Медицинская микробиология, вирусология и иммунология: том 1 : учебник / Под ред. В.В. Зверева, М.Н. Бойченко, 2010.
|
|
|
|
ГЛАВА 5 ГЕНЕТИКА МИКРОБОВ
5.1. Строение генома бактерий
Наследственная информация хранится у бактерий в форме последовательности нуклеотидов ДНК, которые определяют последовательность аминокислот в белке (строение ДНК изложено в разделе 3.1 и показано на рис. 3.1).
Каждому белку соответствует свой ген, т.е. дискретный участок на ДНК, отличающийся числом и специфичностью последовательности нуклеотидов.
Совокупность всех генов бактерий называется геномом. Размеры генома определяются количеством нуклеотидных пар оснований (н.п.). Геном бактерий имеет гаплоидный набор генов. Бактериальный геном состоит из генетических элементов, способных к самостоятельной репликации (воспроизведению), т.е. репликонов. Репликонами являются бактериальная хромосома и плазмиды.
5.1.1. Бактериальная хромосома
Бактериальная хромосома представлена одной двухцепочечной молекулой ДНК. Размеры бактериальной хромосомы у различных представителей домена Procaryotae варьируют. Например, у Е. coli бактериальная хромосома содержит 4,7х106 н.п. На ней располагается около 4300 генов. Для сравнения: размеры ДНК вирусов составляют порядка 103 н.п., дрожжей - 107 н.п., а суммарная длина хромосомных ДНК человека составляет 3х109 н.п.
Бактериальная хромосома у Е. coli представлена 1 кольцевой молекулой ДНК. Одну кольцевую хромосому имеет также ряд других бактерий: Shigella spp, Salmonella spp, P. aeruginosa, B. subtilus. Однако такое строение генома не является универсальным. У некоторых бактерий, в частности у V. cholerae, L. interrhogans, Brucella spp., име-
ется две кольцевых хромосомы. У ряда других бактерий (B. burgdorferi, Streptomyces spp.) обнаружены линейные хромосомы.
Бактериальная хромосома формирует компактный нуклеоид бактериальной клетки. Она кодирует жизненно важные для бактериальной клетки функции.
5.1.2. Плазмиды бактерий
Плазмиды представляют собой двухцепочечные молекулы ДНК размером от 103 до 106 н.п. Они могут быть кольцевой формой и линейными. Плазмиды кодируют не основные для жизнедеятельности бактериальной клетки функции, но придающие бактерии преимущества при попадании в неблагоприятные условия существования.
Среди фенотипических признаков, сообщаемых бактериальной клетке плазмидами, можно выделить следующие:
• устойчивость к антибиотикам;
• продукцию факторов патогенности;
• способность к синтезу антибиотических веществ;
• образование колицинов;
• расщепление сложных органических веществ;
• образование ферментов рестрикции и модификации. Репликация плазмид происходит независимо от хромосомы с
участием того же набора ферментов, который осуществляет репликацию бактериальной хромосомы (см. раздел 3.1.7 и рис. 3.5).
Некоторые плазмиды находятся под строгим контролем. Это означает, что их репликация сопряжена с репликацией хромосомы так, что в каждой бактериальной клетке присутствует одна или, по крайней мере, несколько копий плазмид.
Число копий плазмид, находящихся под слабым контролем, может достигать от 10 до 200 на бактериальную клетку.
Для характеристики плазмидных репликонов их принято разбивать на группы совместимости. Несовместимость плазмид связана с неспособностью двух плазмид стабильно сохраняться в одной и той же бактериальной клетке. Несовместимость свойственна тем плазмидам, которые обладают высоким сходством репликонов, поддержание которых в клетке регулируется одним и тем же механизмом.
Плазмиды, которые могут обратимо встраиваться в бактериальную хромосому и функционировать в виде единого репликона, называются интегративными или эписомами.
Плазмиды, способные передаваться из одной клетки в другую, иногда даже принадлежащую иной таксономической единице, называются трансмиссивными (конъюгативными) Трансмиссивность присуща лишь крупным плазмидам, имеющим tra-оперон, в который объединены гены, ответственные за перенос плазмиды. Эти гены кодируют половые пили, которые образуют мостик с клеткой, не содержащей трансмиссивную плазмиду, по которой плазмидная ДНК передается в новую клетку. Этот процесс называется конъюгацией (подробно он будет рассмотрен в разделе 5.4.1). Бактерии, несущие трансмиссивные плазмиды, чувствительны к «мужским» нитевидным бактериофагам.
Мелкие плазмиды, не несущие tra-гены, не могут передаваться сами по себе, но способны к передаче в присутствии трансмиссивных плазмид, используя их аппарат конъюгации. Такие плазмиды называются мобилизуемыми, а сам процесс - мобилизацией нетрансмиссивной плазмиды.
Особое значение в медицинской микробиологии имеют плазмиды, обеспечивающие устойчивость бактерий к антибиотикам, которые получили название R-плазмид (от англ. resistance - противодействие), и плазмиды, обеспечивающие продукцию факторов патогенности, способствующих развитию инфекционного процесса в макроорганизме. R-плазмиды содержат гены, детерминирующие синтез ферментов, разрушающих антибактериальные препараты (например, антибиотики). В результате наличия такой плазмиды бактериальная клетка становится устойчивой (резистентной) к действию целой группы лекарственных веществ, а иногда и к нескольким препаратам. Многие R-плазмиды являются трансмиссивными, распространяясь в популяции бактерий, делая ее недоступной к воздействию антибактериальных препаратов. Бактериальные штаммы, несущие R-плазмиды, очень часто являются этиологическими агентами внутрибольничных инфекций.
