Ботаника: учебник. Зайчикова С.Г., Барабанов Е.И. 2013. - 288 с. : ил.
|
|
|
|
ГЛАВА VI ЦАРСТВО РАСТЕНИЯ (PLANTAE, EMBRYOPHYTA, PHYTOBIOTA)
Растения - сложные многоклеточные автотрофные организмы, приспособленные к жизни главным образом в наземной среде. В процессе эволюции с выходом растений на сушу изменилось их морфологическое строение. Так, вместо таллома низших растений у высших растений произошла дифференциация тела на органы: побег (стебель и лист) и у большинства - корень. Поэтому растения называют еще листостебельными, или побеговыми (Cormophyta, Cormobionta). Произошли изменения и во внутреннем строении, связанные с возникновением тканей, выполняющих специализированные функции - покровных, проводящих, механических и др. В проводящей системе появляются сосуды и трахеиды, в связи с чем высшие растения получают название - сосудистые. Из зиготы у высших растений формируется многоклеточный зародыш, и они получают еще одно название - зародышевые (Embryobionta).
В цикле развития происходит закономерное чередование 2 поколений: полового - гаметофит и бесполого - спорофит. На гаметофите образуются мужские (антеридии) и женские (архего-
нии) половые органы. Внутри архегония происходит деление зиготы; на ранних этапах развития зародыш развивается под защитой гаметофита и питание полностью зависит от него. Спорофит у преобладающего большинства растений расчленен на побеги (стебли и листья) и корни.
Более примитивные высшие растения (моховидные, плауновидные, хвощевидные, папоротниковидные), несмотря на выход на сушу, еще сохраняют связь с водной средой. Так, у них половой процесс неразрывно связан с водной средой, поскольку без нее невозможно передвижение сперматозоидов; и для существования гаметофитов необходима влажность субстрата и атмосферы.
В процессе эволюции семенные растения как наиболее организованные и приспособленные к наземному образу жизни утратили зависимость размножения от капельно-жидкой среды, и морфологическая эволюция высших растений пошла по пути совершенствования спорофита.
Таким образом, высшие растения подразделяются на 2 неравные по величине и значению группы - высшие споровые и семенные растения.
Высшие споровые растения размножаются спорами, а процессы по- лового и бесполого размножения закономерно сочетаются в жизненном цикле. Бесполое размножение осуществляется с помощью спор, образующихся мейозом в спорангиях, располагающихся на спорофите. Половое размножение происходит путем слияния гамет: сперматозоидов, созревающих в антеридиях, и яйцеклеток - в архегониях, расположенных на гаметофитах.
У семенных растений появляется новый орган размножения - семя с зародышем. Высшие споровые были первыми сосудистыми растениями, заселившими сушу. От них в процессе эволюции произошли семенные растения.
Наиболее известными являются 6 отделов высших растений: Моховидные (Bryophyta), Плауновидные (Lycopodiophyta), Хвощевидные (Equiesetophyta), Папоротниковидные (Polypodiophyta), Голосеменные (Pinophyta) и Покрытосеменные (Magnoliophyta).
ВЫСШИЕ СПОРОВЫЕ РАСТЕНИЯ
Высшие споровые растения появились от водорослевого предка приблизительно 400-430 млн лет назад. Тело примитивных высших растений было дифференцировано на элементарные органы. Надземная часть представляла собой дихотомически ветвящиеся оси, вверху
заканчивающиеся органами спороношения, а снизу - корневищеподобными выростами - ризомоидами (прототип корня) и ризоидами (прототип корневых волосков). Формирование листьев происходило различными путями. У одних высших растений листья сформировались как выросты на осевых органах (листья плауновидных), у других - за счет уплощения и бокового срастания разветвленных осей, несущих на себе спорангии. Поэтому листья выполняли функцию фотосинтеза и бесполого размножения. Со временем произошло разделение функций, и одни листья - спорофиллы (спороносные листья) несли спорангии со спорами, другие - трофофиллы (зеленые листья) выполняли функцию фотосинтеза. Возможно, в процессе эволюции из спороносных листьев развились шишки (стробилы) голосеменных и цветки покрытосеменных.
Усовершенствованию органов сопутствовало и усложнение онтогенеза. Происходит чередование бесполого и полового поколений. Бесполое поколение представлено диплоидным спорофитом, половое - гаплоидным гаметофитом.
