Иммунология : учебник. - Хаитов Р. М. - 2009. - 320 с. : ил.
|
|
|
|
ГЛАВА 16 ПОСЛЕСЛОВИЕ
Это послесловие адресовано главным образом молодым учёным, искренне любящим науку, любящим больше, чем себя в науке, признающих превосходство природы, не нами созданной, над волей человека, переделывающего природу «здесь и сейчас», понимающих, что точный прогноз последствий - это только горделивая иллюзия. Годами обдумывая всё новые и новые экспериментальные и клинические данные, касающиеся иммунитета и иммунной системы, мы пришли к интересному выводу более общего назначения: сколько бы люди ни узнавали нового, количество непознанного не уменьшается хотя бы потому, что сам мир постоянно меняется.
Мы скажем несколько слов о контурах общей теории иммунитета, которые начали просматриваться в конце XX века. Это совсем ещё «младенец», его легко раздавить под грузом сложившихся ранее многолетних представлений. Он никому не бросает вызов и никого не вызывает на научный бой. Но только в таких быстро развивающихся и неканонизированных областях знаний, как иммунология, и могут (а точнее, не могут не) зарождаться с той или иной частотой «младенцы» общих теорий. Почему именно в названные годы начали проступать контуры общей теории иммунитета?
Потому, что к концу XX в. людьми, работающими в науке, был достигнут методический предел исследования биологических объектов - секвенирование генома, клонирование генов, трансгенные организмы и организмы с нокаутом генов. Дальнейшее разложение объекта исследования на составляющие - расщепление гена - уже есть потеря кода биологической индивидуальности, т.е. уже не биология, а химия, физика, возможно что-то ещё. Новый уровень биологического мышления в постгенно-инженерно-клональный период развития человеческой культуры начинает возвращаться к попыткам осознания организма как целого и не только и не столько в пределах отдельной особи, сколько в понимании всей земной природы как це-
лостной системы. Материальный маркёр единства всех форм жизни на Земле - нуклеиновые кислоты - единый код (как у любого ничтожного вируса, так и у всех великих организмов мира сего). Функциональный маркёр единства всех форм жизни на Земле - поразительное постоянство биологических часов: у каждого вида живых организмов свой естественный срок пребывания в живых, а колебания от особи к особи в пределах вида ничтожны. Поэтому можно не слишком опасаться пагубных последствий от работ по клонированию человека и животных. Природный «буфер» биологических часов скорее всего будет надёжной ловушкой для продуктов искусственного клонирования, защищающей естественно развивающиеся организмы в конкуренции с искусственно созданными. Общая теория иммунитета, таким образом, не может не учитывать параметры эволюционного сопряжения многоклеточных организмов, имеющих иммунную систему, с микроорганизмами-патогенами, от которых иммунитет и защищает. На самом деле почти всё, что мы собираемся сформулировать ниже, присутствует в основном тексте учебника с объяснениями и с аргументацией (схема иммунного ответа и другие разделы). Поэтому здесь мы обойдёмся почти без объяснений, изложим свой материал тезисно.
Итак, контуры общей теории иммунитета. Иммунитет - эволюционно самый новый биологический механизм защиты многоклеточных организмов от инфекций. Иммунная система есть примерно у 1,5% видов живых организмов на Земле (начиная с челюстных рыб). Материальные носители иммунитета - лимфоциты. Согласно гипотезе Чарльза Джанвея, лимфоцитарный иммунитет возник в результате ретровирусной инфекции в гаметы древних челюстных рыб, привнёсшей в их геном механизм перестройки соматической ДНК в определённых участках. Современный остаток этих ретровирусов - гены RAG, функционирующие только в лимфоцитах. Отбор закрепил это свойство, и мы изучили его в XX в. как Рц T- и B-лимфоцитов для Аг.
