включается ДНК-полимераза, полимеризуя 5'-фосфат дезокси- рибонуклеотидного остатка с 3' -гидроксильным концом цепи РНК, что приводит к нормальному синтезу цепи ДНК. В последующем «затравочная» последовательность РНК удаляется и брешь заполняется ДНК. Таким образом, роль «затравки» в синтезе фрагментов Оказаки выполняет РНК.
Репликация ДНК эукариотов характеризуется теми же механизмами, что и у прокариотов, хотя скорость полимеризации цепей является меньшей (около 50 нуклеотидов в секунду у млекопитающих). В репликации ДНК эукариотов принимают участие те же ферменты,
что и в случае прокариотов. Размеры фрагментов Оказаки составляют 100-200 нуклеотидов.
Раскручивание двойной цепи ДНК происходит с участием трех разных белков, а именно: а) белки, дестабилизирующие спираль (SS В-белки). Они связываются с одноцепочечными ДНК, помогают ДНК-геликазам раскручивать спираль и обеспечивают протяженный одноцепочечный шаблон для полимеризации; б) ДНК-геликазы, раскручивающие ДНК. Они прямо вовлечены в катализирование раскручивания; в) ДНК-гиразы, которые катализируют формирование негативных супервитков в ДНК.
У эукариотов известно пять ДНК-полимераз (α, β, γ, δ и ε), из которых главную роль в репликации играют полимеразы α и δ. Начинает α-полимераза.
Синтез из ведущей (лидирующей) и запаздывающей цепей, поскольку только она обладает «затравочной» активностью. Дальнейшую элонгацию лидирующей цепи осуществляет δ-фермент, а «запазды- вающей» цепи - ε- или α-ферменты. Завершает репликацию «запаз- дывающей» цепи γ-фермент, который является митохондриальным. При этом он играет роль, присущую в бактериях ферменту poli. Роль β-фермента неясна.
Установлен также белок (циклин), который синтезируется в ^-фазе клеточного цикла и также необходим для репликации ДНК.
Спирализацию ДНК после репликации обеспечивают ферменты ДНК-топоизомеразы. Процесс репликации ДНК характеризу- ется исключительной точностью. Как отмечено выше, фрагменты Оказаки, продуцируемые в ДНК у эукариотов, имеют длину от 100 до 20 пар нуклеотидов. Это, возможно, связано с тем, что у эукариотов синтез ДНК более медленный (1 молекула ДНК в минуту) по сравнению с прокариотами (30 молекул в минуту).
Удвоение хромосом у эукариотов является сложным процессом, поскольку включает не только репликацию гигантских молекул ДНК, но и синтез связанных с ДНК пистонов и негистоновых хромосомных белков. Конечный этап - упаковка ДНК и гистонов в нуклеосомы. Считают, что удвоение хромосом также имеет полуконсервативный характер.
Репликационное поведение хромосом обосновывается на трех фундаментальных свойствах, а именно: непосредственно репликация, сегрегация хромосом при репликации ДНК и делении клеток, а также репликация и предохранение концов хромосом.
0-пункты репликации существуют в хромосомах (сайты инициации репликации) также организмов-эукариотов, состоящих из определенных последовательностей азотистых оснований, причем являются множественными. Эти пункты получили название автономно реплицирующихся последовательностей (ARS-элементов). Определение количества репликационных «вилок» показало, что они удалены один от другого на расстояние 30 000-300 000 пар азотистых оснований. В результате этого по каждой хромосоме двигаются много репликационных «вилок», причем одновременно и независимо одна от другой. Инициацию репликации ДНК обеспечивают белки, связанные с 0-пунктом репликации, а также белки-киназы. Последние ответственны за выход ДНК из репликации. Но как действуют эти механизмы - это вопрос, который еще не получил разрешения.
За сегрегацию хромосом в дочерние клетки ответственны центромеры.
В репликации и предохранении концов хромосом имеют значение так называемые теломеры, представляющие собой повторяющиеся последовательности ДНК длиной 5-10 азотистых оснований. Их роль заключается в обеспечении доступа ДНК-полимеразы к концам цепей ДНК. Вновь образованные хромосомы содержат как старые гистоны, так и вновь синтезированные, контроль которых у млекопитающих осуществляется 20 генными блоками, каждый из которых содержит по 5 гистоновых генов.
