Ботаника: учебник. Зайчикова С.Г., Барабанов Е.И. 2013. - 288 с. : ил.
|
|
|
|
ГЛАВА V ЦАРСТВО ДРОБЯНКИ (MYCHOTA). ЦАРСТВО ПРОТОКТИСТЫ (PROTOCTISTA). ЦАРСТВО ГРИБЫ (MYCOTA, FUNGI)
ТАКСОНОМИЧЕСКИЕ КАТЕГОРИИ И ТАКСОНЫ. БИНАРНАЯ НОМЕНКЛАТУРА
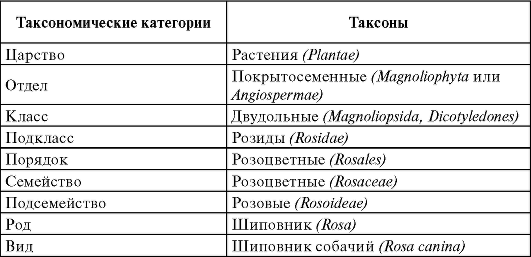
Систематика изучает многообразие расти- тельных организмов, и ее основная задача - классификация этого огромного многообразия. Вид представляет собой основную таксономическую категорию в систематике. Каждый вид принадлежит к какому-либо роду, род - к семейству, семейство - к порядку, порядок - к классу, класс - к отделу, отдел - к царству. При необходимости могут использоваться и промежуточные таксономические категории, например подвид, подрод, подсемейство, надпорядок, надцарство.
Таксонами принято называть реально существующие или существовавшие группы организмов, которые в процессе классификации отнесены к определенным таксономическим категориям. Например, рода или виды являются таксономическими категориями, а род Шипов-
ник и вид Шиповник собачий - конкретные таксоны. Первый таксон охватывает все существующие виды рода Шиповник, второй - все особи, относимые к виду Шиповник собачий (табл. 5.1).
Таблица 5.1. Таксономические категории и таксоны (на примере Ши- повника собачьего)
Научные названия всех таксонов - выше вида - состоят из одного латинского слова, т.е. униноминальны. Для видов начиная с
Подвиды - более мелкие таксоны внутри вида, обладающие своим ареалом (щавель обыкновенный).
Разновидности - менее различаются между собой, чем подвиды, не имеют своего ареала. Но при этом морфологические признаки закреплены наследственно.
Сорт - группа особей внутри вида, подвида, разновидностей, имеющих ряд наследственных, стойких признаков (зимостойкость, крупноплодность, высокая урожайность и др.), не передающихся по
наследству. Чтобы сохранить материнские признаки сорта, обычно проводят вегетативное размножение.
Надцарство Доядерные организмы - Прокариоты (Procariota). Царство Дробянки (Mychota)
Подцарство Настоящие бактерии (Bacteriobonta)
Микроорганизмы, клетки которых не имеют ядра, ограниченного мембраной, относятся к прокариотам (от лат. pro - перед и греч. caryon - ядро). Это очень древние организмы, появившиеся более 3 млрд лет назад. Бактерии - это в основном одноклеточные и колониальные организмы. Условно их относят к царству растений, однако связь бактерий с высшими растениями или водорослями не доказана. Бактерии распространены повсеместно, так, в 1 мл воды их содержится около тысячи, а в 1 мл молока - около миллиона.
Строение бактериальной клетки
В составе клеточной стенки бактерий отсутствуют хитин и целлюлоза, что характерно для грибных и растительных клеток. Опорный каркас стенок образован гликопептидом (мукопротедом) муреином. Поверх клеточной оболочки располагается капсула или слизистый слой. Эта часть клеточной стенки, являясь дополнительной защитой бактерий, также служит для формирования колоний из отдельных клеток. Клеточная стенка сверху покрывает плазматическую мембрану бактерий.