Спорофит - растение, образующее споры. Споры появляются в многоклеточных спорангиях в результате мейоза. Они распространяются ветром, водой, животными. У равноспоровых растений все споры одинаковые по размеру. Более высокоорганизованные растения разноспоровые: в микроспорангиях у них образуются многочисленные мелкие споры - микроспоры, а в мегаспорангиях - крупные мегаспоры. При прорастании спор образуются гаметофиты, поэтому гаметофиты гаплоидны.
Гаметофит - растение, образующее гаметы. Мужские гаметы - сперматозоиды образуются в мужских половых многоклеточных органах - антеридиях, похожих на мешочки, а женские яйцеклетки - в колбообразных архегониях (рис. 6.1). Оплодотворение яйцеклеток происходит при наличии капельно-жидкой среды, необходимой для движения сперматозоидов. После оплодотворения образуется дипло- идная зигота, формирующая многоклеточный зародыш. Зародыш со временем развивается в новый спорофит.
В жизненном цикле только у моховидных доминирует гаметофит, у всех остальных высших растений - спорофит. Гаметофит еще называют заростком. Он имеет вид небольшой пластинки (несколько миллиметров) или клубенька, без дифференциации на органы, прикрепляющегося к почве с помощью ризоидов. Вся эволюция высших растений была направлена на редукцию гаметофита и усовершенствование спорофита.
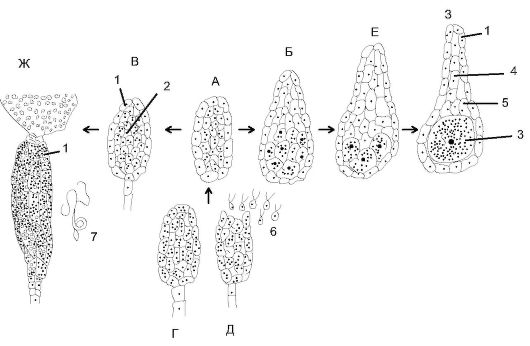
Рис. 6.1. Происхождение и строение антеридиев и архегониев: А, Б - многокамерный гаметангий; В - образование стенки гаметангия; Г, Д - формирование и строение антеридия; Е-З - этапы формирования и строение архегония; 1 - стенка; 2 - сперматогенная ткань; 3 - яйцеклетка; 4 - шейковые канальцевые клетки; 5 - брюшные канальцевые клетки; 6 - изогаметы; 7 - сперматозоиды
ОТДЕЛ МОХОВИДНЫЕ (BRYOPHYTA)
Общая характеристика. В этот отдел входит более 25 тыс. видов сравнительно просто устроенных существующих ныне высших растений. Среди высших растений мхи образуют обособленную группу. Это - единственная в истории растительного мира линия эволюции, связанная с регрессивным развитием спорофита. Мхи представляют собой тупиковую, или слепую, ветвь развития растений, а по общей организации и по экологии они близки и к водорослям.
Характерными признаками мхов являются: 1) отсутствие настоящих корней. У некоторых представителей подземная часть представлена ризоидами; 2) полное преобладание гаметофазы в цикле развития; 3) у более примитивных форм гаметофит представлен слоевищем, у других расчленен на стебель и листья; 4) половое и бесполое поколения существуют вместе, при этом спорофит вырастает на гаметофите после оплодотворения.
По данным многих ученых, моховидные произошли от бурых водорослей. При прорастании спор у них развивается ветвистая зеленая нить - протонема, напоминающая нитчатую водоросль. Половой процесс осуществляется только в водной среде. Наиболее примитивными считаются печеночные мхи, а более высокоорганизованными - листостебельные.
В отделе Моховидные рассмотрим классы: Печеночники (Hepaticopsida) и Листостебельные (Bryopsida).
Класс Печеночники (Hepaticopsida)
Для представителей этого класса, включающего около 8500 видов, характерны чрезвычайно большое разнообразие структуры гамето- фита (слоевище или стебель с листьями простого строения) и однотипность спорофита. Наиболее распространенным представителем класса Печеночников является маршанция обыкновенная (Marchantia polymorpha L.), произрастающая на влажной лесной почве. Ее слоевище имеет вид дихотомически ветвящейся стелющейся зеленой пластинки (размером до 10 см). Слоевища - раздельнополые, прикрепляющиеся к почве с помощью ризоидов (рис. 6.2). Органы полового размножения размещаются на особых подставках и возвышаются над слоевищем. У мужских гаметофитов имеются подставки в виде 8 лопастных дисков, сидящих на ножке. На верхней стороне дисков открываются антеридии - сперматозоиды двужгутиковые. На женских гаметофитах - подставки в виде многолучевой звезды: между лучами подставки группами (шейкой вниз) расположены архегонии. В дождливую погоду или с росой сперматозоиды попадают на женские подставки и проникают в архегоний. После оплодотворения из зиготы развивается спорангий в виде овальной коробочки, сидящей на очень короткой ножке. Внутри коробочки в результате мейоза образуются гаплоидные, но физиологически разные споры. К моменту созревания спор коробочки лопаются, и споры высыпаются. Споры, выпадающие из спорангия, сначала дают пластинчатую слабо развитую протонему, из которой затем развивается новый (мужской или женский) гаметофит. У печеночников есть и вегетативное размножение, осуществляющееся выводковыми почками, образующимися в выводковых корзиночках на слоевищах.
