Медицинская микробиология, вирусология и иммунология: том 1 : учебник / Под ред. В.В. Зверева, М.Н. Бойченко, 2010.
|
|
|
|
ГЛАВА 10 АНТИГЕНЫ И ИММУННАЯ СИСТЕМА ЧЕЛОВЕКА
10.1. Антигены
10.1.1. Общие сведения
Жизнедеятельность каждого макроорганизма проходит в непосредственном контакте с чужеродными для него клетками, доклеточными формами жизни и отдельными биоорганическими молекулами. Будучи чужеродными, эти объекты таят в себе огромную опасность, так как могут нарушить гомеостаз, повлиять на течение биологических процессов в макроорганизме и даже повлечь его гибель. Контакт с чужеродными биологическими объектами представляет собой ранний сигнал опасности для иммунной системы, они являются основным раздражителем и объектом системы приобретенного иммунитета. Такие объекты получили название антигенов (от греч. anti - против, genos - создавать).
Современное определение термина «антиген» - это биополимер органической природы, генетически чужеродный для макроорганизма, который при попадании в последний распознается его иммунной системой и вызывает иммунные реакции, направленные на его устранение. Учение об антигенах является ключевым для понимания основ молекулярно-генетических механизмов иммунной защиты макроорганизма, так как антиген является движущей силой иммунного ответа, а также принципов иммунотерапии и иммунопрофилактики.
Антигены имеют разнообразное происхождение. Они являются продуктом природного биологического синтеза любого чужеродного организма, могут образовываться в собственном организме при структурных изменениях уже синтезированных молекул в ходе биодеградации, нарушении их нормального биосинтеза или генетической мутации клеток. Кроме того, антигены могут быть по-
лучены искусственно в результате научной работы или путем направленного химического синтеза. Однако в любом случае молекулу антигена будет отличать генетическая чужеродность по отношению к макроорганизму, в который она попала. Теоретически антигеном может быть молекула любого органического соединения.
Антигены могут попадать в макроорганизм самыми разными путями: через кожные покровы или слизистые оболочки, непосредственно во внутреннюю среду организма, минуя покровы или образовываясь внутри него. При попадании в макроорганизм антигены распознаются иммунокомпетентными клетками и вызывают каскад разнообразных иммунных реакций, направленных на их инактивацию, разрушение и удаление.
10.1.2. Свойства антигенов
Характерными свойствами антигенов являются антигенность, иммуногенность и специфичность.
Антигенность - это потенциальная способность молекулы антигена активировать компоненты иммунной системы и специфически взаимодействовать с факторами иммунитета (антитела, клон эффекторных лимфоцитов). При этом компоненты иммунной системы взаимодействуют не со всей молекулой антигена, а только с ее небольшим участком, который получил название антигенной детерминанты, или эпитопа.
Различают линейные, или секвенциальные, антигенные детерминанты, например первичная аминокислотная последовательность пептидной цепи, и поверхностные, или конформационные, расположенные на поверхности молекулы антигена и возникшие в результате вторичной или более высокой конформации. На концевых участках молекулы антигена расположены концевые эпитопы, а в центре молекулы - центральные. Существуют также глубинные, или скрытые, антигенные детерминанты, которые проявляются при разрушении биополимера.
Размер антигенной детерминанты невелик. Он определяется характеристиками рецепторной части фактора иммунитета и структурой эпитопа. Например, антигенсвязывающий участок молекулы иммуноглобулина способен распознать линейную антигенную детерминанту, состоящую из 5 аминокислотных остатков. Для образования конформационной детерминанты требуется 6-12 аминокислотных остатков. Рецепторному аппарату Т-киллера для
определения чужеродности требуется нанопептид, включенный в состав MHC I класса, Т-хелперу - олигопептид размером 12- 25 аминокислотньгх остатков в комплексе с MHC II класса.
Молекулы большинства антигенов имеют довольно большие размеры. В их структуре определяется множество антигенных детерминант, которые распознаются разными по специфичности антителами и клонами лимфоцитов. Поэтому антигенность вещества зависит от наличия и числа антигенных детерминант в структуре его молекулы.
Структура и состав эпитопа имеют критическое значение. Замена хотя бы одного структурного компонента молекулы приводит к образованию принципиально новой антигенной детерминанты. Денатурация приводит к потере имеющихся антигенных детерминант или появлению новых, а также специфичности.
Чужеродность является обязательным условием для реализации антигенности. Понятие «чужеродность» относительное, так как иммунокомпетентные клетки не способны напрямую анализировать чужеродный генетический код, а лишь продукты, синтезированные с чужеродной генетической матрицы. В норме иммунная система невосприимчива к собственным биополимерам, если он не приобрел черты чужеродности. Кроме того, при некоторых патологических состояниях в результате нарушения регуляции иммунного ответа (см. аутоантигены, аутоантитела, аутоиммунитет, аутоиммунные болезни) собственные биополимеры могут восприниматься иммунной системой как чужие.
Чужеродность находится в прямой зависимости от эволюционного расстояния между организмом и источником антигенов. Чем дальше в таксономическом плане организмы отстоят друг от друга, тем большей чужеродностью и, следовательно, иммуногенностью обладают их антигены. Чужеродность заметно проявляется даже между особями одного вида, так как замена хотя бы одной аминокислоты эффективно распознается антителами в серологических реакциях.
Вместе с тем антигенные детерминанты даже генетически неродственных существ или веществ могут иметь определенное подобие и способны специфически взаимодействовать с одними и теми же факторами иммунитета. Такие антигены получили название перекрестно реагирующих. Обнаружено также сходство антигенных детерминант стрептококка, сарколеммы миокарда и базальной
мембраны почек, Treponema pallidum и липидной вытяжки из миокарда крупного рогатого скота, возбудителя чумы и эритроцитов человека 0(I) группы крови. Явление, когда один организм маскируется антигенами другого для защиты от факторов иммунитета, получило название антигенной мимикрии.
10.1.2.1. Иммуногенность
Иммуногенность - потенциальная способность антигена вызывать по отношению к себе в макроорганизме специфический продуктивный ответ. Иммуногенность зависит от трех групп факторов: молекулярных особенностей антигена, кинетики антигена в организме, реактивности макроорганизма.
К первой группе факторов отнесены природа, химический состав, молекулярная масса, структура и некоторые другие характеристики.
Природа антигена в значительной степени определяет иммуногенность. Наиболее выраженной иммуногенностью обладают белки и полисахариды, наименьшей - нуклеиновые кислоты и липиды. В то же время их сополимеры - липополисахариды, гликопротеиды, липопротеиды - способны в достаточной мере активировать иммунную систему.
Иммуногенность в определенной мере зависит от химического состава молекулы антигена. Для белковых антигенов важно разнообразие их аминокислотного состава. Монотонные полипептиды, построенные из одной аминокислоты, практически не активируют иммунную систему. Наличие в структуре молекулы белка ароматических аминокислот, таких, как тирозин, триптофан, существенно повышает иммуногенность.
Важна оптическая изомерия структурных компонентов молекулы антигена. Пептиды, построенные из L-аминокислот, высокоиммуногенны. Полипептидная цепочка, построенная из правовращающих изомеров аминокислот, напротив, может проявлять ограниченную иммуногенность при введении в малых дозах.
В спектре иммуногенности существует определенная иерархия антигенных детерминант: эпитопы различаются по способности индуцировать иммунный ответ. При иммунизации некоторым антигеном будут преобладать реакции к отдельным антигенным детерминантам. Это явление получило название иммунодоминантности. По современным представлениям она обусловлена различиями в сродстве эпитопов к рецепторам антигенпрезентирующих клеток.
Большое значение имеют размер и молекулярная масса антигена. Небольшие полипептидные молекулы с массой менее 5 кД, как правило, низкоиммуногенны. Олигопептид, способный индуцировать иммунный ответ, должен состоять из 6-12 аминокислотных остатков и иметь молекулярную массу около 450 Д. С увеличением размера пептида возрастает его иммуногенность, однако эта зависимость на практике не всегда выполняется. Так, при равной молекулярной массе (около 70 кД) альбумин является более сильным антигеном, чем гемоглобин.
Опытным путем было доказано, что высокодисперсные коллоидные растворы антигена плохо индуцируют иммунный ответ. Гораздо большей иммуногенностью обладают агрегаты молекул и корпускулярные антигены - цельные клетки (эритроциты, бактерии и т.д.). Это связано с тем, что корпускулярные и высокоагрегированные антигены лучше фагоцитируются, чем отдельные молекулы.
Оказалась также существенной стерическая стабильность молекулы антигена. При денатурации белков до желатина вместе с конформационной жесткостью теряется иммуногенность. Поэтому растворы желатина широко используются для парентерального введения.
Важным условием иммуногенности является растворимость антигена. Например, высокомолекулярные соединения кератин, меланин, натуральный шелк и др. нерастворимы в воде, не образуют коллоидных растворов в нормальном состоянии и не являются иммуногенами. Благодаря этому свойству конский волос, шелк, кетгут и др. прменяют в клинической практике для сшивания органов и тканей.
Вторая группа факторов связана с динамикой поступления антигена в организм и его выведения. Так, хорошо известна зависимость иммуногенности антигена от места и способа его введения, что обусловлено особенностями строения иммунной системы в местах интервенции антигена.
Сила иммунного ответа зависит от количества поступающего антигена: чем его больше, тем выраженнее иммунная реакция макроорганизма.
Третья группа объединяет факторы, определяющие зависимость иммуногенности от состояния макроорганизма: наследственности и функциональных характеристик. Хорошо известно, что резуль-
тат иммунизации в определенной мере связан с генотипом особи. Существуют чувствительные и нечувствительные к определенным антигенам роды и виды животных. Например, кролики и крысы практически не реагируют на некоторые бактериальные антигены, которые могут вызывать у морской свинки или мыши чрезвычайно бурный иммунный ответ.
10.1.2.2. Специфичность
Специфичностью называют способность антигена индуцировать иммунный ответ к строго определенному эпитопу. Специфичность антигена во многом определяется свойствами составляющих его эпитопов.
10.1.3. Классификация антигенов
Основываясь на отдельных характерных свойствах, все многообразие антигенов можно классифицировать по происхождению, природе, молекулярной структуре, степени иммуногенности, степени чужеродности, направленности активации и обеспеченности иммунного реагирования.
По происхождению различают экзогенные (возникшие вне организма) и эндогенные (возникшие внутри организма) антигены. Среди эндогенных особого внимания заслуживают ауто- и неоантигены. Аутогенные антигены (аутоантигены) - это структурно неизмененные антигены собственного организма, синтезируемые в организме в физиологических условиях. В норме аутоантигены неиммуногенны вследствие сформировавшейся иммунологической толерантности (невосприимчивости) либо их недоступности для контакта с факторами иммунитета - это так называемые забарьерные антигены. При срыве толерантности или нарушении целостности биологических барьеров (воспаление, травма) компоненты иммунной системы начинают специфически реагировать на аутоантигены выработкой специфических факторов иммунитета (аутоантитела, клон аутореактивных лимфоцитов). Неоантигены, в отличие от аутоантигенов, возникают в организме в результате генетических мутаций или модификаций и всегда чужеродны.
По природе: биополимеры белковой (протеиды) и небелковой (полисахариды, липиды, липополисахариды, нуклеиновые кислоты и др.) природы.
По молекулярной структуре: глобулярные (молекула имеет шаровидную форму) и фибриллярные (форма нити).
По степени иммуногенности: полноценные и неполноценные. Полноценные антигены обладают выраженной антигенностью и иммуногенностью - иммунная система чувствительного организма реагирует на их введение выработкой факторов иммунитета. Такие вещества, как правило, имеют достаточно большую молекулярную массу (более 10 кД), большой размер молекулы (частицы) в виде глобулы и хорошо взаимодействуют с факторами иммунитета.
Неполноценные антигены, или гаптены (термин предложен К. Ландштейнером), обладают антигенностью - способны специфически взаимодействовать с уже готовыми факторами иммунитета (антителами, лимфоцитами), но не способны при введении в нормальных условиях индуцировать в организме иммунный ответ. Чаще всего гаптенами являются низкомолекулярные соединения (молекулярная масса менее 10 кД).
Если искусственно укрупнить молекулу гаптена - соединить ее прочной связью с достаточно большой белковой молекулой, удается заставить иммунную систему макроорганизма специфически реагировать на гаптен как на полноценный антиген и вырабатывать факторы иммунитета. Молекула белка-носителя получила название шлеппера (тягача). При этом специфичность в составе молекулы конъюгата определяется гаптенной частью, а иммуногенность - белком-носителем. Используя для иммунизации конъюгаты, получают антитела к гормонам, лекарственным препаратам и другим низкоиммуногенным соединениям.
По степени чужеродности: ксено-, алло- и изоантигены. Ксеногенные антигены (или гетерологичные) - общие для организмов, стоящих на разных ступенях эволюционного развития, например, относящиеся к разным родам и видам. Впервые феномен общности ряда антигенов у животных разных видов был отмечен Д. Форсманом (1911 г.). При иммунизации кролика суспензией органов морской свинки ученый получил иммунную сыворотку, способную взаимодействовать с эритроцитами барана. Позже было установлено, что морская свинка и баран имеют ряд структурно сходных антигенных детерминант, дающих перекрестное реагирование. В дальнейшем перечень подобных ксеногенных антигенов был значительно расширен и они получили обобщенное название «антигены Форсмана».