Плазмиды, детерминирующие синтез факторов патогенности, в настоящее время обнаружены у многих бактерий, являющихся возбудителями инфекционных заболеваний человека. Патогенность возбудителей шигеллезов, иерсиниозов, чумы, сибирской язвы, иксодового бореллиоза, кишечных эшерихиозов связана с наличием у них и функционированием плазмид патогенности.
Некоторые бактериальные клетки содержат плазмиды, детерминирующие синтез бактерицидных по отношению к другим бактери-
ям веществ. Например, некоторые Е. coli владеют Col-плазмидой, определяющей синтез колицинов, обладающих микробоцидной активностью по отношению к колиформным бактериям. Бактериальные клетки, несущие такие плазмиды, обладают преимуществами при заселении экологических ниш.
Плазмиды используются в практической деятельности человека, в частности в генной инженерии при конструировании специальных рекомбинантных бактериальных штаммов, вырабатывающих в больших количествах биологически активные вещества (см. главу 6).
5.1.3. Подвижные генетические элементы
Подвижные генетические элементы обнаружены в составе бактериального генома как в бактериальной хромосоме, так и в плазмидах. К подвижным генетическим элементам относятся вставочные последовательности и транспозоны.
Вставочные (инсерционные) последовательности - IS-элементы (от англ. insertion sequences) - это участки ДНК, способные как целое перемещаться из одного участка репликона в другой, а также между репликонами. IS-элементы имеют размеры 1000 н.п. и содержат лишь те гены, которые необходимы для их собственного перемещения - транспозиции: ген, кодирующий фермент транспозазу, обеспечивающую процесс исключения IS-элемента из ДНК и его интеграцию в новый локус, и ген, детерминирующий синтез репрессора, который регулирует весь процесс перемещения. Эти гены по флангам окружены инвертированными повторами, которые служат сайтами рекомбинации, сопровождающей перемещения вставочной последовательности при участии транспозиционных ферментов, в частности транспозаз.
Инвертированные повторы узнает фермент транспозаза (рис. 5.1), которая осуществляет одноцепочечные разрывы цепей ДНК, расположенных по обе стороны от подвижного элемента. Оригинальная копия IS-элемента остается на прежнем месте, а ее реплицированный дубликат перемещается на новый участок.
Перемещение подвижных генетических элементов принято называть репликативной или незаконной рекомбинацией. Однако, в отличие от бактериальной хромосомы и плазмид, подвижные генетические элементы не являются самостоятельными репликонами,
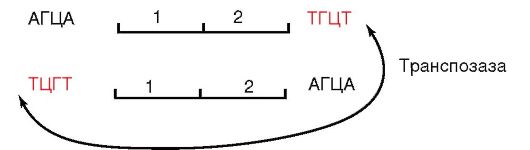 Рис. 5.1. Схема строения IS-элемента: 1 - ген репрессора; 2 - ген транспозазы; стрелками указаны места разрывов
Рис. 5.1. Схема строения IS-элемента: 1 - ген репрессора; 2 - ген транспозазы; стрелками указаны места разрывов
так как их репликация - составной элемент репликации ДНК репликона, в составе которого они находятся.
IS-элементы различаются по размеру, типу и количеству инвертированных повторов.
Транспозоны - это сегменты ДНК, обладающие теми же свойствами, что и IS-элементы, но имеющие в своем составе структурные гены, т.е. гены, обеспечивающие синтез молекул, обладающих специфическим биологическим свойством, например токсичностью, или обеспечивающих устойчивость к антибиотикам.
Перемещение подвижных генетических элементов по репликону или между репликонами вызывает:
• инактивацию генов тех участков ДНК, куда они, переместившись, встраиваются;
• образование повреждений генетического материала;
• слияние репликонов, т.е. встраивание плазмиды в хромосому;
• распространение генов в популяции бактерий, что может приводить к изменению биологических свойств популяции, смене возбудителей инфекционных заболеваний, а также способствует эволюционным процессам среди микробов.
5.1.4. Интегроны
Помимо плазмид и подвижных генетических элементов, у бактерий существует еще одна система, способствующая распространению генов, - система интегронов. Интегроны являются системой захвата малых элементов ДНК, называемых генными кассетами, посредством сайтспецифической рекомбинации и их экспрессии.
Интегрон состоит из консервативного участка, расположенного на 5' конце, который содержит ген, кодирующий фермент интегразу, сайт рекомбинации att и промотор Р (рис. 5.2).
Кассета может существовать в двух формах: линейной, когда кассета интегрирована в интегрон, и в виде маленькой кольцевой двухцепочечной ДНК. Кассеты имеют размеры от 260 до 1500 н.п. Они содержат преимущественно 1 ген антибиотикорезистентности и сайт рекомбинации, состоящий из 59 пар оснований, расположенный на З'-конце.
