Медицинская микробиология, вирусология и иммунология: том 1 : учебник / Под ред. В.В. Зверева, М.Н. Бойченко, 2010.
|
|
|
|
ГЛАВА 3 ФИЗИОЛОГИЯ МИКРОБОВ
3.1. Физиология бактерий
Физиология бактерий включает метаболизм бактерий, т.е. питание, получение энергии, рост и размножения бактерий, а также их взаимодействие с окружающей средой. Метаболизм бактерий лежит в основе изучения и разработки методов их культивирования, получения чистых культур и их идентификации. Выяснение физиологии патогенных и условно-патогенных бактерий важно для изучения патогенеза вызываемых ими инфекционных болезней, постановки микробиологического диагноза, лечения и профилактики инфекционных заболеваний, регуляции взаимоотношения человека с окружающей средой, а также для использования бактерий в биотехнологических процессах с целью получения биологически активных веществ.
3.1.1. Питание бактерий
Химический состав бактериальной клетки
Бактериальная клетка на 80-90% состоит из воды и только 10% приходится на долю сухого вещества. Вода в клетке находится в свободном или связанном состоянии. Она выполняет механическую роль в обеспечении тургора, участвует в гидролитических реакциях. Удаление воды из клетки путем высушивания приводит к приостановке процессов метаболизма, прекращению размножения, а для многих микроорганизмов губительно. В то же время особый способ высушивания микроорганизмов в вакууме из замороженного состояния (лиофилизация) обеспечивает сохранение жизнеспособности большинства микроорганизмов. Лиофилизация используется для приготовления проб, пригодных для длительного хранения.
В сухом веществе бактерий 52% составляют белки, 17% - углеводы, 9% - липиды, 16% - РНК, 3% - ДНК и 3% - минеральные вещества.
Белки являются ферментами, а также составной частью клетки, входят в состав цитоплазматической мембраны (ЦПМ) и ее производных, клеточной стенки, жгутиков, спор и некоторых капсул. Некоторые бактериальные белки являются антигенами и токсинами бактерий. В состав белков бактерий входят отсутствующие у человека D-аминокислоты, а также диаминопимелиновая кислота.
Углеводы представлены в бактериальной клетке в виде моно-, ди-, олигосахаров и полисахаридов, а также входят в состав комплексных соединений с белками, липидами и другими соединениями. Полисахариды входят в состав некоторых капсул, клеточной стенки; крахмал и гликоген являются запасными питательными веществами. Некоторые полисахариды принимают участие в формировании антигенов.
Липиды или жиры входят в состав ЦПМ и ее производных, клеточной стенки грамотрицательных бактерий, а также служат запасными веществами, входят в состав эндотоксина грамотрицательных бактерий, в составе ЛПС формируют антигены. В бактериальных жирах преобладают длинноцепочечные (С14-С18) насыщенные жирные кислоты и ненасыщенные жирные кислоты, содержащие одну двойную связь. Сложные липиды представлены фосфатидилинозитом, фосфатидилглицерином и фосфатидилэтаноламином. У некоторых бактерий в клетке находятся воски, эфиры миколовой кислоты. Микоплазмы - единственные представители царства Procaryotae, имеющие в составе ЦПМ стеролы. Остальные бактерии в составе ЦПМ и ее производных не имеют стеролов.
В бактериальной клетке присутствуют все типы РНК: иРНК, транспортная РНК (тРНК), рРНК, менее известная антисенс РНК (асРНК). Молекулы асРНК пока не обнаружены в клетках эукариот. Информация об асРНК записана в хромосоме, в так называемых антисенс-генах. АсРНК принимает активное участие в регуляции различных клеточных процессов, в том числе репликации ДНК бактерий, вирусов, плазмид и танспозонов. асРНК представляет собой короткую молекулу, комплементарную определенному участку иРНК, и, соединяясь с ней, блокирует процесс синтеза белка. При этом в клетке подобные комплексы могут накапли-
ваться, и при диссоциации асРНК и иРНК одновременно начинается синтез белка на большом числе однотипньгх матриц. Искусственные молекулы асРНК пытаются использовать для борьбы с бактериями за счет угнетения ими синтеза в клетке определенных жизненно важных белков.
Пуриновые и пиримидиновые нуклеотиды - это те строительные блоки, из которых синтезируются нуклеиновые кислоты. Кроме того, пуриновые и пиримидиновые нуклеотиды входят в состав многих коферментов и служат для активации и переноса аминокислот, моносахаров, органических кислот.
ДНК выполняет в бактериальной клетке наследственную функцию. Молекула ДНК построена из двух полинуклеотидных цепочек. Каждый нуклеотид состоит из азотистого основания, сахара дезоксирибозы и фосфатной группы (рис. 3.1, б). Азотистые основания представлены пуринами (аденин, гуанин) и пиримидинами (тимин, цитозин). Каждый нуклеотид обладает полярностью. У него имеется дезоксирибозный З'-конец и фосфатный 5'-конец. Нуклеотиды соединяются в полинуклеотидную цепочку посредством фосфодиэфирных связей между 5'-концом одного нуклеотида и З'-концом другого (рис. 3.1, а). Соединение цепей обеспечивается водородными связями между комплементарными
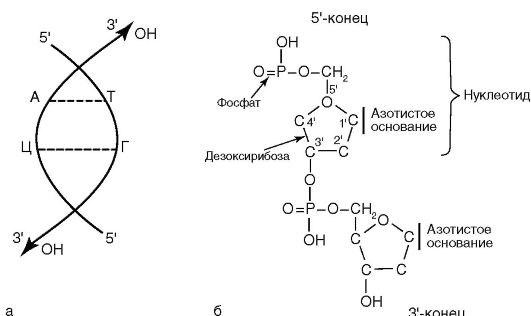 Рис. 3.1. Строение ДНК и ее элементов (объяснение в тексте)
Рис. 3.1. Строение ДНК и ее элементов (объяснение в тексте)
азотистыми основаниями: аденина с тимином, гуанина с цитозином. Нуклеотидные цепи антипараллельны: на каждом из концов линейной молекулы ДНК расположены 5'-конец одной цепи и З'-конец другой цепи. Процентное содержание ГЦ-пар в ДНК определяет степень родства между бактериями и используется при определении таксономического положения бактерий.
Минеральные вещества обнаруживаются в золе, полученной после сжигания клеток. В большом количестве представлены N, S, Р, Са, К, Mg, Fe, Mn, а также микроэлементы Zn, Cu, Co, Ва.
Азот входит в состав белков, нуклеотидов, коферментов. Сера входит в виде сульфгидрильных групп в структуру белков. Фосфор в виде фосфатов представлен в нуклеиновых кислотах, АТФ, коферментах. В качестве активаторов ферментов используются ионы Mg, Fe, Mn. Ионы К и Mg необходимы для активации рибосом. Са является составной частью клеточной стенки грамположительных бактерий. У многих бактерий имеются сидерохромы, которые обеспечивают транспортировку ионов Fe внутрь клетки в виде растворимых комплексных соединений.
Классификация бактерий по типам питания и способам получения энергии
Основной целью метаболизма бактерий является рост, т.е. координированное увеличение всех компонентов клетки. Поскольку основными компонентами бактериальной клетки являются органические соединения, белки, углеводы, нуклеиновые кислоты и липиды, остов которых построен из атомов углерода, то для роста требуется постоянный приток атомов углерода. В зависимости от источника усвояемого углерода бактерии подразделяют на аутотрофы (от греч. autos - сам, trophe - питание), которые используют для построения своих клеток неорганический углерод, в виде СО2, и гетеротрофы (от греч. heteros - другой), которые используют органический углерод. Легкоусвояемыми источниками органического углерода являются гексозы, многоатомные спирты, аминокислоты, липиды.
Белки, жиры, углеводы и нуклеиновые кислоты являются крупными полимерными молекулами, которые синтезируются из мономеров в реакциях поликонденсации, протекающих с поглощением энергии. Поэтому для восполнения своей биомассы бактериям, помимо источника углерода, требуется источник энергии. Энергия запасается бактериальной клеткой в форме молекул АТФ.
Организмы, для которых источником энергии является свет, называются фототрофами. Те организмы, которые получают энергию за счет окислительно-восстановительных реакций, называются хемотрофами.
Среди хемотрофов выделяют литотрофы (от греч. lithos - камень), способные использовать неорганические доноры электронов (Н2, NH3, H2S, Fe2+ и др.) и органотрофы, которые используют в качестве доноров электронов органические соединения.
Бактерии, изучаемые медицинской микробиологией, являются гетерохемоорганотрофами. Отличительной особенностью этой группы является то, что источник углерода у них является источником энергии. Учитывая разнообразие микромира и типов метаболизма, далее изложение материала ограничено рассмотрением метаболизма у гетерохемоорганотрофов.
Степень гетеротрофности у различных бактерий неодинакова. Среди бактерий выделяют сапрофиты (от греч. sapros - гнилой, phyton - растение), которые питаются мертвым органическим материалом и независимы от других организмов, и паразиты (от греч. parasites - нахлебник) - гетеротрофные микроорганизмы, получающие питательные вещества от макроорганизма.
Среди паразитов различают облигатные и факультативные. Облигатные паразиты полностью лишены возможности жить вне клеток макроорганизма. К ним относятся представители родов Rickettsia, Coxiella, Ehrlichia, Chlamydia и др., размножающиеся только внутри клеток макроорганизма. Факультативные паразиты могут жить и без хозяина и размножаться, так же как и сапрофиты, на питательных средах in vitro, т.е. вне организма.
Культивирование бактерий в системах in vitro осуществляется на питательных средах. Искусственные питательные среды должны отвечать следующим требованиям.
• Каждая питательная среда должна содержать воду, так как все процессы жизнедеятельности бактерий протекают в воде.
• Для культивирования гетероорганотрофных бактерий в среде должен содержаться органический источник углерода и энергии. Эту функцию выполняют различные органические соединения: углеводы, аминокислоты, органические кислоты, липиды. Наибольшим энергетическим потенциалом обладает глюкоза, так как она непосредственно подвергается расщеплению с образованием АТФ и ингредиентов для биосинте-
тических путей. Часто используется в этих целях пептон - продукт неполного гидролиза белков, состоящий из поли-, олиго- и дипептидов. Пептон также поставляет аминокислоты для построения бактериальных белков.
• Для синтеза белков, нуклеотидов, АТФ, коферментов бактериям требуются источники азота, серы, фосфаты и другие минеральные вещества, в том числе микроэлементы. Источником азота может служить пептон; кроме того, большинство бактерий способны использовать соли аммония в качестве источника азота. Серу и фосфор бактерии способны утилизировать в виде неорганических солей: сульфатов и фосфатов. Для нормального функционирования ферментов бактериям требуются ионы Са2+, Mg2+, Mn2+, Fe2+, которые добавляют в питательную среду в виде солей, чаще всего фосфатов.
• Решающее значение для роста многих микроорганизмов имеет рН среды. Поддерживание определенного рН имеет значение для предотвращения гибели микроорганизмов от ими же образованных продуктов обмена.
• Среда должна обладать определенным осмотическим давлением. Большинство бактерий способны расти на изотоничных средах, изотоничность которых достигается добавлением NaCl в концентрации 0,87%. Некоторые бактерии не способны расти на средах при концентрации соли в них ниже 1%. Такие бактерии называются галофильными. Так как устойчивость к осмотическому давлению определяется наличием у бактерий клеточной стенки, бактерии, лишенные клеточной стенки, микоплазмы L-формы, могут расти на питательных средах, содержащих гипертонический раствор, обычно сахарозы. При необходимости к питательной среде добавляют факторы роста, ингибиторы роста определенных бактерий, субстраты для действия ферментов, индикаторы.
• Питательные среды должны быть стерильными.
В зависимости от консистенции питательные среды могут быть жидкими, полужидкими и плотными. Плотность среды достигается добавлением агара.
Агар - полисахарид, получаемый из водорослей. Он плавится при температуре 100 °С, но при охлаждении остывает при температуре 45-50 °С. Агар добавляют в концентрации 0,5% для полужидких сред и 1,5-2% для создания плотных сред. В зависимо-
сти от состава и цели применения различают простые, сложные, элективные, минимальные, дифференциально-диагностические и комбинированные среды.
По составу питательные среды могут быть простыми и сложными. К простым средам относятся пептонная вода, питательный бульон, мясопептонный агар. На основе простых сред готовят сложные среды, например сахарный и сывороточный бульоны, кровяной агар.
В зависимости от назначения среды подразделяются на элективные, обогащения, дифференциально-диагностические. Под элективными понимают среды, на которых лучше растет какой-то определенный микроорганизм. Например, щелочной агар, имеющий рН 9,0, служит для выделения холерного вибриона. Другие бактерии, в частности кишечная палочка, из-за высокого рН на этой среде не растут.
Среды обогащения - это среды, которые стимулируют рост какого-то определенного микроорганизма, ингибируя рост других. Например, среда, содержащая селенит натрия, стимулирует рост бактерий рода Salmonella, ингибируя рост кишечной палочки.
Дифференциально-диагностические среды служат для изучения ферментативной активности бактерий. Они состоят из простой питательной среды с добавлением субстрата, на который должен подействовать фермент, и индикатора, меняющего свой цвет в результате ферментативного превращения субстрата. Примером таких сред являются среды Гисса, используемые для изучения способности бактерий ферментировать сахара.
Комбинированные питательные среды сочетают в себе элективную среду, подавляющую рост сопутствующей флоры, и дифференциальную среду, диагностирующую ферментативную активность выделяемого микроба. Примером таких сред служат среда Плоскирева и висмут-сульфитный агар, используемые при выделении патогенных кишечных бактерий. Обе эти среды ингибируют рост кишечной палочки.
3.1.2. Ферменты бактерий
В основе всех метаболических реакций в бактериальной клетке лежит деятельность ферментов, которые принадлежат к 6 классам: оксиредуктазы, трансферазы, гидролазы, лигазы, лиазы, изомеразы. Ферменты, образуемые бактериальной клеткой, могут как
локализоваться внутри клетки - эндоферменты, так и выделяться в окружающую среду - экзоферменты. Экзоферменты играют большую роль в обеспечении бактериальной клетки доступными для проникновения внутрь источниками углерода и энергии. Большинство гидролаз являются экзоферментами, которые, выделяясь в окружающую среду, расщепляют крупные молекулы пептидов, полисахаридов, липидов до мономеров и димеров, способных проникнуть внутрь клетки. Ряд экзоферментов, например гиалуронидаза, коллагеназа, являются ферментами агрессии. Некоторые ферменты локализованы в периплазматическом пространстве бактериальной клетки. Они участвуют в процессах переноса веществ в бактериальную клетку. Ферментативный спектр является таксономическим признаком, характерным для семейства, рода и в некоторых случаях для видов. Поэтому определением спектра ферментативной активности пользуются при установлении таксономического положения бактерий. Наличие экзоферментов можно определить при помощи дифференциально-диагностических сред. Для идентификации бактерий разработаны специальные тест-системы, состоящие из набора дифференциально-диагностических сред.
3.1.3. Энергетический метаболизм
Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных бактерий реакции, связанные с получением энергии в форме АТФ, - это реакции окисления- восстановления, сопряженные с реакциями фосфорилирования. Окисленный в этих реакциях углерод выделяется клеткой в виде СО2. Для удаления отщепившегося в этих реакциях водорода, который находится в форме восстановленного НАД, разные бактерии используют различные возможности в зависимости от конечного акцептора водорода (или электронов, что является эквивалентным понятием). В зависимости от способа получения энергии у бактерий имеется несколько типов метаболизма: окислительный, или дыхание; бродильный, или ферментативный; смешанный. Тип метаболизма определяет не только реакции, в результате которых образуется АТФ, но и конечные продукты этих реакций, которые используются при идентификации бактерий, а также условия культивирования бактерий.