Аллогенные антигены (или групповые) - общие для генетически неродственных организмов, но относящихся к одному виду. На основании аллоантигенов общую популяцию организмов можно подразделить на отдельные группы. Примером таких антигенов у людей являются антигены групп крови (системы АВ0 и др.). Аллогенные ткани при трансплантации иммунологически несовместимы - они отторгаются или лизируются реципиентом. Микробы на основании групповых антигенов могут быть подразделены на серогруппы, что используется в микробиологической диагностике.
Изогенные антигены (или индивидуальные) - общие только для генетически идентичных организмов, например для однояйцовых близнецов, инбредных линий животных. Изотрансплантаты обладают практически полной иммунной совместимостью и не отторгаются. К изоантигенам у людей относятся антигены гистосовместимости, а у бактерий - типовые антигены, не дающие дальнейшего расщепления.
В пределах отдельного организма в определенных органах или тканях обнаруживаются специфичные для них антигены, которые нигде больше не встречаются. Такие антигены получили название органо- и тканеспецифических.
В зависимости от физико-химических свойств антигена, условий его внедрения, характера реакции и реактивности макроорганизма различают иммуногены, толерогены и аллергены. Иммуногены способны индуцировать нормальную продуктивную реакцию иммунной системы - выработку факторов иммунитета (антитела, антигенореактивные клоны лимфоцитов). В клинической практике иммуногены используют для иммунодиагностики, иммунотерапии и иммунопрофилактики многих патологических состояний.
Толероген является полной противоположностью иммуногену. Он формирует иммунологическую толерантность или неотвечаемость на эпитопы данного вещества (см. раздел 11.6). Толероген, как правило, - мономер с низкой молекулярной массой, высокой эпитопной плотностью и высокой дисперсностью. Толерогены используют для профилактики и лечения иммунологических конфликтов и аллергии путем наведения искусственной неотвечаемости на отдельные антигены.
Аллерген, в отличие от иммуногена, формирует патологическую реакцию организма в виде гиперчувствительности немедленного или замедленного типа (см. раздел 11.4). По своим свойствам
аллерген не отличается от иммуногена. В клинической практике аллергены применяют для диагностики инфекционных и аллергических заболеваний.
По направленности активации и обеспеченности иммунного реагирования, т.е. необходимости вовлечения Т-лимфоцитов в индукцию иммунного ответа, выделяют Т-зависимые и Т-независимые антигены. Иммунная реакция в ответ на введение Т-зависимого антигена реализуется при обязательном участии Т-хелперов. К ним относится большая часть известных антигенов. Для развития иммунного ответа на Т-независимые антигены не требуется привлечение Т-хелперов. Эти антигены способны непосредственно стимулировать В-лимфоциты к антителопродукции, дифференцировке и пролиферации, а также вызывать иммунный ответ у бестимусных животных. Т-независимые антигены имеют относительно простое строение. Это крупные молекулы с молекулярной массой более 103 кД, поливалентны и имеют многочисленные однотипные эпитопы. Т-независимые антигены являются митогенами и поликлональными активаторами, например полимерный флагеллин (сократительный белок жгутиков бактерий), липополисахарид, туберкулин и др.
От Т-независимых антигенов следует отличать суперантигены. Это группа веществ, в основном микробного происхождения, которые могут неспецифически вызывать поликлональную реакцию. Молекула суперантигена способна вмешиваться в кооперацию антигенпрезентирующей клетки и Т-хелпера и формировать ложный сигнал распознавания чужеродной субстанции.
Суперантигены способны одновременно неспецифически активировать огромное количество иммунокомпетентных клеток (до 20% и более), вызывать гиперпродукцию цитокинов и низкоспецифичных иммуноглобулинов, массовую гибель лимфоцитов вследствие апоптоза и развитие вторичного функционального иммунодефицита. Свойства суперантигена обнаружены у стафилококкового энтеротоксина, белков вирусов Эпштейна-Барр, бешенства, ВИЧ и некоторых других микробных агентов.
10.1.4. Антигены организма человека
Начало изучению аллоантигенных свойств тканей было положено К. Ландштайнером, который в 1901 г. открыл систему групповых антигенов эритроцитов (АВ0). В организме человека
выделяют множество разнообразных антигенов. Они не только нужны для полноценного развития и функционирования всего организма в целом, но также несут важную информацию при клинико-лабораторной диагностике, определении иммунной совместимости органов и тканей в трансплантологии, а также в научных исследованиях. Наибольший медицинский интерес из числа аллогенных антигенов представляют антигены групп крови, среди изогенных - антигены гистосовместимости, а в группе органо- и тканеспецифических - раково-эмбриональные антигены.
10.1.4.1. Антигены групп крови человека
Антигены групп крови человека располагаются на цитоплазматической мембране клеток, но наиболее легко определяются на поверхности эритроцитов. Поэтому они получили название «эритроцитарные антигены». На сегодняшний день известно более 250 различных эритроцитарных антигенов. Однако наиболее важное клиническое значение имеют антигены системы АВ0 и Rh (резус-фактор): их необходимо учитывать при проведении переливания крови, пересадке органов и тканей, предупреждении и лечении иммуноконфликтных осложнений беременности и т.д.
Антигены системы АВ0 обнаруживаются в плазме крови, лимфе, секретах слизистых оболочек и других биологических жидкостях, но наиболее выражены на эритроцитах. Они синтезируются многими клетками организма, включая ядросодержащие предшественники эритроцитов, и свободно секретируются в межклеточное пространство. На мембране клеток эти антигены могут появиться либо как продукт клеточного биосинтеза, либо в результате сорбции из межклеточных жидкостей.
Антигены системы АВ0 представляют собой высокогликозилированные пептиды: 85% приходится на углеводную часть и 15% - на полипептидную. Пептидный компонент состоит из 15 аминокислотных остатков. Он постоянен для всех групп крови АВ0 и иммунологически инертен. Иммуногенность молукулы антигена системы АВ0 определяется его углеводной частью.
В системе антигенов АВ0 выделяют три варианта антигенов, различающихся по строению углеводной части: Н, А и В. Базовой молекулой является антиген Н, специфичность которого определяют три углеводных остатка. Антиген А имеет в структуре дополнительный четвертый углеводный остаток - N-ацетил-D-галактозу, а антиген В - D-галактозу. Антигены системы АВ0 имеют неза-
висимое аллельное наследование, что определяет наличие в популяции 4 групп крови: 0(I), А(II), B(III) и АВ(IV). Кроме того, антигены А и В имеют несколько аллотипов (например, А1, А2, А3... или В1, В2, В3...), которые встречаются в популяции людей с разной частотой.
Антигены системы АВ0 определяют в реакции агглютинации. Однако, учитывая высокий популяционный полиморфизм данной антигенной системы, перед гемотрансфузией обязательно проводят биологическую пробу на совместимость крови реципиента и донора. Ошибка в определении групповой принадлежности и переливание пациенту несовместимой по группе крови приводят к развитию острого внутрисосудистого гемолиза.
Другой важнейшей системой эритроцитарных антигенов является система резус-антигенов (Rh) или резус-факторов. Эти антигены синтезируются предшественниками эритроцитов и обнаруживаются главным образом на эритроцитах, так как они водонерастворимы. Резус-антиген представляет собой термолабильный липопротеид. Выделяют 6 разновидностей этого антигена. Генетическая информация о его строении закодирована в многочисленных аллелях трех сцепленных между собой локусов (D/d, C/c, E/e). В зависимости от наличия или отсутствия резус-антигена в популяции людей различают две группы: резус-положительных и резус-отрицательных индивидуумов.
Совпадение по резус-антигену важно не только при переливании крови, но также для течения и исхода беременности. При беременности резус-отрицательной матери резус-положительным плодом может развиться резус-конфликт. Это патологическое состояние связано с выработкой антирезусных антител, способных вызвать иммунологический конфликт: невынашивание беременности или желтуху новорожденного (внутрисосудистый иммунный лизис эритроцитов).
Вследствие того что плотность резус-антигена на мембране эритроцитов невысока и его молекула обладает слабой антигенностью, резус-фактор определяют на мембране эритроцитов в реакции непрямой агглютинации (реакция Кумбса).
10.1.4.2. Антигены гистосовместимости
На цитоплазматических мембранах практически всех клеток макроорганизма обнаруживаются антигены гистосовместимости. Большая часть из них относится к системе главного комплекса
гистосовместимости, или MHC (от англ. Main Hystocompatibility Complex). Установлено, что антигены гистосовместимости играют ключевую роль в осуществлении специфического распознавания «свой-чужой» и индукции приобретенного иммунного ответа, определяют совместимость органов и тканей при трансплантации в пределах одного вида и другие эффекты. Большая заслуга в изучении MHC принадлежит Дж. Доссе, П. Догерти, П. Гореру, Г. Снеллу, Р. Цинкернагелю, Р.В. Петрову, ставшими основоположниками иммуногенетики.
Впервые MHC был обнаружен в 60-х годах ХХ века в опытах на генетически чистых (инбредных) линиях мышей при попытке межлинейной пересадки опухолевых тканей (П. Горер, Г. Снелл). У мышей этот комплекс получил название Н-2 и был картирован в 17-й хромосоме.
У человека MHC был описан несколько позже в работах Дж. Доссе. Его обозначили как HLA (от англ. Human Leukocyte Antigen), так как он ассоциирован с лейкоцитами. Биосинтез HLA определяется генами, локализованными сразу в нескольких локусах короткого плеча 6-й хромосомы.
MHC имеет сложную структуру и высокую полиморфность. Антигены гистосовместимости представляют собой гликопротеины, прочно связанные с цитоплазматической мембраной клеток. Их отдельные фрагменты имеют структурное сходство с молекулами иммуноглобулинов и поэтому относятся к единому суперсемейству. Различают два основных класса молекул MHC (I и II), которые объединяют множество сходных по структуре антигенов, кодируемых множеством аллельных генов. На клетках индивидуума могут одновременно экспрессироваться не более двух разновидностей продуктов каждого гена MHC. MHC I класса индуцирует преимущественно клеточный иммунный ответ, а MHC II класса - гуморальный.
MHC I класса состоит из двух нековалентно связанных полипептидных цепей (α и β) с разной молекулярной массой (рис. 10.1). α-Цепь имеет внеклеточный участок с доменным строением (α1-, α2- и а3-домены), трансмембранный и цитоплазматический. β-Цепь представляет собой β2-микроглобулин, адгезированный на α,,-домен после экспрессии α-цепи на цитоплазматической мембране клетки. α1- и α2-Домены α-цепи формируют щель Бъеркмана - участок, ответственный за сорбцию и презентацию молекул
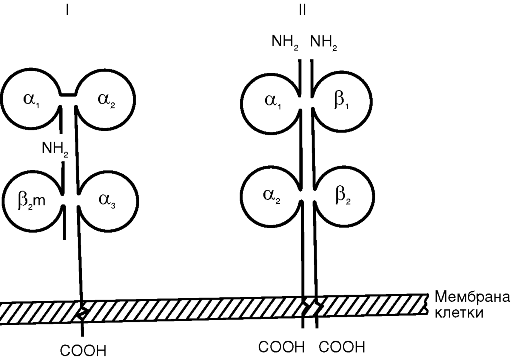 Рис. 10.1. Схема строения антигенов главного комплекса гистосовместимости: I - МНС I класса; II - МНС II класса
Рис. 10.1. Схема строения антигенов главного комплекса гистосовместимости: I - МНС I класса; II - МНС II класса
антигена. Щель Бъеркмана MHC I класса вмещает нанопептид, который легко выявляется специфическими антителами.
Сборка комплекса MHC I класса - антиген протекает внутриклеточно непрерывно в эндоплазматическом ретикулуме. В его состав включаются любые эндогенно синтезированные пептиды, в том числе вирусные, куда они переносятся из цитоплазмы при помощи особого белка, протеосомы. Включенный в комплекс пептид придает структурную устойчивость MHC I класса. В его отсутствие функцию стабилизатора выполняет шаперон (калнексин).
MHC I класса экспрессируются на поверхности практически всех клеток, кроме эритроцитов и клеток ворсинчатого трофобласта (профилактика отторжения плода). Плотность MHC I класса достигает 7000 молекул на клетку, и они покрывают около 1% ее поверхности. Для них характерна высокая скорость биосинтеза - процесс завершается за 6 ч. Экспрессия MHC I класса усиливается под влиянием цитокинов, например γ-интерферона.
В настоящее время у человека различают более 200 различных вариантов HLA I класса. Они кодируются генами, картированными
в трех основных сублокусах 6-й хромосомы и наследуются и проявляются независимо: HLA-A, HLA-B и HLA-C. Локус А объединяет более 60 вариантов, В - 130, а С - около 40. Независимое наследование генов сублокусов в популяции формирует бесконечное множество неповторяющихся комбинаций HLA I класса. Каждый человек строго уникален по набору антигенов гистосовместимости, исключение составляют только однояйцовые близнецы. Основная биологическая роль HLA I класса - они определяют биологическую индивидуальность (биологический паспорт) и являются маркерами «своего» для иммунокомпетентных клеток. Заражение клетки вирусом или ее мутация изменяют структуру HLA I класса, что является сигналом для активации Т-киллеров (CD8+-лимфоциты) к уничтожению объекта.
