Биология: учебник / Пехов А.П., -, 2010. - 664 с.
|
|
|
|
ГЛАВА IX ТЕОРИЯ ЭВОЛЮЦИИ
Масса Земли составляет около 4?1018 тонн, возраст - около 4,5-5 млрд лет. Жизнь возникла на Земле около 3,9 млрд лет назад. Возникнув, она оказала существенное влияние на Землю и ее атмос- феру, которая изменялась от окисляющей к неокисляющей.
Огромное разнообразие живых форм, населяющих сейчас Землю, является результатом длительной эволюции (от лат. evolution - раз-
вертываю), под которой понимают развитие организмов во времени или
процесс исторического преобразования на Земле. В итоге этого процесса
возникло многообразие современного живого мира. Термин «эволюция» был
введен в науку в
Современные представления о происхождении жизни основываются на теории эволюции, сама же теория эволюции основывается на данных об общих закономерностях и движущих силах исторического развития живой природы. Одной из характерных особенностей эволюции является длительность сроков ее для первых живых существ. Например, микроорганизмы были единственными живыми обитателями Земли в течение 3 млрд лет. Затем эволюция пошла быстро. Многоклеточные появились лишь после 4/5 времени существования Земли (в палеозойской эре). Эволюция человека составила всего лишь несколько последних миллионов лет.
Эволюция протекает на всех уровнях организации живой материи и на каждом уровне характеризуется новообразованием структур. Объединение структур одного уровня сопровождается переходом системы на более высокий эволюционный уровень.
ВЗГЛЯДЫ НА ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЮ
ЖИЗНИ В ПРОШЛОМ
Проблемы происхождения и эволюции жизни на Земле принадлежат к числу величайших проблем естествознания. Эти проблемы
привлекали к себе внимание человеческого ума с самых незапамятных времен. Они являлись предметом интереса всех философских и религиозных систем. Однако в различные эпохи и на разных ступенях развития человеческой культуры проблемы происхождения и эволюции жизни решались по-разному, причем всегда были ареной борьбы материализма и идеализма.
Самыми древними взглядами на природу были мистические, по которым жизнь связывали с силами природы. Но уже у самых истоков культуры в Древней Греции на смену мистическим истолкованиям природы приходят начала других представлений. В тот период возникла и стала развиваться доктрина абиогенеза, или спонтанного самозарождения, в соответствии с которой признавалось, что живые организмы возникают спонтанно из неживого материала. Тогда же появились и эволюционные идеи. Например, Эмпедокл (490-430 гг. до н.э.) считал, что первые живые существа возникли из четырех элементов живой материи (огонь, воздух, вода и земля) и что для природы характерно закономерное развитие, выживание тех организмов, которые наиболее гармонично (целесообразно) устроены. Эти мысли были очень важными для дальнейшего распространения идеи о естественном происхождении живых существ.
По мнению Демокрита (460-370 гг. до н.э.), мир состоит из множества мельчайших частиц, которые находятся в движении, и жизнь является не результатом творения, а результатом действия механических сил самой природы, приводящих к самозарождению. По Демокриту, самозарождение живых существ происходит из ила и воды в результате сочетания атомов при их механическом движении, когда мельчайшие частицы влажной земли встречаются и соединяются с атомами огня. Самозарождение представлялось случайным процессом.
Предполагая, что черви, клещи и другие организмы возникают из росы, ила, навоза, волос, пота, мяса, моллюски из влажной земли, а рыбы из морской тины и т. д., Платон (427-347 гг. до н.э.) утверждал, что живые существа образуются в результате соединения пассивной материи с активным началом (формой), представляющим собой душу, которая затем движет организмом.
Аристотель (384-322 гг. до н.э.) утверждал, что растения и животные возникают из неживого материала. В частности, он утверждал, что некоторые животные появляются из разложившегося мяса. Признавая реальность материального мира и постоянство его движе-
ния, сравнивая организмы между собой, Аристотель пришел к заключению о «лестнице природы», отражающей последовательность организмов, начинающуюся с неорганических тел и продолжающуюся через растения к губкам и асцидиям, а затем к свободно живущим морским организмам. Однако признавая развитие, Аристотель не допускал мысли о развитии низших организмов к высшим.
Взгляды Аристотеля оказали влияние на века, ибо последующие греческие и римские философские школы полностью разделяли идею самопроизвольного зарождения, которая все больше и больше наполнялась мистическим содержанием. Описания различных случаев самозарождения даны Цицероном, Овидием, позднее Сенекой, Плинием, Плутархом и Апулеем. Идея изменяемости прослеживается во взглядах древних философов Индии, Китая, Месопотамии, Египта. Раннее христианство обосновывало доктрину абиогенеза примерами из Библии, которая, в свою очередь, заимствовала сказания, сложенные в Египте и Вавилоне. Подчеркивалось, что самозарождение действует от сотворения мира до наших дней.
В течение Средних веков (V-XV вв.) вера в самопроизвольное самозарождение была господствующей среди ученых того времени, ибо философская мысль тогда могла существовать лишь в качестве богословской мысли, а сама философия, как отмечал Ф. Энгельс, была «служанкой богословия». Поэтому сочинения средневековых ученых содержат многочисленные описания самозарождения насекомых, червей, рыб. Тогда часто считали, что даже львы возникли из камней пустыни. Парацельс (1498-1541) приводил рецепт изго- товления гомункулуса путем помещения семени человека в тыкву. Как известно, Мефистофель из трагедии Гете «Фауст» называл себя повелителем крыс, мышей, мух, лягушек, клопов и вшей.
Средневековье не внесло новых идей в представления о развитии органического мира. Напротив, в тот период царило креационистское представление о возникновении живого в результате акта творения, о постоянстве и неизменности существующих живых форм. Вершиной креационизма было создание лестницы тел природы: Бог - ангел - человек - животные - растения - мицеллы.
Гарвей (1578-1667) допускал, что черви, насекомые и другие животные могли зарождаться в результате гниения, но при действии особых сил. Ф. Бэкон (1561-1626) считал, что мухи, муравьи и лягушки могут самопроизвольно возникать при гниении, однако к вопросу подходил материалистически, отрицая непреодолимую грань между неорга-
ническим и органическим. Р. Декарт (1596-1650) также признавал самопроизвольное зарождение, но отрицал участие в нем духовного начала. По Р. Декарту, самозарождение - это естественный процесс, наступающий при определенных (непонятных) условиях.
Оценивая взгляды выдающихся деятелей прошлого, можно сказать, что доктрина самозарождения не подвергалась сомнению вплоть до середины XVIII в. Метафизичность воззрений в XVII-XVIII вв. особенно проявлялась в представлениях об неизменности видов и органической целесообразности, которые считались результатом мудрости творца и жизненных сил. Характеризуя мировоззрение, сложившееся в XVII-XVIII вв., центром которого являлось представление об абсолютной неизменности природы, Ф. Энгельс писал, что «Земля оставалась от века или со дня своего сотворения (в зависимости от точки зрения) неизменно одинаковой. Теперешние "пять частей света" существовали всегда, имели всегда те же самые горы, долины, реки, тот же климат, ту же флору и фауну, если не говорить о том, что изменено или перемещено рукой человека. Виды растений и животных были установлены раз и навсегда при своем возникно- вении, одинаковое всегда порождает одинаковое. В природе отрицали всякое изменение, всякое развитие. Для естествоиспытателей виды были чем-то окостенелым, неизменным, а для большинства чем-то созданным сразу. Наука все еще глубоко увязает в теологии».
Однако вопреки господству метафизических представлений в XVI-XVII вв. все же происходит ломка догматического мышления Средневековья, обостряется борьба против духовной диктатуры церкви, возникает и углубляется процесс познания, который привел в XVIII в. к существенной аргументации против теории абиогенеза и к возбуждению интереса к эволюционизму.
Осуществив ряд экспериментов с мясом и мухами, в
В
В 1861-1862 гг. Л. Пастер представил развернутые доказательства невозможности самозарождения в настоях и растворах органических веществ. Экспериментально он доказал, что источником загрязнений всех растворов являются бактерии, находящиеся в воздухе. Исследования Л. Пастера произвели огромное впечатление на современников. Англичанин Д. Тиндаль (1820-1893) нашел, что некоторые формы микробов очень резистентны, выдерживая нагревание до 5 часов. Поэтому он разработал метод дробной стерилизации, называемый сейчас тиндализацией.
Опровержение доктрины абиогенеза сопровождалось формированием представлений о вечности жизни. В самом деле, если самозарождение жизни невозможно, рассуждали многие философы и ученые, то тогда жизнь вечна, автономна, рассеяна во Вселенной. Но как она попала на Землю? Чтобы ответить на этот вопрос, шведский ученый С. Аррениус (1859-1927) в начале XX в. сформулировал гипотезу панспермии, в соответствии с которой жизнь существует во Вселенной и переносится в простейших формах с одного небесного тела на другое, включая Землю, под давлением световых лучей. Сторонники этой гипотезы считали, что перенос жизни на Землю возможен и с помо- щью метеоритов. Однако гипотеза панспермии вызывала возражения в том плане, что в космическом пространстве действуют факторы, которые губительны для микроорганизмов, и что эти факторы исключают циркуляцию микроорганизмов за пределами земной атмосферы. Становилось все более ясным, что жизнь уникальна, что истоки жизни следует искать на Земле. Еще в XVII-XVIII вв. внимание нату- ралистов и философов привлекал ряд вопросов, обсуждение которых расшатывало догмы о неизменяемости органического мира. Один из таких вопросов касался изменяемости видов. Тогда для многих
мысль о возможном изменении организмов под влиянием климата, пищи, почвы, упражнений и неупражнений органов, гибридизации и других факторов казалась сама собой разумеющаяся. Например, Ф. Бэкон (1561-1626), Д. Рей (1628-1705), Ж. Бюффон (1707-1788) и др. допускали изменение организмов под влиянием климата и почвы, а Ш. Бонне признавал, что паразитические черви произошли от свободно живущих червей. Даже К. Линней не исключал изменяемости видов под влиянием почвы и климата, скрещиваний организмов разных видов между собой. Мысль об изменяемости видов поддерживали Эразм Дарвин (1731-1802), К.Ф. Вольф (1734-1794), французские философы-материалисты Д. Дидро (1713-1784), Ж.О. Ламетри (1709-1751), П. Гольбах (1723-1789). Допущение изменяемости видов часто сопровождалось верой в наследование организмами приобретенных свойств. Однако мысль об изменяемости видов еще была далека от идеи исторической преемственности видов, развития органического мира от низшего к высшему.
Не меньшее значение в то время имел вопрос о «естественном родстве» организмов. Речь шла о группировке организмов на основе их естественного сродства, о допущении, что отдельные организмы могли произойти от общих родоначальников. Например, Ж. Бюффон считал, что могли быть «общие родоначальники» для нескольких семейств, в частности для млекопитающих, им допускалось 38 общих родоначальников. В России мысль о происхождении организмов ряда видов от общих родоначальников развивал П.С. Паллас (1741-1811).