При использовании в качестве источника углерода и энергии глюкозы или других гексоз начальные этапы окисления глюкозы
являются общими, как при оксидативном, так и при бродильном метаболизме. К ним относятся пути превращения глюкозы в пируват (при использовании в качестве источника энергии отличных от глюкозы гексоз, или дисахаридов, они в результате химических превращений вступают в цепь реакций, превращающих глюкозу в пируват). Пируват, образовавшийся при расщеплении глюкозы, превращается при участии кофакторов в активированную уксусную кислоту или ацетилкоэнзим А. Последний окисляется в СО2 с отщеплением водорода в цикле трикарбоновых кислот.
Цикл трикарбоновых кислот не только выполняет функцию конечного окисления питательных веществ, но и обеспечивает процессы биосинтеза многочисленными предшественниками: пируват α-кетоглутаровая, щавелевая и янтарные кислоты - для синтеза аминокислот, щавелевоуксусная - для синтеза пиримидиновых нуклеотидов, малонат - для синтеза аминокислот, пиримидиновых нуклеотидов и жиров (рис. 3.2).
Окислительный метаболизм. Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания. Дыхание - процесс получения энергии в реакциях окисления-восстановления, сопряженных с реакциями окислительного фосфорилирования, при котором донорами электронов могут быть органические (у ор-
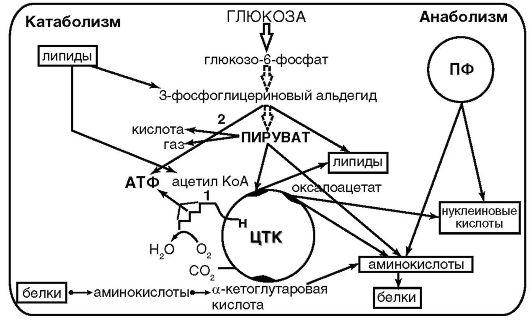 Рис. 3.2. Схема обмена веществ у бактерий
Рис. 3.2. Схема обмена веществ у бактерий
ганотрофов) и неорганические (у литотрофов) соединения, а акцептором - только неорганические соединения.
У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода [Н+]) является молекулярный кислород. В этом случае пируват полностью окисляется в цикле трикарбоновых кислот до СО2. Цикл трикарбоновых кислот выполняет функции поставщика как предшественников для биосинтетических процессов, так и атомов водорода, который в форме восстановленного НАД переносится на молекулярный кислород через серию переносчиков, обладающих сложной структурно оформленной мультиферментной системой - дыхательной цепью. Дыхательная цепь у бактерий локализована в ЦПМ и во внутриклеточных мембранных структурах.
Электрохимическую энергию бактерии получают в процессе переноса электронов по окислительно-восстановительным цепям в мембране, в результате чего происходит неравномерное распределение Н+ по обеим ее сторонам. Переносчики электронов располагаются в мембране таким образом, что во внешней среде происходит накопление ионов водорода (при этом возникает подкисление среды), а в цитоплазме их число уменьшается, что сопровождается подщелачиванием среды. Неравномерное распределение положительно заряженных протонов (большее число на наружной и меньшее на внутренней поверхности плазматической мембраны) приводит к формированию расположенного поперек мембраны электрического поля, мембранного потенциала. В результате при переносе электронов возникает трансмембранный электрохимический градиент ионов водорода, обозначаемый символом ΔμΗ+ и измеряемый в вольтах. Энергия мембранного потенциала используется для синтеза локализованной в мембране АТФазой АТФ.
Энергия в форме ΔμΗ+ не теряется при ее запасании и может образовываться и потребляться клеткой в условиях, когда невозможен синтез АТФ. В последние годы показано, что аналогичным образом перераспределяются и атомы Na+ с образованием энергии, обозначаемой как ΔμNa+. Данные формы энергии тратятся преимущественно на движение бактерий (у подвижных форм) и транспорт веществ в клетку и из нее.
Типичная цепь выглядит следующим образом: ЦТК → НАД(Н2) → флавопротеид → хинон → цитохромы: b c a → О2.
Конечным этапом переноса электронов (протонов) по дыхательной цепи является восстановление цитохромов а + а3 (цитохромоксидазы). Цитохромоксидаза является конечной оксидазой, передающей электроны на кислород. Образующиеся при окислении ФАД или хинонов протоны связываются ионами О2- с образованием воды.
В то время как у эукариотов ферменты дыхательной цепи имеют относительно постоянный состав, у бактерий встречаются вариации в составе дыхательной цепи. У некоторых бактерий цитохромы отсутствуют и при контакте с кислородом происходит непосредственный перенос водорода на кислород с помощью флавопротеидов, конечным продуктом при этом оказывается перекись водорода (Н2О2).
Помимо углеводов, прокариоты способны использовать другие органические соединения, в частности белки, в качестве источника энергии, окисляя их полностью до СО2 и Н2О.
Аминокислоты и белки также могут выступать в качестве энергетических ресурсов. Их использование связано в первую очередь с определенными ферментативными преобразованиями подготовительного характера. Белки вначале вне клетки расщепляются протеолитическими ферментами на пептиды, которые поглощаются клеткой и расщепляются внутриклеточными пептидазами до аминокислот. Аминокислоты могут использоваться в конструктивном метаболизме, а могут у аммонифицирующих бактерий служить основным материалом в энергетических процессах при окислительном дезаминировании, в результате которого происходят выделение аммиака и превращение аминокислоты в кетокислоту, которая через цикл трикарбоновых кислот вступает в конструктивный метаболизм.
Процесс аммонификации известен как гниение, при этом происходит накопление продуктов, обладающих неприятным специфическим запахом образующихся при этом первичных аминов. Гнилостные бактерии осуществляют минерализацию белка, разлагая его до СО2, NH3, H2S. К гнилостным бактериям относятся Proteus, Pseudomonas, Bacillus cereus.
Анаэробное дыхание. Некоторые бактерии обладают способностью использовать в анаэробных условиях нитрат как конечный акцептор водорода. Восстановление нитрата может происходить двумя путями: аммонификацией, при которой нитрат превращает-
ся в аммиак, и денитрофикацией, при которой происходит восстановление нитрата до молекулярного азота или закиси азота. Этот процесс связан с деятельностью фермента нитратредуктазы.
Сульфатное дыхание. Использовать сульфат как конечный акцептор водорода при анаэробном дыхании способна лишь небольшая группа бактерий, включающая только два рода: Desulfovibrio, Desulfotomaculum. Эти бактерии являются строгими анаэробами, они обитают в сероводородном иле и не имеют значения в медицинской микробиологии. Они способны использовать в качестве донора электронов молекулярный водород, поэтому их относят к хемолитотрофам. Этим бактериям принадлежит ведущая роль в образовании сероводорода в природе.
Бродильный (ферментативный) метаболизм. Ферментация, или брожение, - процесс получения энергии, при котором отщепленный от субстрата водород переносится на органические соединения. Кислород в процессе брожения участия не принимает. Восстановленные органические соединения выделяются в питательную среду и накапливаются в ней. Ферментироваться могут углеводы, аминокислоты (за исключением ароматических), пурины, пиримидины, многоатомные спирты. Не способны сбраживаться ароматические углеводороды, стероиды, каротиноиды, жирные кислоты. Эти вещества разлагаются и окисляются только в присутствии кислорода, в анаэробных условиях они стабильны. Продуктами брожения являются кислоты, газы, спирты.
При ферментации гексоз (глюкозы) пируват лишь частично окисляется в цикле трикарбоновых кислот. Последний выполняет только функции поставщика предшественников для биосинтетических процессов. Энергия в форме двух молекул АТФ образуется в результате субстратного фосфорилирования, протекающего при окислении триозофосфата в пируват. Отщепившийся от субстрата водород, находящийся в форме восстановленного НАД, переносится на пируват, превращая его в цепи реакций в этанол, кислоты, газы. Исходя из природы конечных продуктов, различают несколько типов брожения углеводов: спиртовое, молочнокислое, муравьинокислое, маслянокислое.
Спиртовое брожение встречается в основном у дрожжей. Конечными продуктами являются этанол и СО2. Спиртовое брожение используется в хлебопекарной промышленности и виноделии.
Молочнокислое брожение происходит у S. pyogenes, E. faecalis, S. Salivarius, а также у бактерий родов Lactobacillus и Bifidobacterium. Продуктами этого типа брожения являются молочная кислота, этанол и уксусная кислота. Продукты молочнокислого брожения играют большую роль в формировании колонизационной резистентности бактериями рода Lactobacillus и Bifidobacterium, составляющих облигатную флору кишечника. Молочнокислые бактерии широко используются в молочной промышленности для получения молочнокислых продуктов, а также в создании пробиотиков.
Муравьинокислое (смешанное) брожение встречается у представителей семейств Enterobacteriaceae и Vibrionaceae. Различают два типа этого брожения. При первом происходит расщепление пирувата с образованием через цепь реакций муравьиной, янтарной и молочной кислот. Сильное кислотообразование можно выявить реакцией с индикатором метиленовым красным, который меняет окраску в сильнокислой среде. При втором типе брожения образуется целый ряд кислот, однако главным продуктом брожения являются ацетоин и 2,3-бутандиол, образующиеся через цепь реакций из двух молекул пирувата. Эти вещества при взаимодействии с α-нафтолом в щелочной среде вызывают образование окраски бурого цвета, что выявляется реакцией Фогеса-Проскауэра, используемой при идентификации бактерий.
Маслянокислое брожение. Масляная кислота, бутанол, ацетон, изопропанол и ряд других органических кислот, в частности уксусная, капроновая, валериановая, пальмитиновая, являются продуктами сбраживания углеводов сахаролитическими строгими анаэробами. Спектр этих кислот, определяемый при помощи газожидкостной хроматографии, используется как экспресс-метод при идентификации анаэробов.
Ферментация белков. Если для бактерий с бродильным метаболизмом источником энергии служат белки, то такие бактерии называются пептолитическими. Пептолитическими являются некоторые клостридии, в частности С. histolyticum и C. botulinum. Пептолитические бактерии гидролизуют белки и сбраживают аминокислоты. Многие аминокислоты сбраживаются совместно с другими, при этом одна выполняет функцию донора, а другая - функцию акцептора водорода. Аминокислота-донор дезаминируется в кетокислоту, которая в результате окислительного декарбоксилирования превращается в жирную кислоту.
3.1.4. Конструктивный метаболизм
Основные органические компоненты бактериальной клетки, как уже было отмечено, синтезируются в реакциях полимеризации из строительных блоков: аминокислот, фосфатов, сахаров, пуриновых и пиримидиновых оснований, органических кислот. Поставщиками этих строительных блоков являются промежуточные продукты основных путей энергетического метаболизма (см. рис. 3.2). Среди бактерий выделяется группа, называющаяся прототрофами, которые способны синтезировать все компоненты клетки из одного источника углерода и энергии. Если бактерии теряют способность образовывать какое-либо жизненно важное вещество (аминокислоту, витамин, фактор роста и др.), участвующее в биосинтетических процессах, то для их роста и размножения требуется его поступление в готовом виде. Такие вещества называют фактором роста, а бактерии, возникшие, как правило, в результате мутаций, - ауксотрофами.
Факторами роста являются аминокислоты, пуриновые и пиримидиновые основания, витамины, которые входят в состав простетических групп коферментов.
Биосинтез аминокислот и синтез белка. Большинство бактерий обладают способностью синтезировать все 20 аминокислот, из которых состоят белки. Белки в бактериальной клетке выполняют ферментативную функцию, а также являются составной частью структурных образований клетки: ЦПМ и ее производных, клеточной стенки, жгутиков, капсулы и спор у некоторых бактерий.
Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат промежуточные продукты фруктозодифосфатного (ФДФ) и пентозофосфатного (ПФ) путей, цикл трикарбоновых кислот: пируват, кетоглутаровая кислота, оксалоацетат, фумарат, эритрозо-4-фосфат, рибозо- 4-фосфат. Аминогруппы вводятся в результате непосредственного аминирования или переаминирования. Перевод неорганического азота в органические соединения происходит всегда через аммиак. Нитраты и нитриты и молекулярный азот предварительно восстанавливаются в аммиак и только лишь после этого включаются в состав органических соединений. В результате прямого аминирования образуются лишь L-аланин, L-аспартат, L-глутамат и L-глугамин. Все остальные аминокислоты получают свою аминогруппу в результате переаминирования с одной из первичных
аминокислот. Синтез белка осуществляется у бактерий так же, как в клетках эукариот.
Синтез белка происходит на рибосомах и обычно подразделяется на три процесса: инициацию, элонгацию и терминацию. Инициация синтеза белка заключается в связывании мет-тРНК с малой субъединицей рибосомы с последующим встраиванием инициирующего кодона иРНК. Элонгация происходит за счет поочередного присоединения аминокислотных остатков к растущей полипептидной цепи. Терминация наступает, когда синтез полипептида достигает стоп-кодона. У E. coli известны три таких кодона: УАА, УГА и УАГ. В результате действия факторов терминации происходят остановка синтеза белка и диссоциация молекулы иРНК и рибосомы. Скорость синтеза белка в микробной клетке очень велика, так как ДНК бактерий не отграничена мембраной от цитоплазмы, не содержит интронов (участков ДНК, не несущих информации) и соответственно у микробов отсутствуют вырезание их копий из иРНК и сшивание копий экзонов (участков, кодирующих белки). В результате в клетках прокариот не происходит физического разделения процессов синтеза иРНК (транскрипции) и трансляции, поэтому оба процесса часто идут одновременно: трансляция начинается раньше, чем завершена транскрипция. Бактерии также способны одновременно синтезировать несколько идентичных молекул на одной матрице иРНК. При этом иРНК связывается с несколькими рибосомами с образованием комплекса, получившего название «полисомы».
Процесс синтеза белка представляет собой важную мишень для разнообразных антимикробных препаратов. При этом антибиотики имеют различные мишени и механизмы действия, например аминогликозиды и тетрациклины соединяются с малой, а макролиды и линкозамиды - с большой субъединицей рибосом. Белки, синтезируемые клеткой, могут использоваться внутри нее или выделяться в окружающую среду или периплазматическое пространство (грамотрицательные бактерии).
Многие годы считали, что после синтеза на рибосомах молекулы белков сами приобретают нужную форму (третичную структуру). Сейчас мы знаем, что большинство из них приобретают нужную конформацию молекулы с помощью специальных белков, получивших название шаперонов. Молекулы шаперонов не только обеспечивают правильное складывание белков, но и препятствуют неправильному их закручиванию. Эти молекулы абсолютно не-
обходимы для поддержания нормальной жизнедеятельности клетки как эу-, так и прокариот. Процесс укладки белковых молекул является энергетически зависимым и сопровождается расходом энергии АТФ. Количество шаперонов резко возрастает, когда клетка подвергается стрессорному воздействию различных факторов внешней среды (температуры - тепловой шок, токсинов, нарушающих метаболические реакции и др.) При этом шапероны защищают многие белки, включая ДНК-полимеразы от разрушения. Еще одной важной функцией шаперонов является их участие в транспорте белков через мембраны.
Биосинтез нуклеотидов. Пуриновые и пиримидиновые нуклеотиды - это те строительные блоки, из которых синтезируются нуклеиновые кислоты. Кроме того, пуриновые и пиримидиновые нуклеотиды входят в состав многих коферментов и служат для активации и переноса аминокислот, сахаров, липидов в реакциях полимеризации. Исходным соединением для образования пентозной части нуклеотидов служит рибозо-5-фосфат, образующийся в ПФпути. Углеродный скелет пиримидинов происходит из аспартата, который образуется в цикле трикарбоновых кислот. Атомы азота и аминогруппы пуринов и аминосодержащих пиримидинов происходят из аспартата и глутамина.