HLA I класса выявляют на лимфоцитах в реакции микролимфоцитолиза со специфическими сыворотками, которые получают от многорожавших женщин, пациентов после массивной гемотрансфузии, а также с использованием моноклональных антител.
В структуре и функции MHC II класса есть ряд принципиальных отличий. Комплекс образован двумя нековалентно связанными полипептидными цепями (α и β), имеющими сходное доменное строение (см. рис. 10.1). Обе цепи являются трансмембранными пептидами и «заякорены» в цитоплазматической мембране. Щель Бъеркмана в MHC II класса образована одновременно обеими цепями. Она вмещает олигопептид размером 12-25 аминокислотных остатков, недосягаемый специфическими антителами. MHC II класса включает в себя пептид, захваченный из внеклеточной среды путем эндоцитоза, а не синтезированный самой клеткой. Молекулы МНС II класса экспрессируются на поверхности ограниченного числа клеток: дендритных, В-лимфоцитах, Т-хелперах, активированных макрофагах, тучных, эпителиальных и эндотелиальных клетках. Обнаружение MHC II класса на нетипичных клетках расценивается в настоящее время как иммунопатология. Биосинтез MHC II класса протекает в эндоплазматическом ретикулуме и экспрессируется на цитоплазматической мембране клетки в течение 1 ч после эндоцитоза антигена. Экспрессия комплекса может быть усилена γ-интерфероном и снижена простагландином Е2.
У мыши антиген гистосовместимости получил название Ia- антигена, а у человека по аналогии - HLA II класса.
По имеющимся данным, человеческому организму свойствен чрезвычайно высокий полиморфизм HLA II класса, который в большей степени определяется особенностями строения β-цепи. В состав комплекса входят продукты трех основных локусов: HLA- DR, DQ и DP. При этом локус DR объединяет около 300 аллельных форм, DQ - около 400, а DP - около 500.
Наличие и тип MHC II класса определяют в серологических (микролимфоцитотоксический тест) на В-лимфоцитах и клеточных реакциях иммунитета (смешанная культура лимфоцитов). Специфические антитела к MHC II класса получают так же, как и к I классу. Тестирование в смешанной культуре лимфоцитов позволяет выявить минорные компоненты MHC II класса, не определяемые серологически.
MHC II класса участвуют в индукции приобретенного иммунного ответа. Фрагменты молекулы антигена экспрессируются на цитоплазматической мембране особой группы клеток, которая получила название антигенпрезентирующих. Основными являются дендритная клетка, макрофаг и В-лимфоцит. Структура MHC II класса с включенным в него пептидом в комплексе с кофакторными молекулами CD-антигенов воспринимается и анализируется Т-хелперами (CD4+-лимфоциты). В случае распознавания чужеродности Т-хелпер начинает синтез соответствующих иммуноцитокинов, и включается механизм специфического иммунного реагирования: пролиферация и дифференцировка антигенспецифических клонов лимфоцитов.
Помимо описанных выше антигенов гистосовместимости, идентифицирован III класс молекул MHC. Локус, содержащий кодирующие их гены, вклинивается между I и II классами и разделяет их. К MHC III класса относятся некоторые компоненты комплемента (С2, С4), белки теплового шока, факторы некроза опухоли и др.
10.1.4.3. Опухольассоциированные антигены
В 1948-1949 гг. видный отечественный микробиолог и иммунолог Л.А. Зильбер при разработке вирусной теории рака доказал наличие антигена, специфичного для опухолевой ткани. Позже в 60-х годах ХХ века Г.И. Абелев (в опытах на мышах) и Ю.С. Татаринов (при обследовании людей) обнаружили в сыворотке крови больных первичным раком печени эмбриональный вариант сывороточного альбумина - α-фетопротеин. К настоящему моменту обнаружено и охарактеризовано множество опухольассоциирован-
ных антигенов. Однако не все опухоли содержат специфические маркерные антигены, равно как и не все маркеры обладают строгой тканевой специфичностью.
Опухольассоциированные антигены классифицируют по локализации и генезу. Различают сывороточные, секретируемые опухолевыми клетками в межклеточную среду, и мембранные. Последние получили название опухолеспецифических трансплантационных антигенов, или TSTA (от англ. Tumor-Specific Transplantation Antigen).
Выделяют также вирусные, эмбриональные, нормальные гиперэкспрессируемые и мутантные опухольассоциированные антигены. Вирусные - являются продуктами онковирусов, эмбриональные в норме синтезируются в зародышевом периоде. Хорошо известен α-фетопротеин (эмбриональный альбумин), нормальный протеин тестикул (MAGE 1,2,3 и др.), маркеры меланомы, рака молочной железы и др. Хорионический гонадотропин, в норме синтезируемый в плаценте, обнаруживается при хориокарциноме и других опухолях. В меланоме в большом количестве синтезируется нормальный фермент тирозиназа. Из мутантных белков следует отметить протеин Ras - ГТФ-связывающий белок, участвующий в трансмембранном проведении сигнала. Маркерами рака молочной и поджелудочной желез, карцином кишечника являются модифицированные муцины (MUC 1, 2 и др.).
В большинстве случаев опухольассоциированные антигены представляют собой продукты экспрессии генов, в норме включаемых в эмбриональном периоде. Они являются слабыми иммуногенами, хотя в отдельных случаях могут индуцировать реакцию цитотоксических Т-лимфоцитов (Т-киллеров) и распознаваться в составе молекул MHC (HLA) I класса. Синтезируемые к опухольассоциированным антигенам специфические антитела не угнетают рост опухолей.
10.1.4.4. CD-антигены
На мембране клеток обнаруживаются групповые антигены, объединяющие клетки с определенными морфофункциональными характеристиками. Эти молекулы получили название антигенов кластеров дифференцировки клетки, или CD-антигенов (от англ. Cell Differentiation Antigens, или Claster Definition). По структуре они являются гликопротеинами и в большинстве своем относятся к суперсемейству иммуноглобулинов.
Список CD-маркеров довольно обширный и насчитывает около 200 вариантов. Среди многообразия CD-антигенов наиболее широкое распространение получили маркеры иммунокомпетентных клеток. Например, CD3 экспрессируется в популяции Т-лимфоцитов, CD4 - Т-хелперов, а CD8 - цитотоксических Т-лимфоцитов Т-киллеров, CD11a - моно- и гранулоцитов, CD11b - естественных киллеров, CD19-22 - В-лимфоцитов. Информация о структуре закодирована в различных участках генома, а экспрессия зависит от стадии дифференцировки клетки и ее функционального состояния.
CD-антигены имеют значение в диагностике иммунодефицитных состояний. Определение CD-маркеров осуществляется в иммунологических реакциях с использованием моноклональных антител.
10.1.5. Антигены микробов
10.1.5.1. Антигены бактерий
В структуре бактериальной клетки различают жгутиковые, соматические, капсульные и некоторые другие антигены (рис. 10.2). Жгутиковые, или Н-антигены, локализуются в их жгутиках и представляют собой эпитопы сократительного белка флагеллина. При нагревании флагеллин денатурирует и Н-антиген теряет свою специфичность. Фенол не действует на этот антиген.
Соматический, или О-антиген, связан с клеточной стенкой бактерий. Его основу составляют липополисахариды. О-антиген термостабилен и не разрушается при длительном кипячении. Однако альдегиды (например, формалин) и спирты нарушают его структуру.
Если проиммунизировать животное живыми бактериями, имеющими жгутики, то будут вырабатываться антитела одновременно к О- и Н-антигенам. Введение животному прокипяченной культуры стимулирует биосинтез антител к соматическому антигену. Культура бактерий, обработанная фенолом, вызовет образование антител к жгутиковым антигенам.
Капсульные, или К-антигены, встречаются у бактерий, образующих капсулу. Как правило, К-антигены состоят из кислых полисахаридов (уроновые кислоты). В то же время у бациллы сибирской язвы этот антиген построен из полипептидных цепей. По чувствительности к нагреванию различают три типа К-антигена: А, В и L.
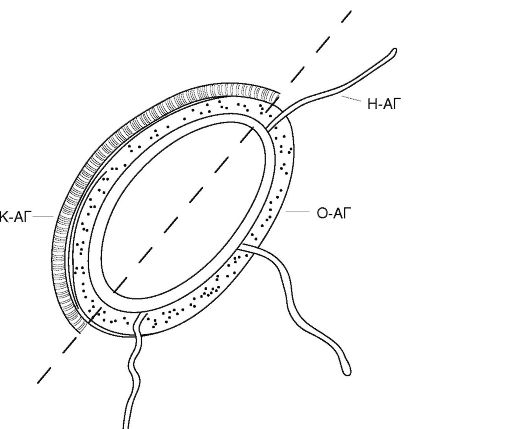 Рис. 10.2. Основные бактериальные антигены (пояснение в тексте)
Рис. 10.2. Основные бактериальные антигены (пояснение в тексте)
Наибольшая термостабильность характерна для группы А - они не денатурируют даже при длительном кипячении. Группа В выдерживает непродолжительное нагревание (около 1 ч) до 60 °С. Группа L быстро разрушается при этой температуре. Поэтому частичное удаление К-антигена возможно путем длительного кипячения бактериальной культуры.
На поверхности возбудителя брюшного тифа и других энтеробактерий, которые обладают высокой вирулентностью, можно обнаружить особый вариант капсульного антигена. Он получил название антигена вирулентности, или Vi-антигена. Обнаружение этого антигена или специфичных к нему антител имеет большое диагностическое значение.
Антигенными свойствами обладают также бактериальные белковые токсины, ферменты и некоторые другие вещества, которые секретируются бактериями в окружающую среду (например, тубер-
кулин). Столбнячный, дифтерийный и ботулинический токсины относятся к числу сильных полноценных антигенов, поэтому их используют для получения молекулярных вакцин - анатоксинов.
В антигенном составе некоторых бактерий выделяется группа антигенов с сильно выраженной иммуногенностью, чья биологическая активность играет ключевую роль в формировании патогенности возбудителя - связывание таких антигенов специфическими антителами практически полностью инактивирует вирулентные свойства микроорганизма и обеспечивает к нему иммунитет. Эти антигены получили название протективных.
10.1.5.2. Антигены вирусов
В структуре вирусной частицы различают ядерные (или коровые), капсидные (или оболочечные) и суперкапсидные антигены. На поверхности некоторых вирусных частиц встречаются особые V-антигены - гемагглютинин и фермент нейраминидаза. Антигены вирусов различаются по происхождению. Часть из них вирусоспецифические, кодируются в нуклеиновой кислоте вируса. Другие, являющиеся компонентами клетки хозяина (углеводы, липиды), формируют суперкапсид вируса при его рождении путем почкования.
Антигенный состав вириона зависит от строения самой вирусной частицы. В просто организованных вирусах антигены ассоциированы с нуклеопротеидами. Эти вещества хорошо растворяются в воде и поэтому обозначаются как S-антигены (от лат. solutio - раствор). У сложноорганизованных вирусов часть антигенов связана с нуклеокапсидом, а другая находится во внешней оболочке, или суперкапсиде.
Антигены многих вирусов отличаются высокой степенью изменчивости, что связано с постоянными мутациями в генетическом материале вирусов. Примером могут служить вирус гриппа,
ВИЧ и др.
10.1.6. Процессы, происходящие с антигеном в макроорганизме
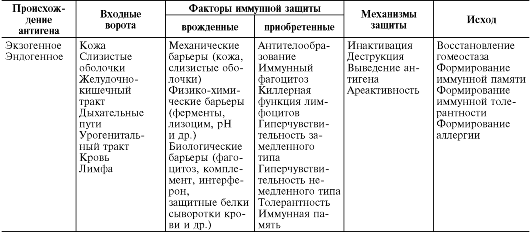
Антигенная интервенция - это процесс, протекающий поэтапно с определенной динамикой во времени. При этом на каждом этапе появления и распространения в макроорганизме антиген сталкивается с мощным противодействием развитой сети разнообразных факторов иммунитета (табл. 10.1).
Таблица 10.1. Процессинг антигена в макроорганизме
 Выделяют
несколько путей проникновения и распространения антигена в
макроорганизме. Они могут появляться внутри самого макроорганизма
(эндогенное происхождение) или поступать извне (экзогенное
происхождение). Экзогенные антигены могут проникнуть в макроорганизм:
Выделяют
несколько путей проникновения и распространения антигена в
макроорганизме. Они могут появляться внутри самого макроорганизма
(эндогенное происхождение) или поступать извне (экзогенное
происхождение). Экзогенные антигены могут проникнуть в макроорганизм:
• через дефекты кожных покровов и слизистых оболочек (как результат ранений, микротравм, укусов насекомых, расчесов и др.);
• путем всасывания в желудочно-кишечном тракте (эндоцитоз эпителиальными клетками);
• межклеточно (при незавершенном фагоцитозе);
• чресклеточно (так распространяются облигатные внутриклеточные паразиты, например вирусы).
В организме антиген может распространяться с лимфой (лимфогенный путь) и кровью (гематогенный путь) по различным органам и тканям. При этом чаще всего он фильтруется в лимфоузлах, селезенке, а также в лимфоидных скоплениях печени, кишечника и других органов, где вступает в контакт с факторами иммунной защиты.