Класс Листостебельные мхи Bryopsida (Musci)
Листостебельные мхи - самый крупный класс моховидных. Они широко распространены (от Арктики до «оазисов» Антарктики), с наибольшим числом представителей (около 700 родов, объединяю-

Рис. 6.2. Маршанция (Marchantia polymorpha): А - таллом с мужскими подставками; Б - разрез через мужскую подставку; В - таллом с женскими подставками; Г - продольный разрез через женскую подставку; Д - молодой спорогоний; Е - взрослый спорогоний со вскрывшейся коробочкой; Ж - споры и элатеры; 1 - антеридий; 2 - архегоний; 3 - гаустория; 4 - ножка спорофита; 5 - коробочка; 6 - споры; 7 - элатера
щих 15 тыс. видов). Класс Листостебельные мхи подразделяется на 2 подкласса: Торфяные, или Белые, мхи (Sphagnidae) и Зеленые мхи (Bryidae).
Подкласс Белые, или Торфяные, мхи (Sphagnidae)
Торфяные мхи имеют единственный род Сфагнум (Sphagnum), к которому относится свыше 300 видов. Все представители беловатозеленого цвета, не имеют ризоидов. От стебля растения отходят боковые веточки, на верхушке собранные в головку (рис. 6.3, А). Веточки сфагнума усажены мелкими листьями. Листья однослойные, состоят из 2 типов клеток: хлорофиллоносных и мертвых водоносных (гиалиновых) клеток. За счет того, что в своем строении листья имеют мертвые гиалиновые клетки (способные удерживать воду), они обла- дают гигроскопическим свойством. Сфагнум в 4 раза гигроскопичнее ваты. Фотосинтез протекает в живых ассимиляционных клетках, содержащих хлоропласты (рис. 6.3, Б). При высыхании мертвые клетки
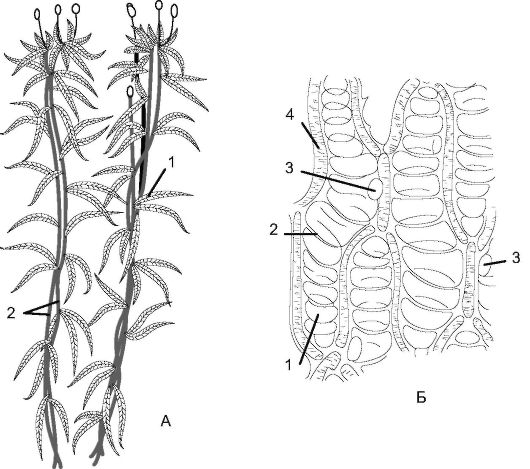
Рис. 6.3. Сфагнум (Sphagnum): А - внешний вид растения: 1 - боковая веточка с листьями; 2 - стебель; Б - анатомическое строение листа (вид сверху): 1 - гиалиновая клетка; 2 - кольчатые утолщения; 3 - пора; 4 - хлорофиллоносная клетка
заполняются воздухом, и мох становится беловатым, отсюда и название - Белый мох. Сфагновые мхи могут быть одно- и двудомными, но в любом случае архегонии и антеридии располагаются на разных боковых веточках. Процесс оплодотворения происходит в присутствии воды с образованием зиготы, из которой развивается бесполое поколение - спорофит в виде шарообразной коробочки и короткой ножки. Внутри коробочки формируется спорангий. В нем в результате мейоза образуются гаплоидные споры. При созревании спор верхушка стебля сильно вытягивается, крышечка отпадает от коро- бочки, а споры высыпаются и разносятся ветром. Споры прорастают в пластинчатую протонему, на которой образуются побеги нового гаметофита.
