Иммунология : учебник. - Хаитов Р. М. - 2009. - 320 с. : ил.
|
|
|
|
ГЛАВА 8 ЭФФЕКТОРНЫЕ МЕХАНИЗМЫ ИММУНИТЕТА
Эффекторные механизмы иммунитета состоят в том, что TCR на поверхности T-лимфоцита и/или иммуноглобулины в растворе физически подводят связанный Аг к таким клеткам или ферментам, которые специально предназначены для расщепления Аг до мелких метаболитов, которые организм может вывести через свои системы выделения (почки, ЖКТ).
В соответствии с двумя типами АГ-связывающих Рц есть и два типа эффекторных механизмов:
• антителозависимые, ранее их называли гуморальным иммунитетом;
• T-лимфоцитзависимые (антителонезависимые), «по старинке» - клеточный иммунитет.
АНТИТЕЛОЗАВИСИМЫЕ МЕХАНИЗМЫ ЗАЩИТЫ Компоненты комплемента
Сам по себе факт связывания патогена АТ является защитным по крайней мере в двух случаях: 1 если патоген - сильный яд, то АТ при связывании нейтрализует токсичность; 2 если патоген инфекционен (вирус, прион, бактерия), то АТ, связав его, препятствует распространению патогена в организме. Но и в этих случаях защитная реакция не заканчивается образованием макромолекулярных комплексов «Аг-АТ». Эти комплексы необходимо расщепить до мелких метаболитов. Для этого АТ фиксируют и активируют компоненты комплемента (IgM >IgG3 >IgG1). Комплексы «Аг-АT-компоненты комплемента» транспортируются эритроцитами, имеющими Рц для компонентов комплемента, в синусоиды селезёнки и печени, где их фагоцитируют и расщепят макрофаги.
Fc-рецепторы
FcR - мембранные молекулы, специфически связывающие Fc- фрагменты Ig. Их (наряду с TCR и BCR) можно отнести к иммуно-
рецепторам, так как клетка-носитель FcR способна связать Аг (пусть и через посредство АТ) и прореагировать на него. FcR присутствуют как на лимфоцитах, так и на всех известных лейкоцитах.
Типы и разновидности FcR. По изотипу связываемых тяжёлых цепей Ig выделяют 4 разновидности FcR: γ, ε, α, μ; а по аффинности связи с лигандом - три типа FcR: I, II и III. Рц типа I способны связывать свободные молекулы АТ (это особенно характерно для IgE), Рц типов II и III - только комплексы «Аг-АТ».
Антителозависимая клеточная цитотоксичность
Феномен АТ-зависимой клеточной цитотоксичности (АЗКЦТ) имеет место, когда АТ связывает Аг на поверхности какой-либо клетки-мишени и через Fc-фрагмент привлекает для деструкции клетки-мишени опредёленные клетки-исполнители (NK и эозинофилы).
• Естественные киллеры. NK имеют Рц (FcγRIII) для Fc-фрагментов IgG. Механизм собственно киллерного действия NK на клеткумишень такой же, как и киллерный механизм ЦТЛ - перфорингранзимовый (см. рис. 1.4 и 6.4).
• Эозинофилы. Механизм санации от гельминтов - это тоже АЗКЦТ, где в качестве эффекторных клеток выступают эозинофилы, имеющие низкоаффинные Рц для IgE - FcεRII и для IgA - FcαRII. Связывание этих Рц с лигандами в сочетании с сигналом от цитокина ИЛ-5 стимулирует синтез и секрецию эозинофилом высокотоксичных белков, направленных на уничтожение гельминтов. Другими словами, активированный эозинофил секретирует ряд биологически активных веществ (БАВ), свойства которых объясняют симптоматику так называемых эозинофильных воспалительных процессов (табл. 8.1).
Таблица 8.1. Биологически активные продукты эозинофилов
 Продолжение табл. 8.1.
Продолжение табл. 8.1.
 T-лимфоцитзависимое
(опосредованное ИЛ-5) иммунное воспаление тканей с участием эозинофилов
характерно для аллергических заболеваний, васкулитов, а также
встречается при отторжении трансплантатов чужеродных органов.