Ключевым промежуточным продуктом для биосинтеза жирных кислот является ацетилкоэнзим А. Ключевыми промежуточными продуктами для синтеза фосфолипидов является продукт ФДФпути - диоксиацетилфосфат, восстанавливающийся в глицерол-3- фосфат, который соединяется с остатками жирных кислот.
Биосинтез углеводов. Углеводы представлены в бактериальной клетке в виде моно-, ди- и полисахаридов, а также комплексных соединений.
Синтез глюкозы происходит из пирувата за счет обратных реакций, путей распада глюкозы. Для обхода реакций, идущих только в одном направлении, имеются обходные пути, например глиоксилатный цикл.
3.1.5. Транспорт веществ
Транспорт веществ в бактериальную клетку
Для того чтобы питательные вещества могли подвергнуться превращениям в цитоплазме клетки, они должны проникнуть в клетку через пограничные слои, отделяющие клетку от окружаю-
щей среды. Ответственность за поступление в клетку питательных веществ лежит на ЦПМ.
Существует два типа переноса веществ в бактериальную клетку: пассивный и активный.
При пассивном переносе вещество проникает в клетку только по градиенту концентрации. Затрат энергии при этом не происходит. Различают две разновидности пассивного переноса: простую диффузию и облегченную диффузию. Простая диффузия - неспецифическое проникновение веществ в клетку, при этом решающее значение имеют величина молекул и липофильность. Скорость переноса незначительна. Облегченная диффузия протекает с участием белка-переносчика транслоказы. Скорость этого способа переноса зависит от концентрации вещества в наружном слое.
При активном переносе вещество проникает в клетку против градиента концентрации при помощи белка-переносчика пермеазы. При этом происходит затрата энергии. Имеется два типа активного транспорта. При первом типе активного транспорта небольшие молекулы (аминокислоты, некоторые сахара) накачиваются в клетку и создают концентрацию, которая может в 100-1000 раз превышать концентрацию этого вещества снаружи клетки. Второй механизм, получивший название «транслокация радикалов» (фосфотрансферазный путь), обеспечивает включение в клетку некоторых сахаров (например, глюкозы, фруктозы), которые в процессе переноса фосфорилируются, т.е. химически модифицируются. Для осуществления этих процессов в бактериальной клетке локализуется специальная фосфотрансферная система, составной частью которой является белок-переносчик, находящийся в активной фосфорилированной форме. Фосфорилированный белок связывает свободный сахар на наружной поверхности мембраны и транспортирует его в цитоплазму, где сахар освобождается в виде фосфата. Поступив в клетку, органический источник углерода и энергии вступает в цепь биохимических реакций, в результате которых образуются АТФ и ингредиенты для биосинтетических процессов. Биосинтетические (конструктивные) и энергетические процессы протекают в клетке одновременно. Они тесно связаны между собой через общие промежуточные продукты, которые называются амфиболитами.
Транспорт веществ из бактериальной клетки
В процессе жизнедеятельности бактериям требуется выделять в окружающую среду различные белки и ферменты. Секретируемые
белки необходимы для жизнедеятельности бактерий. Они участвуют в построении клеточных оболочек, жгутиков, пилей, расщепляют крупные полимерные молекулы, используемые в качестве питательных веществ, до размеров, способных проходить через бактериальную ЦПМ; осуществляют взаимодействие с системами макроорганизма. У грамположительных микробов белки секретируются непосредственно во внешнюю среду. А у грамотрицательных бактерий они должны пересечь наружную мембрану. Наличие наружной мембраны привело к формированию у грамотрицательных бактерий различных по структуре и функциям систем секреции 5 типов.
Белки, секретируемые по I и III путям, пересекают внутреннюю цитоплазматическую и наружные мембраны в один этап без участия sec-белков, тогда как белки, секретируемые по II и IV путям, проходят через внутреннюю и наружную мембрану отдельными этапами при участии sec-белков. Sec-белки (транслоказы) являются небольшими белками в 30 аминокислот, которые способны узнавать сигнальную последовательность, расположенную на N-терминальном конце секретируемого белка, и связываться с ней сразу же после завершения процесса трансляции, предотвращая включение секретируемого белка в метаболизм клетки. В процессе транслокации белка, которая сопровождается поглощением энергии, происходит отщепление пептидазой в периплазматическом пространстве сигнальной последовательности, а в результате взаимодействия с шаперонами происходит формирование четвертичной структуры переносимого белка. Зрелый белок проходит через пору в наружной мембране в окружающую среду. II тип секреции является основным путем для секреции экстрацеллюлярных гидролитических ферментов у грамотрицательных бактерий. У V. cholerae по этому пути выделяются холерный токсин, нейраминидаза, гемагглютининпротеаза, а у P. aeruginosa - эластаза, фосфолипаза С.
V тип секреции отличается от II типа тем, что в периплазматическом пространстве из С-терминальной части секретируемого полипептида формируется β-цилиндрическая структура, выполняющая роль поры, через которую проходит N-терминальный конец. Внеклеточный протеолиз приводит секретируемый белок в активное функциональное состояние. По этому пути секретируются IgA-протеаза у N. gonorrhoeae, белок пертактин у B. pertussis.
I тип секреции протекает одноэтапно и требует наличия трех секреторных белков: белка, формирующего в цитоплазматической мембране пору АТФазы; белка ЦПМ, пронизывающего периплазматическое пространство; белка клеточной стенки, формирующего пору.
Транспортные белки узнают секретируемый белок по наличию сигнальной последовательности, расположенной на С-терминальном конце белка. Данным путем секретируются в основном пороформирующие токсины: гемолизин, металлопротеаза, а также внеклеточная аденилатциклаза у B. pertussis (рис. 3.3).
Особый интерес для медицинской микробиологии представляет III путь секреции, возникший эволюционно у грамотрицательных бактерий для транспорта из клетки компонентов жгутиков. Показано, что он используется также для направленной доставки в клетку эукариот бактериальных белков. В результате действия последних в клетке возникают различные нарушения функций, приводящие в конечном счете к возникновению у человека заболеваний. В процесс выделения молекул из клетки данным способом вовлечено более 20 различных белков. Секреторная система третьего типа (ТТСС) представляет шприцеподобную структуру (рис. 3.4), способную инъецировать эффекторные молекулы непосредственно в цитозоль клетки-хозяина. Белки ТТСС можно разделить на три группы: белки, формирующие «шприц» ТТСС; белки транслокационного комплекса, обеспечивающие транслокацию эффекторных молекул в цитоплазму клеток хозяина; эффекторные белки,
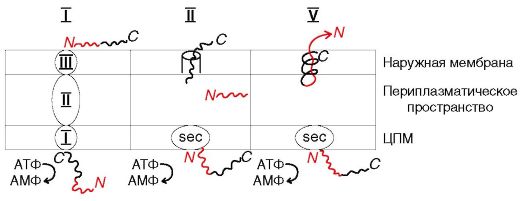 Рис. 3.3. Схема
строения секреторных систем: I тип обеспечивает секрецию формирующих
поры токсинов (гемолизина); II тип обеспечивает секрецию гидролитических
ферментов, некоторых токсинов (энтеротоксин V. cholerae), поверхностных структур (пили 4 типа); V тип обеспечивает аутотранспорт белков у грамотрицательных бактерий
Рис. 3.3. Схема
строения секреторных систем: I тип обеспечивает секрецию формирующих
поры токсинов (гемолизина); II тип обеспечивает секрецию гидролитических
ферментов, некоторых токсинов (энтеротоксин V. cholerae), поверхностных структур (пили 4 типа); V тип обеспечивает аутотранспорт белков у грамотрицательных бактерий
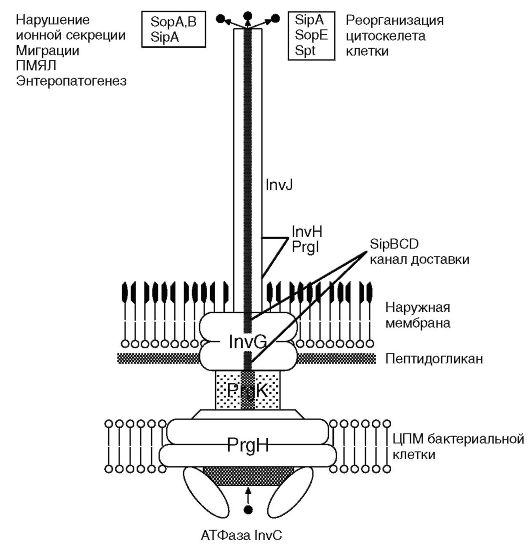 Рис. 3.4. Схема строения секреторной системы III типа у Salmonella: PrgH, PrgK, InvG,H,J; PrgI - белки, формирующие шприц; Sip B, C, D - белки транслокационного комплекса; SopA, B, E SipA; Spt - эффекторные белки
Рис. 3.4. Схема строения секреторной системы III типа у Salmonella: PrgH, PrgK, InvG,H,J; PrgI - белки, формирующие шприц; Sip B, C, D - белки транслокационного комплекса; SopA, B, E SipA; Spt - эффекторные белки
которые непосредственно оказывают модулирующее действие на клетку-хозяина.
Белки транслокационных комплексов формируют поры в мембране клетки-хозяина, создавая канал для доставки эффекторных молекул. Транслокация через сформировавшийся канал в клетку-хозяина эффекторных молекул происходит при участии белков-шоперонов.
Эффекторные белки вызывают реорганизацию цитоскелета клетки-хозяина, что способствует проникновению бактерии в клетку-хозяина, а также вызывают различные нарушения функций
клеток хозяина, приводящие в конечном счете к возникновению патологического процесса.
Экспрессия генов ТТСС регулируется различными транскрипционными регуляторами, которые интегрируют сигналы окружающей среды, такие, как осмомолярность, концентрация кислорода, рН, температура, концентрация ионов Са. ТТСС имеется у Salmonella, Shigella, Yersinia, P. aeruginosa, Chlamydia, некоторых патогенных E. coli.
Выделение молекул из клетки также осуществляется с помощью белков-переносчиков и фосфотрансферазным путем. Одним из вариантов переносчиков можно считать белковые помпы, обеспечивающие выведение из клеток ряда антимикробных препаратов, например тетрациклинов. Фосфотрансферазный путь широко используется при выведении молекул, необходимых для построения различных структур бактерий, расположенных кнаружи от плазматической мембраны, в частности клеточной стенки, капсулы и др. Некоторые из стадий подобного транспорта подавляются используемыми в практике антимикробными препаратами, например транспорт через мембрану N-ацетилглюкозамина блокируется гликопептидным антибиотиком ванкомицином. Особым типом транспорта веществ из клеток бактерий является недавно открытая секреция мембранных пузырьков. Хотя механизм их выделения остается не совсем ясным, показано, что они могут содержать липиды, белки, нуклеиновые кислоты и полисахариды, в том числе некоторые бактериальные токсины.
3.1.6. Регуляция метаболизма у бактерий
Регуляция функций бактерий может приводить к появлению или исчезновению активности, а также изменению ее уровня. Эффективность клеточных регуляторных механизмов очень велика и обеспечивает максимально экономичное использование питательных веществ, предупреждают избыточный синтез промежуточных и конечных метаболитов и способствует адаптации к изменяющимся условиям окружающей среды. Регуляторные процессы в клетке происходят на разных уровнях и начинаются на уровне генома. Изменение числа копий определенного гена приводит к повышению синтеза закодированного в нем белка.
Регуляция на уровне транскрипции сводится к тому, что ряд генов бактерий транскрибируется только в определенных усло-
виях. Регуляция осуществляется специальными транскрипционными факторами и компонентами глобальной регуляторной сети. Под ее контролем находятся ответы микробной клетки на изменения температуры и рН среды, наличие питательных веществ и двухвалентных катионов (особенно Са2+), фаза роста культуры. На уровне транскрипции регулируются III тип транспорта веществ из бактерий в клетку хозяина и процессы, обеспечивающие поддержание жизнедеятельности клетки в экстремальных для нее условиях. К последним относятся системы холодового и теплового шока, адаптационная система, индуцибельная система репарации ДНК (SOS-ответ) и др. Синтез белка этих систем начинается, когда клетка попадает в неблагоприятные условия и образование молекул иРНК других белков замедляется. Количество одного и того же фермента у бактерий в разных условиях может изменяться от 1-2 молекул на клетку до нескольких процентов от ее массы. Известны некоторые механизмы регуляции на уровне транскрипции, связанные с регуляторными генами. Различают негативную и позитивную регуляцию. При позитивной регуляции индуктор взаимодействует с репрессором и освобождает ген-оператор. При негативной регуляции происходит блокирование гена-оператора белком-репрессором.
В регуляции различных свойств бактерий в ответ на изменения условий окружающей среды принимают участие так называемые двухкомпонентные системы передачи сигнала. Основными компонентами системы проведения сигнала являются сенсор, принимающий сигнал, которым обычно является трансмембранная протеинкиназа, изменяющая свою активность под влиянием фактора окружающей среды, и расположенный в цитозоле регулятор, который она активирует и который, в свою очередь, влияет на экспрессию соответствующих оперонов.
Регуляция на уровне ферментов осуществляется регуляцией интенсивности ферментативных реакций. Скорость последних может регулироваться двумя основными способами: путем изменения количества ферментов и путем изменения их активности. Биосинтетические пути, опосредованные конститутивными ферментами, регулируются аллостерическим ингибированием активности первого фермента. Биосинтетические пути, опосредованные индуцибельными ферментами, регулируются путем репрессии их синтеза конечным продуктом.
Катаболические пути, опосредованные индуцибельными ферментами, регулируются индукцией синтеза ферментов и катаболической репрессией, а опосредованные конститутивными ферментами - посредством аллостерических воздействий на их активность. АТФ в этом случае является отрицательным эффектором, а аденозиндифосфат (АДФ) - положительным эффектором.
3.1.7. Морфогенез бактерий и их сообществ
Построение микробной клетки представляет собой сложный процесс, включающий не только синтез компонентов и деление. Для формирования нормальной клетки необходимо также образование всех структур, расположенных кнаружи от плазматической мембраны, т.е. клеточной стенки, жгутиков, ресничек капсулы и т.п. Морфогенез начинается с синтеза молекул-предшественниц в цитоплазме клеток. Эти молекулы представляют собой компоненты будущих внешних структур. Синтезированные молекулы различными путями переносятся через плазматическую мембрану. Компоненты пептидогликана, например, переносятся за счет фосфотрансферазного пути, некоторые белки - по II типу секреции. Компоненты жгутиков переносятся за счет специальных белков - транслоказ, являющихся компонентами III типа вывода белков из клетки. На поверхности мембраны перенесенные молекулы собираются в блоки, которые, в свою очередь, транспортируются к конечному месту своего расположения и формируют ту или иную внешнюю структуру.
Все этапы морфогенеза контролируются специфическими транспортными и связующими белками, регулируются различными внешними и внутренними факторами и являются мишенями действия ряда антимикробных препаратов. Кроме компонентов клетки, во внешнюю среду также выносятся органические молекулы, предназначенные для образования внеклеточного матрикса и поверхностной пленки, отделяющей сообщества от внешней среды. Многие стадии морфогенеза регулируются за счет активности связанных с мембраной ферментов, работа которых, в свою очередь, зависит от различных факторов, например температуры. Так, при 30 °С у мезофильных возбудителей болезней человека не образуются полноценные капсулы и капсулоподобные оболочки, реснички, функционально активные молекулы токсинов. Последние могут накапливаться в клетке или периплазме в виде
гигантских молекул - протоксинов, у которых не происходит разрезания (ограниченный протеолиз), делающего их функционально активными. Синтез самих молекул-предшественников при этом может не изменяться. Интересно, что после возвращения микроба к оптимальной температуре нормальная структура всех компонентов клетки восстанавливается обычно уже через 2-3 ч и микроб вновь становится вирулентным.