Ответная реакция этих факторов возникает практически немедленно. Первыми вступают в действие факторы врожденного иммунитета, так как эта система не требует длительного времени для активации. Если антиген не был инактивирован или элиминирован в течение 4 ч, включается система приобретенного иммунитета: обеспечивается специфическое распознавание «свой-чужой», вырабатываются факторы регуляции (цитокины) и иммунной защиты (специфические антитела, клоны антигенореактивных лимфоцитов).
Совокупный эффект всех звеньев и уровней иммунной защиты макроорганизма, независимо от степени их вовлечения в процесс, направлен на:
• связывание и блокирование биологически активных участков молекулы антигена;
• разрушение или отторжение антигена;
• утилизацию, изоляцию (инкапсуляцию) или выведение остатков антигена из макроорганизма.
В итоге достигается восстановление гомеостаза и структурной целостности макроорганизма. Параллельно формируется иммунная память, толерантность или аллергия.
10.2. Иммунная система человека
Специфическую функцию надзора за генетическим постоянством внутренней среды организма, сохранения его биологической и видовой индивидуальности осуществляет иммунная система.
10.2.1. Структурно-функциональные элементы иммунной системы
Иммунная система - это специализированная, анатомически обособленная лимфоидная ткань. Она распределена по всему организму в виде различных лимфоидных образований и отдельных клеток, и на ее долю приходится 1-2% от массы тела. В анатомическом плане иммунная система подразделена на центральные и периферические органы, в функциональном - на органы воспроизводства и селекции клеток (костный мозг, тимус), контроля внешней среды или экзогенной интервенции (лимфоидные системы кожи и слизистых оболочек), контроля генетического постоянства внутренней среды (селезенка, лимфатические узлы, печень, кровь, лимфа).
Основными функциональными клетками являются лимфоциты. Их количество в организме достигает 1012. К числу функциональных клеток иммунной системы относят также мононуклеарные и гранулярные лейкоциты, тучные и дендритные клетки. Часть клеток сосредоточена в отдельных органах иммунной системы, другие свободно перемещаются по всему организму. Схематическое строение иммунной системы представлено на рис. 10.3.
10.2.1.1. Центральные органы иммунной системы
Центральные органы иммунной системы, костный мозг и вилочковая железа или тимус, - это органы воспроизводства и селекции клеток иммунной системы. Здесь происходят лимфопоэз - рождение, размножение (пролиферация) и дифференцировка лимфоцитов до стадии предшественников или зрелых неиммунных (наивных) клеток, а также их «обучение». У птиц к центральным органам иммунной системы относят сумку Фабрициуса (bursa Fabricii), локализованную в области клоаки.
Костный мозг располагается в губчатом веществе костей (эпифизы трубчатых костей, грудина, ребра и др.). Здесь находятся полипотентные стволовые клетки (ППСК), которые являются ро-
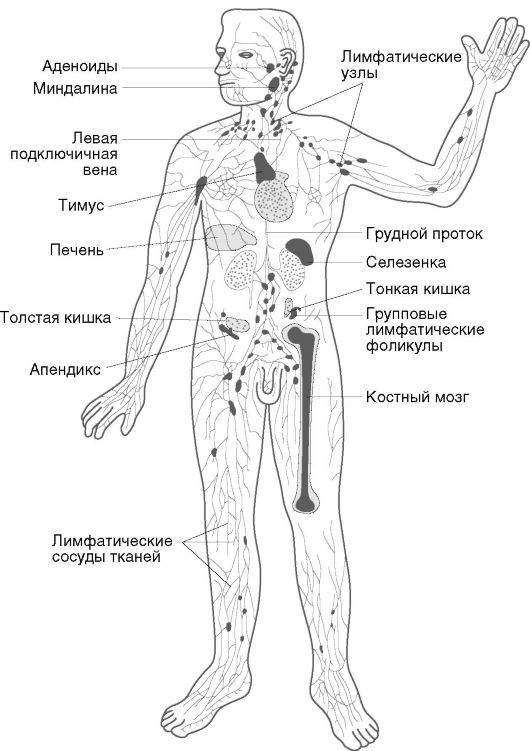 Рис. 10.3. Органы иммунной системы человека
Рис. 10.3. Органы иммунной системы человека
доначальницами всех форменных элементов крови, включая иммунокомпетентные клетки. В строме костного мозга формируются предшественники В- и Т-лимфоцитов, которые впоследствии мигрируют соответственно в В-зоны макроорганизма и тимус. Фагоциты и некоторые дендритные клетки также образуются в костном мозгу. В нем можно также обнаружить плазматические клетки - результат терминальной дифференцировки В-лимфоцитов.
Вилочковая железа, тимус, или зобная железа, располагается в верхней части загрудинного пространства. Этот орган отличается особым морфогенезом. Тимус формируется в период внутриутробного развития. К моменту рождения масса тимуса достигает 10-15 г, окончательно он созревает к пятилетнему возрасту, а максимального размера достигает к 10-12 годам жизни (масса 30-40 г). После периода полового созревания начинается инволюция органа - происходит замещение лимфоидной ткани жировой и соединительной.
Тимус имеет дольчатое строение. В его структуре различают мозговой и корковый слои. В строме коркового слоя находится большое количество эпителиальных клеток коры, названных «клеткиняньки», которые своими отростками образуют мелкоячеистую сеть, где располагаются созревающие лимфоциты. В пограничном, корково-мозговом, слое располагаются дендритные клетки тимуса, а в мозговом - эпителиальные клетки мозгового слоя.
Предшественники Т-лимфоцитов поступают из костного мозга в корковый слой тимуса. Здесь под влиянием тимических факторов они активно размножаются, дифференцируются (превращаются) в зрелые Т-лимфоциты и «учатся» распознавать чужеродные антигенные детерминанты.
Процесс «обучения» включает положительную и отрицательную селекцию. Критерием «обученности» являются качество Т-клеточной антигенной рецепции (специфичность и аффинность) и жизнеспособность клетки.
Положительная селекция происходит в корковом слое при помощи эпителиальных клеток. Суть ее заключается в поддержке клонов Т-лимфоцитов, рецепторы которых эффективно связались с экспрессированными на эпителиальных клетках молекулами MHC, независимо от структуры инкорпорированных собственных олигопептидов. Эпителиоциты коры выделяют ростовые факторы тимуса, активирующие размножение Т-лимфоцитов.
Отрицательную селекцию осуществляют дендритные клетки в пограничной корково-мозговой зоне тимуса. Ее цель - выбраковка аутореактивных клонов Т-лимфоцитов. Клетки, позитивно реагирующие на комплекс MHC-аутологичный пептид, подвергаются уничтожению путем индукции у них апоптоза.
В итоге селекции более 99% Т-лимфоцитов не выдерживают испытаний и погибают. Лишь менее 1% клеток превращается в зрелые формы, способные распознать в комплексе с аутологичными MHC только чужеродные биополимеры. Ежесуточно около 106 зрелых «обученных» Т-лимфоцитов покидают тимус с крово- и лимфотоком и мигрируют в различные органы и ткани.
Созревание и «обучение» Т-лимфоцитов в тимусе имеет важное значение для формирования иммунитета. Отсутствие или недоразвитие тимуса при врожденном дефекте развития вилочковой железы - аплазии или гипоплазии органа, ее хирургическом удалении или радиационном поражении ведет к резкому снижению эффективности иммунной защиты макроорганизма. Между тем тимэктомия у взрослых практически не приводит к серьезным дефектам в иммунитете.
10.2.1.2. Периферические органы иммунной системы
К периферическим органам иммунной системы относят селезенку, лимфатические узлы, аппендикс, печень, миндалины глоточного кольца, групповые лимфатические фолликулы, кровь, лимфу и др. В этих органах проходит иммуногенез - размножение и окончательное созревание предшественников иммунокомпетентных клеток и осуществляется иммунологический надзор. В функциональном плане периферические органы иммунной системы могут быть подразделены на органы контроля внутренней среды организма (лимфатические узлы, селезенка, тканевые мигрирующие клетки) и его кожных и слизистых покровов (аппендикс, лимфатические фолликулы и скопления).
Лимфатические узлы - мелкие округлые анатомические образования бобовидной формы, которые располагаются по ходу лимфатических сосудов. Каждый участок тела имеет региональные лимфоузлы. В общей сложности в организме человека насчитывается до 1000 лимфоузлов. Лимфатические узлы выполняют функцию биологического сита - через них фильтруется лимфа и задерживаются и концентрируются антигены. Через лимфоузел проходит в среднем около 109 лимфоцитов в 1 ч.
В строении лимфоузла различают корковое и мозговое вещество. Строма коры разделена соединительнотканными трабекулами на сектора. В ней выделяют поверхностный корковый слой и паракортикальную зону. В секторах поверхностного коркового слоя расположены лимфатические фолликулы с центрами размножения В-лимфоцитов (герминативные центры). Здесь же обнаруживаются фолликулярные дендритные клетки, способствующие созреванию В-лимфоцитов. Паракортикальный слой - это зона Т-лимфоцитов и интердигитальных дендритных клеток, потомков дермальных клеток Лангерганса. Мозговое вещество образовано тяжами соединительной ткани, между которыми располагаются макрофаги и плазматические клетки.
В пределах лимфоузла происходит антигенная стимуляция иммунокомпетентных клеток и включается система специфического иммунного реагирования, направленная на обезвреживание антигена.
Селезенка - это орган, через который фильтруется вся кровь. Он располагается в левой подвздошной области и имеет дольчатое строение. Лимфоидная ткань образует белую пульпу. В строении различают первичные, периартериальные лимфоидные фолликулы (окружают артерии по их ходу) и вторичные, располагающиеся на границах первичных фолликулов. Первичные лимфоидные скопления заселены преимущественно Т-лимфоцитами, а вторичные - В-лимфоцитами и плазматическими клетками. Кроме того, в строме селезенки обнаруживают фагоциты и ретикулярные дендритные клетки.
В селезенке, как в сите, задерживаются антигены, оказавшиеся в кровотоке, и состарившиеся эритроциты. Этот орган называют кладбищем эритроцитов. Здесь происходят антигенная стимуляция иммунокомпетентных клеток, развитие специфической иммунной реакции на антиген и его обезвреживание.
Печень играет особую роль в иммунной системе. В ней находится более половины всех тканевых макрофагов и большая часть естественных киллеров. Лимфоидные популяции печени обеспечивают толерантность к пищевым антигенам, а макрофаги утилизируют иммунные комплексы, в том числе сорбированные на стареющих эритроцитах.
Групповые лимфатические фолликулы (пейеровы бляшки) являются скоплением лимфоидной ткани в слизистой оболочке тонкой кишки. Такие образования также находятся в червеобразном отростке слепой кишки - аппендиксе. Кроме того, на всем протяже-
нии желудочно-кишечного тракта, начиная с пищевода и кончая анальным отверстием, располагаются единичные лимфатические фолликулы. Они обеспечивают местный иммунитет слизистой оболочки кишки и ее просвета и регулируют видовой и количественный состав ее нормальной микрофлоры.
Скопление лимфоидных элементов в виде миндалин глоточного кольца обеспечивает местный иммунитет в носоглотке, ротовой полости и верхних дыхательных путях, защищает их слизистые оболочки от внедрения микробов и других генетически чужеродных агентов, передающихся воздушно-капельным или воздушнопылевым путем, и регулирует локальную нормофлору.
Лимфа - жидкая ткань организма, которая содержится в лимфатических сосудах и узлах. Она включает в себя все соединения, поступающие из межтканевой жидкости. Основными и практически единственными клетками лимфы являются лимфоциты. В ее составе эти клетки осуществляют кругооборот в организме.
В кровь циркулируют предшественники и зрелые Т- и В-лимфоциты, полиморфно-ядерные лейкоциты, моноциты. Лимфоциты составляют 30% общего количества лейкоцитов. Одномоментно в крови присутствует менее 2% общего количества лимфоцитов.
10.2.1.3. Клетки иммунной системы
Специфическую функцию иммунной защиты непосредственно осуществляет многочисленный пул клеток миелоидного и лимфоидного ростков крови: лимфоциты, фагоциты и дендритные клетки. Это основные клетки иммунной системы. Кроме них, в иммунный ответ может вовлекаться множество других клеточных популяций (эпителий, эндотелий, фибробласты и др.). Перечисленные клетки различаются морфологически, по функциональной активности, маркерам (специфические молекулярные метки), рецепторному аппарату и продуктам биосинтеза. Тем не менее большую часть клеток иммунной системы объединяет близкое генетическое родство: они имеют общего предшественника, полипотентную стволовую клетку костного мозга (рис. 10.4).