Интеграза осуществляет рекомбинацию между участком 59 н.п. кассеты и участком att интегрона, включая гены кассеты в интегрон в такой ориентации, чтобы они могли экспрессироваться с промотора Р интегрона. Интеграция кассет в интегрон является обратимым процессом. Интегроны могут располагаться как на хромосоме, так и на плазмидах. Поэтому возможно перемещение кассет с одного интегрона на другой как в пределах одной бактериальной клетки, так и по популяции бактерий. Один интегрон может захватывать несколько кассет антибиотикорезистентности. Изменения
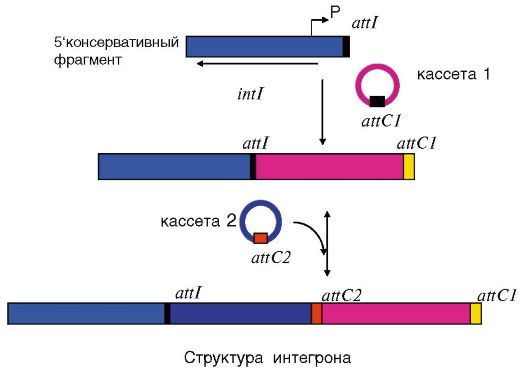 Рис. 5.2. Строение интегрона: attI - сайт рекомбинации интегрона; intI - ген, кодирующий интегразу; P - промотор; attC - сайты рекомбинации кассет антибиотикорезистентности
Рис. 5.2. Строение интегрона: attI - сайт рекомбинации интегрона; intI - ген, кодирующий интегразу; P - промотор; attC - сайты рекомбинации кассет антибиотикорезистентности
бактериального генома, а следовательно, и свойств бактерий могут происходить в результате мутаций и рекомбинаций.
5.1.5. Острова патогенности
В геноме патогенных бактерий (см. главу 8) имеются участки ДНК протяженностью не менее 10 000 пар нуклеотидов, которые отличаются от основного генома составом Г-Ц-пар нуклеотидных оснований. Эти участки ответственны за синтез факторов патогенности, которые обеспечивают развитие патологического процесса в организме хозяина, поэтому были названы островами патогенности. Острова патогенности обычно по флангам имеют прямые повторы последовательностей ДНК или IS-элементы. Некоторые имеют в своем составе участки, характерные для сайтов интеграции, расположенных вблизи генов тРНК. Большинство островов патогенности локализовано на хромосоме бактерий (Salmonella), но также они могут находиться в составе плазмид (Shigella) и фаговых ДНК (V. cholerae O1, O139).
5.2. Мутации у бактерий
Мутации - это изменения в последовательности отдельных нуклеотидов ДНК, которые фенотипически ведут к таким проявлениям, как изменения морфологии бактериальной клетки, возникновение потребностей в факторах роста, например в аминокислотах, витаминах, т.е. ауксотрофности, к устойчивости к антибиотикам, изменению чувствительности к температуре, снижению вирулентности (аттенуация) и т.д.
Мутация, приводящая к потере функции, называется прямой мутацией. У мутантов может произойти восстановление исходных свойств, т.е. реверсия (от англ. reverse - обратный). Если происходит восстановление исходного генотипа, то мутация, восстанавливающая генотип и фенотип, называется обратной или прямой реверсией. Если мутация восстанавливает фенотип, не восстанавливая генотип, то такая мутация называется супрессорной. Супрессорные мутации могут возникать как в пределах того самого гена, в котором произошла первичная мутация, так и в других генах или могут быть связаны с мутациями в тРНК.
По протяженности изменений повреждения ДНК различают мутации точечные, когда повреждения ограничиваются одной
парой нуклеотидов, и протяженные или аберрации. В последнем случае могут наблюдаться выпадения нескольких пар нуклеотидов, которые называются делецией, добавление нуклеотидных пар, т.е. дупликации, перемещения фрагментов хромосомы, транслокации и перестановки нуклеотидных пар - инверсии.
Мутации могут быть спонтанными, т.е. возникающими самопроизвольно, без воздействия извне, и индуцированными.
Точечные спонтанные мутации возникают в результате ошибок при репликации ДНК, что связано с таутомерным перемещением электронов в азотистых основаниях.
Тимин (Т), например, обычно находится в кетоформе, в которой он способен образовывать водородные связи с аденином (А). Но если тимин во время спаривания оснований при репликации ДНК переходит в енольную форму, то он спаривается с гуанином. В результате в новой молекуле ДНК на месте, где раньше стояла пара А-Т, появляется пара Г-Ц.
Спонтанные хромосомные аберрации возникают вследствие перемещения подвижных генетических элементов. Индуцированные мутации появляются под влиянием внешних факторов, которые называются мутагенами. Мутагены бывают физическими (УФ-лучи, γ-радиация), химическими (аналоги пуриновых и пиримидиновых оснований, азотистая кислота и ее аналоги и другие соединения) и биологическими - транспозоны.
Аналоги пуриновых и пиримидиновых оснований, например 2-аминопурин, 5-бромурацил, включаются в нуклеотиды, а следовательно, и в ДНК, но при этом они значительно чаще в силу таутомерных превращений спариваются с «неправильными» партнерами, в результате вызывая замену пурина другим пурином (А-Г) или пиримидина другим пиримидином (Т-Ц). Замена пурина другим пурином, а пиримидина другим пиримидином называется транзицией.