3.1.8. Вторичный метаболизм
Хотя бактерии и клетки животных имеют в большинстве своем сходные пути промежуточного метаболизма, ряду бактерий свойственны дополнительные реакции, в ходе которых синтезируются различные уникальные соединения. Совокупность подобных реакций получила название «вторичный метаболизм», а полученные в результате вещества - вторичные метаболиты. Наиболее характерными примерами вторичных метаболитов являются антибиотики, синтезируемые представителями ограниченного числа родов бактерий, включающего Bacillus, Streptomyces и Nocardia.
3.1.9. Отношение к факторам окружающей среды
Отношение к температуре
Влияние температуры на бактерии в медицинской микробиологии имеет два основных результата: возможность размножаться и сохранение жизнеспособности. В последнем случае речь идет о возможности восстановления способности к росту и размножению после пребывания при экстремальных температурах (повышенных или пониженных) для данного вида. Температурные условия на Земле различаются в широком диапазоне от 90 до 2500 °С, и всюду встречаются микробы, приспособившиеся к ним. Бактерии, вызывающие болезни людей, максимально адаптированы к температуре тела человека. В то же время некоторые из них могут жить и размножаться в окружающей среде (воде, почве, организмах различных животных), в связи с чем оптимальная температура для их роста может быть ниже или выше 37 °С.
По отношению к температуре роста бактерии принято разделять на три основные группы: психрофилы, мезофилы и термофилы. Психрофилы живут и размножаются при пониженных температурах (диапозон температур роста от +10 до -20 °С). При этом
строгие (облигатные) психрофилы неспособны размножаться при температуре выше 20 °С, а факультативные имеют оптимум роста от 22 до 30 °С. Именно в группе факультативных психрофилов обнаружены возбудители болезней человека (например, возбудитель чумы - Yersinia pestis, иерсиниоза - Yersinia enterocolitica, гнойно-воспалительных процессов - Aeromonas spp.). Бактерии, растущие при низких температурах, содержат повышенное количество ненасыщенных жирных кислот и имеют ряд особенностей структуры ферментных белков, что позволяет им расти при низких температурах. Мезофилы включают бактерии, температурный диапазон роста которых находится между 10 и 45 °С, а диапазон оптимальных температур роста лежит между 30 и 40 °С. Именно к этой группе относится большинство возбудителей болезней человека, оптимальный рост которых возможен при 37 °С. Термофилы представляют группу микробов, способных расти при повышенных температурах. Различают термотолерантные формы, для которых оптимальной температурой роста являются 37 °С, но возможность роста, в отличие от мезофилов, сохраняется до 60 °С. Факультативные термофилы проявляют максимальный рост при 50-60 °С, но также растут при 20-40 °С, в то время как облигатные термофилы не могут расти при температуре ниже 40 °С; оптимальная температура для их размножения 70 °С. Известны также экстремальные термофилы, размножающиеся при температуре выше 80 °С. Ряд микробов, являющихся экстремальными термофилами, относятся к доминиону Археи. В строении термофилов одним из важнейших факторов, обеспечивающих термоустойчивость, является структура их белков. Хотя среди термофилов пока не найдены возбудители болезней человека, продукты их жизнедеятельности используются в медицинской промышленности (протеазы, нанесенные на перевязочный материал, для инфицированных ран) и при изготовлении моющих средств (ферменты-биодобавки для очистки тканей от органических молекул).
Сохранение жизнеспособности бактерий при различных температурах зависит от строения клеток. Наиболее устойчивыми следует считать споры, выживающие в широком диапазоне температур - от минусовых до температуры кипящей воды. Вегетативные формы бактерий для выживания в условиях, несколько отличающихся от оптимальных, используют дополнительные индуцибельные генетические программы - системы холодового и теплового
шока, относящиеся к так называемым стрессовым системам. Особенности чувствительности бактерий к температуре учитываются при дезинфекции и стерилизации. Отношение к кислотности среды
Одним из важных факторов, определяющих возможности жизни бактерий, является кислотность среды (концентрация ионов водорода). Большинство возбудителей болезней человека живут при рН среды от 4,0 до 9,0 с оптимумом около 7,0. Вместе с тем известны микробы, предпочитающие щелочную среду рН от 9,0 и выше (алкалофильные бактерии). К их числу можно отнести возбудитель холеры - Vibrio cholera. Некоторые микробы растут только в кислой среде при рН 4,0 и ниже (ацидофильные бактерии). Представители этой группы микроорганизмов используются в пищевой промышленности для получения молочнокислых продуктов. Известны микробы, устойчивые к изменениям рН среды и способные сохранять жизнеспособность как в сильнокислой, так и в сильнощелочной средах. К таким бактериям относятся возбудители туберкулеза, проказы и микобактериозов (Mycobacterium spp), а также актиномицеты и нокардии.
Отношение к молекулярному кислороду
Кислород, широко распространенный в природе, находится в свободном и связанном состоянии. В клетках он находится в связанном состоянии в составе воды и органических соединений. В атмосфере он присутствует в свободном состоянии в виде молекулярной формы, объемная доля которого составляет 21%. По отношению к кислороду, а также по использованию его в процессах получения энергии микроорганизмы подразделяются на три группы: облигатные аэробы, облигатные анаэробы, факультативные анаэробы.
Облигатные аэробы растут и размножаются только в присутствии кислорода, используют кислород для получении энергии путем кислородного дыхания. Энергию получают оксидативным метаболизмом, используя кислород как терминальный акцептор электронов в реакции, катализируемой цитохромоксидазой.
Облигатные аэробы подразделяются на строгие аэробы, которые растут при парциальном давлении воздуха, и микроаэрофилы, которые, используя кислород в процессах получения энергии, растут при его пониженном парциальном давлении. Это связано с тем, что у микроаэрофилов имеются ферменты, которые инак-
тивируются при контакте с сильными окислителями и активны только при низких значениях парциального давления кислорода, например фермент гидрогеназа.
Облигатные анаэробы не используют кислород для получения энергии. Тип метаболизма у них бродильный, за исключением метаболизма у двух видов бактерий: Desulfovibrio и Desulfotomaculum, которые относятся к хемолитотрофам и обладают сульфатным дыханием. Облигатные анаэробы подразделяются на две группы: строгие анаэробы и аэротолерантные. Строгие анаэробы характеризуются тем, что молекулярный кислород для них токсичен: он убивает микроорганизмы или ограничивает их рост. Энергию строгие анаэробы получают маслянокислым брожением. К строгим анаэробам относятся, например, некоторые клостридии (C. botulinum, C. tetani), бактероиды.
Аэротолерантные микроорганизмы не используют кислород для получения энергии, но могут существовать в его атмосфере. К ним относятся молочнокислые бактерии, получающие энергию гетероферментативным молочнокислым брожением.
Факультативные анаэробы способны расти и размножаться как в присутствии, так и при отсутствии кислорода. Они обладают смешанным типом метаболизма. Процесс получения энергии у них может происходить кислородным дыханием в присутствии кислорода, а при его отсутствии переключаться на брожение. Для этих бактерий характерно наличие анаэробного нитратного дыхания.
Различное физиологическое отношение микроорганизмов к кислороду связано с наличием у них ферментных систем, позволяющих существовать в атмосфере кислорода. Следует отметить, что в окислительных процессах, протекающих в атмосфере кислорода, при окислении флавопротеидов образуются токсичные продукты: перекись водорода Н2О2 и закисный радикал кислорода О2- - соединение, имеющее неспаренный электрон. Эти соединения вызывают перекисное окисление ненасыщенных жирных кислот и окисление SH-групп белков.
Для нейтрализации токсичных форм кислорода микроорганизмы, способные существовать в его атмосфере, имеют защитные механизмы. У облигатных аэробов и факультативных анаэробов накоплению закисного радикала О2- препятствует фермент супероксиддисмутаза, расщепляющая закисный радикал на перекись
водорода и молекулярный кислород. Перекись водорода у этих бактерий разлагается ферментом каталазой на воду и молекулярный кислород.
Аэротолерантные микроорганизмы не имеют супероксиддисмутазы, и ее функцию восполняет высокая концентрация ионов марганца, который, окисляясь под действием О2-, убирает тем самым супероксидный ион. Перекись водорода у этих микроорганизмов разрушается ферментом пероксидазой в катализируемых ею реакциях окисления органических веществ.
Строгие анаэробы не имеют ни каталазы, ни пероксидазы. Однако супероксиддисмутаза встречается у многих строгих анаэробов, и наличие этого фермента коррелирует с их устойчивостью к кислороду. Некоторые строгие анаэробы (роды Bacteroides, Fusobacterium) не выносят присутствия даже незначительного количества молекулярного кислорода, тогда как некоторые представители рода Clostridium могут находиться в атмосфере кислорода. Для культивирования строгих анаэробов создаются условия, позволяющие удалять атмосферный кислород: использование специальных приборов, анаэростатов и анаэробных боксов, добавление в питательные среды редуцирующих кислород веществ, например тиогликолята натрия, использование поглотителей кислорода.
Отношение к излучению
Важнейшим естественным источником излучения для Земли является солнечная радиация. Поверхности Земли достигают преимущественно волны длиной от 300 нм и более, поскольку более короткие волны задерживаются атмосферой. Свет в диапазоне от 300 до 1000 нм, приходящийся в основном на видимый свет, оказывает заметное влияние на жизнь различных прокариотов, включая бактерии - возбудителей болезней человека. Излучение в этом диапазоне индуцирует в бактериальной клетке процессы фотореактивации, необходимые для поддержания постоянства состава ДНК и повышения выживаемости (световая репарация ДНК), а также синтез некоторых макромолекул. В медицине излучение используется для дезинфекции воздуха, различных поверхностей оборудования и материалов. Источником излучения в этом случае являются специальные лампы, получившие название бактерицидных ламп. Бактерицидное действие этих ламп связано с действием коротковолнового излучения от 220 до 300 нм. При этом излучение с длиной волны около 220 нм вызывает ионизацию молекул
кислорода с образованием озона (О3). Действие коротковолнового излучения в бактериальных клетках приводит к повреждениям ДНК, сопровождающимся или появлением мутаций, или гибелью клеток и изменению и разрушению других органических макромолекул. Среди бактерий наиболее устойчивыми к действию солнечной радиации и обработке ультрафиолетовым (УФ) светом искусственного происхождения являются их споры.
Радиоактивное излучение в естественных условиях преимущественно связано с излучением горных пород и сильно варьирует в различных географических точках, а также городах и сельской местности. В настоящее время мало известно о роли подобной радиации в изменении свойств бактерий, актуальных для практической медицины. Искусственная радиационная обработка, используемая для лечения ряда заболеваний (прежде всего злокачественных новообразований), может изменять состав нормальной микрофлоры, что требует коррекции для профилактики различных осложнений.
3.1.10. Рост и размножение
Рост бактериальных клеток связан с синтезом и накоплением всех компонентов, входящих в ее состав, и увеличением размера, характерного для данного вида. В условиях, обеспечивающих рост микробов, происходит и процесс их деления. Для большинства бактерий характерно поперечное бинарное деление, приводящее к образованию двух дочерних клеток. У грамположительных бактерий при этом происходит синтез перегородки между делящимися клетками. Перегородка начинает формироваться на периферии и «движется» к центру клетки. Для грамотрицательных бактерий характерно первоначальное формирование перетяжки, отделяющей клетки. После ее образования окончательное разделение дочерних клеток сопровождается синтезом перегородки между ними.
Деление бактериальной клетки начинается спустя некоторое время после завершения цикла репликации хромосомы, которая у бактерий протекает по полуконсервативному механизму. Это означает, что каждая из двух нитей ДНК хромосомы служит матрицей для синтеза комплементарной дочерней цепи ДНК. В процессе репликации бактериальной хромосомы участвует более 20 ферментов. Перед репликацией цепи родительской молекулы матричной цепи ДНК должны быть разделены. В этом процессе участвуют фермент
хеликаза, которая в энергопоглощаемой реакции расплетает двойную спираль, и фермент топоизомераза (гираза), которая предотвращает образование вторичных завитков. SSB-белок связывается с одноцепочечной ДНК, предотвращая повторное скручивание в двойную спираль. В результате образуется репликативная вилка (рис. 3.5). Синтез новых цепей ДНК осуществляется ферментом ДНК-полимеразой. ДНК-полимераза не способна инициировать новые цепи ДНК, а может присоединять комплементарные матрице нуклеотиды к свободному З'-концу растущей цепи. Поэтому для осуществления реакции полимеризации нуклеотидов на матрице родительской цепи полимеразе требуется затравка, праймер (от англ. primer - запал). Праймер представляет собой короткую нуклеотидную цепочку РНК, комплементарную матричной цепи, со свободным 3'-концом. Достраивание осуществляется присоединением к свободной гидроксильной группе 3'-конца затравки нового нуклеотида. Расплетенные цепи ДНК всегда содержат на 5'-конце несколько рибонуклеотидов, т.е. синтез ДНК начинается с синтеза РНК. РНК-затравку для синтеза ДНК образует специальный фермент ДНК-праймаза, способная инициировать синтез РНК по одноцепочечной ДНК матрицы при отсутствии какойлибо затравки. После того как цепь ДНК начала синтезироваться, РНК-затравка удаляется, а удаляющиеся бреши застраиваются ДНК-полимеразой с высокой точностью. Сохранение высокой
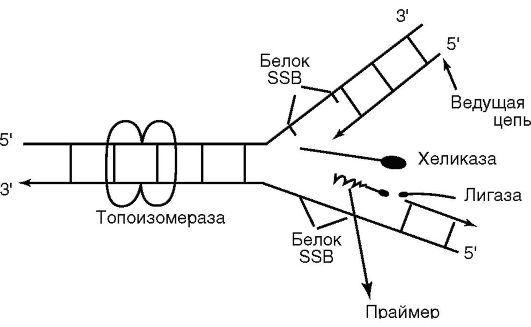 Рис. 3.5. Схема репликативной вилки
Рис. 3.5. Схема репликативной вилки
степени точности, необходимой при репликации, обеспечивается различными функциями ДНК-полимеразы. Кроме полимеразной активности она способна к проверке считывания. В ходе последней фермент проверяет, правильно ли осуществлено присоединение очередного нуклеотида. Если выявляется нарушение правила комплементарности, проявляется третья функция данного фермента - экзонуклеазная и происходит отщепление неправильно присоединенного нуклеотида. После его удаления вновь происходит полимеразная реакция с последующей проверкой ее правильности. В целом в благоприятных условиях синтез ДНК в клетке значительно опережает скорость ее деления. В реальных условиях одна микробная клетка содержит от 2 до 10 копий хромосом. Показано, что многие бактерии без повреждения клетки выделяют избыток ДНК в окружающую среду. Этот процесс играет важную роль в обмене генетической информацией между бактериями.
Процесс репликации ДНК бактерии продолжается до тех пор, пока не удвоится вся ДНК. Репликация начинается в одной избранной области, называемой origin (от англ. origin - начало), имеющей определенную последовательность нуклеотидов. На origin может возникать одна или две репликативные вилки. Последовательность нуклеотидов на origin-участке способствует необходимому для репликации ДНК расплетанию двойной спирали и служит местом «посадки» на ДНК комплекса ферментов, участвующих в репликации. Правильное распределение вновь синтезированных нитей ДНК по дочерним клеткам достигается у бактерий за счет прикрепления ДНК к мембране. Пространственная организация участка прикрепления и зоны роста мембраны и клеточной стенки обеспечивает автоматическое растаскивание двух копий реплицированной ДНК по дочерним клеткам. Размножение бактерий бинарным делением приводит к росту числа бактериальных клеток в геометрической прогрессии.