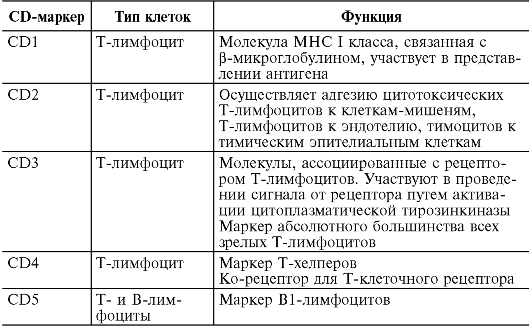
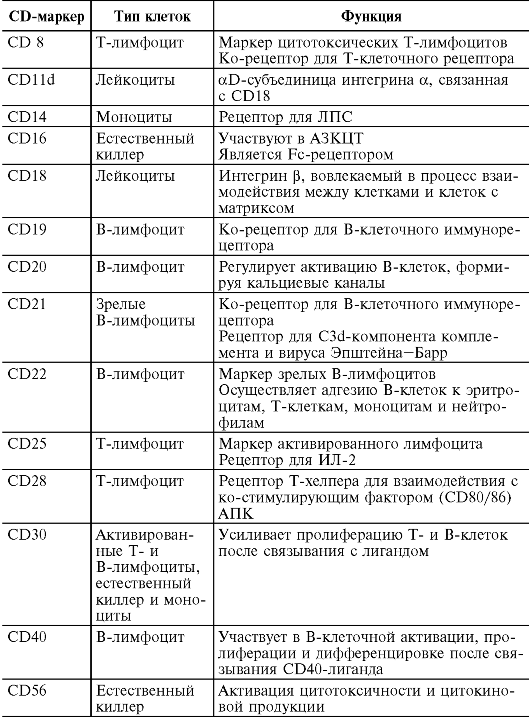
На поверхности цитоплазматической мембраны клеток иммунной системы существуют особые молекулы, которые служат их маркерами. В 80-х годах прошлого века была принята международная номенклатура мембранных маркеров лейкоцитов человека, названных «CD-антигены» (табл. 10.2)
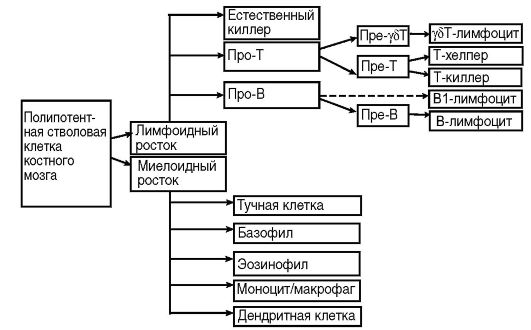 Рис. 10.4. Схема иммуногенеза (пояснения в тексте)
Рис. 10.4. Схема иммуногенеза (пояснения в тексте)
Таблица 10.2. Основные CD-маркеры клеток, участвующих в иммунном ответе
 Продолжение табл. 10.2
Продолжение табл. 10.2
 Окончание табл. 10.2
Окончание табл. 10.2
 Примечание. АЗКЦТ - антителозависимая клеточно-опосредованная цитотоксичность; АПК - антигенпрезентирующие клетки.
Примечание. АЗКЦТ - антителозависимая клеточно-опосредованная цитотоксичность; АПК - антигенпрезентирующие клетки.
По функциональной активности клетки-участники иммунного ответа подразделяют на регуляторные (индукторные), эффекторные и антигенпрезентирующие. Регуляторные клетки управляют функционированием компонентов иммунной системы путем выработки медиаторов - иммуноцитокинов и лигандов. Эти клетки определяют направление развития иммунного реагирования, его интенсивность и продолжительность. Эффекторы являются непосредственными исполнителями иммунной защиты путем прямого воздействия на объект либо путем биосинтеза биологически активных веществ со специфическим эффектом (антитела, токсичные субстанции, медиаторы и пр.).
Антигенпрезентирующие клетки выполняют ответственную задачу: захватывают, процессируют (перерабатывают путем ограниченного протеолиза) и представляют антиген иммунокомпетентным Т-клеткам в составе комплекса с MHC II класса. АПК лишены специфичности в отношении самого антигена. Молекула MHC II класса может включать в себя любые эндоцитированные из межклеточной среды олигопептиды, как свои собственные, так и чужие. Установлено, что большая часть комплексов MHC II класса содержит аутогенные молекулы и лишь малая доля - чужеродный материал.
Помимо MHC II класса, АПК экспрессируют ко-стимулирующие факторы (CD40, 80, 86) и множество молекул адгезии. Последние обеспечивают тесный, пространственно стабильный и продолжительный контакт АПК с Т-хелпером. Кроме того, АПК экспрессируют молекулы CD1, с помощью которых могут презентировать липосодержащие или полисахаридные антигены.
Основными профессиональными АПК являются дендритные клетки костно-мозгового происхождения, В-лимфоциты и макро-
фаги. Дендритные клетки почти в 100 раз эффективнее макрофагов. Функцию непрофессиональных АПК могут также выполнять некоторые другие клетки в состоянии активации - эпителиальные клетки и эндотелиоциты.
Осуществление целенаправленной иммунной защиты макроорганизма возможно благодаря наличию на клетках иммунной системы специфических антигенных рецепторов (иммунорецепторов). По механизму функционирования они подразделяются на прямые и непрямые. Прямые иммунорецепторы непосредственно связываются с молекулой антигена. Непрямые иммунорецепторы взаимодействуют с молекулой антигена опосредованно - через Fc-фрагмент молекулы иммуноглобулина (см. раздел 11.1.2). Это так называемый Fc-рецептор (FcR).
Fc-рецепторы различаются по аффинности. Высокоаффинный рецептор может связываться с интактными молекулами IgE или IgG4 и образовывать рецепторный комплекс, в котором антигенспецифическую ко-рецепторную функцию выполняет молекула иммуноглобулина. Такой рецептор есть у базофилов и тучных клеток. Низкоаффинный FcR распознает молекулы иммуноглобулина, уже образовавшие иммунные комплексы. Он обнаруживается на макрофагах, естественных киллерах, эпителиальных, дендритных и множестве других клеток.
Иммунное реагирование основано на тесном взаимодействии различных клеточных популяций. Это достигается при помощи биосинтеза клетками иммунной системы широкого спектра иммуноцитокинов. Подавляющее большинство клеток иммунной системы постоянно перемещается во внутренних средах организма с крово- и лимфотоком и благодаря амебоидной подвижности.
Клеточно-элементный состав иммунной системы постоянно возобновляется за счет деления стволовых клеток. Состарившиеся, выработавшие свой биологический ресурс, ложно активированные, зараженные и генетически трансформированные клетки уничтожаются.
10.2.1.3.1. Лимфоциты
Лимфоциты - подвижные мононуклеарные клетки. В зависимости от места созревания эти клетки подразделяются на две популяции Т- (тимус) и В- (бурса Фабрициуса, костный мозг) лимфоцитов. Лимфоциты играют ключевую роль в обеспечении приобретенного (адаптивного) иммунитета. Они осуществляют
специфическое распознавание антигена, индукцию клеточного и гуморального иммунного ответа, различные формы иммунного реагирования.
В организме непрерывно идет возобновление популяций лимфоцитов, клетки активно мигрируют между различными органами и тканями. Вместе с тем миграция и расселение лимфоцитов в тканях не являются хаотическим процессом. Он имеет направленный характер и строго регулируется экспрессией на мембране лимфоцитов, эндотелии сосудов и клеточных элементах стромы особых молекул адгезии (интегрины, селектины и др.). Так, незрелые Т-лимфоциты активно мигрируют в тимус. Зрелые неиммунные («наивные») лимфоциты тропны к периферическим лимфоидным органам и тканям. При этом Т- и В-лимфоциты заселяют только «свои» области - это так называемый эффект хоминговой рецепции (от англ. home - дом). Зрелые иммунные (активированные) лимфоциты распознают эпителий в очаге воспаления. Клетки иммунологической памяти всегда возвращаются в места своего происхождения.
Продолжительность жизни неиммунных лимфоцитов достаточно большая. У Т-лимфоцитов она достигает нескольких месяцев или лет, а у В-клеток - недель или месяцев. Дольше всех живут клетки иммунологической памяти (см. раздел 11.5) - до 10 лет и более. Однако активированные или терминально дифференцированные лимфоциты имеют короткую продолжительность жизни (несколько суток). Состарившиеся, ложно активированные и аутореактивные (реагирующие на аутоантигены) лимфоциты подвергаются уничтожению путем индукции у них апоптоза. Погибшие лимфоциты постоянно заменяются новыми за счет их пролиферации в центральных и периферических органах иммунной системы. Численность лимфоидных популяций находится под жестким контролем клеток самой иммунной системы.
Для выполнения специфической функции лимфоциты несут на своей поверхности прямые антигенные рецепторы и являются иммунокомпетентными клетками. Иммунорецептор В-лимфоцита и особого γδТ-лимфоцита распознает нативный эпитоп, т.е. непосредственно отличает чужеродные субстанции. Иммунорецептор традиционного Т-лимфоцита ориентирован на олигопептиды в составе MHC, т.е. распознает измененное «свое».
Антигенспецифические рецепторы лимфоцитов имеют сложное молекулярное строение, уникальное для каждой клетки. Напри-
мер, у Т-лимфоцитов они состоят из нескольких полипептидных субъединиц, имеющих полигенное кодирование. Число генов, детерминирующих структуру V-области этого рецептора (вариабельный участок, ответственный за специфическое распознавание), в незрелой клетке достигает 100. При созревании лимфоцита в результате рекомбинационных перестроек в V-генах, индивидуальных для каждой клетки, образуется бесконечно большое количество вариантов антигенной специфичности рецептора, достигающее 1012, что сопоставимо с общей численностью популяции Т-лимфоцитов. Формирование В-клеточного рецептора имеет те же закономерности. Биологический смысл феномена чрезвычайно важен: в организме постоянно поддерживается широкий репертуар специфической направленности лимфоидных рецепторов, и клетки готовы в любой момент ответить защитной реакцией на любой возможный антиген.
В такой ситуации закономерно появление Т-лимфоцитов, специфичных для антигенов собственного организма. Однако они должны элиминироваться в тимусе на ранних этапах своего развития. Поэтому различают первичный и вторичный антигенраспознающий репертуар лимфоидных популяций. Первичный характеризуется набором рецепторных специфичностей, формирующимся при образовании лимфоцитов в костном мозгу индивидуума. Вторичный, или клональный, репертуар является совокупностью вариантов рецептора после отбраковки аутореактивных клонов клеток.
Антигенспецифическая рецепция в лимфоцитах имеет стандартные механизмы реализации. Полученный внеклеточной частью рецептора сигнал от раздражителя (антигена) передается по трансмембранному участку на его внутриклеточную часть, которая уже активирует внутриклеточные ферменты (тирозинкиназу, фосфорилазу и др.).
Для запуска продуктивной реакции лимфоцита необходима агрегация его рецепторов. Кроме того, для стабилизации рецепторлигандного взаимодействия и восприятия ко-стимулирующего сигнала требуются вспомогательные молекулы.
Среди лимфоцитов встречаются клетки без отличительных признаков Т- и В-лимфоцитов. Они получили название нулевых клеток. В костном мозгу на их долю приходится около 50% всех лимфоцитов, а в крови - примерно 5%. Функциональная активность остается неясной.
В-лимфоциты. В-лимфоциты - это преимущественно эффекторные иммунокомпетентные клетки, на долю которых приходится около 15% всей численности лимфоцитов. Выделяют две субпопуляции В-лимфоцитов: традиционные В-клетки, не имеющие маркера CD5-, и CD5+ В1-лимфоциты.
При электронной микроскопии CD5- В-лимфоциты имеют шероховатую поверхность, на ней определяются CD19-22 и некоторые другие. Функцию антигенспецифического рецептора (BCR) выполняют особые мембранные формы иммуноглобулинов. Клетки экспрессируют MHC II класса, ко-стимулирующие молекулы CD40, 80, 86, FcR к иммунным комплексам и нативным молекулам иммуноглобулина класса G, рецептор к эритроцитам мыши, иммуноцитокинам и др.
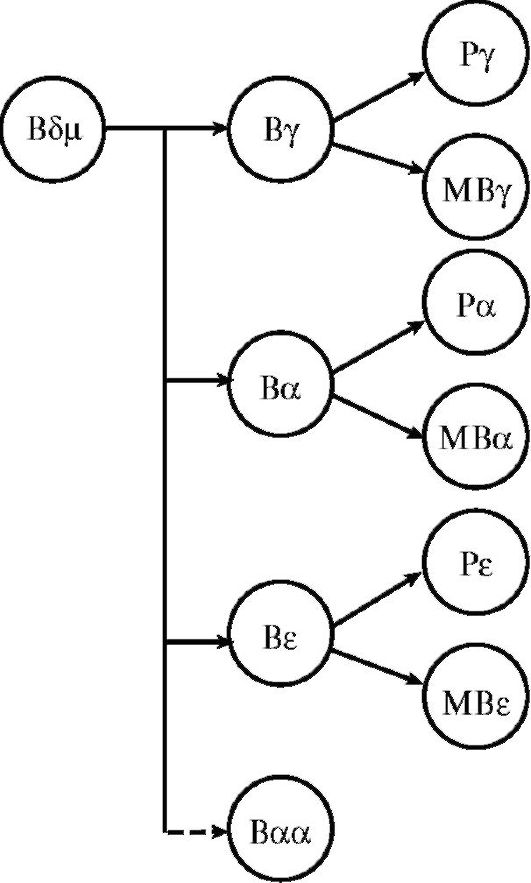 Рис. 10.5. Схема
дифференцировки В-лимфоцита: Р - плазматическая клетка; МВ - В-лимфоцит
иммунологической памяти; Вαα - синтезирует полимерный иммуноглобулин А в
слизистых оболочках
Рис. 10.5. Схема
дифференцировки В-лимфоцита: Р - плазматическая клетка; МВ - В-лимфоцит
иммунологической памяти; Вαα - синтезирует полимерный иммуноглобулин А в
слизистых оболочках
Функцией зрелых CD5- В-лимфоцитов и их потомков (плазмоцитов) является продукция иммуноглобулинов. Кроме того, В-лимфоциты являются профессиональными АПК. Они участвуют в формировании гуморального иммунитета, В-клеточной иммунологической памяти и гиперчувствительности немедленного типа.