Цитоплазма бактерий имеет зернистый вид. В химическом отношении цитоплазма представляет собой сложную смесь белков, жи- ров, углеводов, многочисленных других органических соединений, минеральных веществ, воды и включений - запасных питательных веществ (волютин, содержащий фосфор, гликоген, жир). У фотосинтезируемых бактерий во впячиваниях наружной плазматической мембраны находится пигмент - бактериохлорофилл. Значительная часть органических веществ присутствует в коллоидном состоянии, причем сами вещества всегда тесно перемешаны. В цитоплазме находятся и рибосомы 70 S. Кроме рибосом, в цитоплазме бактерий можно обнаружить сложные мембранные структуры в виде самостоятельных телец - мезосом, образующихся из наружной мембраны. Мезосомы выполняют функцию аппарата Гольджи.
Ядерный аппарат бактерий обычно называют нуклеоидом, представленным одной кольцевой молекулой ДНК. Нить ДНК у бактерий
прикрепляется какой-либо частью к цитоплазматической мембране или ее выростам с помощью специфических белков. Таким образом, цитоплазматическая мембрана у бактерий принимает участие в деле- нии нуклеоида. Кроме кольцевой хромосомы могут находиться более короткие двухцепочечные нити ДНК - так называемые плазмиды (внехромосомные факторы наследственности). Многие бактерии подвижны. Органами движения у них служат жгутики и тонкие палочковидные белковые выросты - пили, или фимбрии (короче и тоньше жгутиков, помогают прилипать к другим клеткам). Жгутики берут свое начало под цитоплазматической мембраной, закрепляясь там с помощью пары дисков (рис. 5.1).
Колонии большинства бактерий, растущих на неокрашенных средах, сероватые или беловатые; иногда они полупрозрачны, ино- гда совсем непрозрачны. Однако для некоторых видов характерно образование самых разнообразных пигментов (бактериохлорофилл, цитохромов, липохромов и др.).
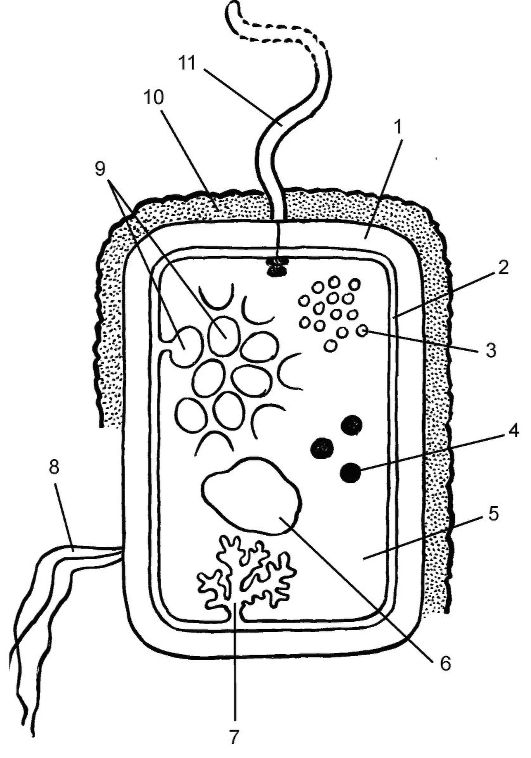
Рис. 5.1. Строение бактериальной клетки:
1 - клеточная стенка;
2 - плазматическая мембрана; 3 - рибосомы; 4 - запасные питательные вещества; 5 - цитоплазма; 6 - кольцевая молекула ДНК; 7 - мезосома;
8 - пили или фимбрии;
9 - фотосинтетические мембраны; 10 - капсула; 11 - жгутик
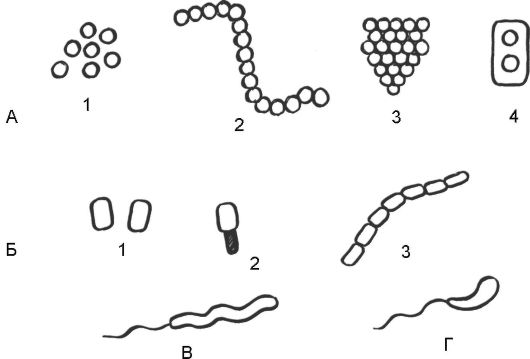
Рис. 5.2. Морфологические типы бактерий:
А - кокки: 1 - кокки; 2 - стрептококки (многие виды вызывают инфекционные заболевания верхних дыхательных путей); 3 - стафилококки (разные штаммы вызывают фурункулез, воспаление легких, пищевые отравления); 4 - диплококки (две клетки в одной капсуле, например пневмококк - возбудитель пневмонии); Б - бациллы (палочковидные): 1 - одиночные палочки (возбудитель брюшного тифа); 2 - бациллы с эндоспорами (например, возбудитель столбняка); 3 - палочки, образующие цепочки клеток (возбудитель сибирской язвы); В - спириллы (спиралевидные) - спиральная палочка с одним жгутиком. По форме клеток на спириллы похожи спирохеты, но есть различия по способу передвижения, например возбудитель сифилиса; Г - вибрионы - короткие палочки, всегда изогнутые в виде запятой (возбудитель холеры)