Азотистая кислота и ее аналоги вызывают дезаминирование азотистых оснований, результатом чего являются ошибки при спаривании и как следствие возникновение транзиции. Аденин в результате дезаминирования превращается в гипоксантин, который спаривается с цитозином, что приводит к возникновению транзиции AT-ГЦ. Гуанин же при дезаминировании превращается в ксантин, который по-прежнему спаривается с цитозином; таким образом, дезаминирование гуанина не сопровождается мутацией.
Акридин и профлавин внедряются между соседними основаниями цепи ДНК, вдвое увеличивая расстояние между ними. Это пространственное изменение при репликации может привести как к утрате нуклеотида, так и к включению дополнительной нуклеотидной пары, что приводит к сдвигу рамки считывания тРНК. Начиная с того места, где произошло выпадение или включение нуклеотида, информация считывается неправильно.
УФ-облучение затрагивает преимущественно пиримидиновые основания, при этом два соседних остатка тимина ДНК могут оказаться ковалентно связанными.
На бактериях, подвергнутых УФ-облучению, было показано, что повреждения, вызванные облучением в бактериальных ДНК, могут частично исправляться благодаря наличию репарационных систем. У различных бактерий имеется несколько типов репарационных систем. Один тип репарации протекает на свету, он связан с деятельностью фотореактивирующегося фермента, который расщепляет тиминовый димер. При темновой репарации дефектные участки цепи ДНК удаляются и образовавшаяся брешь достраивается при помощи ДНК-полимеразы на матрице сохранившейся цепи и соединяется с цепью лигазой.
5.3. Рекомбинация у бактерий
Генетическая рекомбинация - это взаимодействие между двумя ДНК, обладающими различными генотипами, которое приводит к образованию рекомбинантной ДНК, сочетающей гены обоих родителей.
Особенности рекомбинации у бактерий определяет отсутствие полового размножения и мейоза, в процессе которых у высших организмов происходят рекомбинация, гаплоидный набор генов. В процессе рекомбинации бактерии условно делятся на клеткидоноры, которые передают генетический материал, и клеткиреципиенты, которые воспринимают его. В клетку-реципиент проникает не вся, а только часть хромосомы клетки-донора, что приводит к формированию неполной зиготы - мерозиготы. В результате рекомбинации в мерозиготе образуется только один рекомбинант, генотип которого представлен в основном генотипом реципиента, с включенным в него фрагментом хромосомы донора. Реципрокные рекомбинанты не образуются.
По молекулярному механизму генетическая рекомбинация у бактерий делится на гомологичную, сайтспецифическую и незаконную.
5.3.1. Гомологичная рекомбинация
При гомологичной рекомбинации в процессе разрыва и воссоединения ДНК происходит обмен между участками ДНК, обладающими высокой степенью гомологии. Процесс гомологичной рекомбинации находится под контролем генов, объединенных в REC-систему, состоящую из генов recA, B, C, D. Продукты этих генов производят расплетание нитей ДНК и их переориентацию с образованием полухиазмы, структуры Холидея, а также разрезают структуру Холидея для завершения процесса рекомбинации.
5.3.2. Сайтспецифическая рекомбинация
Этот тип рекомбинации не зависит от функционирования генов recA, B, C, D, не требует протяжных участков гомологии ДНК, но для протекания которой необходимы строго определенные последовательности ДНК и специальный ферментативный аппарат, которые специфичны для каждого конкретного случая. Примером этого типа рекомбинации является встраивание плазмиды в хромосому бактерий, которое происходит между идентичными IS- элементами хромосомы и плазмиды, интеграция ДНК фага лямбда в хромосому Е. coli. Сайтспецифическая рекомбинация, происходящая в пределах одного репликона, участвует также в переключении активности генов. Например, у сальмонелл следствием этого процесса являются фазовые вариации жгутикового Н-антигена.
5.3.3. Незаконная или репликативная рекомбинация
Незаконная или репликативная рекомбинация не зависит от функционирования генов recA, B, C, D. Примером ее является транспозиция подвижных генетических элементов по репликону или между репликонами, при этом, как уже было отмечено в разделе 5.1.3, транспозиция подвижного генетического элемента сопровождается репликацией ДНК.
Рекомбинация у бактерий является конечным этапом передачи генетического материала между бактериями, которая осуществляется тремя механизмами: конъюгацией (при контакте бактерий,
одна из которых несет конъюгативную плазмиду), трансдукцией (при помощи бактериофага), трансформацией (при помощи высокополимеризованной ДНК).
5.4. Передача генетической информации у бактерий 5.4.1. Конъюгация
Передача генетического материала от клетки-донора в клеткурепипиент путем непосредственного контакта клеток называется конъюгацией, которая впервые была обнаружена Дж. Ледербергом и Э. Тейтумом в 1946 г.