Мхи растут верхушкой стебля, а его нижняя часть отмирает - «отторфовывается». Так в течение многих лет образуются огромные залежи торфа. Процесс торфообразования происходит благодаря за- стойному переувлажнению, отсутствию кислорода и созданию мхами кислой среды. Эти условия в совокупности оказываются неблагоприятными для процессов гниения, т.е. для развития грибов и бактерий, что препятствует разложению сфагнумов. Сфагнум может использоваться как антисептическое средство благодаря присутствию фенолоподобного вещества - сфагнола и в качестве перевязочного материала.
Подкласс Зеленые (Бриевые) мхи (Bryidae)
Это наиболее обширный (свыше 14 тыс. видов) подкласс из всех листостебельных мхов, распространенный повсюду. Его представи- тели - это, как правило, многолетние растения высотой от 1 мм до 60 см. Преобладающая окраска зеленая, но может быть буроватокрасной и даже черной. Характерный представитель этого подкласса - кукушкин лен обыкновенный (Polytrichum commune) - один из наиболее высокорослых мхов, его стебель достигает в высоту 50 см. Произрастает на сырой почве в лесах, в болотах, образуя крупные подушкообразные дернины. Стебель мха - прямостоячий, неветвя- щийся, густо покрыт жесткими линейно-шиловидными листьями. Листья многослойные, в отличие от других мхов состоят из однородных рядов хлорофиллоносной ткани. Подземная часть представлена многолетними ветвящимися ризоидами.
Кукушкин лен относится к двудомным растениям. На женском растении (гаметофите) между верхними, салатного цвета листьями образуются архегонии - женские половые органы. Архегоний яв-
ляется многоклеточным образованием колбовидной формы. Суженная часть - шейка, расширенная - брюшко, в котором помещается крупная яйцеклетка. На мужском растении (гаметофите) среди верхних красных листьев развиваются антеридии - мужские половые органы, в которых образуются двужгутиковые сперматозоиды. Антеридии имеют вид продолговатых или округлых мешочков на ножке. При созревании архегония шейковые, или канальцевые, клетки ослизняются и на их месте формируется узкий канал, по которому сперматозоид может проникнуть в яйцеклетку. В период обильных дождей или снеготаяния сперматозоиды подплывают к архегониям.
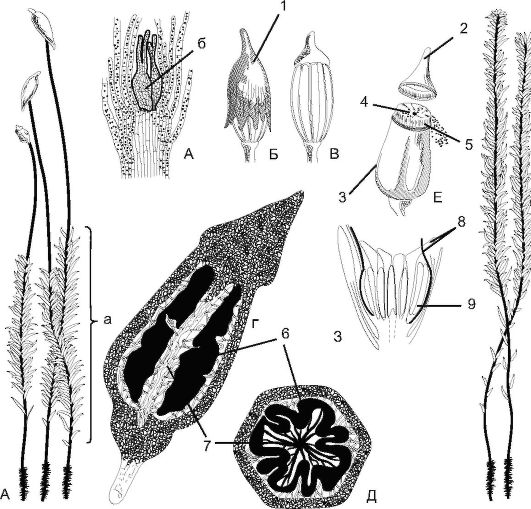
Рис. 6.4. Кукушкин лен (Роlytrichum): А - женский гаметофит (а) с архегониями (б); Б - коробочка с колпачком; В - внешний вид коробочки; Г - продольный разрез коробочки; Д - поперечный разрез коробочки; Е - вскрывшаяся коробочка; Ж - мужской гаметофит; З - вершина мужского гаметофита с антеридиями и парафизами; 1 - колпачок; 2 - крышечка; 3 - урночка; 4 - эпифрагма; 5 - перистом; 6 - спорангий; 7 - колонка; 8 - парафизы; 9 - антеридий
Как предполагают, они обладают хемотаксисом к содержимому слизи архегония. Сперматозоид проникает в архегоний и продолжает движение к яйцеклетке. Слияние гамет и дальнейшее развитие зиготы происходят внутри архегония. Через несколько месяцев из зиготы прорастает спорофит (спорогон), который представляет собой коробочку на длинной ножке (рис. 6.4).
Нижняя часть ножки превращена в гаусторий (присоска), внедряющийся в тело женского гаметофита. Таким образом, спорофит лишен самостоятельности и полностью зависит от гаметофита. Сверху коробочка прикрыта легко отпадающим колпачком (остаток архегония) с тонкими, направленными вниз волосками, напоминающими льняную пряжу (отсюда и название растения). Внутри коробочки - в спорангии - происходит образование спор мейозом. При созревании спор колпачок, а затем и крышечка отделяются и споры высыпаются из отверстий наверху коробочки (урны). Коробочка имеет ряд зубцов - перистом, прикрывающих отверстия во влажную погоду. Одинаковые по форме споры (изоспоры) разносятся ветром, а затем попадают в почву и прорастают в протонему (нитевидное образование), на которой из почек формируются листостебельные побеги. Таким образом, эти побеги с протонемой представляют собой гаметофит, имеющий гаплоидный набор хромосом. На этом цикл замыкается.