При внесении бактерий в питательную среду они растут и размножаются до тех пор, пока содержание какого-нибудь из необходимых компонентов среды не достигает минимума, после чего рост и размножение прекращаются. Если на протяжении всего этого времени не прибавлять питательные вещества и не удалять конечные продукты обмена, то получаем статическую бактериальную культуру. Статическая (периодическая) культура бактерий ведет себя как многоклеточный организм с генетическим ограничением роста. Если построить график, по оси абсцисс которого отложить
время, а по оси ординат - число клеток, то получим кривую, описывающую зависимость числа образующихся клеток от времени размножения, которая называется кривой роста (рис. 3.6).
На кривой роста бактерий в жидкой питательной среде можно различить несколько фаз, сменяющих друг друга в определенной последовательности.
• Начальная - лаг-фаза (от англ. lag - отставать), охватывает промежуток времени между инокуляцией (посевом бактерий) и началом размножения. Ее продолжительность в среднем 2-5 ч и зависит от состава питательной среды и возраста засеваемой культуры. Во время лаг-фазы происходит адаптация бактериальных клеток к новым условиям культивирования, идет синтез индуцибельных ферментов.
• Экспоненциальная (логарифмическая) фаза характеризуется постоянной максимальной скоростью деления клеток. Эта скорость зависит от вида бактерий и питательной среды. Время удвоения клеток называется временем генерации, которое варьирует от вида бактериальной культуры: у бактерий рода Pseudomonas оно равняется 14 мин, а у Mycobacterium - 24 ч. Величина клеток и содержание белка в них во время экспоненциальной фазы остаются постоянными. Бактериальная культура в этой фазе состоит из стандартных клеток.
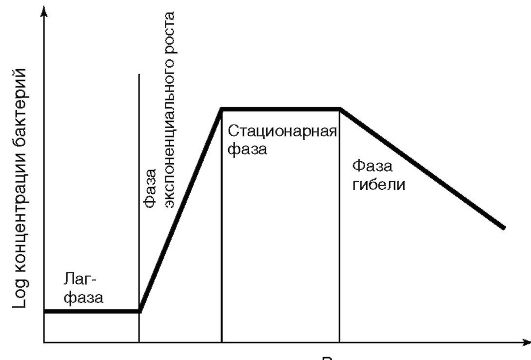 Рис. 3.6. Кривая бактериального роста
Рис. 3.6. Кривая бактериального роста
• Стационарная фаза наступает тогда, когда число клеток перестает увеличиваться. Так как скорость роста зависит от концентрации питательных веществ, то при уменьшении содержания последних в питательной среде уменьшается и скорость роста. Снижение скорости роста происходит также из-за большой плотности бактериальных клеток, снижения парциального давления кислорода, накопления токсичных продуктов обмена. Продолжительность стационарной фазы составляет несколько часов и зависит от вида бактерий и особенностей их культивирования.
• Фаза отмирания наступает вследствие накопления кислых продуктов обмена или в результате аутолиза под влиянием собственных ферментов. Продолжительность этой фазы колеблется от десятка часов до нескольких недель.
Продолжительность жизни бактерий мало изучена. Известно, что мезофилы на питательной среде при комнатной температуре в условиях, когда размножение бактерий минимально, могут сохранять свою жизнеспособность в течение 1-2 лет. Очевидно, что биологическая смерть бактерий в большей степени связана с ограничением числа возможных делений. Считается, что большинство бактерий могут делиться около 50 раз, после чего клетка погибает. Механизмы гибели остаются не до конца изученными, но показано существование у бактерий генов, изменение активности которых специфически направлено на самоуничтожение клеток. Постоянное нахождение бактериальной популяции в логарифмической фазе роста наблюдается в непрерывной культуре, что достигается постепенным дозированием поступления питательных веществ, контролем плотности бактериальной суспензии и удалением метаболитов. Непрерывные бактериальные культуры используются в биотехнологических процессах.
Накопление бактериальной массы (числа бактерий) при культивировании зависит от многих факторов (качество питательных сред, посевная доза, температура выращивания, рН, наличие активирующих рост добавок и др.).
На жидких питательных средах рост и размножение бактерий проявляются в виде диффузного помутнения, образования придонного осадка или поверхностной пленки. Особенностью размножения бактерий роста Leptospira на жидких средах является отсутствие видимых проявлений роста.
На плотных питательных средах бактерии образуют скопление клеток - колонии, которые принято считать потомком одной клетки. Колонии различаются формой, размерами, поверхностью, прозрачностью, консистенцией и окраской. Колонии с гладкой блестящей поверхностью принято называть колониями в S-форме (от англ. smooth - гладкий). Колонии с матовой шероховатой поверхностью называют R-формами (от англ. rough - шероховатый).
Окраска колоний определяется способностью бактерий синтезировать пигменты. Пигменты различаются по цвету, химическому составу и растворимости. Пигменты предохраняют бактериальную клетку от УФ-лучей, обезвреживают токсичные кислородные радикалы, обладают антибиотическими свойствами, принимают участие в реакциях, сопутствующих фотосинтезу в фототрофных бактериях.
Вид, форма, цвет и другие особенности колоний, а также характер роста на плотных питательных средах определяются как культуральные свойства бактерий и учитываются при их идентификации.
Помимо бинарного деления, некоторые представители царства Procaryotae имеют иные способы размножения.
Актиномицеты могут размножаться путем фрагментации гифов. Представители семейства Streptomycetaceae размножаются спорами.
Микоплазмы являются полиморфными бактериями, что обусловлено особенностями их размножения. Помимо поперечного деления, если оно происходит синхронно с синтезом ДНК, микоплазмы могут размножаться почкованием. В этом случае основной морфологической репродуцирующейся единицей являются элементарные тельца сферической или овоидной формы, размножающиеся фрагментацией и почкованием.
Хламидии не обладают способностью к бинарному делению. Они проходят через цикл развития, который предусматривает существование двух форм: внеклеточных инфекционных, малых размеров элементарных телец, не обладающих способностью к бинарному делению, и внутриклеточного, метаболически активного, крупных размеров ретикулярного тельца, способного к бинарному делению. В результате бинарного деления ретикулярного тельца формируются дочерние элементарные тельца, которые выделяются из клетки.
Некоторые спирохеты, например Treponema pallidum, способны образовывать в неблагоприятных условиях цисты, которые, распадаясь на зерна, дают потомство новым бактериальным клеткам.
Некультивируемые формы бактерий. Некоторые неспорообразующие бактерии способны переживать неблагоприятные для размножения условия окружающей среды, переходя в некультивируемое состояние. В этом состоянии бактериальные клетки сохраняют свою метаболическую активность, но не способны к непрерывному клеточному делению, необходимому для роста на жидких и плотных питательных средах. При смене условий существования, в частности при попадании в организм человека или животных, клетки вновь приобретают способность к размножению и сохраняют свой патогенный потенциал. Переход в некультивируемое (покоящееся) состояние обеспечивает сохранение патогенных бактерий в межэпидемические и межэпизоотические периоды. При переходе в некультивируемую форму бактериальные клетки уменьшаются в размерах, приобретают сферическую форму, меняют вязкость ЦПМ. У них сохраняются транспорт электронов по дыхательной цепи и невысокий уровень метаболической активности. На переход в некультивируемую форму влияют температура, концентрация солей, свет, парциальное давление кислорода, содержание питательных веществ, а также метаболиты водорослей, находящихся в биоценозе с бактериями. Выявить наличие бактерий, находящихся в некультивируемой форме, можно с помощью полимеразной цепной реакции (ПЦР) (см. раздел 5.6.3) или красителей, меняющих окраску в окисленной и восстановленной формах. Возврат способности к размножению и росту находящихся в покоящейся форме клеток могут вызвать естественные факторы: простейшие, обитатели почв и водоемов, фитогормоны, выделяемые корневыми волосками растений.
3.1.11. Условия культивирования бактерий
Для культивирования бактерий необходимо соблюдать ряд условий.
• Наличие полноценной питательной среды. Каждая питательная среда независимо от сложности состава и цели применения (см. главу 3.1) должна обладать водной основой, органическим источником углерода и энергии, определенным рН, осмотическим давлением.
• Температура культивирования. Температура влияет на скорость размножения. Для поддержания требуемой температуры используют специальные приборы - термостаты.
• Атмосфера культивирования. Для роста и размножения строгих аэробов необходим кислород. Аэробы хорошо растут на поверхности агара на чашках Петри или в тонком верхнем слое жидкой среды. Для обеспечения роста и размножения строгих аэробов в глубинных слоях жидкой среды необходимо диффузное распределение кислорода по всему объему питательной среды. Это достигается непрерывным перемешиванием или встряхиванием питательной среды, т.е. аэрированием. Аэрирование осуществляется на специальных аппаратах - встряхивателях.
Для культивирования факультативных анаэробов используют те же методы, так как в присутствии кислорода у них преобладает оксидативный метаболизм над ферментацией как наиболее энергетически выгодный.
Микроаэрофилы размножаются при пониженном парциальном давлении кислорода. Этого можно достичь повышением парциального давления СО2 в атмосфере культивирования до 1-5% против 0,03% СО2 в атмосфере воздуха. Для этих же целей используют специальные СО2-инкубаторы или же посевы помещают в эксикаторы, в которых устанавливают горящую свечу.
Облигатные анаэробы для своего роста и размножения требуют исключения доступа кислорода воздуха. Это достигается следующими мерами:
- добавлением к питательным средам редуцирующих кислород веществ: тиогликолевой и аскорбиновой кислот, цистеина, сульфидов;
- регенерацией от кислорода воздуха жидких питательных сред путем их кипячения с последующим плотным закупориванием сосудов, в которые налиты среды, резиновыми пробками;
- использованием поглотителей кислорода, щелочного пирогаллола, и других средств, помещая их в герметически закрываемые емкости газ-паки. Этот метод используется для культивирования аэротолерантных бактерий;
- механическим удалением кислорода воздуха с последующим заполнением емкости инертным газом (для этих целей используют анаэростаты и анаэробные боксы).
Для культивирования хемо- и фотоавтотрофных бактерий создается атмосфера, насыщенная СО2.
• Время культивирования зависит от времени генерации. Большинство бактерий культивируют для получения видимого
роста в течение 18-48 ч. Для культивирования возбудителя коклюша требуется 5 сут, для культивирования М. tuberculosis - 3-4 нед.
• Освещение. Для выращивания фототрофных микроорганизмов необходим свет. Некоторые условно-патогенные микобактерии в зависимости от освещенности образуют пигмент, что используется при их идентификации. Культивирование абсолютных внутриклеточных паразитов, бактерий, относящихся к родам Rickettsia, Ehrlichia, Coxiella, Chlamydia, осуществляют на культурах клеток или в организме животных и членистоногих, а также в куриных эмбрионах (за исключением эрлихий). Куриные эмбрионы используют также для культивирования бактерий, обладающих высоким уровнем гетеротрофности, например родов Borrelia, Legionella.
3.1.12. Поведение бактерий в бактериальных сообществах
Популяция бактерий, будь то в окружающей среде или в организме хозяина, представляет собой не совокупность отдельных клеток, а сообщество, живущее по социальным законам, члены которого общаются между собой посредством понятного им языка. В настоящее время стало известно явление, получившее название «quorum sensing», или чувство кворума. «Quorum sensing» - это межклеточный механизм бактериального общения, предназначенный для контроля экспрессии генов в зависимости от плотности бактериальной популяции
Регуляторные системы «quorum sensing» обычно состоят из двух компонентов: небольшой диффундирующей сигнальной молекулы и транскрипционного активаторного белка. В грамотрицательных бактериях сигнальные молекулы называются аутоиндукторами. По типу «quorum sensing» регулируется широкий ряд физиологических процессов, включая биолюминесценцию, синтез антибиотиков, детерминант вирулентности, перенос конъюгативных плазмид (см. главу 5). Такой тип межклеточной коммуникации позволяет индивидуальным бактериям следить за плотностью собственной популяции в окружающей среде, будь то внешняя среда или организм хозяина, и регулировать экспрессию специфических генов. Патогенным бактериям, которые вызывают развитие заболевания, необходимо достичь критической плотности для эффективного распространения и заселения соответствующих ниш в организме
хозяина. Патогенные бактерии чувствуют необходимость экспрессии детерминант вирулентности при достижении определенной концентрации.
Социальным поведением патогенных бактерий объясняется такое важное для медицины явление, как образование биопленок. Биопленки представляют высокоорганизованные сообщества бактерий, необратимо прикрепленных к субстрату и друг к другу и защищенных продуцируемым этими клетками внеклеточным полимерным матриксом. Они снабжены каналами для водоснабжения, распределения питательных веществ между членами сообщества и удаления отходов жизнедеятельности. Биопленки могут быть образованы бактериями одного или нескольких видов и состоят из активно функционирующих и покоящихся (некультивируемых) клеток. Образование биопленки является одной из основных стратегий выживания бактерий в окружающей среде, поскольку в составе биопленки они защищены от антибактериальных препаратов, включая антибиотики, дезинфектанты, бактериофаги. Многие хронические инфекции, возникновение которых связано с использованием медицинского имплантированного оборудования - катетеров, протезов, искусственных клапанов сердца, обусловлены способностью бактерий расти в виде биопленок на поверхности этих устройств.
Образование биопленки начинается с прикрепления к твердой поверхности отдельной бактериальной клетки, которая выделяет полисахариды. Делящиеся затем клетки образуют микроколонию уже внутри полисахаридного матрикса. Эти микроколонии сливаются или включают в свой состав бактерии этого или другого вида, и процесс заканчивается образованием биопленки. В процессе образования биопленки важная роль принадлежит полярно расположенным пилям IV типа (рис. 3.7).
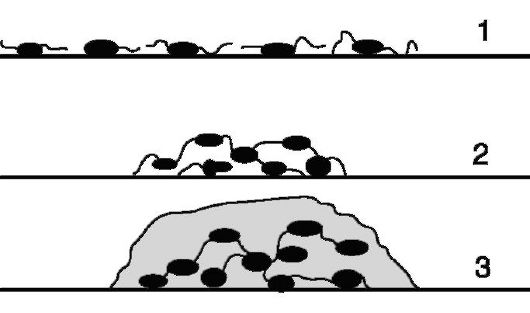 Рис. 3.7. Образование
биопленки: 1 - прикрепление к твердой поверхности отдельных
бактериальных клеток; 2 - агрегация клеток с участием пилей IV типа; 3 -
синтез экзополисахарида
Рис. 3.7. Образование
биопленки: 1 - прикрепление к твердой поверхности отдельных
бактериальных клеток; 2 - агрегация клеток с участием пилей IV типа; 3 -
синтез экзополисахарида
3.2. Физиология вирусов
Вирусы растут только внутриклеточно, т.е. являются облигатными внутриклеточными паразитами. В клетке они могут находиться в различных состояниях.
Нарушения, вызываемые вирусами, весьма разнообразны: от продуктивной инфекции с образованием вирусного потомства и гибелью клетки до продолжительного взаимодействия вируса с клеткой в виде латентной инфекции или злокачественной трансформации клетки. Инфицирование клетки вирусом может иметь следующие последствия:
• разрушение клетки (некроз) в результате цитоцидной инфекции, т.е. репродукция вируса приводит к цитоцидному действию (в культуре клеток происходит цитопатический эффект - клетки округляются, отделяются от соседних клеток, образуются многоядерные гигантские клетки, вакуоли и включения);
• разрушение клетки (апоптоз) в результате инициации вирусом програмированной клеточной гибели, при этом вирусный репликативный цикл часто прерывается;
• разрушение клетки в итоге не самим вирусом, а иммунными реакциями организма;
• вирус находится внутри клетки, но не разрушает ее (латентная инфекция);
• вирус трансформирует клетку организма в раковую клетку. Хорошо изучены три основных типа взаимодействия вируса с
клеткой: продуктивный, абортивный и интегративный.