Необходимым условием для конъюгации является наличие в клетке-доноре трансмиссивной плазмиды. Трансмиссивные плазмиды кодируют половые пили, образующие конъюгационную трубочку между клеткой-донором и клеткой-реципиентом, по которой плазмидная ДНК передается в новую клетку. Механизм передачи плазмидной ДНК из клетки в клетку заключается в том, что специальный белок, кодируемый tra-опероном, узнает определенную последовательность в ДНК плазмиды (называемую от англ. origin - начало), вносит в эту последовательность одноцепочечный разрыв и ковалентно связывается с 5'-концом. Затем цепь ДНК, с которой связан белок, переносится в клетку-реципиент, а неразорванная комплементарная цепь остается в клетке-доноре. Клеточный аппарат синтеза ДНК достраивает одиночные цепи и в доноре, и в реципиенте до двухцепочечной структуры. Белок, связанный с 5'-концом перенесенной цепи, способствует замыканию плазмиды в реципиентной клетке в кольцо. Этот процесс представлен на рис. 5.3, а на примере переноса в реципиентную клетку плазмиды F (от англ. fertility - плодовитость), которая является как трансмиссивной, так и интегративной плазмидой. Клеткидоноры, обладающие F-фактором, обозначаются как F+-клетки, а клетки-реципиенты, не имеющие F-фактора, обозначаются как F-- клетки. Если F-фактор находится в клетке-доноре в автономном состоянии, то в результате скрещивания F+ * F- клетка-реципиент приобретает донорские свойства.
Если F-фактор или другая трансмиссивная плазмида встраиваются в хромосому клетки-донора, то плазмида и хромосома начинают функционировать в виде единого трансмиссивного репликона, что делает возможным перенос бактериальных генов в бесплаз-
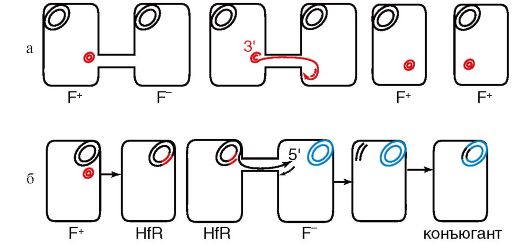 Рис. 5.3. Схема конъюгации у бактерий: а - передача F плазмиды из F+- в F--клетку; б - передача бактериальной хромосомы Hfr * F-
Рис. 5.3. Схема конъюгации у бактерий: а - передача F плазмиды из F+- в F--клетку; б - передача бактериальной хромосомы Hfr * F-
мидную клетку-реципиент, т.е. процесс конъюгации. Штаммы, в которых плазмида находится в интегрированном состоянии, переносят свои хромосомные гены бесплазмидным клеткам с высокой частотой и поэтому называются Hfr (от англ. high frequency of recombination - высокая частота рекомбинации) (рис. 5.3, б).
Процесс переноса хромосомных генов в случае скрещивания Hfr χ F- всегда начинается с расщепления ДНК в одной и той же точке - в месте интеграции F-фактора или другой трансмиссивной плазмиды. Одна нить донорской ДНК передается через конъюгационный мостик в реципиентную клетку. Процесс сопровождается достраиванием комплементарной нити до образования двунитевой структуры. Перенос хромосомных генов при конъюгации всегда имеет одинаковую направленность, противоположную встроенной плазмиде. Сама трансмиссивная плазмида передается последней. Переданная в реципиентную клетку и достроенная до двунитевой структуры нить ДНК донора рекомбинирует с гомологичным участком реципиентной ДНК с образованием стабильной генетической структуры. Из-за хрупкости конъюгационного мостика половой фактор редко передается в клетку-реципиент, поэтому образовавшийся рекомбинант донорскими функциями, как правило, не обладает.
Вследствие направленности передачи генов конъюгация используется для картирования генома бактерий и построения генетической карты.
5.4.2. Трансдукция
Трансдукцией называют передачу бактериальной ДНК посредством бактериофага. Этот процесс был открыт в 1951 г. Н. Циндером и Дж. Ледербергом. В процессе репликации фага внутри бактерий (см. раздел 3.3) фрагмент бактериальной ДНК проникает в фаговую частицу и переносится в реципиентную бактерию во время фаговой инфекции. Существует два типа трансдукции: общая трансдукция - перенос бактериофагом сегмента любой части бактериальной хромосомы - происходит вследствие того, что в процессе фаговой инфекции бактериальная ДНК фрагментируется, и фрагмент бактериальной ДНК того же размера, что и фаговая ДНК, проникает в фаговую головку, формируя дефектную фаговую частицу. Этот процесс происходит с частотой приблизительно 1 на 1000 фаговых частиц (рис. 5.4, а). При инфицировании клетки-реципиента дефектной фаговой частицей ДНК клеткидонора «впрыскивается» в нее и рекомбинирует гомологичной рекомбинацией с гомологичным участком хромосомы-реципиента с образованием стабильного рекомбинанта. Этим типом трансдукции обладают Р-фаги. Специфическая трансдукция наблюдается в том случае, когда фаговая ДНК интегрирует в бактериальную хромосому с образованием профага. В процессе исключения ДНКфага из бактериальной хромосомы в результате случайного процесса захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы, становясь дефектным фагом (рис. 5.4, б). Так как большинство умеренных бактериофагов интегрирует в бактериальную хромосому в специфических участках, для таких бактериофагов характерен перенос в клетку-реципиент определенного участка бактериальной ДНК клетки-донора. ДНК дефектного фага рекомбинирует с ДНК клетки-реципиента сайтспецифической рекомбинацией. Рекомбинант становится меродиплоидом по привнесенному гену. В частности, бактериофаг передает специфической трансдукцией gal-ген у Е. coli.