Далее привлекал внимание вопрос о факторе времени в изменении организмов. В частности, значение фактора времени для существования Земли и формирования на Земле органических форм признавали И. Кант (1724-1804), Д. Дидро, Ж. Бюффон, М.В. Ломоносов (1711-1765), А.Н. Радищев (1749-1802), А.А. Каверзнев (?-1748). И. Кант определял возраст Земли в несколько миллионов лет, а М.В. Ломоносов писал, что время, которое было необходимо для создания организмов, является большим церковного исчисления. Признание фактора времени имело несомненное значение для исторического понимания развития организмов. Однако представления о времени в тот период сводились лишь к мысли о неодновременности появления организмов разных видов, но не к при- знанию развития организмов во времени.
Важное значение тогда имел вопрос о последовательности природных тел. Значительный вклад в формирование идеи последовательно-
сти природных тел принадлежит Ш. Бонне и Г. Лейбницу. В России эту идею поддерживал А.Н. Радищев. Не имея достаточных знаний об организмах, Ш. Боннэ, Г. Лейбниц и другие натуралисты того времени возродили аристотелевскую «лестницу природы». Расположив на ней организмы по ступеням (на главной ступени оказался человек), они создали «лестницу существ», в которой имелись непрерывные переходы от Земли и камней к Богу. Ступеней в лестнице было столько, сколько есть животных. Отражая мысль о единстве и связи живых форм, об усложнении организмов, «лестница существ» в целом явилась порождением метафизического мышления, ибо ее ступени отражали простое соседство, но не результат исторического развития.
Существенное внимание в те времена привлекал вопрос о «прототипе» и единстве плана строения организмов. Допуская существование исходного существа, многие признавали единый план строения организмов. Дискуссии по этому вопросу имели важное значение для последующих представлений об общности происхождения организмов.
Для многих большой интерес вызывал вопрос о трансформации организмов. Например, французский натуралист Б. де Майе (1696-1738) считал, что в море живут вечные семена жизни, которые дают начало морским живым формам, трансформирующимся затем в земные организмы. Отмечая позитивную роль трансформизма в эволюционизме, следует все же отметить, что он был механическим и исключал мысль о развитии, об историзме.
Наконец, центром внимания в то время был вопрос о возникновении органической целесообразности. Многие философы и натуралисты признавали, что целесообразность не изначальна, что она воз- никла естественным путем в результате браковки дисгармонических организмов. Обсуждение этого вопроса продвигало эволюционизм, но не достигало существенного результата, ибо появление одной формы рассматривалось независимо от появления другой.
Итак, к концу XVIII в. появились идеи, противоречащие представлениям о неизменности видов, но они не сложились в систему взгля- дов, а метафизичность мышления мешала полностью отвергнуть религию и взглянуть на природу по-новому. Первым, кто специально обратился к изучению проблем эволюции, был французский ученый Ж.Б. Ламарк (1744-1829). Созданное им учение явилось завершением предыдущих поисков многих натуралистов и философов, пытавшихся осмыслить возникновение и развитие органического мира.
Ж.-Б. Ламарк был деистом, так как считал, что первопричиной материи и движения является творец, но дальнейшее развитие проис- ходит благодаря естественным причинам. По Ламарку, творец делал лишь первый акт, создавая самые простые формы, которые затем развивались, дав начало всему многообразию на основе естественных законов. Ламарк был также антивиталистом, считая, что живое возникает из неживого. Он рассматривал самозарождение в качестве естественного закономерного процесса, являющегося начальным пунктом эволюции. Признавая развитие от простого к сложному и опираясь на «лестницу существ», Ламарк пришел к заключению о градации, в которой он увидел отражение истории жизни, развитие одних форм из других. Ученый считал, что развитие от простейших форм до самых сложных составляет главное содержание истории всего органического мира, включая и историю человека. Однако, доказывая эволюцию видов, Ламарк полагал, что они текучи и между ними нет границ, т. е. фактически он отрицал существование видов.
Главными причинами развития живой природы, по Ламарку, является врожденное постоянное стремление организмов к усложнению через совершенствование. Эволюция идет на основе внутреннего стремления организмов к прогрессу, а положения об упражнениях и неупражнениях органов и о передаче по наследству приобретенных под влиянием среды признаков являются законами. По мнению Ламарка, факторы среды влияют на растения и простые организмы прямо, «вылепливая» из них, как из глины, нужные формы, т. е. изме- нения среды приводят к изменениям видов. На животных факторы среды влияют косвенно. Изменения среды приводят к изменению потребностей животных, изменение потребностей ведет к изменению привычек, а изменение привычек сопровождается использованием или неиспользованием тех или иных органов. В обоснование этих взглядов Ламарк приводил многие примеры. Например, форма тела змеи, как он считал, является результатом привычки этих животных ползать по земле, а длинная шея жирафа обязана необходимости доставать плоды на деревьях. Использование (упражнение) органа сопровождается его дальнейшим развитием, тогда как неиспользование органов - деградацией. Изменения, индуцированные внешними условиями (обстоятельствами), наследуются потомством, накапливаются и ведут к переходу от одного вида в другой.
Исторические заслуги Ламарка заключаются в том, что ему удалось показать развитие от простого к сложному и обратить внимание
на неразрывную связь организма со средой. Однако обосновать эволюционное учение Ламарку все же не удалось, ибо он не смог выяснить подлинные механизмы эволюции. Как отмечал К.А. Тимирязев (1843-1920), Ламарк не объяснил важнейший вопрос, касающийся целесообразности организмов. Учение Ламарка содержало элементы натурфилософии и идеализма, поэтому ему не удалось убедить современников в том, что эволюция действительно имеет место в природе.
Ч. ДАРВИН И ЭВОЛЮЦИЯ
Творцом первой подлинно научной теории эволюции является великий английский ученый Чарлз Дарвин (1809-1882). Главный труд Ч. Дарвина - «Происхождение видов путем естественного отбора, или Сохранение благоприятствуемых пород в борьбе за жизнь» (1859), существенным дополнением к которому являются труды «Изменение домашних животных и культурных растений» (1868) и «Происхождение человека и половой отбор» (1871).
Эволюционное учение Дарвина состоит из трех разделов, а именно: совокупность доводов в пользу того, что историческое развитие организмов действительно имеет место; положение о движущих силах эволюции; представления о путях эволюционных преобразований.
Доводы в пользу того, что эволюция действительно имеет место, Ч. Дарвин черпал из разных наук. Наиболее убедительные доказа- тельства были взяты им из палеонтологии. Например, обнаружение в древнейших слоях организмов, сильно отличающихся от современных, и постепенное увеличение сходства организмов из позднейших слоев для Ч. Дарвина было летописью эволюции. Далее он использовал данные эмбриологии того времени, которые свидетельствовали о единстве происхождения организмов, а также о закономерностях распределения организмов на суше и в воде и явной зависимости организации животных и растений от условий обитания (на материках и островах), которые свидетельствовали в пользу эволюции и разных ее направлений на материках и островах. Наконец, ученый широко использовал достижения сельскохозяйственной практики.
Движущими силами эволюции Ч. Дарвин назвал наследственность, изменчивость и естественный отбор. Он считал, что наслед- ственность и изменчивость позволяют фиксировать изменения и трансформировать их в поколениях. Изменчивость вызывает раз-
нообразие, а наследственность передает эти изменения потомству. Следовательно, для всех живых существ в результате изменений и скрещиваний характерна наследственная гетерогенность.
Уделяя внимание изменчивости и стремясь объяснить причины закономерных преобразований живых существ в процессе эволюции, Ч. Дарвин пришел вначале к заключению, что в искусственных условиях породы животных и сорта растений созданы искусственным отбором. Что же касается естественных условий, то в поисках ответов на вопросы об источниках целесообразной организации и жизнедеятельности живых существ в природе, о механизмах возникновения новых форм, сохраняющихся в данных условиях существования и дающих начало новым разновидностям, затем новым видам и более крупным систематическим подразделениям, Ч. Дарвин сформулировал представления о борьбе за существование и естественном отборе.
Поскольку все организмы очень склонны к интенсивному размножению, то в пределах каждого вида производится потомства больше, чем оно выживает. Излишнее потомство гибнет в результате борьбы за существование, формы которой очень разнообразны. По Дарвину, борьба за существование происходит как между видами, так и внутри видов, причем внутривидовая борьба является более ожесточенной по сравнению с межвидовой, ибо особи обитают в одной местности, нуждаются в одинаковой пище, подвергаются одинаковым опасностям и т. д. В результате борьбы за существование выживают наиболее приспособленные, т. е. те организмы, которые обладают каким-то признаком, обеспечивающим приспособление. Следовательно, неизбежным следствием борьбы за существование является естественный отбор. «Сохранение благоприятных индивидуальных различий и изменений и уничтожение вредных я назвал естественным отбором, или переживанием наиболее приспособленных» (Ч. Дарвин). Дарвин считал, что естественный отбор является главным механизмом эволюции.
Основу представлений Ч. Дарвина о путях эволюционных преобразований составляла мысль о дивергенции (расхождении) при- знаков. По Дарвину, в связи с изменяющимися условиями жизни и благодаря естественному отбору выживали те формы, которые наиболее полно отклонялись от исходной, притом в разных направлениях. Это отклонение происходило на основе расхождения признаков и вело к уменьшению конкуренции, так как благодаря дивергенции организмы получили возможность использовать разнородные усло-
вия существования. Ученый считал, что в результате дивергенции образуются разновидности, которые являются зачатком вида, т. е. дивергенция создает новые виды. По Дарвину, изменения организмов происходят в силу естественной закономерности, но в то же время они случайны по отношению к результатам естественного отбора. Являясь результатом отбора, органическая целесообразность носит относительный характер. Этим Дарвин преодолел метафизическое противопоставление случайности необходимости, что до него нико- му не удавалось.
Заслуги Ч. Дарвина перед естествознанием имеют непреходящее значение. Он всесторонне обосновал исторический метод в применении к природе, создал теорию эволюции и изгнал из науки креационизм. Теория эволюции существенно подорвала влияние религии. Вклад Ч. Дарвина в естествознание, по оценке Ф. Энгельса, равен вкладу в социальные науки. Ф. Энгельс писал: «Ч. Дарвин открыл закон развития органического мира на нашей планете, закон, определяющий движение и развитие человеческой истории». Ф. Энгельс назвал теорию эволюции крупнейшим достижением XIX в. Оценивая взгляды Дарвина, он отмечал, что Ч. Дарвин поставил биологию на научную основу.
СИНТЕЗ ДАРВИНИЗМА И ГЕНЕТИКИ
Учение Дарвина касалось эволюции крупных групп организмов, включая все таксоны, в большие отрезки времени, измеряемые геологическими масштабами, и на обширных территориях. Следовательно, классический дарвинизм является учением о макроэволюции. Однако оно оказалось недостаточно разработанным в генетическом плане. Дарвин имел в виду эволюцию особей, но ведь особи живут, как сейчас известно, в популяциях. Поэтому через 8 лет после выхода книги «Происхождение видов» англичанин Ф. Дженкин поставил такой вопрос: «Если отбор оставляет в живых те особи, которые лишь незначительно отличаются от других, то уже при последующем скрещивании наступает "поглощение" новых признаков, т.е. партнер по скрещиванию, вероятнее всего, не имеет этого нового свойства - произойдет растворение признаков в потомстве». Дарвин называл этот вопрос кошмаром Дженкина, так как никогда не мог дать удовлетворительный ответ.