T-лимфоцитзависимое
(опосредованное ИЛ-5) иммунное воспаление тканей с участием эозинофилов
характерно для аллергических заболеваний, васкулитов, а также
встречается при отторжении трансплантатов чужеродных органов.
Гиперчувствительность немедленного типа
Сосудистые и миоконстрикторные реакции, опосредованные медиаторами тучных клеток и базофилов, приводят к развитию гиперчувствительности немедленного типа.
Базофильные лейкоциты и тучные клетки. В этих реакциях АТ привлекают к «работе» базофилы и тучные клетки. Существенные функциональные признаки этих клеток схожи: наличие на мембране высокоаффинного Рц для IgE (FcεRI), и одинаковый список биологически активных медиаторов.
• Тучные клетки локализованы в соединительной ткани собственного слоя слизистых оболочек, в подкожной соединительной ткани и соединительной ткани по ходу всех кровеносных сосудов, т.е. «подстилают» различные барьеры. Выделяют по крайней мере две тканевые разновидности тучных клеток.
♦ Тучные клетки слизистой оболочки из сериновых протеаз экспрессируют и триптазу, и химазу, секретируют минимум гистамина; из протеогликанов в них преобладает хондроитинсульфат; из метаболитов арахидоновой кислоты - лейкотриен C4 (LTC4). По-видимому, дифференцировка этих клеток зависит от T-
лимфоцитов, а именно от местной стимуляции клеток-предшественниц цитокином ИЛ-3.
♦ Тучные клетки соединительной ткани локализованы в серозных оболочках полостей тела и в лёгких. Из сериновых протеаз экспрессируют преимущественно триптазу, из протеогликанов - гепарин, секретируют много гистамина, из метаболитов арахидоновой кислоты в них преобладает простагландин D2. Дифференцировку этих тучных клеток стимулируют фибробласты.
• Базофилы циркулируют в крови и мигрируют в ткани только в очаг воспаления (как нейтрофилы). На базофилах экспрессированы молекулы адгезии, важные для хоминга в очаг: LFA-1 (CD11a/ CD18), Mac-1 (CD11b/CD18), CD44.
• Активация. Следующие сигналы активируют и тучные клетки, и
базофилы:
♦ Гомотипная агрегация FcεRI (активируют комплекс IgE с Аг или АТ к Рц);
FcεRI способен связывать свободные АТ класса Е (до того, как они свяжут свой Аг в комплекс). Клетки с комплексом «IgE-FcεRI» на мембране находятся «на низком старте» и готовы в считанные секунды и минуты выбросить содержимое гранул в ответ на поступивший Аг (рис. 8.1).
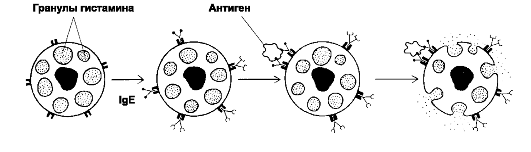 Рис. 8.1. Дегрануляция тучных клеток. На
поверхности тучных клеток молекулы IgE связаны Fc-рецепторами. Аг
взаимодействует с Fab-фрагментами IgE. Активированная этим сигналом
тучная клетка подвергается дегрануляции.
Рис. 8.1. Дегрануляция тучных клеток. На
поверхности тучных клеток молекулы IgE связаны Fc-рецепторами. Аг
взаимодействует с Fab-фрагментами IgE. Активированная этим сигналом
тучная клетка подвергается дегрануляции.
♦ Анафилатоксины - активированные компоненты комплемента (C5a >C4a >C3a).
♦ Медиаторы из активированных нейтрофилов.
♦ Нейромедиаторы (норадреналин, вещество Р).
• Медиаторы тучных клеток и базофилов различны по биохимическим свойствам, по предназначению и по срокам выброса из
активированной тучной клетки. Медиаторы, депонируемые в гранулах, первыми выбрасываются из клетки по активирующему сигналу. Другие медиаторы синтезируются de novo и вступают в процесс позже.
♦ Гистамин. На разных клетках есть различные Рц для гистамина - Н1, Н2 и Н3. Вазоактивные эффекты гистамина состоят в следующем: эндотелиальные клетки претерпевают констрикцию, и сыворотка выходит из сосуда в ткани; гистамин стимулирует синтез в клетках эндотелия простациклина и радикала оксида азота (NO), которые вызывают расслабление гладких мышц сосудистой стенки и, следовательно, вазодилатацию.