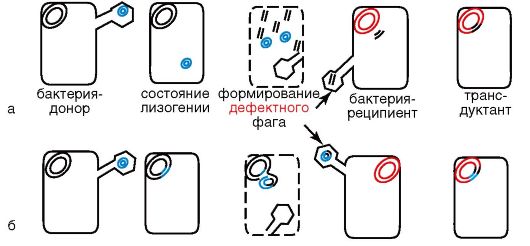 Рис. 5.4. Схема трансдукции: а - неспецифическая (общая); б - специфическая
Рис. 5.4. Схема трансдукции: а - неспецифическая (общая); б - специфическая
5.4.3. Трансформация
Феномен трансформации впервые был описан в 1928 г. Ф. Гриффитсом, обнаружившим превращение бескапсульного R-штамма пневмококков (Streptococcus pneumoniae) в штамм, образующий капсулу S-формы. Гриффитс ввел мышам одновременно небольшое количество авирулентных R-клеток и убитых нагреванием S-клеток. R-клетки были получены от штамма, капсульное вещество которого принадлежало к типу S II, а убитые нагреванием S-штаммы - к типу S III. Из крови погибших мышей были выделены вирулентные пневмококки с капсулой S III.
В 1944 г. О. Эвери, К. Мак-Леод, М. Мак-Карти установили природу трансформирующего фактора, показав, что ДНК, экстрагированная из инкапсулированных пневмококков, может трансформировать некапсулированные пневмококки в инкапсулированную форму. Таким образом, было доказано, что именно ДНК является носителем генетической информации.
Процесс трансформации может самопроизвольно происходить в природе у некоторых видов бактерий, B. subtilis, H. influenzae, S. pneumoniae, когда ДНК, выделенная из погибших клеток, захватывается реципиентными клетками. Процесс трансформации зависит от компетентности клетки-реципиента и состояния донорской трансформирующей ДНК. Компетентность - это спо-
собность бактериальной клетки поглощать ДНК. Она зависит от присутствия особых белков в клеточной мембране, обладающих специфическим аффинитетом к ДНК. Состояние компетентности у грамположительных бактерий связано с определенными фазами кривой роста. Состояние компетенции у грамотрицательных бактерий приходится создавать искусственным путем, подвергая бактерии температурному или электрошоку.
Трансформирующей активностью обладает только двунитевая высокоспирализованная молекула ДНК. Это связано с тем, что в клетку-реципиент проникает только одна нить ДНК, тогда как другая - на клеточной мембране - подвергается деградации с высвобождением энергии, которая необходима для проникновения в клетку сохранившейся нити. Высокая молекулярная масса трансформирующей ДНК увеличивает шанс рекомбинации, так как внутри клетки трансформирующая нить ДНК подвергается воздействию эндонуклеаз. Интеграция с хромосомой требует наличия гомологичных с ней участков у трансформирующей ДНК. Рекомбинация происходит на одной нити, в результате чего образуется гетеродуплексная молекула, одна нить которой имеет генотип реципиента, а другая - рекомбинантный генотип. Рекомбинантные трансформанты формируются только после цикла репликации (рис. 5.5).
В настоящее время этот метод является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
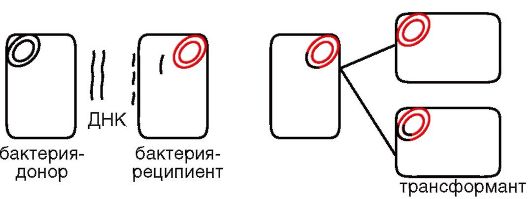 Рис. 5.5. Схема трансформации
Рис. 5.5. Схема трансформации
5.5. Особенности генетики вирусов
Особенность строения вирусного генома заключается в том, что наследственная информация может быть записана как на ДНК, так и на РНК в зависимости от типа вируса.
Мутации у вирусов могут возникать спонтанно в процессе репликации нуклеиновой кислоты вируса, а также под влиянием тех же внешних факторов и мутагенов, что и у бактерий.
Фенотипически мутации вирусного генома проявляются изменениями антигенной структуры, неспособностью вызывать продуктивную инфекцию в чувствительной клетке, чувствительностью продуктивного цикла к температуре, а также изменением формы и размера бляшек, которые образуют вирусы в культуре клеток под агаровым покрытием (см. главу 3.2).
Свойства вирусов могут изменяться при одновременном заражении несколькими вирусами чувствительной клетки, причем изменения свойств при таких условиях могут происходить в результате как обмена между материалами нуклеиновых кислот, принадлежащих разным вирусам (генетическая рекомбинация и генетическая реактивация), так и процессов, не сопровождающихся обменом генетического материала (комплементация и фенотипическое смешивание).
Генетическая рекомбинация встречается чаще у ДНК-содержащих вирусов. Среди РНК-содержащих вирусов она наблюдается у тех из них, которые обладают фрагментированным геномом, например у вируса гриппа. При рекомбинации происходит обмен между гомологичными участками генома.
Генетическая реактивация наблюдается между геномами родственных вирусов, имеющих мутации в разных генах. В результате перераспределения генетического материала формируется полноценный дочерний геном.
Комплементация встречается в том случае, когда один из двух вирусов, инфицирующих клетку, в результате мутации синтезирует нефункциональный белок. Немутантный вирус, синтезируя полноценный белок, восполняет его отсутствие у мутантного вируса.