Дарвинизму длительное время не хватало генетической основы, но генетика очень быстро сама подошла к эволюции. Используя
методологию
классического генетического анализа, генетики стали анализировать роль
отдельных факторов эволюции путем вычленения элементарных единиц и
протекающих в них процессов. Первый шаг на пути объединения дарвинизма и
генетики заключался в законе Харди-Вайнберга, которые в
Исходя из закона Харди-Вайнберга и учитывая влияние отбора и возникновение новых мутаций, в
Позднее Н.П. Дубинин и Д.Н. Ромашов показали, что, когда популяции малы, в них происходят процессы, получившие название генетико-автоматических или дрейфа генов, в результате которого изменяются частоты встречаемости генов, устраняются гетерозиготы и появляются гомозиготы. Изолированная популяция становится доминантной гомозиготной или рецессивной гомозиготной. Если дрейфует мутантный летальный ген, это ведет к вымиранию организма. Таким образом, структура популяции зависит не только от появления новых мутаций, но и от простого изменения частоты встречаемости данного гена.
Современное эволюционное учение представляет собой синтез достижений дарвинизма, физико-химической биологии, генетики, систематики, морфологии, физиологии, экологии, биогеоценологии и других наук. В наше время эволюционное учение, основу которого составляет дарвинизм, - это наука об общих законах развития органической природы. Это учение стало методологической основой всех
специальных биологических дисциплин, выдающимся естественнонаучным обоснованием диалектического материализма.
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О ПРОИСХОЖДЕНИИ ЖИЗНИ
Традиционно происхождение жизни рассматривают в качестве процесса образования живых форм (необиогенеза) из неорганических веществ, т. е. в виде химической эволюции. В наше время абиогенезу противопоставлена концепция постепенного возникновения и развития живого материала в течение длительного периода времени. Эта концепция получила название археобиоза или археогенеза.
Общепризнанной является теория происхождения жизни, впервые предложенная в
На первой стадии происхождения жизни осуществлялся абиогенный синтез карбидов (соединений углерода с металлами) и графита из неорганических веществ. Из карбидов и графита затем формировались простейшие углеводороды.
На второй стадии происходило освобождение углеводородов в первичную атмосферу Земли, где они реагировали с водяными парами, аммиаком и другими газами. Эти реакции интенсифицировались коротковолновым УФ-излучением и электрическими разрядами и происходили в среде, содержащей очень мало кислорода, но много водорода, метана, аммиака и паров воды. Под влиянием Солнца вода разлагалась на кислород и водород. Водород улетучивался, тогда как кислород окислял аммиак до молекулярного азота, а углеводороды - до спиртов, альдегидов, кетонов и органических кислот. Эти вещества с дождями выпадали из влажной холодной атмосферы в моря
и океаны, где накапливались, а благодаря абиогенной полимеризации и конденсации приближались по строению к тем химическим соединениям, которые входят в состав живых организмов. Вода морей и океанов становилась как бы «первичным бульоном».
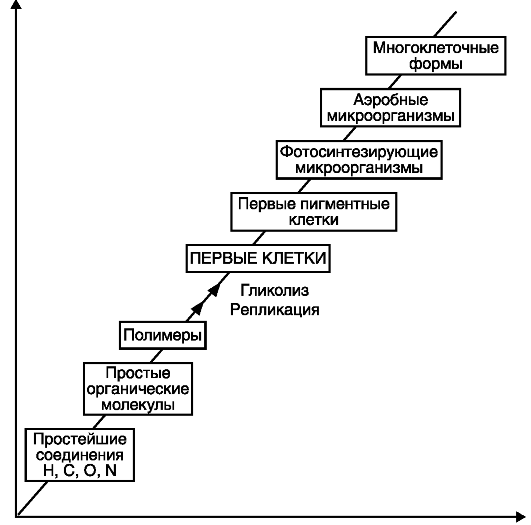
Рис. 80. Происхождение жизни
На третьей стадии в «первичном бульоне» происходило образование коацерватных капель, которые, достигнув определенной величины, приобретали свойство открытых систем, реагирующих со средой. В дальнейшем шло совершенствование и селекция коацерватов, которые становились устойчивыми системами, обособленными от среды и способными к реакциям (рис. 81).
На четвертой стадии совершенствовался метаболизм, упорядочивались мембраны, наступило постепенное установление корреляций
между нуклеиновыми кислотами и белками, что означало появление первичных организмов. Из-за отсутствия кислорода в примитивной земной атмосфере и гидросфере метаболизм первичных организмов был анаэробным. Они питались за счет органических веществ, растворенных в первичном океане. Однако запасы абиогенно возникших органических субстанций постепенно истощались, что давало преимущество тем организмам, которые обладали способностью поглощать свет и создавать органическое вещество из неорганических соединений углерода. В результате отбора в условиях повышенной концентрации СО2 в среде возник биологический путь синтеза органического вещества - фотосинтез. Это произошло около 2,7 млрд лет назад. Вслед за фотосинтезом наступило расхождение организмов на растения и животных.
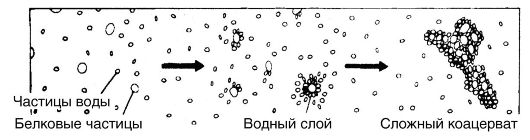
Рис. 81. Формирование коацерватов
Прогрессивное развитие первичных организмов и естественный отбор обеспечили возникновение автотрофов, использующих сол- нечную (световую) энергию для синтеза органических соединений из простейших неорганических. В свою очередь, это способствовало дальнейшему непрерывному синтезу органических веществ, появлению свободного газообразного кислорода в атмосфере, что полностью изменило жизненные процессы на Земле.
Близкие взгляды развивал Н.Г. Холодный (1882-1953), который тоже считал, что вначале образовывались карбиды, в результате взаимодействия которых с водными парами возникали углеводороды, а затем из последних посредством окисления образовались органические кислоты, спирты и т. д. В отличие от А.И. Опарина Н.Г. Холодный считал, что жизнь возникла не в Мировом океане, а в мелководьях после появления суши, что способствовало концентрации органических веществ и образованию коацерватов. Превращение пробиотических структур (пробионтов) в примитивные организмы произошло с окончательным высыханием.
Близкие взгляды развивал также английский ученый Д. Бернал (1901-1971), который полагал, что неорганическими предшествен-
никами были углекислота, неорганические ортофосфаты, сероводород и что первичный синтез органических соединений заключался в образовании очень простых молекул из воды, метана и аммиака. В последующем в результате полимеризации возникли более сложные структуры, связанные с клеточной организацией. По Д. Берналу, возникновение жизни шло по схеме атом - молекула - мономер - полимер - организм. Но в отличие от А.И. Опарина он считал, что конденсация органических молекул происходила не путем образования коацерватов, а за счет адсорбции первых полимеризованных структур на минеральных частицах.
Теория А.И. Опарина имеет ряд доказательств, среди которых важнейшими являются результаты экспериментов, выполненные американским ученым С. Миллером еще в 50-е гг. XX в. Пропуская электрический разряд через смесь, состоящую из метана, водорода, аммиака и водяных паров, С. Миллер получил абиогенным путем аминокислоты (глицин и аланин), альдегиды, молочную, уксусную и другие кислоты. В исследованиях ученых, которые вместо электрического разряда использовали ионизирующее излучение, УФ-излучение и тепло, было показано абиогенное образование других аминокислот, азотистых оснований, полисахаридов.
Установлено также, что при повышенных температурах и удалении свободной воды из реакционных смесей имеет место поликонденсация аминокислот с образованием протеиноидов молекулярной массой 4 000-10 000 дальтон. Под действием УФ-излучения или ионизирующего излучения в водных растворах происходит поликонденсация нуклеотидов с образованием связей 3'-5'.
В последние годы сложилось представление о том, что жизнь возникла не в океане, а в геотермальных источниках в ранний период истории Земли (архей), и что она началась с РНК. Предполагают, что местами абиогенной поликонденсации азотистых оснований РНК (формирование цепей РНК) могли служить такие минералы, как цеолиты, характеризующиеся трехмерной сетчатой структурой. Эти минералы могли быть катализаторами (катализирующими свойствами обладала их поверхность), сходными с белками-ферментами, появившимися значительно позднее. Поэтому, располагаясь в трещинах вулканических пород вблизи гидротермальных источников, цеолиты «плавали» в горячих растворах (40 ?С) с высоким содержанием метана, аммиака, фосфатов и рН порядка 8,0, катализируя при этом сборку олигонуклеотидов и полинуклеотидов РНК.
В последующем молекулы РНК развились в самореплицирующиеся структуры, которые затем стали изолированными от цеолитов. Наконец, они объединялись с липидами, что способствовало подготовке их к жизни в океане, где завершалось образование первичных жизненных форм.
Представление о том, что жизнь начиналась с РНК, согласуется с открытием энзиматической активности ряда молекул РНК (рибо- зимов). Исходя из данных о том, что молекулы РНК обладают одновременно информативными и каталитическими свойствами, стали предполагать, что они были первыми ферментами и что в нуклеотидном бульоне они сами себя собирали. Первичные молекулы РНК развивались затем в самореплицирующиеся структуры, используя мутации и рекомбинацию. На следующей стадии РНК-молекулы стали синтезировать белки, вначале путем развития адапторных молекул РНК, которые могли связывать активированные аминокислоты, затем посредством расположения их соответственно РНКшаблону, используя молекулы РНК рибосом. Впоследствии появилась ДНК, информация в которой копировалась РНК посредством реверсивной (обратной) транскрипции, после чего РНК приобрела промежуточную роль.
Предполагают, что мир РНК связывает «пробиотический мир» с ДНК. Олигонуклеотиды РНК обеспечивают синтез комплементарных цепей так называемой пептидной нуклеиновой кислоты (ПНК), в которой вместо фосфодиэфирных скелетных связей скелетом является амид.
В пользу представлений о пробиотической стадии служат данные и о том, что одноцепочечная ДНК способна сенлигировать олигодезоксинуклеотиды в присутствии некоторых ионов. Следовательно, ДНК тоже обладает каталитической активностью, действуя как металлофермент (ДНК-зим); РНК и ДНК обладают двойной способностью: катализировать реакции и хранить генети- ческую информацию.
Итак, возникновение жизни можно представить в ряде последовательных связей, а именно: пробиотические химические процессы -- про -- РНК -- мир (первые самореплицирующиеся молекулы) -- РНК - мир (биохимические реакции, катализируемые генетически кодируемыми рибозами) - ДНК - белок - мир (биохимические реакции, катализируемые ДНК-кодируемыми белковыми ферментами - первые живые одноклеточные организмы.