Существуют различные морфологические типы бактерий (рис.
5.2).
Питание и энергетический обмен
Бактерии, как и грибы, способны поглощать питательные вещества всей поверхностью через плазматическую мембрану, которая проницаема для некрупных молекул белков, полисахаридов и др. Для этого в окружающую среду выделяются экзоферменты, расщепляю-
щие полимеры до более простых молекул. Такое явление называется внеклеточным перевариванием.
По типу питания бактерии подразделяются на автотрофы (хемосинтетики, фотосинтетики) и гетеротрофы (сапрофиты, симбионты, паразиты). Источником энергии для хемосинтетиков является окис- ление минеральных веществ, для фотосинтетиков - свет.
К хемосинтетикам относятся: серобактерии, получающие энергию при окислении серы или сероводорода; железобактерии - при окислении двухвалентного железа; нитрифицирующие бактерии - при окислении аммиака или нитритов. К фотосинтетикам относятся зеленые и пурпурные серобактерии, живущие в соленых, пресных и серных водоемах. Фотосинтез у них проходит по анаэробному типу, т.е. без выделения О2, но при этом задействована фотосистема I (ФС I) в отличие от сине-зеленых водорослей и растений.
К гетеротрофным организмам относятся получающие углерод за счет разложения готового органического вещества мертвых организмов сапрофиты, а также использующие вещество живых растений, животных и человека - паразиты. Бактерии, живущие в симбиозе с растительными или животными организмами, являются симбионтами (например, клубеньковые азотфиксирующие бактерии бобовых). Энергетический обмен (катаболизм) у гетеротрофных бактерий связан с окислением органических веществ в форме аэробного дыхания и брожения. По типу дыхания бактерии бывают анаэробами (столбнячная палочка) и аэробами (туберкулезная палочка).
Размножение бактерий
Бесполое размножение бактерий. После достижения определенных (критических) размеров клетка подвергается делению. Для подавляющего большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию 2 одинаковых дочерних клеток. Весь цикл деления прокариот можно разделить на 3 стадии:
I - редупликация (начинается в точке прикрепления кольцевой хромосомы к цитоплазматической мембране, которая определяет начало и конец ее репликации) (см. рис. 5.3, А);
II - синтез мембраны в области контакта ДНК с цитоплазматической мембраной. Это приводит к разделению (растаскиванию) дочерних молекул ДНК и оформлению обособленных хромосом (см. рис. 5.3, Б);
III - образование поперечной перегородки. Синтез поперечной перегородки идет от периферии к центру. Она разделяет 2 дочерние
хромосомы, каждая из которых прикреплена к цитоплазматической мембране (рис. 5.3).