Продуктивный тип взаимодействия завершается воспроизводством вирусного потомства - многочисленных вирионов и гибелью зараженных клеток (цитоцидное действие). Некоторые вирусы выходят из клеток, не разрушая их (нецитоцидное действие).
Абортивный тип взаимодействия не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип взаимодействия, или вирогения, характеризуется встраиванием (интеграцией), вирусной ДНК в виде провируса в хромосому клетки и их совместной репликацией.
3.2.1. Продуктивный тип взаимодействия вируса с клеткой
Продуктивный тип взаимодействия вируса с клеткой, т.е. репродукция вируса (от лат. re - повторение, productio - производство), проходит несколько стадий:
1) адсорбция вириона на клеточной мембране;
2) проникновение вириона в клетку, «раздевание» и высвобождение вирусного генома (депротеинизация вируса);
3) синтез вирусных компонентов;
4) сборка реплицированной нуклеиновой кислоты и новых капсидных белков;
5) выход вирусного потомства из клетки.
Адсорбция вириона, т.е. его прикрепление к клеточной мембране, - первая стадия репродукции вирусов. Она происходит в результате взаимодействия поверхностных молекул (белковых лигандов) вируса с мембранными рецепторами клеток вирусов. Белки поверхности вирусов, например гликопротеины липопротеиновой оболочки, узнающие специфические клеточные рецепторы и взаимодействующие с ними, называются прикрепительными белками.
Лиганды вирусов и специфические рецепторы клеток имеют различную природу. Так, гемагглютининовые шипы вируса гриппа связываются с сиаловой кислотой в составе гликопротеинов и гликолипидов (ганглиозидов) клеток дыхательных путей. Гликопротеины вируса иммунодефицита человека взаимодействуют с CD4- молекулами и хемокиновыми рецепторами Т-хелперов, моноцитов и дентритных клеток. Капсидные белки вируса полиомиелита связываются с CD155-молекулой, а капсидные белки риновирусов - с ICAM (молекулой адгезии) клеток. На клетке находятся десятки тысяч специфических рецепторов, поэтому на ней могут адсорбироваться десятки и сотни вирионов, но проникают в клетку только определенные вирионы или их содержимое.
В основе избирательности поражения вирусами определенных клеток, тканей и органов (так называемого тропизма) лежат специфичность рецепторов поражаемой клетки и возможность развития в ней репродуктивного цикла вируса (пермиссивные условия клетки). Например, вирусы, репродуцирующиеся преимущественно в клетках печени, называются гепатотропными, в нервных клетках - нейротропными, в иммунокомпетентных клетках - иммунотропными и т.д.
Проникновение вирусов в клетку возможно в результате рецепторзависимого эндоцитоза или слияния оболочки вируса с клеточной мембраной. Возможно также сочетание этих механизмов. Рецепторзависимый эндоцитоз происходит в результате захватывания и поглощения вириона клеткой: клеточная мембрана с прикрепленным вирионом впячиваются с образованием эндосомы (внутриклеточной вакуоли). Эндоцитоз вирусов осуществляется с помощью везикул, покрытых клатрином («ямки, окаймленные клатрином»). Содержимое эндосомы закисляется, что приводит к слиянию липопротеиновой оболочки сложного вируса с мембраной эндосомы и выходу вирусного нуклеокапсида в цитозоль клетки. Эндосомы объединяются с лизосомами, которые разрушают оставшиеся вирусные компоненты. Пенетрация компонентов вируса в цитозоль обычно происходит в ранних или поздних эндосомах при уменьшенном значении pH.
Стали известны новые клатриннезависимые, альтернативные пути соединения вируса с эндосомами. Одним из них может быть макропиноцитоз с образованием крупной вакуоли, окруженной плазматической мембраной, наполненной в основном жидкостью. Другим путем попадания вируса может быть вовлечение эндоплазматического ретикулума. Этот путь начинается с формирования различных везикул (кальвеолярный эндоцитоз). Вируснесущие везикулы, сформированные в плазматической мембране, маленькие (диаметр около 70 нм). В результате проникновение вируса в цитозоль может происходить на уровне плазматической мембраны, эндосомы, кальвеосомы и эндоплазматического ретикулума.
Слияние вириона с клеточной мембраной характерно только для некоторых оболочечных вирусов (герпесвирусов, парамиксовирусов, ретровирусов), в составе которых имеются белки слияния. В результате взаимодействия вирусного белка слияния с липидами клеточной мембраны вирусная липопротеиновая оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса попадает в цитозоль клетки.
Существует три варианта проникновения безоболочечных вирусов в клетку: мембранный прокол (вирион образует пору в мембране, через которую геном попадает в цитозоль, а капсид в него не попадает; перфорация (капсид переносится через мембрану без основного лизиса мембраны); лизис (вирионы индуцируют поломку мембраны цитоплазматических органелл, что способствует
проникновению вируса и его компонентов в цитозоль). Выход безоболочечных (простых) вирусов из эндосомы в цитозоль остается малоизученным.
Попав в клетку, вирусы лишаются многих белков («раздевание», или депротеинизация вирусов). В результате депротеинизации удаляются поверхностные структуры вируса и высвобождается его внутренний компонент, способный вызывать инфекционный процесс. Первые этапы «раздевания» вируса начинаются в процессе его проникновения в клетку путем слияния вирусных и клеточных мембран или же при выходе вируса из эндосомы в цитозоль. Последующие этапы «раздевания» вируса тесно взаимосвязаны с их внутриклеточным транспортом к местам депротеинизации. Для разных вирусов существуют свои специализированные участки «раздевания» в клетке: для пикорнавирусов - в цитоплазме с участием лизосом, аппарата Гольджи, для герпесвирусов - околоядерное пространство или поры ядерной мембраны, для аденовирусов - сначала структуры цитоплазмы, а затем ядро клетки. Конечными продуктами «раздевания» могут быть нуклеиновая кислота, нуклеопротеид (нуклеокапсид) или сердцевина вириона. Так, конечным продуктом «раздевания» пикорнавирусов является нуклеиновая кислота, ковалентно связанная с одним из внутренних белков. А у многих оболочечных РНК-содержащих вирусов конечными продуктами «раздевания» могут быть нуклеокапсиды или сердцевины, которые не только не препятствуют экспрессии вирусного генома, а, более того, защищают его от клеточных протеаз и регулируют последующие биосинтетические процессы.
Следующей стадией репродукции является синтез белков и нуклеиновых кислот вируса, который разобщен во времени и пространстве.
Синтез вирусных белков. В вирусинфицированной клетке синтезируются две группы белков: неструктурные белки, обслуживающие разные этапы репродукции вируса; структурные белки, которые входят в состав вириона (нуклеопротеины, связанные с геномом вируса, капсидные и оболочечные белки). К неструктурным белкам относятся: ферменты синтеза нуклеиновых кислот (РНКили ДНК-полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома; белки-регуляторы; предшественники вирусных белков, отличающиеся своей нестабильностью в результате быстрого нарезания на структурные белки; ферменты,
модифицирующие вирусные белки, например протеиназы и протеинкиназы.
Синтез белков в клетке осуществляется в соответствии с хорошо известными процессами транскрипции путем «переписывания» генетической информации с нуклеиновой кислоты в нуклеотидную последовательность иРНК, или мРНК, и трансляции - считывания иРНК на рибосомах с образованием белков. Передача наследственной информации в отношении синтеза иРНК у разных групп вирусов неодинакова.
• ДНК-содержащие вирусы имеют ДНК-геном, транскрибирующийся в ядре клетки с помощью клеточной РНК-полимеразы, в результате чего образуется иРНК, которая транслируется с образованием белка вируса. Особенностью этого процесса является синтез иРНК в ядре с помощью клеточной РНКполимеразы (у аденовирусов, папилломавирусов, герпесвирусов) или в цитоплазме с помощью собственной РНКполимеразы (у поксвирусов). Таким образом, синтез белка реализуется по схеме: геномная ДНК вируса → транскрипция иРНК → трансляция белка вируса.
• Плюс-нитевые РНК-содержащие вирусы (пикорнавирусы, флавивирусы, тогавирусы) имеют геном, выполняюший функцию иРНК; он распознается и транслируется рибосомами. Белки этих вирусов синтезируются без процесса транскрипции по схеме: геномная РНК вируса - трансляция белка вируса.
• Минус-нитевые РНК-содержащие вирусы (минусоднонитевые - ортомиксовирусы, парамиксовирусы, рабдовирусы и двунитевые - реовирусы) имеют геном, выполняющий роль матрицы, с которой транскрибируется иРНК, при участии РНК-полимеразы, связанной с нуклеиновой кислотой вируса. Синтез белка у них происходит по схеме: геномная РНК вируса - транскрипция иРНК - трансляция белка вируса.
• Ретровирусы (ВИЧ, онкогенные ретровирусы) имеют диплоидный геном, состоящий из двух идентичных молекул РНК. В состав ретровируса включена вирионная обратная транскриптаза, или ревертаза, с помощью которой осуществляется процесс обратной транскрипции, т.е. на матрице геномной РНК синтезируется комплементарная однонитевая ДНК. Комплементарная нить ДНК копируется с образованием двунитевой
комплементарной ДНК, которая интегрирует в клеточный геном и в его составе транскрибируется в иРНК с помощью клеточной ДНК-зависимой РНК-полимеразы. Синтез белков ретровирусов осуществляется по схеме: геномная РНК вируса - комплементарная ДНК - транскрипция иРНК - трансляция белка вируса. Репликация вирусных геномов, т.е. накопление копий вирусных геномов, которые используются при сборке вирионов в клетке, отличается у вирусов, имеющих двунитевую ДНК, однонитевую ДНК, плюс-однонитевую РНК, минус-однонитевую РНК, двунитевую РНК и идентичные плюс-нитевые РНК (ретровирусы).
• Двунитевые ДНК-вирусы. К ним относятся вирусы, содержащие двунитевую ДНК в линейной (герпесвирусы, аденовирусы и поксвирусы) или кольцевой (папилломавирусы и полиомавирусы) форме. В репликации вирусных геномов (см. табл. 2.2) участвуют вирусные ДНК-зависимые ДНК-полимеразы (у аденовирусов, герпесвирусов и поксвирусов) или клеточные ДНК-зависимые ДНК-полимеразы (у папилломавирусов, полиомавирусов и анелловирусов). Двунитевые вирусные ДНК реплицируются обычным полуконсервативным механизмом: после расплетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре. Своеобразный механизм репродукции с включением процесса обратной транскрипции имеют гепаднавирусы (см. ниже).
• Однонитевые ДНК-вирусы представлены парвовирусами, которые используют клеточную ДНК-полимеразу для создания двунитевого вирусного генома, так называемой репликативной формы последнего. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
• Плюс-однонитевые РНК-вирусы включают большую группу вирусов (пикорнавирусы, флавивирусы, тогавирусы), у которых геномная плюс-нить РНК выполняет функцию иРНК - матрицы для синтеза белка. На ее основе синтезируется по-
липротеин, который расщепляется на фрагменты: вирусные РНК-зависимую РНК-полимеразу, протеазы и капсидные белки. Вирусная РНК-полимераза транскрибирует геномную плюс-нить РНК в минус-нить РНК, на матрице которой синтезируется геномная плюс-нить РНК. Вирионы формируются в цитоплазме.
• Минус-однонитевые РНК-вирусы (аренавирусы, борнавирусы, рабдовирусы, парамиксовирусы, ортомиксовирусы, филовирусы) имеют вирионную РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус-нить РНК трансформируется вирионной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства. Вирионы формируются в цитоплазме.
• Двунитевые РНК-вирусы. Репликация этих вирусов (реовирусы и ротавирусы) сходна с репликацией минус-однонитевых РНК-вирусов. Образовавшиеся в процессе транскрипции плюс-нити РНК не только функционируют как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
• Вирусы с обратной транскрипцией. К обратнотранскрибирующимся вирусам относятся представители семейств Retroviridae и Hepadnaviridae. Ретровирусы, в частности ВИЧ, являются плюс-нитевыми диплоидными РНК-содержащими вирусами. Вирионная обратная транскриптаза ретровирусов синтезирует (на матрице РНК вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо. Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. В результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы образуются вирионные РНК.
Формирование вирионов. Белки и нуклеиновые кислоты вируса синтезируются в разных частях клетки, вследствии чего этот спо-
соб репродукции вирусов получил название дисъюнктивного (от лат. disjunctus - разобщенный). Синтезированные компоненты вириона транспортируются в различные участки ядра или цитоплазмы клетки - места сборки вируса, которая происходит с участием гидрофобных, ионных, водородных связей и стерического соответствия. Формирование вирионов - многоступенчатый процесс с образованием промежуточных форм, отличающихся от зрелых вирионов по составу полипептидов. Сборка простых вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов. У сложных вирусов сначала формируются нуклеокапсиды, которые окружаются модифицированной мембраной клетки (будущей липопротеиновой оболочкой вируса). В процессе сборки вирионов в их структуры включаются отдельные липиды и углеводы клетки-хозяина. Так, формирование вирионов в ядре клетки происходит с участием мембраны ядра, а формирование вирионов в цитоплазме - с участием мембран эндоплазматической сети или плазматической мембраны, куда встраиваются гликопротеины и другие белки оболочки вируса. У ряда сложных минус-нитевых РНК-вирусов (ортомиксовирусов, парамиксовирусов) в сборку вовлекается матриксный белок (М-белок), расположенный под модифицированной клеточной мембраной - будущей оболочкой вириона. Обладая гидрофобными свойствами, он выполняет роль посредника между нуклеокапсидом и липопротеиновой оболочкой вируса.
Выход вирусов из клетки. Продолжительность цикла вирусной репродукции колеблется от 6-8 ч (вирус гриппа, пикорнавирусы) до более чем 40 ч (некоторые герпесвирусы). Вирусное потомство составляет 10-1000 зрелых вирионов и в несколько раз большее количество дефектных вирионов. Репродукция вирусов заканчивается выходом их из клетки, который происходит взрывным путем почкованием или экзоцитозом.
• Взрывной путь характерен для простых (безоболочечных) вирусов: из погибающей клетки одновременно выходит большое количество вирионов.
• Почкование, экзоцитоз присущи сложным вирусам, имеющим липопротеиновую оболочку, которая является производной от клеточных мембран. Сначала образовавшийся нуклеокапсид или сердцевина вириона транспортируется к участкам клеточных мембран, в которые уже встроены вирусспецифические
белки, после чего начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложного вируса, а клетка может длительно оставаться жизнеспособной, продуцируя вирусное потомство. Вирусы, формирующиеся в ядре клетки (например, герпесвирусы) почкуются в перинуклеарное пространство через модифицированную ядерную мембрану, приобретая таким образом липопротеиновую оболочку. Затем они транспортируются в составе цитоплазматических везикул на поверхность клетки.
Почкование вирусов, формирующихся в цитоплазме, может происходить либо через плазматическую мембрану (например, парамиксовирусы, тогавирусы), либо через мембраны эндоплазматической сети с последующим их выходом на поверхность клетки (буньявирусы).
3.2.2. Программируемая клеточная смерть (апоптоз)
Вирусы могут вызывать апоптоз инфицированной клетки, предотвращая распространение инфекционного процесса на другие клетки. С другой стороны, многие вирусы имеют механизмы, предотвращающие апоптоз клетки. Так, некоторые ДНК-вирусы кодируют белки, сходные с клеточными Bcl-2-белками, контролирующими апоптоз в результате усиления клеточной пролиферации.