ХОД, ГЛАВНЫЕ НАПРАВЛЕНИЯ И ДОКАЗАТЕЛЬСТВА
ЭВОЛЮЦИИ
Классические представления о ходе эволюции и доказательства эволюции получены прежде всего в палеонтологии в результате изу- чения ископаемых организмов, живших в прошлые эпохи. В более глубоких слоях Земли обнаруживаются останки более древних форм жизни, тогда как в поверхностных слоях находят останки более поздних форм. Можно сказать, что история жизни на Земле написана на языке ископаемых останков. Палеонтологический материал дает основание судить также о темпах и направлениях эволюции.
Прецизионность данных палеонтологии повышается при сравнении их с данными других наук. В частности, посредством изучения ископаемых останков и использования радиоактивных изотопов (метода радиодатирования), а также использования данных геологии, биогеографии, систематики и других наук сконструирована шкала геологического времени, с помощью которой филогенез, т. е. изменения земной фауны и флоры, можно проследить во времени (табл. 27). В соответствии с этой шкалой историю Земли подразделяют на несколько последовательных эр, которые, в свою очередь, подразделяют на периоды, а периоды на эпохи. Эрами являются катархей, архей, протерозой, палеозой (возраст рыб и амфибий), мезозой (возраст рептилий) и кайнозой (возраст млекопитающих). Конец одной эры и начало другой сопровождались существенными преобразованиями лика Земли. Менялось соотношение между размерами суши и морей, происходили горообразовательные процессы, менялся климат. Изменения лика Земли сопровождались сменой флор и фаун, изменениями в структуре биогеоценозов. В отдельных случаях имеется возможность проследить происхождение видов из древних форм на протяжении нескольких эволюционных стадий. Наиболее убедительным примером является происхождение лошади.
Предполагают, что жизнь возникла на границе катархея и архея, который начался около 3,5 млрд лет назад, а его длительность составляет около 2 млрд лет. Эта эра характеризуется широкой вулканической деятельностью. Обнаружение в архейских породах графита и чистого углерода предположительно указывает на существование в архее живых организмов растительной и животной природы.
В протерозое происходило отложение большого количества осадков, формировалось ледниковое покрытие Земли. Обнаружены иско-
Эра | Период | Эпоха | Продолжитель- ность | Время от на чала периода до наших дней, млн лет | Преобладающие формы | Климат | Геологические условия |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
Кайнозой (новая) | Четвертичный | Современная | 0,02 | 11 тыс. | Травянистые растения, млекопитающие. Эпоха человека | Теплый | Конец последнего ледникового периода |
Плейстоцен | 1 | 1,7 | Вымирание многих видов растений и крупных млекопитающих животных. Появление человека | Холодный | Повторные оледенения (четыре ледниковых периода) | ||
Третичный | Плиоцен | 12 | 13 | Цветковые растения. Млекопитающие (появление слонов, лошадей, верблюдов, сходных с современными) | Прохладный | Поднятие отдельных гор. Остепенение суши. Вулканическая активность | |
Миоцен | 13 | 25 | Появление первых человекообразных обезьян | Прохладный | Образование гор. Вулканическая активность | ||
Олигоцен | 11 | 36 | Леса. Однодольные цветковые растения. Предшественники многих современных родов млекопитающих (включая человекообразных обезьян и людей) | Теплый | Материки низменные |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
Кайнозой (новая) | Третичный | Эоцен | 22 | 58 тыс. | Плацентарные млекопитающие | Теплый | Горы размыты. Внутрикон гинентальных морей нет |
Палеоцен | 5 | 63 | Примитивные млекопитающие | - | - | ||
Мезозой | Мел | 72 | 136 | Однодольные растения. Динозавры. Примитивные млекопитающие | Образуются горы, отложения мела, сланцев | ||
Юра | 46 | 181 | Цикадофиты и хвойные. Динозавры, древнейшие птицы, насекомоядные, сумчатые | Материки довольно возвышены. Мелководные моря | |||
Триас | 49 | 230 | Голосеменные. Динозавры | Теплый, усиливается засушливость | Материки приподняты над уровнем моря | ||
Палеозой (древний) | Пермь | 50 | 280 | Голосеменные, насекомоядные, пресмыкающиеся | Аридный | Материки приподняты, периодическое оледене- ние | |
Карбон (каменноугольный) | 65 | 345 | Папоротникообразные, плауны, голосеменные. Земноводные | Теплый, влажный | Болота, поднятие суши, материки низменные | ||
Девон | 60 | 405 | Первые леса (голосеменные). Рыбы | Аридный | Небольшие внутриконтинентальные моря. Оледенение |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
Палеозой (древний) | Силур | 20 | 425 | Водоросли. Морские паукообразные (скорпионы) | Засушливый | Обширные внутриконтинентальные моря. Поднятие суши | |
Ордовик | 75 | 500 | Морские водоросли. Наутиловидные. Трилобиты, моллюски | Теплый, включая Арктику | Заметное погружение суши | ||
Кембрий | 100 | 600 | Морские водоросли. Трилобиты и плеченогие | Умеренный | Материки низменные | ||
Протерозой | 1000 | 1600 | Примитивные водоросли и водные грибы. Морские простейшие. В конце - моллюски, черви и другие морские беспозвоночные | Интенсивное осадкообразование. Вулканическая активность, обширная эрозия и многократные обледенения | |||
Первое великое горообразование | |||||||
Архей | 2000 | 3600 | Ископаемые не обнаружены (обнаружено органическое вещество в породах) | Значительная вулканическая деятельность. Слабое осадкообразование. Обширная эрозия | |||
паемые останки медуз, грибов, водорослей, плеченогих, членистоногих (ракоскорпионов) и других организмов, живших в эту эру. Эти находки свидетельствуют о том, что уже тогда существовали микроорганизмы, многие типы водных простейших и беспозвоночных. Предполагают, что первыми микроорганизмами были анаэробы. Земных форм жизни еще не было, но уже тогда произошли переходы от неядерных к ядерным формам и от одноклеточных к многокле- точным.
В палеозойской эре возникли представители основных групп растений и почти всех типов и классов животных (кроме птиц и млекопитающих). Можно сказать, что эта эра знаменовала важнейший этап в развитии жизни, связанный с растениями и животными суши.
В первом периоде этой эры (кембрии) жизнь по-прежнему существовала лишь в воде. В морях были широко распространены одноклеточные и многоклеточные водоросли, примитивные ракообразные и паукообразные формы, например мечехвосты. Морское дно было заселено губками, кораллами и иглокожими, брюхоногими и двухстворчатыми моллюсками. Очень распространены были плеченогие (сидячие животные с двухстворчатой раковиной) и трилобиты (примитивные членистоногие). В то время уже существовали все виды морских животных, кроме хордовых. Материки оставались безжизненными.
В ордовикский период продолжалось начавшееся в кембрии погружение материков в воду. Теплел климат. Начался выход растений на сушу. В пресных водах обитали гигантские скорпионы. В воде появились первые позвоночные. В пресных водах обитали щитковые, представлявшие собой примитивных бесчелюстных рыб, не имевших парных плавников.
В течение силура продолжался выход растений на сушу. Первыми растениями были травянистые псилофиты и древесные формы ликопсидов, сходные с папоротниками. Существовали все ветви беспозвоночных. Начался выход на сушу и животных (паукообразных). Появились животные, дышащие воздухом.
В девонский период этой эры поднималась суша, сокращалась площадь морей. Климат стал более континентальным. На суше впервые возникли примитивные голосеменные растения (семенные папоротники), появились первые леса гигантских папоротников, хвощей и плаунов. В пресных водах появились настоящие акулы. Предки костных рыб дали начало двоякодышащим, кистеперым и лучеперым
рыбам. Лучеперые дали начало в более поздние периоды костным рыбам, а кистеперые стали предками наземных позвоночных.
В течение каменноугольного периода (карбона) этой эры происходит небольшое поднятие суши и потепление климата. Заболоченные леса состояли из обычных и семенных папоротников и широко- лиственных зеленых растений, давших начало залежам каменного угля. В изобилии были земноводные, широко распространились пресмыкающиеся. Появились первые насекомые (предки стрекоз и тараканов).
В течение пермского периода этой эры происходили резкие изменения рельефа Земли и климата. Поднятие суши сопровождалось похолоданием климата. Вымирают папоротникообразные, шире стали распространяться голосеменные растения. Типичными растительными формами в этот период были цикадофиты и хвойные. На Земле в изобилии обитали пресмыкающиеся (пеликозавры и терапсиды). Вымирает более 80% видов морских беспозвоночных, но в морях увеличилось количество костных рыб и головоногих моллюсков. Значительного развития достигают пресмыкающиеся (растительноядные и хищные), среди которых большое место занимали динозавры, принадлежащие к типам птицетазовых (травоядных) и ящеротазовых (хищных).
В мезозойской эре продолжалось поднятие суши, иссушался климат. В эту эру происходило возникновение и вымирание разных пре- смыкающихся. Мезозой часто называют эпохой рептилий.
В триасовом периоде этой эры достигают расцвета семенные папоротники, цикадовые и хвойные. В морях в еще большей мере возросло количество костных рыб и головоногих моллюсков.
В юрском периоде этой эры появляются покрытосеменные растения (цветковые), древнейшие птицы, расширяется многообразие морских пресмыкающихся (ихтиозавров, плезиозавров). Уже суще- ствует четыре отряда млекопитающих, все они были яйцекладущими животными.
В меловой период этой эры были распространены покрытосеменные растения (магнолии, пальмы, клены, дубы и др.). Отмечено появление современных костных рыб, исчезновение морских пресмыкающихся. Вымирают динозавры. Крупные пресмыкающиеся (крокодилы, черепахи, гаутерии) сохраняются лишь в экваториальном поясе. Впервые появляются змеи, птицы. Распространяются млекопитающие, способные рождать живых детенышей (сумчатые).
В кайнозойской эре произошло интенсивное горообразование, похолодание климата. Эта эра характеризуется временем цветковых растений, насекомых, птиц, млекопитающих и появлением человека. Возникли морские организмы всех современных групп, на Земле появились травянистые покрытосеменные растения, птицы, современные млекопитающие, включая обезьян.
В третичном периоде этой эры различают эпохи палеоцен, эоцен, олигоцен, миоцен и плиоцен. В палеоцене уже существовали примитивные плацентарные млекопитающие. В палеоцене и эоцене возникли первые плотоядные хищники и древнейшие копытные. В эоцене и олигоцене на смену первым хищникам пришли формы, давшие начало современным кошкам, собакам, медведям и ласкам, а также тюленям и моржам. Что касается копытных (лошадей, верблюдов и слонов), то их эволюция шла в сторону увеличения размеров тела и изменения количества пальцев. Копытные разошлись на группу с четким числом пальцев (коровы, овцы, верблюды, олени, жирафы, свиньи и гиппопотамы) и группу с нечетким числом пальцев (лошади, зебры, тапиры, носороги). В эоцене существовали китообразные формы, давшие начало китам и дельфинам. В олигоцене широко были распространены предковые формы человекообразных обезьян и человека.
В миоцене и плиоцене древесная растительность вытесняется травянистыми формами. Наступает расцвет цветковых растений, членистоногих и млекопитающих. Конец третичного периода датируется появлением всех современных семейств животных и растений.