Дифференцировка и созревание В-лимфоцитов (рис. 10.5) происходят сначала в костном мозге, а затем в периферических органах иммунной системы, куда они отселяются на стадии предшественников. Потомками В-лимфоцитов являются клетки иммунологической памяти и плазматические клетки. Основные морфологические признаки последних - развитый эндоплазматический ретикулум и аппарат Гольджи с большим количеством рибо-
сом. Плазмоцит имеет короткий период жизни - не более 2-3 сут.
В1-лимфоциты считают филогенетически наиболее древней ветвью антителопродуцирующих клеток. Предшественники этих клеток рано мигрируют в ткани слизистых оболочек, где автономно от центральных органов иммунной системы поддерживают численность своей популяции. Клетки экспрессируют CD5, синтезируют низкоаффинные IgA и IgM к полисахаридным и липидным антигенам микробов и обеспечивают иммунную защиту слизистых оболочек от условно-патогенных бактерий.
Функциональной активностью В-лимфоцитов управляют молекулярные антигены и иммуноцитокины Т-хелпера, макрофага и других клеток.
Т-лимфоциты. Т-лимфоциты - это сложная по составу группа клеток, которая происходит от полипотентной стволовой клетки костного мозга, а созревает и дифференцируется в тимусе из предшественников. На долю этих клеток приходится около 75% всей лимфоидной популяции. На электронограмме все Т-лимфоциты имеют гладкую поверхность, их общим маркером являются CD3, а также рецептор к эритроцитам барана. В зависимости от строения антигенного рецептора (TCR) и функциональной направленности сообщество Т-лимфоцитов может быть разделено на группы.
Различают два типа TCR: αβ и γδ. Первый тип - гетеродимер, который состоит из двух полипептидных цепей - α и β. Он характерен для традиционных Т-лимфоцитов, известных как Т-хелперы и Т-киллеры. Второй обнаруживается на поверхности особой популяции γδТ-лимфоцитов.
Т-лимфоциты функционально также разделяются на две субпопуляции: иммунорегуляторов и эффекторов. Задачу регуляции иммунного ответа выполняют Т-хелперы. Ранее предполагалось существование Т-супрессоров, способных тормозить развитие иммунной реакции (супрессия). Однако до сих пор клетка морфологически не идентифицирована, хотя сам супрессорный эффект существует. Эффекторную функцию осуществляют цитотоксические лимфоциты Т-киллеры.
В организме Т-лимфоциты обеспечивают клеточные формы иммунного ответа (гиперчувствительность замедленного типа, трансплантационный иммунитет и т.д.), определяют силу и продолжительность иммунной реакции. Их созреванием, дифференцировкой и активностью управляют цитокины и макрофаги.
Т-хелперы. Т-хелперы или Т-помощники - субпопуляция Т-лимфоцитов, которые выполняют регуляторную функцию. На их долю приходится около 75% всей популяции Т-лимфоцитов. Они несут маркер CD4, а также αβ TCR, с помощью которого анализируют природу антигена, представляемую им АПК.
Рецепция антигена Т-хелпером, т.е. анализ его чужеродности, - весьма сложный процесс, требующий высокой точности. Ему способствуют (рис. 10.6) молекула CD3 (комплексируется с TCR), ко-рецепторные молекулы CD4 (имеют сродство к молекулярному комплексу MHC II класса), молекулы адгезии (стабилизируют межклеточный контакт), рецепторы (взаимодействуют с ко-стимулирующими факторами АПК - CD28, 40L).
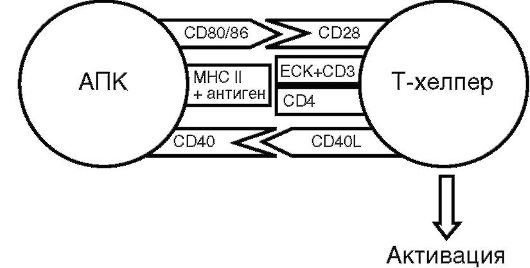 Рис. 10.6. Схема активации Т-хелпера (пояснение в тексте)
Рис. 10.6. Схема активации Т-хелпера (пояснение в тексте)
Активированный Т-хелпер продуцирует широкий спектр иммуноцитокитов, при помощи которых он управляет биологической активностью множества клеток, вовлеченных в иммунный ответ.
Популяция Т-хелперов гетерогенна. Активированный CD4+ Т-лимфоцит (ТΩ-хелпер) дифференцируется в одного из своих потомков: T1- или Т2-хелпер (рис. 10.7). Эта дифференцировка является альтернативной и направляемой цитокинами. Т1- или Т2-хелперы различаются лишь функционально по спектру продуцируемых цитокинов.
Т1-хелпер образует ИЛ-2, 3, γ-ИФН, ФНО и др., необходимые для развития клеточного иммунного ответа, гиперчувствительности замедленного типа, иммунного воспаления. Формирование этой клетки определяют активированный макрофаг, естественный и Т-киллеры, синтезирующие ИЛ-12 и γ-ИФН.
Т2-хелпер продуцирует ИЛ-4, 5, 6, 9, 10, 13 и др., которые поддерживают гуморальный иммунный ответ, а также гиперчув-
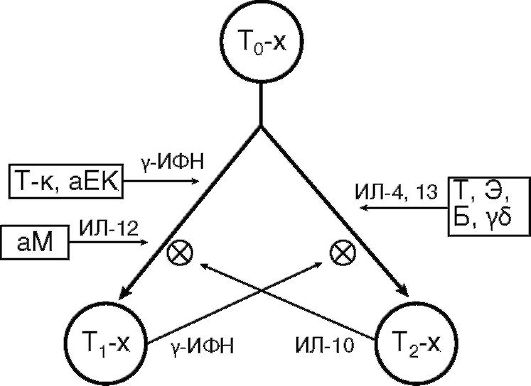 Рис. 10.7. Схема
дифференцировки Т-хелпера: Т-х - Т-хелпер; аМ - активированный
макрофаг; Т-к - Т-киллер; аЕК - активированный естественный киллер; Э -
эозинофил; Б - базофил; Т - тучная клетка; γδТ - γδТ-лимфоцит
Рис. 10.7. Схема
дифференцировки Т-хелпера: Т-х - Т-хелпер; аМ - активированный
макрофаг; Т-к - Т-киллер; аЕК - активированный естественный киллер; Э -
эозинофил; Б - базофил; Т - тучная клетка; γδТ - γδТ-лимфоцит
ствительность немедленного типа. Дифференцировку в сторону Т2-хелпера потенцируют γδТ-клетки, базофилы, тучные клетки и эозинофилы, синтезирующие ИЛ-4 и 13.
В организме поддерживается баланс Т1-/Т2-хелперов, который необходим для развития адекватного иммунного ответа. Т1- и Т2- хелперы являются антагонистами и тормозят развитие друг друга. Установлено, что в организме новорожденных преобладают Т2-хелперы. Нарушение заселения желудочно-кишечного тракта нормальной микрофлорой тормозит развитие субпопуляции Т1- хелперов и ведет к аллергизации организма.
Т-киллеры (цитотоксические Т-лимфоциты). Т-киллер - субпопуляция Т-лимфоцитов-эффекторов, на долю которых приходится примерно 25% всех Т-лимфоцитов. На поверхности Т-киллера определяются молекулы CD8, а также αβTCR к антигену в комплексе с MHC I класса, по которому «свои» клетки отличаются от «чужих». В рецепции принимают участие молекула CD3, комплексирующая с TCR, и ко-рецепторные молекулы CD8, тропные к MHC I класса (рис. 10.8).
Т-киллер анализирует клетки собственного организма в поисках чужеродного MHC I класса. Клетки мутантные, пораженные вирусом, или аллогенного трансплантата несут на своей поверхности такие признаки генетической чужеродности, поэтому являются мишенью Т-киллера.
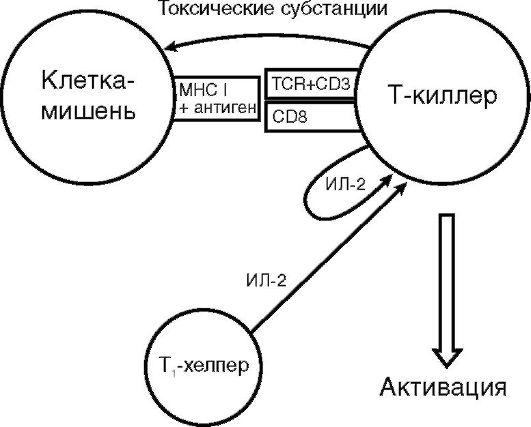 Рис. 10.8. Схема активации Т-киллера (пояснения в тексте)
Рис. 10.8. Схема активации Т-киллера (пояснения в тексте)
Т-киллер устраняет клетки-мишени путем антителонезависимой клеточно-опосредованной цитотоксичности (АНКЦТ) (см. раздел 11.3.2), для чего синтезирует ряд токсичных субстанций: перфорин, гранзимы и гранулизин. Перфорин - токсичный белок, который синтезируют цитотоксические лимфоциты-Т-киллеры и естественные киллеры. Обладает неспецифическим свойством. Вырабатывается только зрелыми активированными клетками. Перфорин образуется в виде растворимого белка-предшественника и накапливается в цитоплазме в гранулах, которые сосредоточиваются около TCR, связавшегося с клеткой-мишенью для обеспечения локального, адресного поражения клетки-мишени. Содержимое гранул высвобождается в узкую синаптическую щель, образованную тесным контактом цитотоксического лимфоцита и клеткимишени. За счет гидрофобных участков перфорин встраивается в цитоплазматическую мембрану клетки-мишени, где в присутствии ионов Са2+ полимеризуется в трансмембранную пору диаметром 16 нм. Образовавшийся канал может вызвать осмотический лизис клетки-мишени (некроз) и/или обеспечить проникновение в нее гранзимов и гранулизина.
Гранзимы - это обобщающее название сериновых протеаз, синтезируемых зрелыми активированными цитотоксическими лимфоцитами. Различают три типа гранзимов: А, В и С. После синтеза гранзимы накапливаются в гранулах подобно перфорину и вместе
с ним выделяются из клетки в синаптическую щель. В клеткумишень проникают через поры, образованные перфорином. Мишенью для гранзимов являются внутриклеточные ферменты, инициирующие апоптоз и обладающие широкой нуклеазной активностью, в том числе способностью разрушать нуклеиновые кислоты внутриклеточных паразитов. Таким образом, гранзимы индуцируют гибель клетки путем апоптоза и санацию организма от зараженных клеток.
Гранулизин - эффекторная молекула с ферментативной активностью, синтезируемая цитотоксическими лимфоцитами. Способен запускать в клетках-мишенях апоптоз, повреждая мембрану их митохондрий.
Т-киллер обладает огромным биологическим потенциалом - его называют серийным убийцем. За короткий срок он может уничтожить несколько клеток-мишеней, затрачивая на каждую около 5 мин. Эффекторную функцию Т-киллера стимулирует Т1-хелпер, хотя в ряде случаев его помощь не требуется. Помимо эффекторной функции, активированный Т-киллер синтезирует γ-ИФН и ФНО, стимулирующие макрофаг и потенцирующие иммунное воспаление.
γδТ-лимфоциты. Среди Т-лимфоцитов существует малочисленная популяция клеток с фенотипом CD4-CD8-, которые несут на своей поверхности особый TCR γδ-типа - γδТ-лимфоциты. Локализуются в эпидермисе и слизистой оболочке желудочнокишечного тракта. Их общая численность не превышает 1% общего пула Т-лимфоцитов, однако в покровных тканях она может достигать 10%.
γδТ-лимфоциты происходят из автономного ростка стволовых клеток, мигрировавших в покровные ткани на ранних этапах эмбриогенеза. В созревании минуют тимус. Активируются клетками поврежденного эпителия желудочно-кишечного тракта и эпидермиса, размножение усиливается ИЛ-7.
Антигенный рецептор γδТ-лимфоцита сходен с BCR, его активный центр непосредственно связывается с эпитопом антигена без его предварительного процессинга и участия MHC. Антигенные детерминанты могут быть представлены, например, молекулами CD1. γδTCR ориентированы на распознавание некоторых широко распространенных микробных антигенов (липопротеинов, белков теплового шока, бактериальных суперантигенов и др.).
γδТ-лимфоциты могут быть как эффекторными, цитотоксическими клетками (принимают участие в удалении патогенов на ранних этапах антиинфекционной защиты), так и регуляторами иммунореактивности. Синтезируют цитокины, активирующие местный иммунитет и локальную воспалительную реакцию, в том числе усиливают образование Т2-хелперов. Кроме того, γδ-клетки продуцируют ИЛ-7 и управляют численностью собственной популяции.
10.2.1.3.2. Другие клетки иммунной системы
Помимо лимфоцитов, в развитии иммунного ответа участвует множество различных клеточных популяций, относящихся в основном к миелоидному ростку. Особого внимания заслуживают естественные киллеры, гранулоциты, тучные и дендритные клетки.
Естественные или нормальные киллеры, NK-клетки (от англ. Natural killer) были изначально описаны как большие гранулярные лимфоциты, способные распознать в организме некоторые виды раково-трансформированных клеток и уничтожить их без предварительной подготовки, что обусловило название клеток.