3.2.3. Непродуктивные инфекции
Иногда вирус заражает клетку, но его репродукция не завершается. Если вирусный геном персистирует в клетке, то говорят о латентной инфекции.
Латентная инфекция поддерживается в инфицированной клетке в виде последовательности вирусной ДНК, интегрированной в геном клетки, или в виде множественных копий ковалентно замкнутой циркулярной ДНК вируса. Например, герпесвирусы обычно вызывают латентные инфекции, при которых вирусный геном поддерживается как циркулярная эписома в ядре, экспрессируя только несколько вирусных генов и не образуя инфекционного вируса. В эукариотической клетке вирусная ДНК связана с гистонами этой клетки, которые играют стабилизирующую роль в латенции.
Абортивный тип взаимодействия вирусов с клеткой не завершается образованием вирусного потомства. Причины развития абор-
тивного типа разнообразны: заражение чувствительных клеток дефектными вирусами или дефектными вирионами; заражение стандартным вирусом генетически резистентных к нему клеток; заражение стандартным вирусом чувствительных клеток в непермиссивных (неразрешающих) условиях. Различают дефектные вирусы и дефектные вирионы.
• Дефектные вирусы существуют как самостоятельные виды, которые репродуцируются лишь при наличии вируса-помощника (например, вирус гепатита D репродуцируется только в присутствии вируса гепатита B).
• Дефектные вирионы обычно лишены части генетического материала и могут накапливаться в популяции многих вирусов при множественном заражении клеток. Имеются дефектные интерферирующие частицы (ДИ-частицы), которые интерферируют с репродукцией стандартного вируса и подавляют воспроизводство вирусного потомства. Таким образом, ДИ-частицы могут защищать организм от болезнетворного вируса.
Абортивный тип взаимодействия чаще наблюдается при заражении непермиссивных (нечувствительных) клеток стандартным вирусом. Механизм генетически обусловленной резистентности клеток к вирусам широко варьирует. Он может быть связан: с отсутствием на плазматической мембране специфических рецепторов для вирусов; неспособностью данного вида клеток инициировать трансляцию вирусной иРНК; отсутствием специфических протеаз или нуклеаз, необходимых для синтеза вирусных макромолекул и т.д. Абортивный тип взаимодействия может также возникать при изменении условий, в которых происходит репродукция вирусов: повышение температуры организма, изменение рН в очаге воспаления, введение в организм противовирусных препаратов и др. Некоторые абортивные инфекции могут приводить к уничтожению клетки хозяина. При устранении неразрешающих условий абортивный тип переходит в продуктивный тип взаимодействия вирусов с клеткой.
Интегративный тип взаимодействия вирусов с клеткой (вирогения) заключается во взаимном сосуществовании вируса и клетки в результате интеграции (встраивания) генома вируса в хромосому клетки хозяина. При этом интегрированный геном вируса реплицируется и функционирует как составная часть генома клетки.
Интегративный тип взаимодействия характерен для умеренных ДНК-содержащих бактериофагов, онкогенных вирусов и некоторых инфекционных как ДНК-содержащих (например, вируса гепатита В), так и РНК-содержащих (например, ВИЧ) вирусов. С геномом клетки интегрирует двунитевая ДНК вируса в кольцевой форме, которая прикрепляется к клеточной ДНК в месте гомологии нуклеотидных последовательностей и встраивается в определенный участок хромосомы при участии ферментов (рестриктаз, эндонуклеаз, лигаз).
Более сложным является процесс интеграции у РНК-содержащих вирусов. Он начинается с механизма обратной транскрипции, который заключается в синтезе комплементарной нити ДНК на матрице вирусной РНК с помощью вирионной обратной транскриптазы (ревертазы). После образования двунитевой ДНК и замыкания ее в кольцо происходит интеграция ДНК-транскрипта в хромосому клетки. Встроенная в хромосому клетки ДНК вируса называется провирусом, или провирусной ДНК. Провирус реплицируется в составе хромосомы и переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. Однако под влиянием некоторых физических или химических факторов провирус может исключаться из хромосомы клетки и переходить в автономное состояние с развитием продуктивного типа взаимодействия с клеткой.
Дополнительная генетическая информация провируса при вирогении сообщает клетке новые свойства, что может быть причиной онкогенной трансформации клеток и развития опухолей, а также развития аутоиммунных и хронических заболеваний. Сохранение вирусной информации в виде провируса в составе клеточного генома и передача ее потомству лежат в основе персистенции (от лат. persistentia - упорство, постоянство) вирусов в организме и развития латентных (скрытых) вирусных инфекций.
3.3. Культивирование вирусов
Вирусы культивируют для лабораторной диагностики вирусных инфекций, изучения патогенеза и иммунитета при вирусных инфекциях, а также для получения вакцин и диагностических препаратов. Поскольку вирусы являются облигатными внутриклеточными паразитами, их культивируют в организме лабораторных животных, развивающихся куриных эмбрионах и других птиц, а также на культурах клеток (тканей).
Присутствие вируса в исследуемом материале определяют с помощью методов индикации и идентификации. Индикация вирусов, т.е. неспецифическое обнаружение факта инфицирования, основано на выявлении биологических свойств вирусов и особенностей их взаимодействия с чувствительными клетками. Идентификация означает установление вида или типа вируса. Она осуществляется в основном с помощью иммунных реакций или молекулярногенетических методов (ПЦР и др.).
Вирусы можно культивировать в организме восприимчивых к ним лабораторных животных (белые мыши, хомячки, кролики, обезьяны и др.), которых заражают вируссодержащим материалом различными способами в зависимости от тропизма вирусов (подкожно, внутримышечно, интраназально, интрацеребрально и т.д.). Наличие вирусов выявляют по развитию у животных клинических проявлений заболевания, патоморфологическим изменениям органов и тканей, а также на основании реакции гемагглютинации (РГА) с вируссодержащим материалом. РГА основана на способности многих вирусов склеивать (агглютинировать) эритроциты своими гликопротеиновыми шипами (гемагглютининами).
Другой моделью культивирования вирусов являются куриные эмбрионы (5-12-дневные), которых заражают исследуемым материалом в различные полости и ткани зародыша. Таким образом можно культивировать вирусы гриппа, герпеса, натуральной оспы и др. Свидетельством репродукции вирусов в куриных эмбрионах являются специфические поражения оболочек и тела эмбриона (оспины, кровоизлияния), гибель эмбриона, положительная РГА с вируссодержащей жидкостью, полученной из полостей зараженного зародыша.
Часто вирусы культивируют на культуре клеток. Метод культур клеток был впервые разработан в 1949 г. Дж. Эндерсом и соавт., получившими за разработку техники культивирования вируса полиомиелита Нобелевскую премию в 1954 г. Изолированные клетки, полученные из различных органов и тканей человека, животных, птиц и других биологических объектов, размножают на искусственных питательных средах в специальной лабораторной посуде. Широко распространены культуры клеток из эмбриональных и опухолевых (злокачественно перерожденных) тканей, обладающих по сравнению с нормальными клетками взрослого организма более активной способностью к росту и размножению.
Культуры клеток обычно состоят из одного-двух основных типов клеток. Так, фибробласты имеют вытянутую форму, тогда как эпителиальные клетки имеют многоугольную форму.
Культуру клеток выращивают с соблюдением оптимальной температуры (36-38,5 °С) роста клеток и асептических условий в специальной лабораторной посуде (пробирки, флаконы, матрасы) или в реакторах для получения биотехнологической продукции. При этом используют сложную питательную среду Игла, среду 199 и другие среды, содержащие необходимые для роста клеток аминокислоты, минеральные соли, витамины, глюкозу, сыворотку крови животных или человека, буферные растворы для поддержания стабильного рН. Для подавления роста посторонней микрофлоры в среды для культивирования клеток добавляют антибиотики.
Различают однослойные, суспензионные и органные культуры клеток. Клетки однослойной культуры клеток прикрепляются и размножаются на поверхности лабораторной посуды в виде монослоя. Они получили наибольшее применение в вирусологии. Клетки суспензионных культур клеток размножаются во всем объеме питательной среды при постоянном ее перемешивании с помощью магнитной мешалки или вращающегося барабана. Метод применяют для получения большого количества клеток, например, при промышленном получении вирусных вакцин. Органные культуры применяются ограниченно. Они представляют собой цельные кусочки органов и тканей, сохраняющие при культивировании исходную структуру.
По свойствам жизнеспособных генераций культуры клеток подразделяют на первичные, или первично-трипсинизированные, перевиваемые, или стабильные и полуперививаемые.
Первичные культуры клеток размножаются только в первых генерациях, т.е. выдерживают не более 5-10 пассажей после выделения из тканей. Их получают при обработке кусочков тканей (эмбриональных, опухолевых или нормальных) протеолитическими ферментами, которые разрушают межклеточные связи в тканях и органах с образованием изолированных клеток.
Перевиваемые, или стабильные, культуры клеток способны размножаться в лабораторных условиях неопределенно длительный срок (десятки лет), т.е. выдерживают многочисленные пассажи. Их получают преимущественно из опухолевых или эмбриональных тканей, обладающих большой потенцией роста. Перевиваемые
культуры клеток имеют преимущества перед первичными культурами: продолжительность их культивирования, высокая скорость размножения опухолевых и эмбриональных клеток, меньшая трудоемкость, способность культур сохранять свои свойства в замороженном состоянии в течение многих лет, возможность использования международных линий культур во многих лабораториях мира. Однако злокачественный характер клеток и соматические мутации, претерпеваемые нормальными клетками в процессе многочисленных генераций, ограничивают использование этого вида культур, в частности их нельзя применять в производстве вирусных вакцин.
Полуперививаемые культуры клеток имеют ограниченную продолжительность жизни и выдерживают 40-50 пассажей. Их обычно получают из диплоидных клеток эмбриона человека. В процессе пассажей эти культуры сохраняют диплоидный набор хромосом, характерный для соматических клеток исходной ткани, и не претерпевают злокачественную трансформацию. Поэтому полуперививаемые культуры клеток могут быть использованы как в диагностике, так и в производстве вакцин. О репродукции вирусов в культуре клеток, зараженных вируссодержащим материалом, можно судить на основании следующих феноменов: цитопатогенного действия (ЦПД) вирусов, или цитопатического эффекта (ЦПЭ), образования внутриклеточных включений, образования бляшек, реакций гемадсорбции и гемагглютинации; цветной реакции.
ЦПД, или ЦПЭ, - видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения от стекла), возникающие в результате внутриклеточной репродукции вирусов. В зависимости от особенностей репродуцирующихся вирусов ЦПД может различаться. В одних случаях быстро вакуолизируется цитоплазма, разрушаются митохондрии, округляются и гибнут клетки, в других формируются гигантские многоядерные клетки (так называемые симпласты) или наблюдается явление клеточной пролиферации, которое в итоге заканчивается деструкцией клеток. Таким образом, характер ЦПД позволяет использовать этот феномен не только для индикации вирусов, но и для их ориентировочной идентификации в культуре клеток. Другим проявлением ЦПЭ является образование внутриклеточных включений в ядре или цитоплазме зараженных клеток. Часто включения представляют собой скопления вирионов или их компонентов, иногда они могут содержать клеточный материал. Выявляют включения с помощью
светового или люминесцентного микроскопа после окрашивания зараженных клеток соответственно анилиновыми красителями или флюорохромами. Включения могут отличаться по величине (от 0,2 до 25 мкм), форме (округлые или неправильные) и численности (одиночные и множественные). Характерные цитоплазматические включения формируются в клетках, инфицированных вирусом натуральной оспы (тельца Гварниери), бешенства (тельца Бабеша-Негри), а внутриядерные включения - при заражении аденовирусами или вирусами герпеса.
Бляшки, или негативные, колонии представляют собой ограниченные участки разрушенных вирусами клеток в сплошном монослое культур клеток (рис. 3.8). Они видны невооруженным глазом в виде светлых пятен на фоне окрашенного монослоя живых клеток. Добавление агара в питательную среду ограничивает распространение вирусов по всему монослою после выхода из разрушенной клетки и обеспечивает взаимодействие вирусов только с соседними клетками. Каждая бляшка образуется потомством одного вириона. Подсчитав количество бляшек, можно определить концентрацию вирусов в исследуемом материале. Кроме того, бляшки разных групп вирусов отличаются по размеру, форме, срокам появления. Поэтому метод бляшек используют для дифференциации вирусов,
 Рис. 3.8. Образование «бляшек» в культуре клеток, зараженных вирусом
Рис. 3.8. Образование «бляшек» в культуре клеток, зараженных вирусом
а также для селекции штаммов и получения чистых линий вирусов. Однако не все вирусы могут вызывать цитопатический эффект, тогда их выявляют другими методами.
В основе реакции гемадсорбции лежит способность культур клеток, инфицированных вирусами, адсорбировать на своей поверхности эритроциты. Целый ряд вирусов (гриппа, парагриппа и др.) обладают гемадсорбирующими свойствами, что позволяет использовать реакцию гемадсорбции для индикации этих вирусов даже при отсутствии выраженного ЦПД в культуре клеток. Механизмы реакции гемадсорбции и гемагглютинации сходны. Поэтому для обнаружения репродукции некоторых вирусов в культуре клеток можно использовать реакцию гемагглютинации с культуральной жидкостью, т.е. с питательной средой, содержащей размножившиеся вирусы.
Присутствие в культуре клеток популяции вирусов можно также выявить с помощью цветной реакции, которая регистрируется по цвету индикатора питательной среды для культур клеток. При репродукции вирусов нормальный метаболизм клеток нарушается (клетки гибнут) и среда сохраняет свой первоначальный цвет. Если же вирусы не размножаются в культуре клеток, то клетки, оставаясь жизнеспособными, в процессе метаболизма выделяют кислые продукты, изменяющие рН среды и соответственно цвета индикатора.
3.4. Бактериофаги (вирусы бактерий)
Бактериофаги (от «бактерия» и греч. phagos - пожирающий) - вирусы, специфически проникающие в бактерии, использующие их биосинтетические системы для своей репродукции и вызывающие их лизис (растворение, разрушение клеток). Впервые явление самопроизвольного лизиса сибиреязвенных бактерий наблюдал один из основоположников отечественной микробиологии Н.Ф. Гамалея (1898). Английский бактериолог Ф. Туорт (1915) описал способность фильтрата стафилококков растворять свежую культуру этих же бактерий. Однако лишь французский ученый Ф. д'Эрелль (1917) правильно оценил это явление, выделив фильтрующийся литический агент из испражнений больных дизентерией. Добавление литического агента к мутной бульонной культуре дизентерийных бактерий приводило к полному просветлению среды. Ана-
Физиология микробов
121
логичный эффект д'Эррель наблюдал и на плотных питательных средах, засеянных смесью литического агента с соответствующими бактериями. На фоне сплошного бактериального роста появлялись стерильные пятна круглой или неправильной формы - участки лизиса бактерий, названные негативными колониями, или бляшками. Предположив, что имеет дело с вирусами, д'Эрелль выделил этот литический агент с помощью бактериальных фильтров и назвал его бактериофагом - пожирателем бактерий.
Бактериофаги широко распространены в природе. Они обнаружены в воде, почве, пищевых продуктах, различных выделениях из организма людей и животных (фекалии, моча, мокрота, гной и т.д.). Особенно большое количество бактериофагов выделяется в период выздоровления больного человека. В настоящее время эти вирусы выявлены у большинства бактерий, а также у некоторых других микроорганизмов, в частности у грибов. Поэтому бактериофаги в широком смысле слова часто называют просто фагами.
Бактериофаги принято обозначать буквами латинского, греческого или русского алфавита, часто с цифровым индексом, перед которым стоит название вида бактерий (например, фаги Е. coli T2). Для обозначения группы родственных фагов используют родовые и видовые названия микробов, из которых выделены соответствующие фаги: колифаги, стафилофаги, актинофаги, микофаги и т.д.