ОТДЕЛ ПЛАУНОВИДНЫЕ (LYCOPODIOPHYTA)
Плауновидные - один из самых древних отделов высших растений; их ископаемые остатки известны из силурийского периода па- леозойской эры. Это были огромные, достигавшие в высоту 40 м и в диаметре до 2 м растения, которые занимали господствующее положение в растительности на всем земном шаре. В настоящее время плауновидные являются обитателями хвойных лесов и представляют собой многолетние вечнозеленые травы, реже - полукустарники; всего их насчитывается до 1 тыс. видов (4 рода). Это самые первые сосудистые растения, имеющие хорошо развитые, дихотомически ветвящиеся, олиственные побеги. Их листья возникли как поверхностные боковые выросты оси. Все листья мелкие - микрофилия - с центральной жилкой. Листорасположение супротивное, спиральное и мутовчатое. Плауновидные нарастают за счет верхушечной меристемы, деятельность которой со временем угасает, за счет чего они ограничены в росте. Подземная часть представлена придаточными корнями.
Отдел делится на 2 класса: равноспоровые Плауновые (Lycopodiopsida) и разноспоровые Полушниковые (Isoetopsida).
Класс Плауновые (Lycopodiopsida)
До настоящего времени сохранился один порядок (Lycopodiales), одно семейство (Lycopodiaceae), представленное 2 родами. Наиболее значим род Плаун (Lycopodium), который насчитывает около 200 видов, распространенных от арктических областей до тропиков. В тро- пиках вертикальные стебли плауновидных достигают в высоту 1,5 м. Типичным представителем зеленомошных хвойных лесов умеренного пояса Северного полушария является плаун булавовидный (L.clavatum). Ползучий, дихотомически ветвящийся побег этого вечнозеленого, многолетнего травянистого растения достигает в длину 3 м. Стебель густо покрыт мелкими линейно-ланцетными листьями. От стебля отходят тонкие придаточные корни, а вверх - дихотомически ветвящиеся вертикальные невысокие побеги. На верхушках побегов к середине лета появляются спороносные колоски, как правило, по 2 на одной ножке. Колосок состоит из споролистиков (спорофиллов), прикрепляющихся к оси и имеющих на своем основании почкообразные спорангии на короткой ножке. В спорангиях в результате мейоза образуются гаплоидные споры. Морфологически и физиологически все споры одинаковы (изоспоры) - округло-тетраэдрической формы, покрыты толстой желтой оболочкой (рис. 6.5).
Споры высыпаются из спорангия и при благоприятных условиях прорастают (примерно в течение 5 лет) в маленький 2-3- миллиметровый заросток - обоеполый, клубеньковой формы гаметофит, лишенный хлорофилла (половое поколение). В клетки заростка внедряются гифы грибов. В присутствии гиф почвенного гриба гаметофит, питаясь сапрофитно, медленно растет и развивается в течение 12 лет. На верхней стороне гаметофита образуются много- численные антеридии и архегонии, погруженные в ткань заростка, и только шейки архегониев выдаются наружу. Оплодотворение яйцеклетки, находящейся в архегонии, двужгутиковым сперматозоидом происходит в капельно-жидкой среде. После оплодотворения из зиготы образуется зародыш спорофита, развивающийся в брюшке архегония, а из него - взрослое растение. Взрослое растение плауна является спорофитом и представляет собой бесполое поколение. Некоторые виды плауна содержат парализующий яд, сходный по своему действию с ядом кураре. Очевидно поэтому позвоночные животные не употребляют плауны в пищу. Споры плауна булавовидного содер-
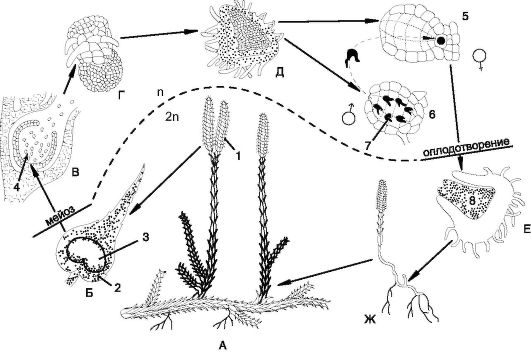
Рис. 6.5. Чередование поколений в жизненном цикле плаунов (Lycopodium clavatum): А - взрослый спорофит со спороносным колоском (1); Б - споролистик (2) со спорангием (3) спороносного колоска; В - образование спор (4) в спорангии; Г - прорастание спор в заросток; Д - обоеполый заросток (гаметофит) с архегониями (5), антеридиями (6) со сперматозоидами (7); Е - зародыш спорофита (8) на заростке; Ж - молодой спорофит
жат до 50% невысыхающих масел; они использовались в медицине в качестве детской присыпки, а также для обсыпки пилюль.