Однако репликация эукариотической ДНК имеет и существенное отличие от репликации прокариотической ДНК. Когда ДНК эукариотов метят 3Н-тимидином, а затем экстрагируют из хромосом и изучают методом радиографии, то при этом наблюдают тандемный порядок радиоактивности. Это свидетельствует о том, что одиночные молекулы ДНК содержат множественные 0-пункты репликации. Например, в ДНК клеток млекопитающих 0-пункты встречаются через каждые 40 000-200 000 пар оснований. Экспериментальные данные указывают на то, что репликация хромосомы эукариотов происходит в двух направлениях, поскольку репликационные «вилки» двигаются в двух направлениях из центральных 0-пунктов к репликационным терминусам (пунктам остановки репликации). Сегмент хромосомы, чья репликация находится под контролем одного 0-пункта и двух тер- минусов, является единицей репликации и ее называют репликоном. Размеры эукариотических репликонов зависят от вида организмов, но в общем они составляют около 10-100 нм.
ВОПРОСЫ ДЛЯ ОБСУЖДЕНИЯ
1. В чем заключается классический генетический анализ и применим ли он для изучения наследственности всех организмов?
2. На чем основаны представления о том, что генетическим материалом являются нуклеиновые кислоты?
3. Существует ли связь между размерами генома (в количестве нуклеотидных пар) и видовой принадлежностью организмов? Приведите примеры в обоснование своей точки зрения.
4. Что вы знаете о путях увеличения генома клеток в процессе развития организмов от низших форм к высшим?
5. Определите в сантиметрах общую длину ДНК в клетках человека.
6. ДНК стабильна при рН 11, но РНК деградирует до нуклеотидов при щелочной реакции. Используя учебники по биохимии, объясните причину этого явления.
7. Если субъединицы (β и β' РНК-полимеразы составляют 0,005 доли от массы общего белка в клетках E. coli, то сколько будет в клетке молекул РНК-полимеразы при условии, что каждая субъединица β и β' представляет цельную молекулу этого фермента?
8. Почему мочевина денатурирует РНК?
9. Состав оснований (фракция Г + Ц) двухцепочечной молекулы ДНК отражается на показателях плавучей плотности в хлориде цезия и температуры плавления (Тп), при которой половина молекул «плавится» на отдельные цепи. Было найдено, что плавучая плотность равна 1,660 + (0,093 χ фракция Г + Ц), фракция Г + Ц = 2,44 (Тп - 69,3), причем Тп определена в стандартном солевом растворе. Плавучая плотность ДНК крысы составляет 1,702, D. melanogaster - 1,698 и дрожжей - 1,699. Определите фракцию Г+Ц и температуру плавления для ДНК каждого вида.
10. Каково значение митохондриальной ДНК у человека?
11. Что представляют собой транспозируемые генетические элементы? Как их классифицируют?
12. Что представляют собой плазмиды?
13. Что представляют собой повторяющиеся последовательности ДНК и как часто они повторяются в геноме человека?
14. В чем заключается полуконсервативный способ репликации ДНК и каково биологическое значение такого способа репликации?
15. Какова роль ферментов в репликации ДНК?
16. Есть ли разница между репликацией ДНК и репликацией хромосом?
17. Что такое нуклеосома и каковы ее размеры? Какова роль белков в упаковке ДНК в хромосомы?
18. Вычислите число нуклеотидных пар в 1 мегадальтоне двухцепочечной ДНК.
19. Как вы думаете, сколько генов имеется в одной клетке человека при условии, что длина одного гена составляет около 500 пар нуклеотидов?
20. По данным кислотного гидролиза, препарат ДНК, выделенной из клеток мертворожденного плода человека, характеризовался следующим составом (в %): аденин - 25, тимин - 32, гуанин - 22, цитозин - 21. Каким образом можно объяснить этот необычный результат исследования, руководствуясь данными о структуре ДНК?
21. Какими будут длина и суммарная масса ДНК, если в ней объединить молекулы ДНК из всех клеток новорожденного ребенка, организм которого состоит из 2-1012 клеток?
22. Что представляют собой фрагменты Оказаки и какова их роль в репликации ДНК?
23. Можете ли вы назвать экспериментальные данные, подтверждающие антипараллельную ориентацию цепей в молекуле ДНК?
24. После денатурации ДНК ренатурировали, позволив регибридизацию цепей до 1% их последовательностей. Затем эту ДНК подвергли обработке нуклеазой I до полного переваривания молекул, после чего ее «разогнали» путем электрофореза в агарозном геле. Каковы результаты электрофореза?
25. Каким образом можно определить генетическую локализацию ^-последовательностей в геноме Е. coli?
26. Почему наследование митохондриальных генов у человека идет по материнской линии?
27. Насколько связана репликация ДНК с хромосомами в клетках человека?