Морфология и химический состав. Морфологию бактериофагов изучают с помощью электронной микроскопии. Фаги, как и просто организованные вирусы человека, состоят из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки - капсида. Однако между собой они в значительной степени различаются по морфологии. В зависимости от формы, структурной организации и типа нуклеиновой кислоты фаги подразделяют на несколько морфологических типов (рис. 3.9). К I типу относятся нитевидные ДНК-содержащие фаги, взаимодействующие с мужскими особями бактерий (см. раздел 2.2 и главу 5). Геном фагов представлен однонитевой ДНК, заключенной в спиральный капсид. II тип включает мелкие РНК-содержащие и однонитевые ДНК-содержащие фаги, геном которых находится внутри икосаэдрического капсида (головки) с аналогом отростка. К III типу относятся икосаэдрические фаги с коротким отростком, содержащие двунитевую ДНК. IV и V типы - сложные по морфологии ДНК-содержащие фаги, имеющие форму сперматозоида: икосаэдрический капсид головки
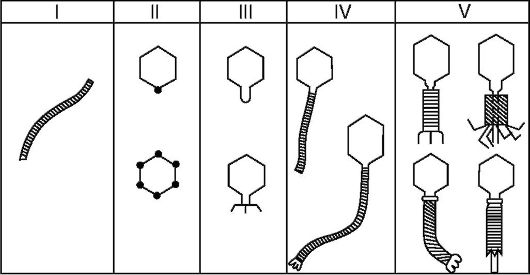 Рис. 3.9. Морфологические типы бактериофагов (объяснение в тексте)
Рис. 3.9. Морфологические типы бактериофагов (объяснение в тексте)
соединен с длинным хвостовым отростком. V тип фагов отличается от VI типа тем, что чехол их отростков способен к сокращению. Размеры фагов колеблются от 20 до 800 нм (нитевидный тип).
 Рис. 3.10. Строение Т-четного фага (электронограмма)
Рис. 3.10. Строение Т-четного фага (электронограмма)
Наиболее изучены крупные бактериофаги, имеющие форму сперматозоида и сокращающийся чехол отростка (рис. 3.10), например колифаги T2, Т4, Т6 (от англ. type - типовые). У этих фагов молекула двунитевой суперспирализованной ДНК находится внутри головки размером 65-100 нм и защищена капсидом. Капсид состоит из белковых молекул - идентичных полипептидных субъединиц, уложенных по икосаэдрическому (кубическому) типу симметрии. В состав головки также входит полипептид, состоящий из аспарагиновой, глутаминовой кислот и лизина. У некоторых фагов внутри головки находится внутренний гистоноподобный белок, обеспечивающий суперспирализа-
цию ДНК. Хвостовой отросток длиной более 100 нм имеет внутри полый цилиндрический стержень, сообщающийся с головкой, а снаружи - чехол (футляр), способный к сокращению наподобие мышцы. Чехол хвостового отростка образован белковыми субъединицами, уложенными по спиральному типу симметрии, содержит АТФ и ионы Ca. На дистальном конце отростка имеется шестиугольная базальная пластинка с шипами, от которых отходят нитевидные структуры - фибриллы.
У некоторых фагов (например, T2) в дистальной части отростка содержится фермент лизоцим.
Антигенные свойства. Бактериофаги содержат группоспецифические и типоспецифические антигены, обладают иммуногенными свойствами, вызывая синтез специфических антител в организме. Антитела, взаимодействуя с бактериофагами, могут нейтрализовать их литическую активность в отношении бактерий. По типоспецифическим антигенам фаги делят на серотипы.
Резистентность. По сравнению с вирусами человека бактериофаги более устойчивы к факторам окружающей среды. Они инактивируются под действием температуры 65-70 °С, УФ-облучения в высоких дозах, ионизирующей радиации, формалина и кислот. Длительно сохраняются при низкой температуре и высушивании.
Взаимодействие фагов с бактериальной клеткой. Взаимодействие фагов с бактериями может протекать, как и у других вирусов, по продуктивному, абортивному и интегративному типам. При продуктивном типе взаимодействия образуется фаговое потомство, бактерии лизируются; при абортивном типе фаговое потомство не образуется и бактерии сохраняют свою жизнедеятельность, при интегративном типе геном фага встраивается в хромосому бактерии и сосуществует с ней. В зависимости от типа взаимодействия различают вирулентные и умеренные бактериофаги.
Вирулентные бактериофаги взаимодействуют с бактерией по продуктивному типу. Проникнув в бактерию, они репродуцируются с образованием 200-300 новых фаговых частиц и вызывают лизис бактерий. Процесс взаимодействия с бактериями в достаточной мере изучен у бактериофагов, имеющих отросток с сокращающимся чехлом. Он состоит из последовательно сменяющих друг друга стадий и весьма схож с процессом взаимодействия вирусов человека и животных с клеткой хозяина. Однако имеются и некоторые особенности.
Специфическая адсорбция фагов происходит только при соответствии прикрепительных белков вирусов и рецепторов бактериальной клетки липополисахаридной или липопротеиновой природы, находящихся в ее клеточной стенке. На бактериях, лишенных клеточной стенки (протопласты, сферопласты), бактериофаги не могут адсорбироваться. Фаги, имеющие хвостовой отросток, прикрепляются к бактериальной клетке свободным концом отростка (фибриллами базальной пластинки). В результате активации АТФ чехол хвостового отростка сокращается и стержень с помощью лизоцима, растворяющего прилегающий фрагмент клеточной стенки, как бы просверливает оболочку клетки. При этом ДНК фага, содержащаяся в его головке, проходит в форме нити через канал хвостового стержня и инъецируется в клетку, а капсидные оболочки фага остаются снаружи бактерии.
Инъецированная внутрь бактерии нуклеиновая кислота подавляет биосинтез компонентов клетки, заставляя ее синтезировать нуклеиновую кислоту и белки фага. Процесс синтеза вирусных белков и репликация фаговых геномов в бактериальной клетке аналогичны процессу репродукции других вирусов, содержащих двунитевую ДНК. РНК-полимераза клетки транскрибирует некоторые гены фаговой ДНК, в результате чего образуются ранние иРНК. Рибосомы клетки транслируют иРНК, при этом синтезируется целый ряд ферментов, включая те, которые необходимы для репликации фаговой ДНК. Репликация двунитевой ДНК фагов протекает в соответствии с общим механизмом репликации. После начала репликации фаговой ДНК начинается синтез поздних вирусных иРНК, в результате трансляции которых образуется второй набор вирусспецифических белков, в том числе капсидных белков фагов.
После образования компонентов фага происходит самосборка частиц: сначала пустотелые капсиды головок заполняются нуклеиновой кислотой, затем сформированные головки соединяются с хвостовыми отростками. При литической инфекции в клетке появляется еще один поздний вирусспецифический белок - фаговый лизоцим. Этот фермент воздействует на пептидогликановый слой стенки бактерии, делая ее менее прочной. В конце концов под действием внутриклеточного осмотического давления оболочка клетки разрывается и фаговое потомство выходит в окружающую среду вместе с остальным содержимым бактериальной клетки. Весь литический цикл от адсорбции бактериофага на бактерии до его выхода из нее занимает 20-40 мин.
У некоторых фагов механизм адсорбции, проникновения и высвобождения из клеток совершенно иной. Например, у нитевидных фагов на концах капсидной оболочки имеются минорные белки, с помощью которых эти фаги прикрепляются к половым пилям бактерии (см. главу 5). Фаговая ДНК вместе с минорным белком проникают в цитоплазму клетки через ее половые пили. После репликации нуклеиновой кислоты фагов вновь синтезированные белки фаговой оболочки располагаются на клеточной мембране. Сборка и высвобождение нитевидных фагов происходят путем просачивания ДНК через цитоплазматическую мембрану и клеточную стенку бактерии, во время которого они приобретают белковые капсиды. Бактериальная клетка при этом сохраняет свою жизнеспособность.
Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности, что явилось основанием для подразделения их на поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными типами (вариантами) данного вида бактерий.
Умеренные бактериофаги, в отличие от вирулентных, взаимодействуют с чувствительными бактериями либо по продуктивному, либо по интегративному типу. Продуктивный цикл умеренного фага идет в той же последовательности, что и у вирулентных фагов, и заканчивается лизисом клетки. При интегративном типе взаимодействия ДНК умеренного фага встраивается в хромосому бактерии, причем в строго определенную гомологическую область хромосомы, реплицируется синхронно с геномом размножающейся бактерии, не вызывая ее лизиса. ДНК бактериофага, встроенная в хромосому бактерии, называется профагом, а культура бактерий, содержащих профаг, - лизогенной. Само же биологическое явление сосуществования бактерии и умеренного бактериофага носит название лизогении (от греч. lysis - разложение, genea - происхождение). Профаг, ставший частью хромосомы размножающейся бактерии, передается по наследству от клетки к клетке неограниченному числу потомков.
Лизогенные бактерии не образуют структурные вирусные белки и, следовательно, фаговое потомство. В основе сдерживающего механизма репродукции фагов лежит образование в бактерии специфического репрессора -- низкомолекулярного белка, по-
давляющего траскрипцию фаговых генов. Биосинтез репрессора детерминируется генами профага. Наличием репрессора можно объяснить способность лизогенных бактерий прибретать иммунитет (невосприимчивость) к последующему заражению гомологичными или близкородственными фагами. Под иммунитетом в данном случае понимается такое состояние бактерии, при котором исключаются процесс вегетативного размножения вышеуказанных фагов и лизис клетки. Однако термин «лизогения» отражает потенциальную возможность лизиса бактерии, содержащей профаг. Действительно, профаги некоторой части лизогенной культуры бактерий могут спонтанно (самопроизвольно) или направленно под действием ряда физических или химических факторов дерепрессироваться, исключаться из хромосомы и переходить в вегетативное состояние. Этот процесс заканчивается продукцией фагов и лизисом бактерий. Частота спонтанного лизиса бактерий в лизогенных культурах весьма незначительна. Частоту лизиса бактерий можно значительно увеличить, воздействуя на лизогенную культуру индуцирующими агентами: УФ-лучами, ионизирующим излучением, перекисными соединениями, митомицином С и др. Сам же феномен воздействия, приводящий к инактивации репрессора, называется индукцией профага. Явление индукции используют в генетической инженерии. Однако спонтанный лизис лизогенных культур может нанести вред микробиологическому производству. Так, если микроорганизмы - продуценты биологически активных веществ - оказываются лизогенными, существует опасность перехода фага в вегетативное состояние, что приведет к лизису производственного штамма этого микроба.
Геном профага может придавать бактерии новые, ранее отсутствовавшие у нее свойства. Этот феномен изменения свойств микроорганизмов под влиянием профага получил название фаговой конверсии (от лат. conversion - превращение). Конвертироваться могут морфологические, культуральные, биохимические, антигенные и другие свойства бактерий. Например, только лизогенные культуры дифтерийной палочки способны вызвать болезнь (дифтерию), так как содержат в хромосоме профаг, ответственный за синтез белкового экзотоксина.
Умеренные фаги могут быть дефектными, т.е. неспособными образовывать зрелые фаговые частицы ни в естественных усло-
виях, ни при индукции. Геном некоторых умеренных фагов (Р1) может находиться в цитоплазме бактериальной клетки в так называемой плазмидной форме, не включаясь в ее хромосому. Такого рода умеренные фаги используют в качестве векторов в генетической инженерии.
Практическое применение фагов. Бактериофаги используют в лабораторной диагностике инфекций при внутривидовой идентификации бактерий, т.е. определении фаговара (фаготипа). Для этого применяют метод фаготипирования, основанный на строгой специфичности действия фагов: на чашку Петри с плотной питательной средой, засеянной «газоном» чистой культурой возбудителя, наносят капли различных диагностических типоспецифических фагов. Фаговар бактерии определяется тем типом фага, который вызвал ее лизис (образование стерильного пятна, бляшки или негативной колонии). Метод фаготипирования позволяет выявить источник инфекции и проследить путь возбудителя от источника до восприимчивого организма (эпидемиологическое маркирование).
По содержанию бактериофагов в объектах окружающей среды можно судить о присутствии в них соответствующих патогенных бактерий. Подобные исследования проводят при санитарномикробиологическом исследовании воды. Например, в системах из поверхностных источников воды перед подачей ее в распределительную сеть определяют наличие колифагов. Колифаги являются одними из санитарно-показательных микробов, характеризующих фекальное загрязнение воды.
Фаги применяют также для лечения и профилактики ряда бактериальных, чаще всего кишечных инфекций. Производят брюшнотифозный, дизентерийный, синегнойный, стафилококковый фаги и комбинированные препараты (колипротейный, пиобактериофаги и др.). Бактериофаги назначают по показаниям перорально, парентерально или местно в виде жидких, таблетированных форм, свечей или аэрозолей. Отличительной чертой фагов является полное отсутствие у них побочного действия. Однако лечебный и профилактический эффект фагов умеренный, поэтому их необходимо применять в комплексе с другими лечебными и профилактическими мероприятиями. Бактериофаги широко применяют в генетической инженерии в качестве векторов для получения рекомбинантных ДНК.
Задания для самоподготовки (самоконтроля)
A. Назовите процесс, при котором бактерии получают энергию путем ферментации глюкозы:
1. Гниение.
2. Брожение.
3. Денитрофикация.
4. Анаэробное дыхание.
Б. Большинство болезнетворных бактерий, называемых мезофилами, растут при температуре:
1. 15-20 °С.
2. 20-30 °С.
3. 30-37 °С
4. 50-55 °С.
B. Назовите процесс, при котором в присутствии кислорода происходит минерализация белка:
1. Денитрофикация.
2. Брожение.
3. Лиофилизация.
4. Гниение.
Г. Назовите механизм, который используется бактериями для доставки внутрь цитозоля клетки эффекторных молекул:
1. Активный транспорт.
2. Секреция по III типу.
3. Секреция по II типу.
4. Транслокация радикалов.
Д. Некоторые вирусы в составе своих вирионов имеют РНКзависимую РНК-полимеразу. Назовите тип нуклеиновой кислоты этих вирусов:
1. Двунитевая ДНК кольцевой формы.
2. Плюс-однонитевая РНК.
3. Минус-однонитевая РНК.
4. Двунитевая ДНК линейная.
Е. Назовите последствия интегративного типа взаимодействия вируса и клетки:
1. Вирусоносительство.
2. Трансформация клетки.
3. Гибель клетки.
4. Образование нового поколения вирионов.
Ж. Взвесь культуры E. coli была засеяна в 2 колбы, одна из которых содержала среду ? 1, а другая - среду ? 2. Посевы были поставлены в термостат с температурой 37 °С. Каждый час отбирали пробы для определения плотности бактериальной популяции, на основании чего были построены кривые роста, которые показали, что продолжительность лаг-фазы в среде ? 1 равнялась 20 мин, а в среде ? 2 - 50 мин. Назовите более эффективную среду.
З. На 3 чашки с кровяным агаром был произведен посев 4 бактериальных культур: А, Б, В, Г. Чашка ? 1 была поставлена в термостат с температурой 37 °С. Чашка ? 2 была помещена в анаэростат, из которого откачали воздух, и поставили в термостат с температурой 37 °С. Чашка ? 3 была поставлена в СО2-инкубатор с температурой 37 °С. Через сутки инкубации были получены следующие результаты. Бактериальная культура А выросла на всех 3 чашках. Бактериальная культура Б выросла только на чашке ? 3 (культивирование в атмосфере 5% СО2). Бактериальная культура В выросла только на чашке ? 1. Бактериальная культура Г выросла только на чашке ? 2. Охарактеризуйте каждый тип культур. Ответ обоснуйте.