Класс Полушниковые, или Шильниковые (Isoetopsida)
К разноспоровым плаунам относится род Селагинелла (Selaginella), насчитывающий около 700 видов, преимущественно тропических. Это нежные многолетние травянистые растения; большинство из них - мелких размеров - до 15 см в высоту, но имеются виды с ла- зающими и вьющимися побегами, достигающими 20 м в длину. Прикрепляются они к почве с помощью тонких дихотомически разветвленных корней, образующихся на особых выростах стебля - ризофорах (корненосцах).
Селагинеллы - разноспоровые растения. В спороносных колосках (стробилах) в мегаспорангиях образуются по 4 мегаспоры и многочисленные микроспоры в микроспорангиях. При прорастании микроспоры появляется сильно редуцированный мужской заросток
(гаметофит), состоящий из маленькой проталлеальной (остаток вегетативного тела заростка) и большой антеридиальной клеток. Антеридиальная клетка дает начало антеридию, где формируются двужгутиковые сперматозоиды. Мегаспора развивается в женский гаметофит, состоящий из многоклеточной ткани с архегониями и ризоидами. После оплодотворения яйцеклетки развивается зародыш, состоящий из стебелька, листочков и ризофора. У некоторых видов оплодотворение совершается в колоске, и на почву падает зародыш.
В отличие от плаунов сильная редукция гаметофита, связанная с разноспоровостью, представляет собой основное направление эволюции высших растений.
ОТДЕЛ ХВОЩЕВИДНЫЕ (EQUISETOPHYTA)
В геологическом прошлом хвощевидные были весьма разнообразны. Ископаемые хвощевидные (например, древовидные каламиты) достигали в высоту 20 м, в их стволах находили вторичную ксилему. Наряду с древними плауновидными и древовидными папоротниками они образовывали леса каменноугольного периода. Современные хвощи - травянистые растения, представленные в растительном мире единственным классом Хвощевые (Equiesetopsida), одним порядком (Equesetales), одним семейством (Equesetaceae) и одним родом Хвощ (Equiesetum).
Класс Хвощевые (Equisetopsida)
Род Хвощ (Equiesetum) представлен многолетними травянистыми растениями, встречающимися в условиях избыточного увлажнения лесов, полей, лугов, болот. Ранней весной у хвоща полевого из глу- боко залегающих корневищ вырастают однолетние спороносные побеги, заканчивающиеся спороносными колосками. Клетки эпидермы побегов пропитаны кремнеземом. На узлах летних побегов сидят бурые чешуйчатые листья, срастающиеся основаниями в листовое влагалище, и мутовки боковых побегов. Боковые побеги выполняют ассимилирующую функцию. Спороносные колоски состоят из оси, перпендикулярно которой крепятся щитки - спорангиофоры (видоизмененные боковые побеги); под ними находится 6-10 спорангиев, содержащих споры, образующиеся в результате мейоза. Вначале щитки сидят плотно, без зазоров, но позднее, к моменту созревания спор, стержень колоска удлиняется. Между щитками образуются промежутки, через которые и высыпаются споры из созревших спорангиев.
Шаровидная зеленая спора обмотана 4 пружинами - элатерами. При подсыхании элатеры раскручиваются, с их помощью споры сцепля- ются в крупные рыхлые комочки и лучше разносятся потоками воздуха. Попадая во влажную среду, споры прорастают целыми группами заростков, что увеличивает вероятность оплодотворения. Из физиологически различных спор развиваются разнополые гаметофиты. Заростки очень мелкие (всего несколько миллиметров), имеют вид небольших зеленых рассеченных пластинок с ризоидами. Через 3-5 недель на одних заростках созревают антеридии с многожгутиковыми сперматозоидами, на других - архегоний с яйцеклеткой. Во влажной среде происходит оплодотворение. Из образовавшейся зиготы развивается зародыш, а из него - взрослый спорофит (рис. 6.6).