Сейчас установлено, что естественные киллеры (ЕК) имеют морфологию малых лимфоцитов, на долю которых приходится около 15% всех лимфоцитов. Они образуются из ППСК костного мозга, мигрируют с кровотоком, но отсутствуют в лимфе. Обнаруживаются в печени, селезенке, слизистых оболочках, матке. По маркерам, местам типичной локализации и эффекторным механизмам выделяют две субпопуляции ЕК: кровяную и тканевую. ЕК - главный защитник макроорганизма от внутриклеточных паразитов. Он срабатывает задолго до активации адаптивного иммунитета. Вместе с тем биологические возможности ЕК весьма ограничены.
Кровяные ЕК - это активно циркулирующие в кровотоке клетки. Обнаруживаются в красной пульпе селезенки. Несут на себе маркер CD16+CD56мало, FcR к иммуноглобулину класса G, связанному в иммунный комплекс, и рецептор к MHC I класса. При активации синтезируют, накапливая в гранулах, перфорин, гранзимы и гранулизин. Эффекторная функция кровяных ЕК в отношении меченных иммуноглобулинами клеток реализуется в АЗКЦТ (см. раздел 11.3.1). Мишенями являются клетки, инфицированные внутриклеточными паразитами (бактерии, вирусы, простейшие), аллогенные клетки трансплантата и некоторые опухоли.
Рецептор к MHC I класса анализирует плотность его экспрессии на мембране клетки. Дефицит этих молекул, наблюдающийся при раковой трансфорации клеток, также потенцирует цитотоксичность ЕК.
Тканевые ЕК ведут более оседлый образ жизни и обнаруживаются в большом количестве в печени и децидаульной оболочке беременной матки. Несут маркер CD16-CD56много и много Fas-лиганда. Реализуют АНКЦТ (см. раздел 11.3.2). Клетками-мишенями являются лимфоциты, активированные, например, пищевыми антигенами или аллоантигенами плода и экспрессирующие Fas.
Помимо цитотоксических функций, ЕК вырабатывают цитокины (ИЛ-5, 8, γ-ИФН, ФНО, гранулоцит-моноцит-колониестимулирующий фактор-ГМ-КСФ и др.), активирует макрофагально-фагоцитарное звено, развитие иммунного ответа и иммунного воспаления. Эффекторная функция ЕК усиливается цитокинами (ИЛ-2, 4, 10, 12, γ-ИФН и др.).
Фагоциты (см. раздел 9.2.3.1) - самая многочисленная морфологически гетерогенная фракция иммунокомпетентных клеток. Выполняют регуляторную и эффекторную функции. Вырабатывают иммуноцитокины, ферменты, ион-радикалы и другие биологически активные вещества, осуществляют вне- и внутриклеточный киллинг и фагоцитоз. Кроме того, макрофаги являются АПК - обеспечивают процессинг и презентацию антигена Т-хелперам.
Эозинофилы - гранулярные лейкоциты крови. Содержатся в крови, рыхлой соединительной ткани, в большом количестве накапливаются в очагах местного воспаления, вызванного гельминтами, и обеспечивают АЗКЦТ.
Эозинофилы несут на мембране низкоаффинные FcR к IgA или IgE, распознающие паразитов, отмеченных такими антителами. Активированная клетка выделяет ферменты (эозинофильная пероксидаза и коллагеназа) и белковые токсины (главный щелочной протеин, эозинофильный катионный белок и нейротоксин), губительно действующие на гельминты.
Эозинофилы также синтезируют цитокины (ИЛ-3, 5, 8, ГМ-КСФ и др.), стимулирующие клеточное звено иммунитета и образование Т2-хелпера, и липидные медиаторы (лейкотриены, тромбоцитактивирующий фактор и др.), запускающие воспалительную реакцию в месте внедрения гельминта.
Тучные клетки - немигрирующие морфологические элементы неясного происхождения, располагаются оседло вдоль барьерных тканей (lamina propria слизистых оболочек, в подкожной соединительной ткани) и в соединительной ткани кровеносных сосудов. По набору синтезируемых биологически активных соединений и локализации выделяют две разновидности тучных клеток - клетки слизистых оболочек и соединительной ткани.
Базофилы - гранулоциты, происходящие из костно-мозговой стволовой ППСК и родственные эозинофилам. Их дифференцировка альтернативно определяется цитокинами. Постоянно мигрируют с кровотоком, привлекаются в очаг воспаления анафилотоксинами (С3а, С4а и С5а) и задерживаются там с помощью соответствующих хоминговых рецепторов.
Базофил и тучная клетка синтезируют сходный набор биологически активных веществ. Вырабатывают, накапливая в гранулах, вазоактивные амины (гистамин у человека и серотонин у грызунов), сульфатированные глюкозаминогликаны (хондроитинсульфат, гепарин), ферменты (сериновые протеазы и др.), а также цитокин α-ФНО. Напрямую выделяют в межклеточное пространство лейкотриены (С4, Д4, Е4), простагландины (PGD2, PGE2), цитокины (ИЛ-3, 4, 5, 13 и ГМ-КСФ) и фактор активации тромбоцитов.
На поверхности базофил и тучная клетка несут высокоаффинный FcR к IgE и G4. Образованный рецепторный комплекс специфически взаимодействует с эпитопом антигена/аллергена. Экспрессируют также FcR к IgG в составе иммунного комплекса. Базофил и тучная клетка активируются аллергенами, анафилотоксинами, медиаторами активированных нейтрофилов, норадреналином, ингибируются иммунными комплексами.
Связывание аллергена с рецепторным комплексом вызывает дегрануляцию базофила и тучной клетки - залповый выброс биологически активных соединений, содержащихся в гранулах, в межклеточное пространство, которые вызывают развитие гиперчувствительности немедленного типа (аллергической реакции I типа).
Базофил и тучная клетка направляют дифференцировку Т-хелперов в сторону Т2-субпопуляции и усиливают эозинофилогенез.
Дендритные клетки - отростчатые клетки костно-мозгового происхождения. Локализуются в лимфоидных органах и барьерных тканях. Экспрессируют на своей поверхности MHC II класса и ко-стимулирующие факторы (CD40, 80, 86). Способны погло-
щать путем эндоцитоза, перерабатывать (процессировать) и представлять (презентировать) антиген Т-хелперам в комплексе с MHC II класса. Является наиболее активной АПК. Из числа дендритных клеток хорошо известны клетки Лангерганса (в эпидермисе), интердигитальные клетки (в лимфатических узлах) и дендритные клетки тимуса.
10.2.2. Организация функционирования иммунной системы
Иммунная система имеет сложную организацию - для осуществления специфической функции задействовано множество различных клеточных популяций и растворимых факторов иммунитета. Клетки постоянно циркулируют в организме, погибают в процессе жизнедеятельности и воспроизводятся.
В зависимости от конкретной потребности специфическая функция иммунной системы может быть активирована либо подавлена (супрессирована). Однако любое реагирование иммунной системы осуществляется только при постоянном взаимодействии практически всех типов ее клеток, т.е. в условиях межклеточной кооперации. Раздражителем (активирующим сигналом) является антиген. В развитии любого иммунного реагирования прослеживается каскад последовательно сменяющихся этапов.
10.2.2.1. Взаимодействие клеток иммунной системы
Необходимым условием функционирования иммунной системы является тесная межклеточная кооперация, основу которой составляет рецептор-лигандное взаимодействие. Для связи между собой клетки используют различные дистантные растворимые факторы и прямой контакт.
Синтез растворимых факторов является одним из универсальных способов коммутации клеток между собой. К таковым относятся цитокины, которых в настоящее время известно более 25. Они представляют собой гетерогенное семейство разнообразных по структуре и функции биологически активных молекул, имеющих ряд общих свойств:
• как правило, цитокины не депонируются в клетке, а синтезируются после соответствующего стимула;
• для восприятия цитокинового сигнала клетка экспрессирует соответствующий рецептор, который может взаимодействовать с несколькими различными цитокинами;
• цитокины синтезируются клетками разных ростков, уровней и направлений дифференцировки;
• субпопуляции клеток иммунной системы различаются по спектру синтезируемых цитокинов и их рецепторов;
• цитокины обладают универсальностью, множественностью эффектов и синергизмом;
• цитокины могут воздействовать как на рядом расположенную клетку (паракринная регуляция), так и на сам продуцент (аутокринная регуляция);
• цитокиновая регуляция носит каскадный характер: активация клетки одним цитокином вызывает синтез другого;
• в подавляющем большинстве это короткодистантные медиаторы - их эффекты проявляются на месте выработки. Вместе с тем ряд провоспалительнъгх цитокинов (ИЛ-1, 6, α-ФНО и др.) могут оказывать системное действие.
Цитокины различаются по ведущей функциональной направленности:
• медиаторы доиммунного воспаления (ИЛ-1, 6,12, α-ФНОидр);
• медиаторы иммунного воспаления (ИЛ-5, 9, 10, γ-ИФН
и др.);
• стимуляторы пролиферации и дифференцировки лимфоцитов (ИЛ-2, 4, 13, трансформирующий фактор роста - β-ТФР
и др.);
• факторы роста клеток, или колониестимулирующие факторы
(ИЛ-3, 7, ГМ-КСФ и др.);
• хемокины, или клеточные хемоаттрактанты (ИЛ-8 и др.). Краткая характеристика некоторых цитокинов приведена в
табл. 10.3.
Прямое межклеточное взаимодействие основано на рецепции структур, экспрессированных на мембране клетки-оппонента. Для этого требуется достаточно продолжительный и стабильный контакт клеток. Такой способ коммутации используют Т-хелперы и Т-киллеры при анализе чужеродности презентированных структур. Механизм действия ко-стимулирующих факторов (пары CD40- CD40-лиганд, CD28-CD80, 86) также требует непосредственного контакта.
10.2.2.2. Активация иммунной системы
Активация иммунной системы подразумевает развитие продуктивной иммунной реакции в ответ на антигенное раздражение
Таблица 10.3. Характеристика основных цитокинов
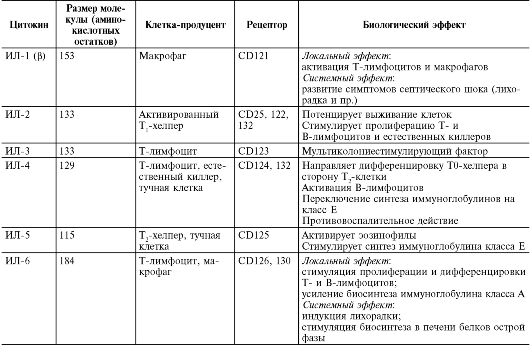 Продолжение табл. 10.3
Продолжение табл. 10.3
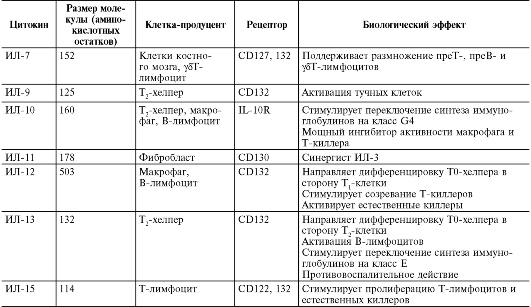 Продолжение табл. 10.3
Продолжение табл. 10.3
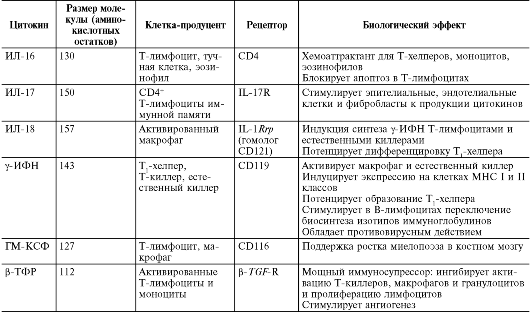 Окончание табл. 10.3
Окончание табл. 10.3
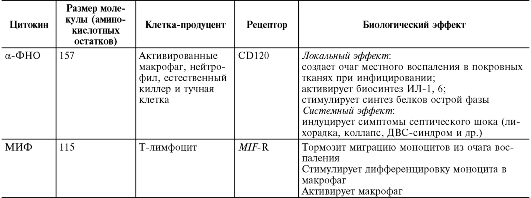 Примечание. МИФ - миграцию ингибирующий фактор.
Примечание. МИФ - миграцию ингибирующий фактор.
и появление продуктов деструкции тканей макроорганизма. Это сложный многоступенчатый процесс, требующий продолжительного времени для своей индукции - около 4 сут. Критическим событием является невозможность элиминации антигена факторами врожденного иммунитета в течение указанного срока.
Пусковым механизмом адаптивного иммунитета является распознавание «свой-чужой», которое осуществляют Т-лимфоциты при помощи своих прямых иммунорецепторов - TCR. В случае установления чужеродности биоорганической молекулы включается второй этап реагирования - запускается интенсивное тиражирование клона высокоспецифичных для антигена лимфоцитовэффекторов, способных прервать антигенную интервенцию. Это явление получило название «экспансия клона». Параллельно, но несколько позже пролиферации стимулируются дифференцировка иммунных лимфоцитов и формирование из него клеток иммунологической памяти, гарантирующих выживание в будущем.