- Если процесс происходит в коже, то клинически он проявляется в виде волдырей и покраснения (крапивница). В случае аллергической патологии снять симптомы помогают лекарственные средства (ЛС) - блокаторы Н1-Рц для гистамина.
- Если гистамина выделяется достаточно много, то он вызывает клинически значимые сокращения гладких мышц кишки (перистальтику) и бронхов (бронхоспазм), но этот эффект непродолжителен, так как гистамин быстро распадается во внеклеточной среде.
♦ Липидные медиаторы. При стимуляции тучных клеток в них активируются ферменты метаболизма липидов, а именно фосфолипаза А2. Этот фермент использует в качестве субстратов фосфолипиды клеточных мембран и липиды (в первую очередь арахидоновую кислоту). В результате образуются следующие биологически активные медиаторы.
- Простагландин D2 - действует как вазодилататор и бронхоконстриктор. В биосинтезе простагландина D2 из арахидоновой кислоты участвует циклооксигеназа. Фармакологическими ингибиторами этого фермента являются ацетилсалициловая кислота и ряд других нестероидных противовоспалительных средств.
- Лейкотриены (LTC4, LTD4, LTE4) - альтернативные продукты метаболизма арахидоновой кислоты, которые образуются под воздействием 5-липооксигеназы. Комплекс лейкотриенов - медленно реагирующая субстанция анафилаксии. Именно этот комплекс медиаторов в наибольшей мере ответственен за бронхоконстрикцию при бронхиальной астме. Это объясняет усугубление астматических приступов ацетилсалициловой кислотой: блокируя синтез простагландина D2, ацетилсали-
циловая кислота высвобождает метаболический шунт арахидоновой кислоты в пользу лейкотриенов.
- Фактор активации тромбоцитов (ФАТ) вызывает бронхоконстрикцию, а также релаксацию гладких мышц сосудов и ретракцию эндотелия. ФАТ продуцируют не только (а может быть и не столько) тучные клетки, сколько клетки эндотелия, стимулированные гистамином и лейкотриенами.
- Ферменты тучных клеток и базофилов - сериновые протеазы (триптаза и химаза), катепсин G, карбоксипептидаза - способствуют перестройке матрикса соединительной ткани.
- Цитокины тучных клеток и базофилов. К ним относятся интерлейкины и GM-CSF.
• Гиперчувствительность немедленного типа (ГНТ). Цитокины тучных клеток и базофилов поддерживают иммунное отклонение в дифференцировке субпопуляций CD4+ T-лимфоцитов в пользу Th2 (ИЛ- 4, ИЛ-13), а также поддерживают дифференцировку и активацию эозинофилов (ИЛ-5, ИЛ-3, GM-CSF). В случае патологии именно эти клетки (Th2, IgE, тучные клетки, базофилы, эозинофилы) и IgE составляют «дружный» и самоподдерживающийся ансамбль, ответственный за реакции гиперчувствительности немедленного типа. Мишени для цитокинов - гладкие мышцы и эндотелий (следовательно, сосуды, бронхи, ЖКТ). Системная реакция ГНТ - анафилактический шок.
Реликтовые свойства антител
Помимо связывания Аг, комплемента и FcR, молекулы Ig имеют ещё несколько свойств (видимо, унаследованных от каких-то предковых белков).
• Пептидазная активность - в некоторых случаях АТ сами расщепляют свой Аг, функционируя как протеазы.
• Способность связывать нуклеотиды выявлена у доменов VL и VH: пуриновое кольцо связывается с остатками триптофана и тирозина. При этом связи с нуклеотидами могут не мешать связыванию с Аг в Аг-связывающем центре. Некоторые АТ, связывающие нуклеиновые кислоты, обладают способностью расщеплять полинуклеотиды (ДНКазная активность).
• Способность связывать металлы (например, ртуть и свинец). Это свойство АТ может иметь отношение к патогенезу заболеваний: АТ, связывая жизненно необходимые ионы металлов-микроэлементов, вызывают дефицит этих металлов во внутренней среде.