У разных видов хвощей строение побегов различное. Так, у хвоща полевого после рассеивания спор весенние неветвящиеся бесхлорофильные спороносные побеги отмирают, и на смену им вырастают летние зеленые ассимилирующие побеги (рис. 6.7). У других видов
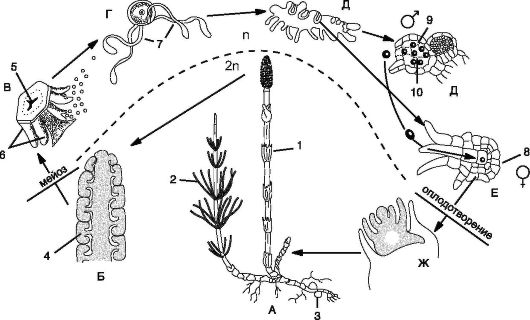
Рис. 6.6. Чередование поколений в жизненном цикле хвощей (Equisetum arvense): А - взрослое растение хвоща (спорофит): генеративный побег со спороносным колоском (1); 2 - вегетативный побег; 3 - клубеньки; Б - спороносный колосок со спорангиоформами (4); В - спорангиофор: 5 - щиток спорангиофора; 6 - спорангии; Г - спора с элатерами (7); Д - мужской заросток с антеридиями (9); 10 - сперматозоид; Е - женский заросток с архегониями (8); Ж - зародыш будущего спорофита
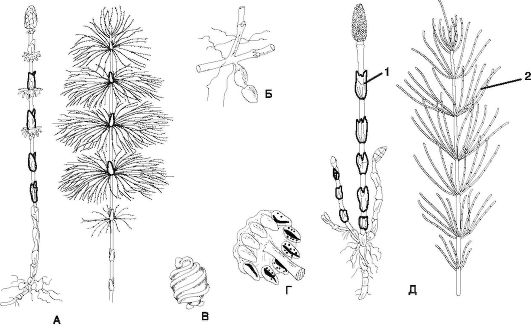
Рис. 6.7. Семейство Хвощевые: A - хвощ лесной (Equisetum sylvaticum), спороносный (слева) и вегетативный (справа) побеги; Б - клубеньки на корневище; В - споры с прижатыми элатерами; Г - спорангиофор со спорангиями; Д - хвощ полевой (Equisetum arvense), спороносный (1) и вегетативный (2) побеги
хвощей (лесной, луговой) спороносные колоски образуются на зеленых ассимилирующих побегах.
Хвощи, быстро размножаясь корневищами, становятся сорняками пастбищ, так как являются несъедобными для животных растениями, поскольку содержащиеся в них сапонины и алкалоиды могут вызывать отравления. Хвощ полевой применяют как кровоостанавливающее и мочегонное средство.
ОТДЕЛ ПАПОРОТНИКОВИДНЫЕ (POLYPODIOPHYTA)
Папоротники по возрасту уступают только плауновидным и имеют приблизительно один геологический возраст с хвощевыми. Современные папоротники насчитывают около 300 родов (12 тыс. видов). Обитают они в самых разных местах, но преимущественно - в условиях повышенной влажности: в Азии, Австралии, Южной Америке. Наибольшее их разнообразие характерно для влажных тропических лесов. У древовидных тропических форм стебли могут достигать в высоту 25 м.
Для всех папоротников характерно преобладание многолетнего листостебельного спорофита над временно образующимся примитивным гаметофитом.
Класс Полиподиопсиды (Polypodiopsida)
У папоротников нашей флоры - таких, как орляк (Pteridium aguilinum), кочедыжник женский (Athyrium filix-femina), щитовник мужской (Dryopteris filix-mas) и других, надземный стебель отсутствует, и внешне растение представляет собой пучок листьев - вайи, отходящих от хорошо развитого корневища (рис. 6.8). Вайями листья папоротника называют из-за происхождения, поскольку они возникли в результате уплощения крупных ветвей предковых растений. Это доказывается тем, что вайи папоротника долго сохраняют верхушечный рост, образуя при этом характерную разворачивающуюся улитку, что не свойственно листьям. Рассмотрим цикл развития папоротника на примере щитовника мужского.