У многоклеточных есть ряд биологических механизмов защиты от инфекций. Доиммунные механизмы есть у всех многоклеточных. Это покровные ткани, фагоциты, микробоцидные гуморальные вещества (протеазы, сильные окислители и свободные радикалы, продуцируемые фагоцитами, эндогенные пептиды-антибиотики и др.), сосудистые реакции. Первичные Рц для микробных патогенов, которые распознают чужое, пытающееся проникнуть или проникшее в покровные ткани и внутреннюю среду, - это Рц PRR и TOLL клеток доиммунного воспаления, локализованных в покровных тканях, - дендритных клеток и B-лимфоцитов, макрофагов, а также растворимые Рц - ряд белков
сыворотки крови (СРБ, СМЛ, компоненты комплемента). Первичные Рц для патогенов на клетках доиммунного воспаления - носители эволюционной памяти многоклеточных, памяти о том, чем отличаются микробы от собственных клеток. Количество известных на сегодня первичных Рц для патогенов порядка 20. Собственные возможности санировать организм от проникшего патогена у клеток доиммунной резистентности слабы. Множество патогенов приспособилось выживать в присутствии факторов доиммунной резистентности к инфекциям. Поэтому лимфоцитарный иммунитет был не отброшен, а закреплён естественным отбором. Сопряжение доиммунных «распознавателей чужого» с лимфоцитарным иммунитетом начинается с того, что дендритные клетки, связавшие и поглотившие патоген, активируются, на них экспрессируются дополнительные мембранные молекулы для контактов с лимфоцитами, инициируется биосинтез цитокинов доиммунного воспаления, клетки метаболизируют патоген и экспрессируют на своей мембране комплексы MHC с пептидами - фрагментами вещества патогена. Дендритные клетки покидают покровные ткани и мигрируют на территорию периферических лимфоидных органов. Таким образом, дендритные клетки превращаются в АПК для T-лим- фоцитов. Что же распознают лимфоциты ?
Рецепторы лимфоцитов для Аг транскрибируются и транслируются с генов, формирующихся как продукты соматической рекомбинации ДНК - уникальной в каждом отдельном дифференцирующемся лимфоците. Расчётное разнообразие вариантов Рц T-лимфоцитов для Аг - 1018. Структура Рц лимфоцитов не находится под влиянием естественного отбора, но находится под влиянием сильных механизмов селекции во время лимфопоэза в онтогенезе. Мощную положительную селекцию T-лимфоцитов можно считать доказанной. В первую очередь селекция идёт на связывание со структурами MHC (в комплексе с эндогенными пептидами) собственного организма, находящимися под контролем естественного отбора. Как бы ни было велико число вариантов Рц лимфоцитов для Аг, число вариантов патогенов может быть и больше (они свободны в своей изменчивости). Но для каждого Рц лимфоцитов возможно много различных лигандов, если их химический состав таков, что есть возможность установить ионные, водородные, ван-дер-ваальсовы и гидрофобные связи с Рц с авидностью 10-6 М и выше.
Существование положительной селекции В-лимфоцитов представляется весьма вероятным, но механизмы её не известны, и это развивающееся направление исследований.
Все T-лимфоциты, выживающие в периферических тканях, постоянно получают по Рц для Аг сигнал для выживания от собственных нормальных тканей (распознают своё), и это поддерживает их в состоянии минимальной частичной активации, но не выводит в режим иммунного воспаления. Связывание лимфоцита только по Рц для Аг никогда не приводит к продуктивной активации лимфоцита.