Подобные подозрения возникли в отношении склеродермии и АТ, связывающих свинец.
• Способность связывать суперантигены. Свойства суперантигенов для иммуноглобулинов описаны для 3 веществ: белка А стафилококка (SpA), gp120 ВИЧ-1 и кишечного сиалопротеина. Один такой суперантиген может связать более 80% всех иммуноглобулинов крови, при этом иммуноглобулин теряет способность связывать свой специфический Аг. Считают, что таков один из компонентов патогенеза иммунодефицита при ВИЧ-инфекции.
T-ЛИМФОЦИТЗАВИСИМЫЕ ЭФФЕКТОРНЫЕ МЕХАНИЗМЫ
T-лимфоцитзависимые (антителонезависимые) эффекторные механизмы иммунитета реализуют цитотоксические T-лимфоциты (ЦТЛ). К ним относят CD8+ Tαβ-лимфоциты и часть CD4+. Есть T-киллеры и среди Tуδ.
Главное предназначение ЦТЛ - санация организма от внутриклеточных инфекций, реализуемая киллерной функцией ЦТЛ и цитокинами.
• Киллерная функция. ЦТЛ осуществляют киллерную функцию (см. рис. 1.4 и 6.4) с помощью особых белков - цитотоксинов, к которым относят перфорин, гранзимы и недостаточно изученный цитолизин.
♦ Синтез цитотоксинов происходит de novo после вовлечения ЦТЛ в иммунный ответ и распознавания ими своего Аг.
♦ Накопление цитотоксинов. В виде функционально неактивных молекул-предшественников цитотоксины накапливаются в гранулах, которые сконцентрированы вблизи TCR.
♦ Дегрануляция ЦТЛ происходит в области межклеточного контакта, образующегося при связывании TCR с Аг на поверхности клетки-мишени. Этот процесс облигатно зависит от Ca2+.
♦ Перфорин накапливается в гранулах в виде растворимого предшественника. Будучи высвобожденным из гранул и в присутствии Ca2+, перфорин в считанные секунды полимеризуется в мембране клетки-мишени: липофильные участки молекул перфорина ориентируются наружу, гидрофобные - внутрь клетки. В результате образуется пора диаметром около 16 нм.
♦ Гранзимы и апоптоз. Через пору, образованную перфорином, ЦТЛ инъецирует внутрь клетки-мишени гранзимы. Охарактеризовано 3 гранзима ЦТЛ - А, В и С. Это специализированные
сериновые протеазы, субстратами которых являются ферменты, инициирующие в клетке-мишени программу апоптоза. При этом происходит разрушение ДНК и белков не только клетки, но и инфицировавшего её вируса.
♦ Лизис мишени. Если в механизмах апоптоза клетки-мишени есть какие-либо дефекты, ЦТЛ всё равно разрушает клетку - путём осмотического лизиса через сформированные перфорином поры. Однако в этом случае неповреждённые вирусные частицы и нуклеиновые кислоты могут заразить другие клетки, что и происходит в случаях наиболее «коварных» инфекций.
• Цитокины. CD8+ ЦТЛ продуцируют цитокины - ИФНγ , TNFα и TNFβ (лимфотоксин). В частности, ИФНγ проявляет следующие эффекты:
♦ прямо ингибирует репликацию вирусов;
♦ индуцирует в клетках-мишенях повышенную экспрессию молекул MHC-I и -II, способствуя более эффективному представлению вирусных Аг T-лимфоцитам (и для распознавания, и для киллинга);
♦ активирует макрофаги и NK;
♦ наконец, ИФНу - кофактор индукции дифференцировки иммунных CD4+ T-лимфоцитов в Th1, а последнее означает, что CD8+ ЦТЛ вносят свой вклад в развитие других эффекторных механизмов иммунного ответа - варианта с участием Th1.