Такое простое деление клетки в благоприятных условиях наступает каждые 15-20 мин, что обеспечивает большую интенсивность размножения. Таким образом, при равновеликом бинарном делении материнская клетка, делясь, дает начало 2 дочерним клеткам, а сама исчезает.
Особым способом защиты от неблагоприятных условий является процесс спорообразования. Каждая клетка образует за счет потери воды одну спору внутри - эндоспору. При этом протопласт сжима- ется, покрывается плотной оболочкой, а прежняя оболочка клетки разрушается, и спора высвобождается. Так споры могут длительное время пережидать неблагоприятные условия (сотни и даже тысячи лет) и сохранять жизнеспособность.
Половое размножение бактерий. У некоторых бактерий известен половой процесс, при котором происходит лишь генетический обмен между клетками, но не происходит образование новых клеток. Он состоит в прямом контакте 2 клеток, при этом клеткой-донором,
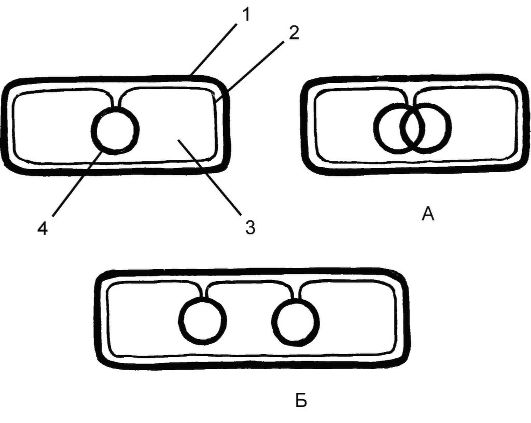
Рис. 5.3. Равновеликое бинарное поперечное деление бактерий:
1 - клеточная стенка; 2 - плазматическая мембрана; 3 - цитоплазма; 4 - молекула ДНК; А - редупликация ДНК; Б - синтез мембраны в области контакта ДНК с цитоплазматической мембраной
выполняющей мужские функции, формируется специальный вырост - копуляционный канал, по которому генетический материал (ДНК) передается в клетку-реципиент, имеющую женскую потенцию. Такой процесс называется конъюгацией. Очень часто наблюдается передача не всей молекулы ДНК, а только ее отдельных фрагментов.
У бактерий имеются и другие способы передачи наследственного материала: трансформация и трансдукция. Трансформация осуществляется путем внесения ДНК разрушенных клеток одной культуры в живую культуру другой бактерии. Трансдукция проявляется в переносе генетического материала от одной культуры к другой с помощью бактериофагов. Эти способы передачи наследственного материала расцениваются как хромосомные мутации.
Значение бактерий в природе
1. Бактерии участвуют в разрушении мертвого органического материала в экосистемах и тем самым принимают непосредственное участие в круговороте углерода, азота, фосфора, серы, железа и других элементов. Роль бактерий в процессе разложения определяющая, так как ими разлагаются практически все природные соединения.
2. Многие виды бактерий человек использует в народном хозяйстве для получения органических продуктов в результате брожения (уксуснокислые, молочнокислые). Большая группа молочнокислых бактерий производит молочнокислое брожение за счет анаэробного окисления сахара молока и других углеводов в молочную кислоту. Так, молочнокислый стрептококк (Streptococcus lactis) широко используется для приготовления простокваши; сырная палочка (Lactobacterium bulgaricum) - для приготовления кефира, сметаны, сыров.
3. Бактерии служат источником для получения антибиотиков (стреп томицин, грамицидин).
4. Бактерии используются для создания новых способов получения важнейших для промышленности веществ, в том числе спиртов, органических кислот, сахаров, полимеров, аминокислот и ряда фер- ментов.
5. Симбиотические бактерии кишечника млекопитающих (микрофлора) участвуют в синтезе ряда витаминов группы В и витамина К, а также расщепляют клетчатку.
6. Многие виды бактерий служат причиной болезней животных и человека (дизентерии, холеры, туберкулеза, сифилиса, бруцеллеза и др.), а также растений (бактериозы - увядание, пятнистость и др.).