Получив комплекс сигналов от клеток доиммунного воспаления - по Рц для Аг и по облигатным корецепторам - T-лимфоциты активируются к иммунному ответу, что выражается в их пролиферации и интенсивной продукции цитокинов иммунного воспаления. Можно сказать, что Рц T-лимфоцитов, начинающих иммунный ответ, как-то комплексно распознают повреждения поверхности своих клеток, большое разнообразие вариантов этих повреждений. Иммунное воспаление гораздо более агрессивное, чем доиммунное. Иммунное воспаление направляется лимфоцитами на собственные повреждённые ткани и приводит к их деструкции. Если при этом разрушается и источник размножающегося патогена, то наступают выздоровление и регенерация тканей в очаге деструкции. Излишняя и/или неспецифическая (поликлональная) активация лимфоцитов патогенна (цитокиновая системная интоксикация, повышенная гибель активированных лимфоцитов, реакции гиперчувствительности). Затяжная во времени активация одних и тех же клонов лимфоцитов - тоже патология (болезни с синдромами хронического иммунного воспаления, в том числе аутоиммунные).
Кроме существенной интенсификации защитного воспалительного процесса, лимфоцитарный иммунитет в отличие от доиммунных механизмов резистентности к инфекциям привнёс ещё одно новое свойство, чрезвычайно полезное для выживания и закреплённое естественным отбором - иммунологическую память. При каждом привлечении к действию доиммунные защитные механизмы только изнашиваются и медленно восстанавливаются не выше исходного уровня при прекращении действия патогенного фактора. При лимфоцитарной же реакции на повреждение тканей патогеном вовлё- ченные клоны лимфоцитов размножаются в немалой степени, часть лимфоцитов не расходуется в текущем ответе, а экспрессирует ингибирующие активность лимфоцита Рц и остаётся персистировать в организме на какой-то срок. Вероятно, на тот срок, в течение которого в организме в какой-то форме (вопрос мало изучен) остаются следы искомого патогена. Иммунные лимфоциты памяти во многих случаях способны быстро и активно ликвидировать на ранних стадиях очаг
инфекционного повреждения тканей при повторной инфекции того же патогена (вторичный ответ, протективный иммунитет).
Вторая «ветвь» лимфоцитарного иммунитета - растворимые антитела, продукты B-лимфоцитов, закрывают собой другой фронт атаки патогенных факторов на организм. В отличие от Рц T-лимфоцитов Ig связываются с любыми комплементарными им нативными структурами как в растворе, так и на твёрдой фазе. Ig не «требуют» предобработки и представления им Аг в особом виде. Ig обеспечивают быструю «физико-химическую сорбцию» растворимых продуктов патогенов в жидких средах организма (в первую очередь в крови) и подведение патогенных субстанций к клеткам, имеющим специальные биохимические механизмы деструкции (нейтрофилам, макрофагам, нормальным киллерам, эозинофилам, тучным клеткам). Ig самостоятельны и уникальны по свойствам на этапе исполнения иммунного ответа. Но на этапах инициации В2-лимфоцитов к иммунному ответу они жёстко сопряжены с T-лимфоцитами и, следовательно, на В2-лимфоциты опосредованно распространяются закономерности развития ответа T-лимфоцитов.
Таким образом обновлённая схема иммунного ответа будет выглядеть так:
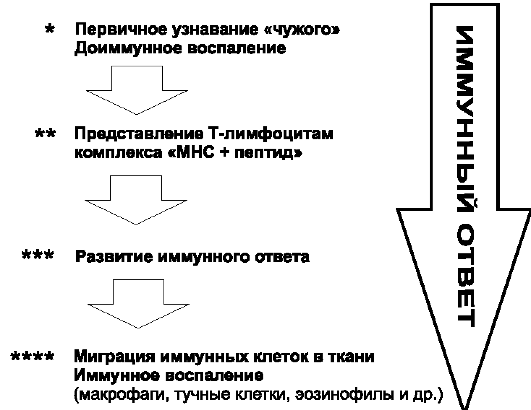 Рис. 16.1. Схема иммунного ответа (сравни с рис. 1.2).
Рис. 16.1. Схема иммунного ответа (сравни с рис. 1.2).
* Первичное узнавание чужого Рц клеток доиммунного воспаления (PRR и TOLL). Развитие доиммунного воспаления.