Вайи папоротника дважды (щитовник мужской) или трижды перисто-рассеченные (кочерыжник женский). Взрослое растение является спорофитом (бесполое поколение 2n). На нижней стороне вайи образуются сорусы - собрания спорангиев на ножке на выросте листа - плаценте, снизу прикрытые покрывальцем - индузием. Стенка спорангия однослойная, состоит из кольца с внутренними и радиальными утолщениями; она охватывает спорангий на 2/3 и на 1/3 (в устье) остается неутолщенной. В спорангиях мейозом образуются гаплоидные споры. При созревании спор внешние стенки кольца клеток ссыхаются, стенка спорангия разрывается поперек в области устья, и споры высыпаются. Из спор прорастают гаплоидные заростки, или обоеполые гаметофиты (половое поколение). Заросток представляет собой зеленую сердцевидную пластинку (около 1 см), которая с помощью ризоидов прикрепляется к почве. Внизу на заростке среди ризоидов формируются антеридии, а позже на верхней части пластинки заростка - архегонии, причем брюшко погружено в тело заростка, а шейки выступают на его поверхности. Во время дождя или при выпадении росы антеридии вскрываются, и штопорообразно извитые сперматозоиды с пучком жгутиков проникают в архегонии и оплодотворяют яйцеклетку. Из зиготы развивается зародыш, переходящий со временем (стебелек с листочком и корешок) к самостоятельной жизни спорофита (рис. 6.9).
Значение папоротников велико. Они выступают в роли важнейшего компонента многих лесных сообществ. Корневище щитовника
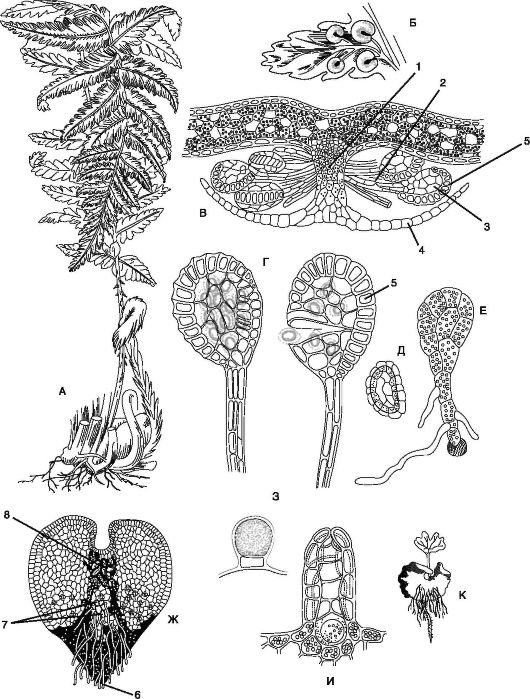
Рис. 6.8. Щитовник мужской (Dryopleris filix-mas): А - спорофит; Б - часть вайи с сорусами; В - поперечный срез через сорус; Г - спорангий; Д - спора; Е - молодой гаметофит; Ж - зрелый гаметофит-заросток; З - антеридий; И - архегоний; К - молодой спорофит: 1 - плацента; 2 - ножка спорангия; 3 - спорангий; 4 - индузий (покрывальце соруса); 5 - кольцо утолщения; 6 - ризоиды; 7 - антеридий; 8 - архегоний
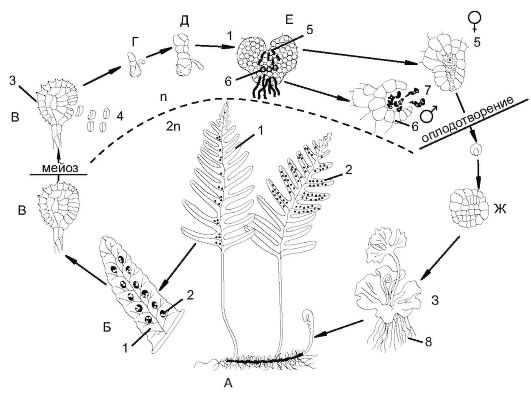
Рис. 6.9. Чередование поколений и смена ядерных фаз у папоротника (Роlуроdium sр.):
А - взрослое растение папоротника (спорофит): 1 - вайя; 2 - сорусы; Б - вайя папоротника с сорусами; В - спорангии: 3 - кольцо утолщения; 4 - споры; Г - прорастание споры; Д - формирование заростка; Е - обоеполый заросток (гаметофит): 5 - архегонии; 6 - антеридии; 7 - сперматозоиды; Ж - образование зиготы на заростке; З - заросток с развивающимся зародышем: 8 - ризоиды
мужского оказывает антигельминтное действие, молодые побеги орляка в некоторых странах употребляют в пищу.
Разноспоровые папоротники объединяют в 2 порядка водных папоротников: Марсилеевые (Marsiliales) и Сальвиниевые (Salviniales). Биологическое значение разноспоровости заключается в обеспечении развивающегося гаметофита питательными веществами, накопленными в мегаспоре.