Следующий период (четвертичный) этой эры, длящийся последние 1-1,7 млн лет, состоит из плейстоценовой и современной эпох. Для плейстоцена характерны четыре последовательных ледника. Растения и животные, которые были распространены в плейстоцене, сходны с современными растениями и животными. В плейстоцене вымерли многие растения (особенно лесные) и млекопитающие (мамонты, саблезубые тигры). Появились много травянистых форм и человек. Современная эпоха началась около 11 000 лет назад, когда отступил последний ледник.
Хотя и существуют многие проблемы, тем не менее палеонтологическая летопись полностью раскрывает происхождение многих существующих в настоящее время видов животных и растений. Например, очень хорошо прослежено происхождение лошадей, слонов, парнокопытных хищников.
Основополагающее представление об эволюции в целом сводится к тому, что все современные живые существа являются измененными потомками одной или нескольких форм, существовавших ранее. Более молодые по происхождению формы животных и растений произошли от предков, менее отличавшихся друг от друга, а те, в свою очередь, развились от общего единого источника. Первыми возникли бактерии, а от них ответвились вирусы, красные, сине-зеленые водоросли и зеленые жгутиковые. От последних ответвились зеленые водоросли (из которых позднее произошли высшие растения) и бесцветные ризофлагелляты, из которых развились диатомовые водоросли, плесневые грибы, губки и простейшие. Часть простейших стала паразитической. Вторым крупнейшим этапом эволюции была трансформация одноклеточных организмов в многоклеточные. Из цилиарных простейших (цилиофор) развились многоклеточные (синтициальные) плоские черви. Последние разветвились на пять линий, одна из которых ведет к иглокожим и хордовым. Остальные линии повели к появлению большинства других типов животного мира.
Первыми позвоночными были остракодермы, которые развились из беспозвоночных, а из остракодерм развились миноги и панцирные акулы. Хрящевые рыбы (акулы и скатовые) и костные рыбы развились из панцирных акул, из костных рыб - надотряд кистеперых рыб (Crossopterygii), которые являлись примитивными рыбами и затем разошлись на две ветки, одна из них привела к дипловертебронам - ранним земноводным, называемым стегоцефалами. Дипловертеброны были предшественниками современных амфибий и пресмыкающихся. Первые пресмыкающиеся (котилозавры) разделились на ветви, давшие начало динозаврам и морским пресмыкающимся, которые затем вымерли, а также черепахам, крокодилам, ящерицам, змеям, птицам и млекопитающим.
Линия, ведущая к человеку, начинается от тупай и ведет к ранним приматам, разветвляясь затем на обезьян Нового света, обезьян Старого Света, понгид и гоминид. Из понгид (настоящих обезьян) произошли гориллы, орангутаны и шимпанзе. Из гоминид произошел непосредственный предшественник современного человека.
Учение о главных направлениях эволюции сформулировано А.Н. Северцовым (1866-1936), который впервые четко определил различия между прогрессивным и регрессивным развитием, обосновал понятия о морфофизиологическом и биологическом прогрессе и регрессе.
Морфофизиологический прогресс - это изменение структуры и общей жизнедеятельности на пути эволюции от простых форм к сложным, от одноклеточных к многоклеточным. Напротив, морфофизиологический регресс - это упрощение организации и жизнедеятельности, выражающееся в редукции органов (например, хорды у оболочников) на пути эволюции.
Биологический прогресс присущ организмам, которые характеризуются возрастанием приспособленности к окружающей среде, что ведет к увеличению их численности и расширению ареалов. Биологический прогресс может быть результатом морфофизиологического прогресса, но иногда и морфофизиологического регресса. В конечном итоге биологический прогресс ведет к процветанию вида. В современную эпоху процветающими являются насекомые, костные рыбы, птицы, млекопитающие, покрытосеменные растения.
Биологический регресс - это снижение приспособленности организмов к среде, сокращение их численности и ареалов. Результатом биологического регресса является вымирание организмов. Значение биологического прогресса заключается в том, что он играет важную роль в эволюции и обеспечивает главные направления эволюции в виде ароморфозов, идиоадаптаций и общей дегенерации.
Ароморфозы - это изменения организмов, которые поднимают на новый более высокий уровень их морфофизиологическую организацию и жизнедеятельность. Кроме того, они открывают организмам путь к проникновению в новую среду обитания. Ароморфозы возникают скачками. Примерами ароморфозов А.Н. Северцов называл развитие легких и четырехкамерного сердца, превращение парных плавников рыб в парные конечности земноводных. По А.Н. Северцову, ароморфозы обеспечили возникновение новых классов и типов организмов.
Идиоадаптация - это изменения, которые не приводят к повышению уровня организации, но сопровождаются приданием организации большей приспособленности к существующим условиям жизни. Примерами идиоадаптаций А.Н. Северцов называл приспособления насекомых к разным условиям жизни (в воде, почве, воздухе), разным типам питания и т. д.
Общая дегенерация - это изменения, которые затрагивают организацию живых существ и заключаются в снижении активности отдельных или ряда органов. Примерами общей дегенерации А.Н. Северцов называл биологию саккулины из группы паразитических ракообраз-
ных, а также оболочников. Выживаемость этих организмов обеспечивается чрезвычайным разнообразием их личиночных форм.
Учение А.Н. Северцова о главных направлениях эволюционного процесса было развито в трудах И.И. Шмальгаузена (1884-1963), который рассматривал организм как целое в историческом и индивидуальном развитии. Среди главных направлений эволюционного процесса И.И. Шмальгаузен большое внимание уделял различным формам ароморфозов.
Доказательства в эволюции получены в разных науках.
В биогеографии различают шесть биогеографических областей, каждая из которых, если говорить о животных, характеризуется специфическими обитателями.
В Палеоарктической области (Европа, Африканский север от Сахары, часть Азии к северу от Гималаев, Азорские острова и острова Зеленого Мыса) эндемичными являются кроты, олени, быки, овцы, козы, скворцы и сороки.
В Неоарктической области (Гренландия и Северная Америка) эндемичными являются горные козлы, луговые собачки, опоссумы, скунсы, еноты, сойки и американские грифы. Кроме того, здесь встречаются формы, эндемичные для палеоарктической области.
В Неотропической области (Южная и Центральная Америка, юг Мексики и острова Вест-Индии) обитают альпаки, ламы, цепкохвостые обезьяны, тапиры, ленивцы, вампиры, муравьеды и многие виды птиц, не встречающиеся в других частях земного шара.
В Эфиопской области (Африка к югу от Сахары, Мадагаскар) обитают гориллы, шимпанзе, зебры, носороги, бегемоты, жирафы и многие виды рыб и птиц, не обнаруживаемые в других областях.
В Восточной области (Индия, Цейлон, Индокитай, юг Китая, Малайский полуостров и отдельные острова Малайского архипелага) обитают орангутаны, черные пантеры, индийские слоны, гиббоны и долгопяты.
В Австралийской области (Австралия, Новая Зеландия, Новая Гвинея и другие острова Малайского архипелага) эндемичными являются утконосы, кенгуру, вомбаты, коала и другие животные. Специфичны бескрылые птицы эму и казуал, птица-лира и какаду.
Одно из основных положений биогеографии заключается в том, что каждый вид растений и животных возникал только однажды и только в одном месте (центре происхождения), откуда он расселялся до тех пор, пока не натыкался на какую-нибудь преграду, например
географическую, климатическую, пищевую и т. д. Географические ареалы близких видов, как правило, не совпадают, но они и не очень отдалены один от другого.
Особенности географического распространения животных и растений указывают на специфику эволюции каждого вида. Например, наличие в Австралии однопроходных и сумчатых, которых нет ни в одной из других областей, является результатом того, что в мезозое этот континент был изолирован от других частей суши. Примитивным млекопитающим-аборигенам не довелось конкурировать с более приспособленными плацентарными млекопитающими, поэтому они дали начало формам, легко приспособившимся к местным условиям.
Аллигаторы встречаются только в реках юго-востока США и в реке Янцзы (Китай), тогда как магнолии произрастают только в восточной части США, в Китае и Японии. В начале кайнозоя Северная Америка соединилась с Азией (на месте теперешнего Берингова пролива). При тогдашнем теплом климате аллигаторы и магнолии были распространены повсеместно. В дальнейшем из-за поднятия Скалистых гор и похолодания магнолии и аллигаторы погибли повсеместно, кроме юго-восточной части США и Восточного Китая, где не было обледенения. Поскольку магнолии и аллигаторы были разделены в отдаленных частях земного шара на протяжении многих миллионов лет, то в современную эпоху, оставаясь близкородственными видами, они несколько различаются между собой.
Еще Ч. Дарвин отмечал различия островных и материковых флор и фаун. Например, на Галапагосских островах, возраст которых составляет всего лишь 3-5 млн лет и которые никогда не были связаны с материком, отсутствуют лягушки и жабы, ибо ни они, ни их яйца не смогли выжить в морской воде, с которой неизбежно пришлось бы столкнуться мигрирующим на этот остров животным. На этих островах нет также наземных млекопитающих, но на одном из них встречаются гигантские ящерицы и черепахи, которые очень близки к некоторым формам, обитающим на побережье Эквадора. Это указывает на эволюцию растений и животных после заселения этих островов.
В систематике главное доказательство эволюции заключается в том, что все живые существа можно расположить в иерархическую систему таксономических единиц - виды, роды, семейства, отряды, классы и типы. Это означает, что все организмы связаны
между собой филогенетически в результате существующих между ними эволюционных взаимоотношений. Принадлежность организ- мов к тем или иным систематическим группам свидетельствует о том, что большинство промежуточных форм, существовавших в прошлом, вымерло. Если бы виды всех существовавших в прошлом организмов жили до настоящего времени, то классифицировать живой мир на таксономические группы было бы невозможно. Будучи генетически различными, виды представляют собой независимо эволюцинирующие и репродуктивно изолированные единицы. Поскольку можно предполагать, что у генетически сходных видов общий предок существовал в менее отдаленном прошлом по сравнению с генетически различными видами, то степень генетических различий является, по существу, мерой, на основе которой строят филогенетическое древо (рис. 82).