ГИПЕРЧУВСТВИТЕЛЬНОСТЬ ЗАМЕДЛЕННОГО ТИПА
Гиперчувствительность замедленного типа (ГЗТ) - иммунное воспаление тканей, которое «организуют» CD4+ T-лимфоциты субпопуляции Th1 - продуценты ИФНγ, а клетками-исполнителями являются активированные макрофаги. Если макрофаг в очаге инфекции активирован взаимодействием с иммунным CD4+ Th1-лим- фоцитом, то микробицидные возможности макрофага существенно повышаются, и активированный макрофаг с большой вероятностью разрушит патогены (к сожалению, в макрофаге погибают не все патогены, особенно жизнеспособны вирусы, например ВИЧ).
• Активация макрофага. Для иммунной активации макрофага необходимы 2 типа воздействия на макрофаг со стороны лимфоцитов: - контактное: молекула CD40L на Th1-лимфоците вступает в связь с молекулой CD40 на макрофаге; - цитокиновое: ИФНγ, продуцируемый Th1, CD8+ ЦТЛ или NK, связывает Рц на макрофаге.
♦ Инфицированный макрофаг имеет больше шансов вступить во взаимодействие с иммунным Th1, так как последний своим TCR свяжет Аг именно на поверхности макрофага и именно на него же направит свои ИФН и CD40L.
♦ Ингибитор активации макрофагов - ИЛ-10.
• Характеристики активированного макрофага. Макрофаг, активированный взаимодействием с Th1, приобретает следующие признаки и функциональные способности.
♦ Увеличивается число иммунорецепторов FcyR, которыми макрофаг связывает комплексы Аг-АТ и фагоцитирует их.
♦ ИФНу в макрофагах индуцирует биосинтез ферментов, генерирующих радикалы активных форм кислорода, которые окисляют фагоцитированный Аг.
♦ В макрофагах под воздействием ИФНу, TNFa и, возможно, ИЛ-1 индуцируется экспрессия NO-синтазы, продуцирующей радикал NO-, который также окисляет фагоцитированный материал.
♦ В макрофагах индуцируется синтез липидных медиаторов воспаления - ФАТ, простагландинов и лейкотриенов (LTE4).
♦ Макрофаг синтезирует тканевый фактор коагуляции. В начавшемся процессе коагуляции активируется сывороточный тромбин - протеаза, стимулирующая клетки эндотелия сосудов, а также нейтрофилы к синтезу ФАТ, что ещё больше способствует прогрессированию воспалительного процесса.
♦ ИФНу является самым сильным из известных индукторов синтеза и экспрессии молекул MHC-II. Кроме того, на активированных макрофагах (в отличие от неактивированных) индуцируется экспрессия костимуляторной молекулы B7, что делает активированные макрофаги более эффективными АПК. Кроме того, на активированных макрофагах возрастает экспрессия молекул адгезии ICAM-1 и LFA-3.
♦ Активированные макрофаги продуцируют ИЛ-12, который способствует дифференцировке Th1.
• Очаг воспаления. Цитокины активированных макрофагов - TNFa, ИЛ-1 и хемокины - создают очаг воспаления в виде плотных на ощупь узелков разного размера (симптом индурации). Плотность очага обусловлена выпотом из сосудов фибриногена и полимеризацией его в фибрин. Среди клеток, присутствующих в очаге, в первые 6-8 ч преобладают нейтрофилы, затем макрофаги и Th1. Плотность клеток в свежем очаге ГЗТ невелика.
• Сроки развития реакции. ГЗТ названа замедленной, так как между моментом попадания Аг в ткань и развитием характерного очага плотного воспаления проходит не менее 24-48 ч. После связывания Аг TCR Th1 примерно один час требуется для индукции первых биосинтезов цитокинов, а также синтеза и экспрессии на мембране молекулы CD40L.
• Эффекты факторов роста. Среди цитокинов, продуцируемых активированными макрофагами, присутствуют факторы роста, что может значительно изменить состояние прилегающих к очагу тканей. Стандартная защитная реакция - развитие очага воспаления по типу ГЗТ, но в патологических случаях цитокины из активированных макрофагов вызывают фиброзное перерождение тканей: тромбоцитарный фактор роста (Platelet-derived Growth Factor) стимулирует пролиферацию фибробластов, а вырабатываемый Т4- лимфоцитами и макрофагами ТФР-β стимулирует синтез коллагена. Кроме того, факторы роста из макрофагов вызывают образование дополнительных кровеносных сосудов - ангиогенез.