** Представление комплексов MHC с пептидами T-лимфоцитам. Распознавание лимфоцитами Аг и повреждённого своего.
*** Развитие иммунного ответа (пролиферация лимфоцитов и продукция ими провоcпалительных цитокинов и иных эффекторных молекул). **** Миграция иммунных лимфоцитов в ткани, поражённые патогеном; развитие там иммунного воспаления; деструкция Аг и тканей, повреждённых патогенами.
Немного прикладных следствий. У всякой особи есть эволюционные ограничения защитных возможностей иммунного ответа: 1) унаследованные от родителей инвариантные первичные Рц узнавания чужого на клетках доиммунного воспаления - PRR и TOLL и 2) унаследованные от родителей инвариантные молекулы MHC. И первое, и второе - результат естественного отбора в поколениях, живших в определённой биосфере (в контакте с определёнными микробами). Вакцинация создаёт протективный иммунитет на уровне размножающихся лимфоцитов, но не может «в лоб» исправить дефекты первичного восприятия патогенов клетками доиммунной резистентности и, возможно, никак не может исправить несоответствие MHC какому-либо конкретному патогену (поэтому никогда стандартные вакцины не будут равно подходящи всем и каждому). Некоторые воздействия на иммунный ответ на этапах доиммунного воспаления тем не менее эмпирически подобраны людьми много лет назад. Это, во-первых, адъюванты и, во-вторых, процедуры закаливания. Известные адъюванты - продукты микроорганизмов или их синтетические аналоги и некоторые полимеры, имеющие химическое сродство к мембране дендритных клеток или макрофагов. Адъюванты - непосредственно не лимфотропны и лимфоциты не активируют. Адъюванты активируют непосредственно клетки доиммунного воспаления. Закаливание «тренирует» пластичность и оперативность сосудистых реакций в покровных тканях и на уровне центральных механизмов регуляции со стороны нервной и эндокринной систем. Стенка сосуда - непременный участник всех этапов развития иммунного ответа. Только дендритные клетки с Аг мигрируют в периферические лимфоидные органы по путям лимфодренажа не через стенку сосудов. Лечебные процедуры, в разумных пределах улучшающие лимфодренаж (физиотерапия, массаж и т.п.), также достаточно эффективно могут ускорять и облегчать санацию очагов инфекционного (и постинфекционного) воспаления тканей, о чем и свидетельствует многолетний опыт медицины.
Поскольку всякий иммунный ответ завершается деструкцией собственных тканей, повреждённых патогеном, то чем выше дозы патогенов, тем больше очаги деструкции и тем больнее воспринимается иммунный ответ (иммунное воспаление) и выше вероятность срыва его в патологический режим прогрессирующего иммунного воспаления. Поэтому лечебные воздействия, прямо направленные на сдерживание размножения инфекционных микробов (антибактериальные, противовирусные, противогрибковые, антипротозойные,
противогельминтные химиопрепараты) могут эффективно помочь иммунной системе радикально санировать инфекцию или инвазию (если только токсичность самих препаратов не повредит организм раньше инфекции).
Исследование иммунной системы продолжается. Нам ещё предстоит порадоваться многим открытиям. Опыт и время покажут, правы мы или нет, полагая, что цели современной науки изменились по сравнению с целями, которые преследовала наука на протяжении всего XX в. В XX в. переделывали природу и беспрецедентно интенсифицировали все антропогенные процессы, бесстрашно (или ослеплённо) замахнувшись на невосстановимые ресурсы планеты. Поскольку геном человека как саморазвивающееся целое тоже, скорее всего, невосстановимый ресурс, то настала пора узнавать новое, чтобы понимать, как сохранить то, что осталось от природы. Только наука может открыть и аргументировать то, чего нельзя делать (не на уровне исследований - там можно всё, а на уровне внедрения), чтобы не ликвидировать своими руками нашу форму жизни на Земле.