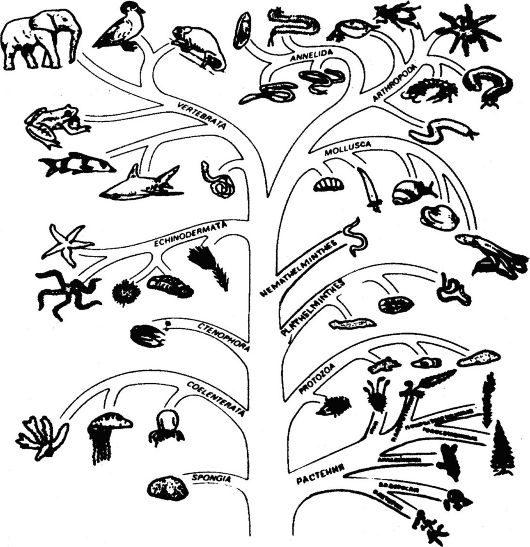
Рис. 82. Филогенетическое древо органического мира
Степень генетических различий между видами определяют либо прямым путем изучения последовательностей нуклеотидов в генах, либо косвенным путем - последовательностей аминокислот в белках. Результаты сравнения последовательностей ДНК разных организмов позволяют определить количество пар нуклеотидов, в которых в ходе эволюции имели место замены азотистых оснований, тогда как сравнение белков от разных организмов позволяет определить различия в аминокислотных последовательностях и их связь со скоростью эволюции (табл. 28). На основе данных о филогении отдельных белков строят филогенетическое древо, которое, как свидетельствует пример с цитохромом С, совпадает с филогенетическим древом, построенным по ископаемым останкам. На основе реконструкции филогении и определения степени генетических различий по аминокислотным последовательностям ряда белков считают, что гены, кодирующие эти белки у животных, происходят от общего предка.
Таблица 28. Связь между аминокислотными различиями в α- и β-глобинах со скоростью эволюционной дивергенции
Животные | α-цепь | β-цепь | α + β (среднее) | Миллионы лет с начала дивергенции |
Плацентарные в сравнении между собой | 16,1 | 16,7 | 16,4 | 100 |
Кенгуру в сравнении с плацентарными млекопитающими | 21,7 | 26,9 | 24,3 | 160 |
Куры в сравнении с млекопитающими (сумчатыми и плацентарными) | 29,6 | 31,7 | 30,6 | 215 |
Змеи в сравнении с теплокровными | 39,2 | 290 | ||
Земноводные в сравнении с наземными животными | 46,7 | 48,9 | 47,8 | 380 |
Костные рыбы в сравнении с четвероногими (рептилиями, амфибиями и млекопитающими) | 49,3 | 49,6 | 49,5 | 400 |
С целью определения степени сходства белков используют также иммунологическое и электрофоретическое сравнение белков. Степень иммунологического сходства белков выражают в иммунологическом
расстоянии, которое приближенно можно связать с различиями по аминокислотным последовательностям. Электрофоретические исследования позволяют выявить электрофоретическое сходство белков и на основе этих данных также определить генетические расстояния между видами.
Эволюция подтверждается данными сравнительной морфологии, эмбриологии, физиологии, биохимии и генетики.
Результаты сравнительного изучения строения животных в пределах систематических групп показывают, что оно имеет общий план. Особенно это проявляется в случае гомологичных органов, которые характеризуются общим строением, сходством эмбрионального развития, иннервации, кровоснабжения и отношения к другим органам. Например, гомологичными являются передний ласт тюленя, крыло летучей мыши, передняя нога лошади и рука человека. Эти органы состоят из почти одинакового количества костей и мышц, их сосуды и нервы имеют сходную топографию. Гомологичные органы с несомненностью указывают на общность происхождения животных, у которых есть эти органы.
Помимо гомологичных органов морфологическим свидетельством в пользу эволюции являются данные о рудиментарных органах, пред- ставляющих собой «останки» развитых органов, имевшихся в прошлом у предковой формы. Например, у человека имеется свыше 100 разных рудиментов, типичными примерами которых являются червеобразный отросток слепой кишки (аппендикс), копчик (слившиеся хвостовые позвонки), волосяной покров туловища и конечностей, третье веко. Червеобразный отросток слепой кишки является результатом перехода человека на диету с большим содержанием мяса и меньшим количеством клетчатки. Слепой вырост в таких условиях уменьшился и стал рудиментом. Напротив, у травоядных животных он представляет собой активно функционирующий орган. Кости задних конечностей китов в связи с переходом их к водному образу жизни стали рудиментами, располагающимися в толще брюшных мышц.
На эмбриологические доказательства эволюции обращал внимание еще Ч. Дарвин. В
животных, но не со взрослыми особями низших форм, как это считал Э. Геккель.
Эмбриологические данные в пользу эволюции сводятся к тому, что на ранних стадиях развития между зародышами млекопита- ющих, рыб, земноводных и пресмыкающихся существует много сходных признаков. В то же время эти организмы обладают рядом собственных признаков, присущих только им и обеспечивающих приспособленность к развитию в утробе материнского организма, но не под скорлупой яйца.
Данные эмбриологии свидетельствуют о том, что после гаструляции развитие зародышей проходит в одном из двух направлений. С одной стороны, бластопор либо превращается в анальное отверстие, либо занимает положение около этого отверстия, как это имеет место в случае иглокожих и хордовых. С другой стороны, бластопор развивается в ротовое отверстие либо занимает положение около этого отверстия, как это имеет место в случае кольчатых червей, моллюсков, членистоногих и др. При развитии как в одном направлении, так и в другом между эктодермой и энтодермой развивается мезо- дерма, которая у иглокожих и позвоночных закладывается частично (карманами или выпячиванием первичной кишки), а у кольчатых червей и других организмов закладывается из специальных клеток, дифференцирующихся на ранних этапах развития.
Вслед за появлением мезодермы у всех хордовых развиваются дорзальный полый нервный тяж, а также нотохорд (основа скелета тела) и жаберные щели (отверстия в глотке). На ранних стадиях развития у зародышей, например, человека, как и у зародышей рыб, имеются жаберные щели, дуги аорты, пересекающие жаберные перегородки, сердце с одним предсердием и одним желудочком, примитивная почка (пронефрос) и хвост с мышцами. Таким образом, на ранних стадиях развития зародыши человека напоминают зародыши рыб.
На более поздних стадиях развития зародыши человека приобретают сходство с зародышами пресмыкающихся, в частности зарастают жаберные щели, сливаются кости, из которых состоят позвонки, исчезает старая почка (пронефрос) и образуется новая почка (мезонефрос), разделяется предсердие на правую и левую части. Еще позднее у зародышей человека развиваются четырехкамерное сердце и метанефрос (совершенно новая почка), исчезает нотохорд. На седьмом месяце внутриутробного развития плод человека покрыт волосами и имеет «обезьянье соотношение» в размерах
тела и конечностей. Явление рекапитуляции признаков у человека объясняют действием сохранившихся «рыбьих», «земноводных» и «пресмыкающихся» генов, существовавших у предковых форм, вслед за которыми действуют гены млекопитающих, а затем приматов. Следовательно, на начальных стадиях развития человека действуют гены предков и проявляются признаки, сходные с признаками предков, на поздних же стадиях действуют гены, характерные для млекопитающих и приматов, в результате чего проявляются признаки млекопитающих и приматов. Закономерность здесь такова, что у зародышей человека раньше проявляются признаки, присущие типам и классам, а позднее признаки, присущие родам и видам. Например, у человека и свиньи процессы развития дивергируют раньше, чем у человека и человекообразных обезьян, поскольку предки человека разошлись с предками свиней раньше, чем произошло расхождение человека и человекообразных обезьян, эволюционировавших от одного предка.
Основные физиологические доказательства эволюции заключаются в сходстве физиологических процессов - дыхания, пищеварения, кровообращения, выделения, реакций на раздражения у организмов разных систематических групп.
Данные биохимии в пользу эволюции очень значительны. Во-первых, химический состав всех живых организмов является сход- ным. В элементарном составе организмов преобладают три элемента - С, О и Н, в атомарном составе 50-60% приходится на С (в земной коре он составляет 0,35%), что указывает на естественное родство всего органического мира. Далее, у всех организмов белки построены из 20 аминокислот. Между белками животных и растений имеется сходство. Например, между гемоглобином позвоночных, гемоцианином беспозвоночных и хлорофиллом растений существует принципиальное сходство, что указывает на единство происхождения всех организмов. Кроме того, имеется иммунологическое сходство между белками крови разных животных. Например, ближайшими родственниками человека являются человекообразные обезьяны, затем обезьяны Старого Света, цепкохвостые обезьяны Нового Света и, наконец, долгопяты. Кошки, собаки и медведи образуют одну родственную группу, тогда как коровы, овцы, козы, олени и антилопы - другую. Тюлени и морские львы стоят ближе к хищным.
Отмечается сходство ферментов и биосинтетических путей у различных организмов. У всех животных и растений в качестве уни-
версального звена в энергетических реакциях используется АТФ. Налицо также сходство продуктов выделения. У человека и других приматов конечным продуктом обмена пуринов является мочевая кислота, у других млекопитающих - аллантоин, у земноводных и у большинства беспозвоночных - мочевина, а у некоторых беспозвоночных - аммиак. Эволюция позвоночных сопровождалась постепенной потерей способности синтезировать ферменты, необхо- димые для постепенного распада мочевой кислоты.
Основное доказательство эволюции, представляемое генетикой, заключается в установлении универсального характера генетического кода. Можно сказать, что это важнейшее доказательство дарвинов- ской идеи единства происхождения всех форм жизни.
Наконец, в пользу эволюции свидетельствует практика сельского хозяйства. Например, все сорта листовой и цветной капусты, кольраби, брокколи и брюссельской капусты происходят от дикой капусты. Кукуруза берет начало от травянистого растения (теосинта), произрастающего в Андах (Боливия) и в Мексике. Все существующие ныне породы домашних животных происходят от диких предков.
УЧЕНИЕ О МИКРОЭВОЛЮЦИИ
Эволюция на уровне вида получила название микроэволюции. В рамках учения о микроэволюции различают элементарные эволю- ционные единицы и явления, элементарный эволюционный материал и элементарные факторы эволюции.
Элементарной единицей эволюции является популяция. Популяция (от фр. population) - это совокупность особей одного вида, длительно (в течение большого числа поколений) населяющих определенную территорию, свободно скрещивающихся между собой и отделенных друг от друга популяций этого же вида давлением изоляции. Для популяций характерны такие показатели, как ареал, численность особей, размеры в пространстве и по числу особей, генетическая гетерогенность, возраст, половой состав.
Эволюция протекает на уровне популяций. Совокупность всех генов во всех их аллельных формах в гаметах организмов, составляющих популяцию, представляет собой пул генов. В случае индивидуального диплоидного организма частота какого-либо аллеля может составлять 100, 50 или 0%, но в популяции организмов частота этого
аллеля является функцией количества (%) индивидуумов в популяции, которые имеют этот ген. Поэтому в популяции организмов частота аллеля может составлять от 0 до 100%. Стойким считается тот генетический пул, в котором частоты генов, независимо от их функций, являются постоянными во всех следующих одно за другим поколениях организмов.
В популяционной генетике различают разницу между частотой генов (аллелей) и частотой генотипов. Допустим, какая-либо воображаемая популяция состоит из 100 индивидуумов, из которых 20 гомо- зиготны по аллелю а, 60 гомозиготны по аллелю А, а 20 гетерозиготны. Следовательно, в этой популяции количество людей с генотипом АА составит 60%, с генотипом Аа - 20% и с генотипом аа - тоже 20%. Из этих частот возможно вычисление общего числа генов и частот генов в популяции (табл. 29).