Фенотипическое смешивание наблюдается в том случае, если при смешанном заражении чувствительной клетки двумя вирусами часть потомства приобретает фенотипические признаки, присущие двум вирусам, при сохранении неизменности генотипа.
5.6. Применение генетических методов в диагностике инфекционных болезней
Генетические методы применяются в практических целях как для обнаружения микроба в исследуемом материале без выделения чистой культуры, так и для определения таксономического положения микроба и проведения внутривидовой идентификации.
5.6.1. Методы, используемые для внутривидовой идентификации бактерий
Рестрикционный анализ основан на применении ферментов, носящих название рестриктаз. Рестриктазы представляют собой эндонуклеазы, которые расщепляют молекулы ДНК, разрывая фосфатные связи не в произвольных местах, а в определенных последовательностях нуклеотидов. Особое значение для методов молекулярной генетики имеют рестриктазы, которые узнают последовательности, обладающие центральной симметрией и считывающиеся одинаково в обе стороны от оси симметрии. Точка разрыва ДНК может или совпадать с осью симметрии, или быть сдвинута относительно нее.
В настоящее время из различных бактерий выделено и очищено более 175 различных рестриктаз, для которых известны сайты (участки) узнавания (рестрикции). Выявлено более 80 различных типов сайтов, в которых может происходить разрыв двойной спирали ДНК. В геноме конкретной таксономической единицы находится строго определенное (генетически задетерминированное) число участков узнавания для определенной рестриктазы. Если выделенную из конкретного микроба ДНК обработать определенной рестриктазой, то это приведет к образованию строго определенного количества фрагментов ДНК фиксированного размера. Размер каждого типа фрагментов можно узнать с помощью электрофореза в агарозном геле: мелкие фрагменты перемещаются в геле быстрее, чем более крупные фрагменты, и длина их пробега больше. Гель окрашивают бромистым этидием и фотографируют в УФ-излучении. Таким способом можно получить рестрикционную карту определенного вида микробов.
Сопоставляя карты рестрикции ДНК, выделенных из различных штаммов, можно определить их генетическое родство, выявить принадлежность к определенному виду или роду, а также обнару-
жить участки, подвергнутые мутациям. Этот метод используется также как начальный этап метода определения последовательности нуклеотидных пар (секвенирования) и метода молекулярной гибридизации.
Определение плазмидного профиля бактерий. Плазмидный профиль позволяет произвести внутривидовую идентификацию бактерий. Для этого из бактериальной клетки выделяют плазмидную ДНК, которую разделяют электрофорезом в агарозном геле для определения количества и размеров плазмид.
Риботипирование. Последовательность нуклеотидных оснований в оперонах, кодирующих рРНК, характеризуется наличием как консервативных участков, которые подверглись малым изменениям в процессе эволюции и имеют сходное строения у различных бактерий, так и вариабельных последовательностей, которые родо- и видо-специфичны и являются маркерами при генетической идентификации. Эти опероны представлены на бактериальной хромосоме в нескольких копиях. Фрагменты ДНК, полученные после обработки ее рестриктазами, содержат последовательности генов рРНК, которые могут быть обнаружены методом молекулярной гибридизации с меченой рРНК соответствующего вида бактерий. Количество и локализация копий оперонов рРНК и рестрикционный состав сайтов как внутри рРНК-оперона, так и по его флангам варьируют у различных видов бактерий. На основе этого свойства построен метод риботипирования, который позволяет производить мониторинг выделенных штаммов и определение их вида. В настоящее время риботипирование проводится в автоматическом режиме в специальных приборах.
5.6.2. Методы, используемые для обнаружения микроба без выделения его в чистую культуру
Метод молекулярной гибридизации позволяет выявить степень сходства различных ДНК. Применяется при идентификации микробов для определения их точного таксономического положения, а также для обнаружения микроба в исследуемом материале без его выделения в чистую культуру. Метод основан на способности двухцепочечной ДНК при повышенной температуре (90 °С) в щелочной среде денатурировать, т.е. расплетаться на две нити, а при понижении температуры на 10 °С вновь восстанавливать исходную двухцепочечную структуру. Метод требует наличия молекулярного зонда.
Зондом называется одноцепочечная молекула нуклеиновой кислоты, меченная радиоактивными нуклидами, ферментом, флюорохромным красителем, с которой сравнивают исследуемую ДНК.
Для проведения молекулярной гибридизации исследуемую ДНК расплетают указанным выше способом, одну нить фиксируют на специальном фильтре, который затем помещают в раствор, содержащий зонд. Создаются условия, благоприятные для образования двойных спиралей. При наличии комплементарности между зондом и исследуемой ДНК они образуют между собой двойную спираль, наличие которой фиксируют методами в зависимости от типа метки зонда: подсчетом радиоактивности, иммуноферментным анализом (ИФА) или денситометрией.
Определение наличия микроба в исследуемом материале при помощи микрочипа
Микрочип представляет стеклянную пластинку, с которой связано от 100 до 1000 молекулярных ДНК-зондов, которые представляют последовательность нуклеотидов, специфичных для данной таксономической единицы, локализованных в определенных участках (рис. 5.6).