Таким образом, продуктивная активация иммунной системы связана с размножением и дифференцировкой антигенореактивных клонов иммунокомпетентных клеток. Антигену в этом процессе отведена роль индуктора и фактора клональной селекции. Механизмы основных этапов активации иммунной системы рассмотрены ниже.
Активация Т-хелпера. Процесс (см. рис. 10.6) осуществляется при непосредственном участии АПК (дендритные клетки, В-лимфоциты и макрофаги). После эндоцитоза и процессинга антигена во внутриклеточных везикулах АПК встраивает образовавшийся олигопептид в молекулу MHC II класса и выставляет полученный комплекс на наружной мембране. На поверхности АПК также экспрессируются ко-стимулирующие факторы - молекулы CD40, 80, 86, мощным индуктором которых являются продукты разрушения покровных тканей на этапе доиммунного воспаления.
Т-хелпер при помощи молекул адгезии прочно соединяется с поверхностью АПК. Иммунорецептор Т-хелпера совместно с молекулой CD3 при поддержке ко-рецепторной молекулы CD4 взаимодействует с комплексом антиген-MHC II класса и анализирует чужеродность его структуры. Продуктивность рецепции зависит от ко-стимулирующих воздействий в парах CD28-CD80/86 и CD40- лиганд-CD40.
В случае признания чужеродности комплекса антиген-MHC II класса (точнее, «не своего») Т-хелпер активируется. Он экспресси-
рует рецептор к ИЛ-2 и начинает синтезировать ИЛ-2 и другие цитокины. Итогом активации Т-хелпера являются его размножение и дифференцировка в одного из своих потомков - T1- или Т2-хелпер (см. рис. 10.2). Любое изменение условий рецепции прекращает активацию Т-хелпера и может индуцировать в нем апоптоз.
Активация В-лимфоцита. Для активации В-лимфоцита (рис. 10.9) необходима суммация трех последовательных сигналов. Первый сигнал - результат взаимодействия молекулы антигена со специфичным для него BCR, второй - интерлейкиновый стимул активированного Т-хелпера и третий - результат взаимодействия ко-стимулирующих молекул CD40 с CD40-лигандом.
Активация инициирует размножение и дифференцировку специфичного для конкретного антигена В-лимфоцита (см. рис. 10.2). В итоге в пределах зародышевых (герминативных) центров лимфоидных фолликулов появляется клон специфических антителопродуцентов. Дифференцировка позволяет переключить биосинтез иммуноглобулинов с классов M и D на более экономные: G, A или Е (редко), повысить аффинность синтезируемых антител и образовать В-клетки иммунологической памяти или плазматические клетки.
Активация В-лимфоцита - весьма тонкий процесс. Отсутствие хотя бы одного из стимулов (нарушение межклеточной кооперации, неспецифичность рецептора В-лимфоцита или элиминация антигена) блокирует развитие антительного иммунного ответа.
Активация Т-киллера. Для исполнения надзорной функции Т-киллер вступает в тесный и прочный контакт с потенциальной
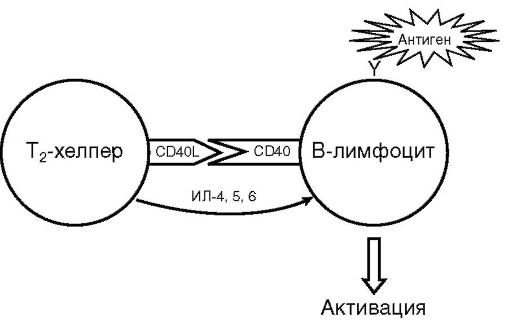 Рис. 10.9. Схема активации В-лимфоцита (пояснения в тексте)
Рис. 10.9. Схема активации В-лимфоцита (пояснения в тексте)
клеткой-мишенью, используя молекулы адгезии (см. рис. 10.8). Затем иммунорецептор Т-киллера (αβTCR) совместно с молекулой CD3 при поддержке ко-рецепторной молекулы CD8 взаимодействует с антигенным комплексом MHC I класса и анализирует его структуру. Обнаружение отклонений в пользу аллогенности активирует Т-киллер к экспрессии рецептора к ИЛ-2 и синтезу ИЛ-2 и высвобождение эффекторных молекул (перфорин, гранзимы, гранулизин) из цитоплазматических гранул в синаптическую щель межклеточного контакта.
Для адекватного развития клеточной формы иммунного ответа требуются активизирующие стимулы со стороны Т1-хелпера. Т-киллер может функционировать автономно, самостоятельно инициируя и поддерживая клонообразование за счет аутокринной стимуляции ИЛ-2. Однако это свойство реализуется редко.
10.2.2.3. Супрессия иммунного ответа
Супрессия или подавление иммунного ответа является физиологической реакцией организма, которая в норме завершает иммунный ответ и направлена на торможение экспансии антигенспецифических клонов лимфоцитов. В отличие от иммунологической толерантности, супрессии подвергается уже инициированное иммунное реагирование. Различают три механизма иммуносупрессии: уничтожение клонов иммунокомпетентных клеток, торможение активности иммунокомпетентных клеток, элиминация антигенного стимула.
Устранить иммунокомпетентные клетки можно путем апоптоза. При этом элиминации подвергаются следующие группы клеток:
• терминально дифференцированные лимфоциты, завершившие свою биологическую программу;
• активированные лимфоциты, не получившие антигенного стимула;
• «изношенные» лимфоциты;
• аутореактивные клетки.
Естественными факторами, инициирующими апоптоз, яляются глюкокортикоидные гормоны, Fas-лиганд, α-ФНО и другие иммуноцитокины, гранзимы и гранулизин. Апоптотическое уничтожение клеток-мишеней могут активировать Т-киллеры, ЕК с фенотипом CD16-CD56много и Т1-хелперы.
Помимо апоптоза возможен антителозависимый лимфоцитолиз. Например, с медицинской целью применяют антилимфоцитарную
сыворотку, которая в присутствии комплемента вызывает лизис лимфоцитов. Устранить лимфоидную популяцию возможно также воздействием ионизирующего излучения или цитостатиков.
Функциональная активность иммунокомпетентных клеток может быть ингибирована растворимыми факторами их конкурентов или потомков. Ведущая роль принадлежит иммуноцитокинам с множественными эффектами. Известно, например, что Т2- хелперы, γδТ-лимфоциты и тучные клетки при помощи ИЛ-4, 13 препятствуют дифференцировке Т0-хелпера в Т1-клетку. Последний, в свою очередь, может блокировать образование Т2-хелпера, синтезируя γ-ИФН. Пролиферацию Т- и В-лимфоцитов ограничивает β-ТФР, который продуцируют терминально дифференцированные Т-хелперы. Уже упомянутые продукты Т2-хелпера (ИЛ-4, 13 и β-ТФР) подавляют биологическую активность макрофагов.
Супрессия гуморального звена иммунитета может быть вызвана иммуноглобулинами. Избыточные концентрации иммуноглобулина класса G, связываясь со специальными рецепторами на мембране В-лимфоцита, тормозят биологическую активность клетки и ее способность дифференцироваться в плазмоцит.
Устранение из организма антигена в природе наблюдается при полном освобождении организма от патогена при развитии стерильного иммунитета. В клинической практике эффект достигается очищением организма плазмоили лимфосорбцией, а также нейтрализацией антигена антителами, специфичными для высокоиммуногенных эпитопов.
10.2.2.4. Возрастные изменения иммунной системы
В развитии иммунной системы четко прослеживаются два этапа. Первый, антигеннезависимый, который начинается с эмбрионального периода развития и частично продолжается всю жизнь. В течение этого периода образуются стволовые клетки и разнообразные антигенспецифические клоны лимфоцитов. Предшественники γδT и В1-лимфоцитов мигрируют в покровные ткани и формируют автономные лимфоидные ростки.
Второй этап, антигензависимый, продолжается с момента рождения особи до ее гибели. В этот период идет «ознакомление» иммунной системы с многообразием окружающих нас антигенов. По мере накопления биологического опыта, т.е. количества и качества продуктивных контактов с антигенами, происходят селекция
и тиражирование отдельных клонов иммунокомпетентных клеток. Особенно интенсивная экспансия клонов характерна для детского возраста. В течение первых 5 лет жизни иммунной системе ребенка приходится усваивать примерно 90% биологической информации. Еще 9% воспринимается до наступления пубертата, на взрослое состояние остается лишь около 1%.
Иммунной системе ребенка приходится справляться с чудовищными нагрузками, которые в основном падают на гуморальное звено иммунитета. В местах с повышенной плотностью населения и частыми межиндивидуальными контактами (крупные города) создаются условия для длительной персистенции высокой концентрации разнообразных патогенов. Поэтому дети в мегаполисах часто болеют. Однако создается впечатление о тотальном иммунодефиците, порожденном крайним экологическим неблагополучием. Между тем эволюционно заложенные механизмы иммунной защиты позволяют организму ребенка успешно справиться с трудными естественными испытаниями на жизнеспособность и адекватно отреагировать на вакцинопрофилактику.
С возрастом иммунная система меняет свою структуру. В организме взрослого до 50% всего лимфоидного пула представлено клонами клеток, прошедших антигенную стимуляцию. Накопленный иммунной системой биологический опыт проявляется образованием узкой «библиотеки» жизненно важных (актуальных) клонов лимфоцитов, специфичных для основных патогенов. Благодаря долгоживучести клеток иммунологической памяти актуальные клоны со временем становятся самодостаточными. Они приобретают способность к самоподдержанию и независимость от центральных органов иммунной системы. Функциональная нагрузка на тимус снижается, что проявляется его возрастной инволюцией. Тем не менее в организме сохраняется широкий набор невостребованных «наивных» клеток. Они способны отреагировать на любую новую антигенную агрессию.
Структура популяции Т-лимфоцитов также претерпевает возрастные изменения. Установлено, что в организме новорожденных преобладают Т2-хелперы, необходимые для развития антительной защиты. Однако со временем перед организмом все острее встает проблема внутриклеточного паразитизма, различных инвазий, мутаций, что требует надежного и хорошо организованного иммунологического надзора за морфогенетическим постоянством кле-
точных элементов организма. Поэтому после рождения начинает усиленно развиваться система адаптивного клеточного иммунитета, а вместе с ним образование клонов Т1-хелперов и Т-киллеров. Отмечено, что нарушение постнатальной колонизации желудочнокишечного тракта нормальной флорой тормозит процесс адекватного формирования популяции Т1-хелперов в пользу Т2-клеток. Избыточная активность последних оборачивается аллергизацией детских организмов.
Продуктивный иммунный ответ после своего завершения (нейтрализации и элиминации антигена из организма) также сопровождается изменениями клональной структуры антигенореактивных лимфоцитов. При отсутствии активирующих стимулов клон инволюционирует. Невостребованные клетки со временем погибают от старости или индукции апоптоза, причем этот процесс начинается с более дифференцированных лимфоцитов-эффекторов. Численность клона постепенно снижается и проявляется постепенным угасанием иммунного ответа. Однако в организме длительно персистируют клетки иммунологической памяти.
Старческий период жизни характеризуется доминированием в иммунной системе актуальных клонов антигенспецифических лимфоцитов в сочетании с нарастающей иммунодепрессией и снижением общей реактивности. Инфекции, вызванные даже условно-патогенными микробами, зачастую принимают затяжной или угрожающий характер. Клеточный иммунитет также теряет эффективность, постепенно нарастает объем злокачественно трансформированных клеток. Поэтому у пожилых людей часто встречаются новообразования.
Задания для самоподготовки (самоконтроля)
А. Отметьте эффекторные клетки иммунной системы:
1. Дендритные клетки.
2. В-лимфоциты.
3. Т-хелперы.
4. Т-киллеры. Б. Отметьте АПК:
1. Дендритные клетки.
2. В-лимфоциты.
3. Макрофаги.
4. Т-хелперы.
В. Отметьте клетки, на которых экспрессируется рецептор 2-го класса МНС:
1. Т-киллеры.
2. Дендритные клетки.
3. Макрофаги.
4. В-лимфоциты.
Г. Отметьте маркеры В-лимфоцитов:
1. МНС 2-го класса.
2. CD40.
3. CD80.
4. CD28.
Д. Отметьте рецепторные молекулы Т-хелперов:
1. CD4.
2. CD3.
3. CD28.
4. CD40L.
Е. Назовите клетки и медиаторы, принимающие участие в формировании Т1-хелперов:
1. ИЛ-12.
2. Т-киллеры.
3. γ-Интерферон.
4. Активированный макрофаг.
5. Тучная клетка.
Ж. Назовите клетки и медиаторы, принимающие участие в формировании Т2-хелперов:
1. Базофилы.
2. Т-киллеры.
3. Тучные клетки.
4. ИЛ-4.
5. ФНО.
З. Назовите рецептор-лигазную пару, необходимую для костимуляции Т-хелперов АПК. Без этой ко-стимуляции представление антигена Т-хелперу может привести к его функциональной инактивации:
1. CD80/CD28.
2. MHC класс2/CD4.
3. MHC класс1CD8.
4. MHC класс2/TCR
И. Назовите рецептор-лигазную пару, необходимую для стимуляции Т-киллера (CD8):
1. MHC класс 2/CD4.
2. MHC класс 1/CD8.
3. CD40/CD40L.
4. CD80/CD28.
К. Некоторые вирусы и бактериальные токсины обладают свойством суперантигенов, вызывая неспецифическую активацию лимфоцитов, приводящую их к гибели. Объясните механизм их действия.