Таблица 29. Частоты генов популяции
Генотип | Количество индивидов, % | Количество аллелей каждого вида в генотипе | Общее число генов, % | Частота генов |
А /А | 60 | 120 А/20 А | (140/200) =70 | А = 0,7 |
А /а | 20 | 20 а | (60/200) =30 | а = 0,3 |
а /а | 20/100 | 40 а/200 | 100 | 0,1 |
Закон Харди-Вайнберга, в соответствии с которым в постоянной популяции частота доминантных и рецессивных аллелей была, есть и будет константной, предсказывает, что частоты генов порядка 0,7 для А и 0,3 для гена а будут поддерживаться и в будущих поколениях, если популяция окажется устойчивой. Более того, частоты генотипов будут изменяться, если в популяции начнутся случайные скрещивания, но до тех пор, пока не достигнут равновесия, которое остается неизменным, пока популяция устойчива.
Изменения отдельных особей не приводят к эволюционным изменениям. Эволюционируют не особи, а группы особей, составляющие популяцию. Что же касается особей, то они являются объектом действия естественного отбора. В эволюционном процессе популяция - это неделимая единица, характеризующаяся экологическим, морфофизиологическим и генетическим единством. В существовании элементарных эволюционных единиц (популяций) проявляется такое важнейшее свойство жизни, как дискретность.
Элементарным эволюционным явлением служит мутация. В результате мутаций популяции становятся генетически гетерогенными, т. е. представляют собой смесь разных генотипов. Если условия существования популяции постоянны, ее генетический состав в среднем будет неизменным. Однако если имеется давление со стороны каких-либо внешних факторов, причем в одном направлении, то происходит векториальное изменение генотипического состава популяции, т. е. начинается элементарное эволюционное явление.
Мутации случайны во времени и пространстве применительно к отдельным организмам. Мутационные изменения организмов в популяции не являются направленными. Направления изменений организмов определяются естественным отбором. Накопление в генеративных клетках мутаций и их отбор обеспечивают не только микроэволюцию, но и макроэволюцию. Таким образом, случайность в виде мутаций ведет к беспорядку, тогда как закономерность в виде естественного отбора создает упорядоченность.
Современные данные о подвижности геномов свидетельствуют о том, что многие гены могут изменять свой статус и частоту в течение одной или нескольких генераций без дифференциальной репродукции целых генотипов, т. е. неменделевским путем. Например, многократное повторение коротких последовательностей нуклеотидов приводит к образованию новых генов. Так, ген коллагена цыплят содержит несколько десятков эксонов, являющихся повторами одной и той же последовательности, состоящей из 9 нуклеотидов.
Мутационный процесс в качестве элементарного фактора эволюции важен тем, что он является поставщиком элементарного эво- люционного материала, поддерживая генетическую гетерогенность природных популяций. Однако важно заметить, что, выполняя эту роль, мутационный процесс в качестве фактора эволюции не направляет ход эволюционных изменений. Эту функцию осуществляет естественный отбор.
Помимо мутаций, эволюционным явлением служит также рекомбинация генов, но ее вклад в микроэволюцию меньший.
Элементарным эволюционным материалом являются любые генотипически различные особи и группы особей, т.е. мутантные орга- низмы или особи - носители мутаций, различающиеся между собой генетически. С эволюционной точки зрения важнейшей характеристикой мутантных организмов как элементарного эволюционного материала является частота их появления в природных популяциях.
Наличие организмов - носителей мутаций создает различия между близкими природными группами особей (популяциями, подвидами, видами).
Элементарные факторы эволюции - естественный отбор, мутационный процесс, популяционные волны, изоляция, с действия которых начинается эволюция в популяциях.
Важнейшим элементарным фактором эволюции является естественный отбор, основная функция которого заключается в устра- нении из популяций организмов с неудачными комбинациями генов и сохранение генотипов, не нарушающих процесса приспособительного формообразования. Действие естественного отбора проявляется в пределах популяции, но объектами приложения естественного отбора служат отдельные мутантные особи, являющиеся элементарными материалами для естественного отбора.
Специфика действия естественного отбора определяется тем, что организмы, которым благоприятствует отбор, характеризуют- ся повышенной эффективностью размножения и, следовательно, большей приспособленностью. Таким образом, приспособленность организмов является отражением их эффективности размножения. Скорость, с которой снижается частота организмов - обладателей тех или иных генотипов, называют коэффициентом отбора. Отбор действует как против, так и в пользу какого-либо аллеля (организма - носителя этого аллеля). Поэтому результатом действия естественного отбора является либо элиминация того или иного аллеля, либо появление полиморфизма, заключающегося в том, что в популяции будут присутствовать организмы - носители двух или более аллелей одного и того же гена. Отбор может действовать в нескольких случаях, в частности, против рецессивного или доминантного аллеля, в пользу или против гетерозигот.
Различают три основные формы естественного отбора: движущий (или обновляющий), стабилизирующий и дизруптивный.
Представления о движущем отборе были сформулированы Ч. Дарвином. Этот отбор обеспечивает преобразование старых, выработку и накопление новых приспособлений (адаптацию иных приспособлений организмов), что ведет к смене нормы реакции. По существу, он заключается в адаптациогенезе и взаимодействии популяций со средой. В этом проявляется его творческая роль, а вслед за его действием и творческий харарактер эволюции. Один из известных примеров движущегося отбора, действующего про-
тив рецессивных гомозигот, связан с индустриальным меланинозом березовой пяденицы (Biston betularia). До середины прошлого века бабочки этого вида на Британских островах имели светло-серую окраску и были гомозиготными по рецессивному аллелю, контро- лирующему окраску тела. Однако после того, как в промышленных районах Англии стволы деревьев стали чернеть от копоти, выбрасываемой из заводских труб, начала появляться разновидность этих бабочек, окрашенных в темный цвет, т. е. гомозиготных доминантных и гетерозиготных. Бабочки темного цвета почти полностью вытеснили бабочек светлой разновидности, ибо последние оказались более доступными для питающихся ими птиц. Следовательно, отбор «подхватил» бабочек с темной окраской тела. Другим примером отбора против рецессивных гомозигот является отбор при фенилкетонурии человека в то время, когда еще неясен был патогенез этой болезни и еще не прибегали к диетотерапии. Рецессивные гомози- готные организмы (дети) без соответствующей диеты оказывались неприспособленными к жизни и погибали еще до достижения половой зрелости.
Отбор в пользу гетерозигот часто называют сверхдоминированием, поскольку гетерозиготы превосходят по выживанию гомози- гот. Обычно он завершается созданием устойчивого полиморфного равновесия в популяции. Примером такого отбора является отбор при серповидноклеточной анемии, которая возникает в результате того, что в эритроцитах индивидов, гомозиготных по аллелю HbS, синтезируется аномальный гемоглобин. Нормальный гемоглобин вырабатывается эритроцитами индивидов с аллелем Hb* Индивидуумы с генотипом HbSHbS по причине невозможности приспособления к среде из-за аномального гемоглобина в большинстве случаев умирают еще до достижения половой зрелости. Между тем в тех районах тропической Африки и Среднего Востока, где распространена малярия, вызываемая P. faciparum, частота аллеля HbS является довольно большой (до 30%). Причина заключается в том, что гетерозиготы Hb*/ HbS более устойчивы к тропической малярии по сравнению с нормальными гомозиготами Hba/ Hb*. Следовательно, в малярийных районах гетерозиготы Hb*/ HbS обладают селективны- ми преимуществами по сравнению с гомозиготами обоих типов, для которых характерна значительно большая смертность от анемии (HbS/HbS) или от малярии (Hb*/ Hb*). Гетерозиготы имеют предпочтение к выживанию и размножению, поддерживая в популяциях
высокую частоту аллеля EIbS. Таким образом, в данном случае отбор благоприятствует гетерозиготам.
Эволюция - не только исторический процесс, ибо она действует в настоящее время. Например, продолжающаяся эволюция возбудителей болезней человека и животных создает проблемы здравоохранению. Многие штаммы бактерий стали резистентными и неэффективными для антибиотиков в результате естественной селекции резистентных форм. Особенно высокую резистентность для антибиотиков и других лекарственных веществ, используемых в прошлом, показывают возбудители малярии, гонореи, туберкулеза, сальмонеллеза и многих других болезней. Описаны случаи естественной селекции и большинства других организмов. Например, описаны резистентные формы крыс к варфарину. Очень много видов насекомых приобрели резистентность к пестицидам. Многие виды растений приобрели толерантность к токсическим металлам или возникли в результате скрещиваний между природными формами растений и растениями, устойчивыми к ядам.
Понятие о стабилизирующем отборе принадлежит И.И. Шмальгаузену (1946). Если исторически сложившаяся определенная приспо- собительная форма («адаптивная норма») сохраняет свою полезность в жизни организмов, то отбор автоматически отсекает все отклонения от «нормы» и последняя как бы берется под охрану отбора. В противоположность движущемуся отбору стабилизирующий отбор закрепляет норму реакции и уровень, достигнутый организмами в процессе эволюции. Стабилизирующий отбор действует обычно в условиях среды, долго остающейся однотипной.
Дизруптивный естественный отбор - это такой отбор, когда ни одна из групп генотипов в популяции не имеет преимуществ из-за одновременного изменения условий среды. В этом случае у одних организмов отбор идет по одному признаку, у других - по другому, в результате чего популяция как бы разрывается на группы особей, каждая из которых затем эволюционирует самостоятельно. Этот отбор действует против средних промежуточных форм организмов и бывает как индивидуальным, так и групповым.
Диалектическая взаимосвязь между разными формами естественного отбора является отражением чрезвычайной противоречивости эволюции как процесса, но в любом случае творческая роль принадлежит движущему дарвиновскому отбору. Стабилизирующий и дизруптивный отборы играют второстепенную роль.
Определенное значение в эволюции имеет половой отбор, направленный на успех в размножении организмов, но этот отбор также имеет подчиненное значение.
Популяционные волны, или волны жизни (колебания численности особей в популяциях в сторону от средней численности), являются элементарным фактором эволюции по той причине, что этот фактор также служит поставщиком элементарного эволюционного материала для естественного отбора. Примером их служат колебания численности вредителей полей и огородов в разные годы.
Эволюционное значение волн жизни состоит в том, что они резко изменяют концентрацию в популяциях редко встречающихся алле- лей (генотипов). В результате волн жизни под отбор попадают редкие гены, которые обычно ускользают от действия отбора.
Изоляция в качестве элементарного фактора эволюции обеспечивает барьеры, исключающие репродукцию. Различают географи- ческую, экологическую и генетическую изоляцию, каждая из которых ведет в конечном итоге к репродуктивной изоляции животных на межвидовом уровне.
Различают прозиготический и постзиготический механизмы, предотвращающие их скрещивания. К прозиготическим изолирующим механизмам относят географическую и экологическую изоляцию, которая исключает встречи потенциальных брачных партнеров. Известны также поведенческая изоляция, когда потенциальные брачные партнеры встречаются, но из-за недостаточного полового влечения эти встречи заканчиваются отрицательно, и сезонная изоляция, когда партнеры встречаются, но не спариваются, ибо спа- ривание у них в нормальных условиях обычно происходит в разное время. Наконец, известна гаметическая изоляция, при которой после совокупления животных не происходит осеменения, так как сперматозоиды утрачивают жизнеспособность в женских половых путях.