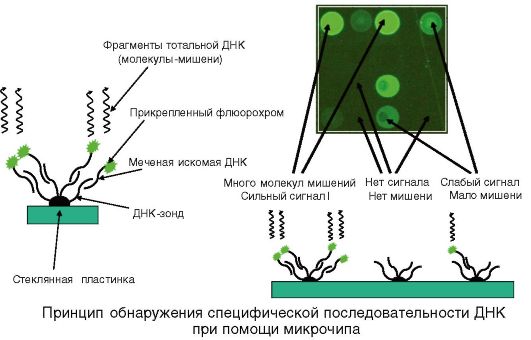 Рис. 5.6. Принцип обнаружения специфической последовательности ДНК при помощи микрочипа
Рис. 5.6. Принцип обнаружения специфической последовательности ДНК при помощи микрочипа
Из исследуемого образца выделяют общую ДНК, которую можно амплифицировать по стабильной последовательности 16S РНКгену. Выделенную ДНК метят флуорохромом или ферментом и обрабатывают ею микрочип, создавая условия для гибридизации. Отмывают несвязавшуюся ДНК, определяют локализацию молекулярных гибридов постановкой ИФА или денситометрией.
Полимеразная цепная реакция позволяет обнаружить микроб в исследуемом материале (воде, продуктах, материале от больного) по наличию в нем ДНК микроба без выделения последнего в чистую культуру.
Для проведения этой реакции из исследуемого материала выделяют ДНК, в которой определяют наличие специфичного для данного микроба гена. Обнаружение гена осуществляют его накоплением. Для этого необходимо иметь праймеры (затравки) комплиментарного З'-концам ДНК исходного гена. Накопление (амплификация) гена выполняют следующим образом. Выделенную из исследуемого материала ДНК нагревают. При этом ДНК распадается на 2 нити. Добавляют праймеры. Смесь ДНК и праймеров охлаждают. При этом праймеры при наличии в смеси ДНК искомого гена связываются с его комплементарными участками. Затем к смеси ДНК и праймера добавляют ДНК-полимеразу и нуклеотиды. Устанавливают температуру, оптимальную для функционирования ДНК-полимеразы. В этих условиях в случае комплементарности ДНК гена и праймера происходит присоединение нуклеотидов к З'-концам праймеров, в результате чего синтезируются две копии гена. После этого цикл повторяется снова, при этом количество ДНК гена увеличивается каждый раз вдвое (рис. 5.7). Проводят реакцию в специальных приборах - амплификаторах. Результат оценивают последующей денситометрией амплифицированной ДНК или ее электрофорезом в полиакриламидном геле. ПЦР применяют для диагностики вирусных и бактериальных инфекций.
ПЦР в реальном времени представляет ускоренный метод ПЦР, при котором амплификацию и определение продукта амплификации проводят одновременно. Для этой цели в амплификационную пробирку вводят молекулярный зонд, который в случае связывания с амплифицированной цепью генерирует флюоресцентный сигнал определенной длины волны. Реакцию проводят в автоматическом режиме.
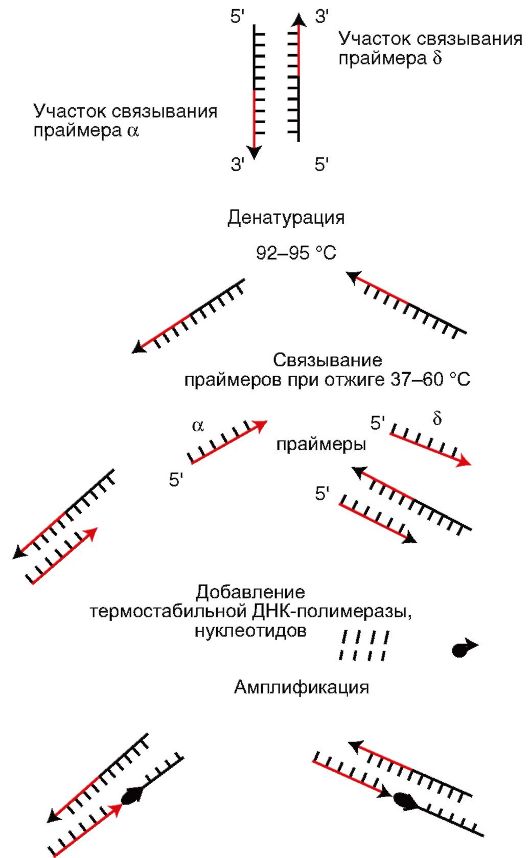 Рис. 5.7. Полимеразная цепная реакция (схема)
Рис. 5.7. Полимеразная цепная реакция (схема)
Опосредованная транскрипцией амплификация рРНК используется для диагностики смешанных инфекций. Этот метод основан на обнаружении с помощью молекулярной гибридизации амплифицированных рРНК, специфичных для определенного вида бактерий. Исследование проводят в три этапа:
• амплификация пула рРНК на матрице выделенной из исследуемого материала ДНК при помощи ДНК-зависимой РНКполимеразы;
• гибридизация накопленного пула рРНК с комплиментарными видоспецифическими рРНК олигонуклеотидами, меченными флюорохромом или ферментами;
• определение продуктов гибридизации методами денситометрии, ИФА.
Реакцию проводят в автоматическом режиме в установках, в которых одномоментное определение рРНК, принадлежащих различным видам бактерий, достигается разделением амплифицированного пула рРНК на несколько проб, в которые вносят для гибридизации комплементарные видоспецифическим рРНК меченые олигонуклеотиды.