Постзиготические изолирующие механизмы также различны. Например, после встречи партнеров яйцеклетки могут оплодотворяться, но зиготы погибают. Если зиготы развиваются, то гибриды все же не достигают половой зрелости. Далее, могут формироваться гибриды, но их жизнеспособность оказывается невысокой либо они могут быть стерильными (не способными продуцировать гаметы, обладающие нормальными свойствами).
Важнейшей особенностью изоляции как элементарного фактора эволюции является ее длительность. Нарушая скрещивания, закре-
пляя различия в генотипах, изоляция как бы расчленяет популяцию на генетически различные группы организмов, полностью исключая среди них обмен генами. Изоляция является, по существу, усилителем начальных стадий генотипической дифференциации, развертывающихся в популяциях, подставляя разделенные части популяций под действие естественного отбора. Однако изоляция тоже не направляет эволюцию. Это делает естественный отбор.
ПРОБЛЕМЫ ВИДООБРАЗОВАНИЯ
Вид - это популяция особей, которые имеют сходное строение и характеризуются сходными функциями, в природе скрещиваются только между собой, приспособлены к жизни в определенных условиях, имеют характерный ареал распространения и общее происхождение. Появление нового вида - это центральный и важнейший завершающий этап эволюции.
Эволюция - это процесс в двух измерениях. Один из них является анагенезом, под которым понимают эволюцию организмов в одном направлении, тогда как другой - кладогенезом, представляющим собой эволюцию в разных направлениях. Примером анагенеза является, видимо, эволюция H. erectus в H. sapiens, тогда как для кладогенеза характерно расщепление одной эволюционной линии на две или больше. Поэтому главное содержание кладогенетической эволюции заключается в видообразовании путем разделения одного вида на два и более.
Образование новых видов происходит постепенно (географическое видообразование) и измеряется длительными отрезками времени либо мгновенно (квантами) на протяжении короткого периода времени.
Постепенное (географическое) видообразование начинается с дивергенции признаков и осуществляется в два этапа. На первом этапе такого видообразования происходит географическое разделение популяций реками, морями, горами, пустынями, различными особенностями ландшафта (рис. 83). Популяции приспосабливаются к существующим условиям благодаря естественному отбору. Между разными популяциями прерывается обмен генами. Это углубляет генетическую дифференциацию, в которой некоторую роль еще может играть дрейф генов, особенно когда популяции имеют небольшие размеры, вплоть до нескольких особей. Возникновение генетических различий сопровождается появлением репродуктивных изолирующих механизмов. В результате действия последних проис-
ходит накопление в каждой из популяций своего рабочего набора генов и достижение популяциями того состояния, которое уже может быть определено в качестве подвида.
На втором этапе постепенного видообразования завершается репродуктивная изоляция, а генетические различия достигают такого уровня, когда гибриды еще будут появляться, но полностью нежизнеспособными. Каждый подвид продолжает эволюционировать независимо. Наконец, наступает превращение подвидов в настоящие виды.

Рис. 83. Географическая изоляция популяций, сопровождающаяся видообразованием
Возникновение нового вида завершается лишь тогда, когда организмы в течение определенного периода изоляции приобретают признаки, которые способствуют или обеспечивают репродуктивную изоляцию даже после разрушения изолирующих преград. Таким образом, в генетическом смысле сущность видообразования заключается в возникновении двух хорошо интегрированных генных комплексов из одного родительского комплекса.
Постепенное видообразование применительно к разным организмам происходит с разной скоростью. Например, гоминиды эволю- ционировали очень быстро.
Новые виды образуются не из самых высокоразвитых видов, а из более простых. Эволюция не всегда идет от простого к сложно-
му. Напротив, есть и «регресс». Например, большинство паразитов произошли от свободноживущих организмов, для которых характерна более сложная организация, а бескрылые насекомые - от крылатых, киты - от четвероногих млекопитающих.
Мгновенное (квантовое) или скачкообразное (сальтационное) видообразование наблюдается в результате мутаций и сокращения сроков действия репродуктивных изолирующих механизмов. Примером такого видообразования является возникновение в течение одного поколения полиплоидов растений. Видообразование через полиплоидию известно и у животных, хотя у последних этот механизм очень редок. Полиплоиды считают самостоятельными видами не только по причине резкого отличия их от исходных видов, но и потому, что они репродуктивно изолированы от исходных видов.
Эволюция всегда необратима. Это означает, что исчезнувшие виды никогда не возвращаются. С другой стороны, существенное место в теории эволюции имеет проблема устойчивости существующих видов. Устойчивость видов - это историческая реальность. В видах удерживается все то, что достигнуто в процессе эволюции. Если бы виды не обладали устойчивостью, то закрепление результатов эволюции просто было бы невозможно. Следовательно, невозможной была бы и сама эволюция. Однако устойчивость видов не является абсолютной. Виды подвергаются изменчивости, которая создает возможности для гибкого приспособления организмов к меняющейся среде. Можно сказать, что посредством изменчивости виды могут «маневрировать», не теряя основных характеристик.
Эволюция протекает на всех уровнях организации живой материи, причем каждый эволюционный уровень является результатом низшего и материалом для высшего уровня. Ресурсы естественного отбора также имеются на всех уровнях. Организмы каждого нового эволюционного уровня обладают потенциальной возможностью для более быстрого развития по сравнению с предыдущим. Например, млекопитающие имеют больше возможностей для дальнейшей эволюции, чем одноклеточные животные.
ГИПОТЕЗА НЕЙТРАЛЬНОСТИ МОЛЕКУЛЯРНОЙ ЭВОЛЮЦИИ
В последние десятилетия стала известна гипотеза, в соответствии с которой большинство изменений, наблюдаемых в популяциях,
обусловлено мутациями, которые не оказывают влияния на приспособление организмов и заключаются в случайных заменах оснований. Скорости замен оснований в ДНК (аминокислот в белках) могут быть постоянными, так как преобладающее число таких замен селективно нейтрально. При одинаковой способности альтернативных аллелей придавать одинаковую приспособленность организмам изменение частот этих аллелей в поколениях организмов может происходить лишь в результате дрейфа генов. Следовательно, скорость замены аллелей в популяциях и скорость происходящих отсюда замен ами- нокислот будет постоянной.
По гипотезе нейтральности молекулярной эволюции большинство мутаций любых генов вредно для организмов и мутанты элиминируются с помощью естественного отбора. Однако могут возникать мутации, которые не изменяют ни морфологических, ни функциональных признаков, в результате чего организмы - носители таких мутаций не подвержены естественному отбору. Следовательно, эволюция на молекулярном уровне в основном заключается в замене одних нейтральных генных аллелей другими, но такими же нейтральными аллелями. Сторонники гипотезы нейтральности эволюции на молекулярном уровне считают, что она не противоречит дарвинизму.
Многие эволюционисты придают большое значение генамрегуляторам. Предполагают, что мутации регуляторных генов очень важны для адаптивной эволюции, однако данные в пользу этого предположения еще незначительны.
АНТИДАРВИНОВСКИЕ КОНЦЕПЦИИ ЭВОЛЮЦИИ
С самого начала теория эволюции оказалась ареной острой идеологической борьбы, в которой дарвинизму стали противопоставлять неоламаркизм или подменять его социал-дарвинизмом.
Среди неоламаркистских взглядов различают психоламаркизм и механоламаркизм. Психоламаркисты в качестве главного фактора эволюции называли психику. По их мысли эволюцию определяет выработка привычек к новым действиям, постепенное превращение сознательных актов в автоматические. Психоламаркисты отрицали естественный отбор и считали, что наследственность - это память, которая запечатлевает все воздействия внешней среды, вос- производящиеся в последующих поколениях в адекватной форме.
Механоламаркисты полагали, что в основе эволюции лежит переход материи от недифференцированного состояния в более дифференцированное. Находясь в состоянии взаимодействия, организм и среда стремятся к равновесию, которое часто нарушается под воздействием факторов среды. Равновесие восстанавливается прямым уравновешиванием в результате непосредственного приспособления и косвенным уравновешиванием через естественный отбор. Основная методологическая ошибка неоламаркистских представителей заключалась в том, что они считали органическую целесообразность свойством организмов, данным изначально.
В XIX в. возник и перешел в XX в. социал-дарвинизм, основателем которого был немецкий ученый Э. Геккель (1834-1919), много сделавший для защиты и пропаганды дарвинизма. Однако в трактовке эволюции Э. Геккель и другие социал-дарвинисты приближались к Ламарку, признавая на равных условиях отбор и прямое приспособление на основе унаследования полезных признаков. Но главная их ошибка заключалась в том, что они стремились использовать биологические закономерности для объяснения закономерностей развития общества. Главным двигателем общественного развития они считали борьбу за существование и естественный отбор. На основе естественного отбора они пытались обосновать возникновение классов и классового общества, объяснить низкий жизненный уровень народов развивающихся стран биологическими причинами. В прошлом социал-дарвинисты часто смыкались с неомальтузианством, объяснявшим трудности развивающихся стран чрезмерным ростом населения. В новейшее время социал-дарвинизм слился с евгеникой, став одной из форм современной биократической идеологии.
В новейшее время оживился также креационизм (см. выше).
ВОПРОСЫ ДЛЯ ОБСУЖДЕНИЯ
1. В чем заключается эволюция?
2. На чем основываются современные доводы в пользу эволюции?
3. Почему современную теорию эволюции называют синтетической и в чем состоит ее отличие от классического дарвинизма?
4. В чем заключается сущность современного понимания происхождения жизни? Почему современная теория происхождения жизни является материалистической?
5. Что является главным в учении Ч. Дарвина?
6. Какова роль случайности в эволюции?
7. Назовите главные направления эволюции, приведите примеры.
8. Дайте определение макро- и микроэволюции. Каково соотношение между макро- и микроэволюцией?
9. Назовите элементарные единицы, явления, материал и факторы эволюции.
10. Дайте определение популяции. Почему элементарной единицей эволюции является популяция?
11. Что представляет собой естественный отбор? Какие формы естественного отбора вы знаете? Какова роль движущегося отбора?
12. Что вы знаете об отборе в пользу гетерозигот? Расскажите о серповидноклеточной анемии человека и объясните, почему в малярийных районах гетерозиготы HbAHbS обладают селективными преимуществами.
13. Дайте определение вида.
14. Дайте определения анагенеза и кладогенеза, есть ли разница между этими процессами?
15. Что вы знаете о скорости эволюции? Как вы понимаете механизм видообразования?
16. Обратима ли эволюция? Почему виды устойчивы, какое это имеет значение для эволюции?
17. Происходит ли образование новых видов в современную эпоху?
18. Можно ли считать новыми видами организмы, создаваемые методом генной и клеточной инженерии?
19. Что вы знаете о гипотезе нейтральности молекулярной эволюции?
20. Какие вы знаете антидарвиновские концепции эволюции?
21. В чем заключается общенаучное значение теории эволюции? Каково мировоззренческое значение теории эволюции?
22. Какова роль теории эволюции в медицине?