ПОДЦАРСТВО ОКСИФОТОБАКТЕРИИ (OXYPHOTOBACTERIA)
ОТДЕЛ ЦИАНОБАКТЕРИИ (СИНЕ-ЗЕЛЕНЫЕ ВОДОРОСЛИ) - Cyanobacteria
Сине-зеленые водоросли (цианобактерии) - очень древние организмы, возникшие свыше 3 млрд лет назад, относящиеся к фототрофным прокариотам. Несмотря на близость их организации к прокариотам (строение клеток, наличие муреина, способность фиксировать азот), имеются и отличия от бактерий: наличие фотосистемы II; организация пигментной системы, аналогичной красным водорослям; фотосинтез с выделением кислорода; водный (в основном пресноводный) образ жизни.
Строение. Клетки (по форме округлые, эллиптические, цилин- дрические, бочонковидные) могут оставаться одиночными, объединяться в колонии или образовывать многоклеточные нити.
Клеточная стенка довольно толстая, содержит некоторое количество целлюлозы, но главными ее компонентами являются иные полисахариды и пектиновые вещества. Подобно многим прокариотам, клеточная стенка сине-зеленых водорослей содержит муреин (гликопептид). В клеточной стенке есть поры - для сообщения между клетками. Поверх клеточной стенки часто выделяется слизь в виде толстого чехла, предохраняющего клетку от высыхания и облегчающего скольжение.
Кроме того, в цианобактериях может фиксироваться атмосферный азот. Настоящие вакуоли с клеточным соком редки, но в цитоплазме многих видов этих водорослей имеются газовые вакуоли, или псевдовакуоли. Нитчатые формы (осциллатория, носток), помимо обычных клеток, имеют более крупные вакуоли, наполненные азо- том, - гетероцисты (рис. 5.4). Гетероцисты способны фиксировать азот, снабжая таким образом азотистыми веществами прочие клетки. Считается, что эти вакуоли регулируют плавучесть клетки и позволяют ей парить в толще воды.
У цианобактерий найдены пигменты: хлорофилл а, несколько каротинов и ксантофиллов, а также фикобиллины, модифицирующие их окраску от сине-зеленой, фиолетовой и красноватой до почти черной (синий фикобиллин - фикоцианин и красный - фикоэритрин). Пигменты располагаются в одиночных тилакоидах. Фотосинтезирующий аппарат цианобактерий имеет фотосистемы I и II, поэтому способен к аэробному фотосинтезу с выделением кислорода. Продукты фотосинтеза могут накапливаться, хотя и в небольших количествах. Чаще всего это гликопротеид, похожий по химическому
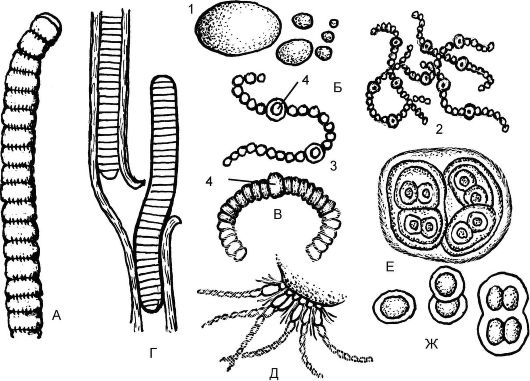
Рис. 5.4. Сине-зеленые водоросли (цианобактерии) (Суаnophyta):
А - осциллатория (Oscillatoria); Б - носток (Nostoc); В - анабена (Аnabaena); Г - лингбия (Lyngbуа); Д - ривулярия (Rivularia); Е - глеокапса (G1eосарsа); Ж - хроококк (Chroococcus); 1 - общий вид; 2 - вид при малом увеличении микроскопа; 3 - отдельная нить при большом увеличении; 4 - гетероцисты анабены
составу на гликоген. Большинство цианобактерий, будучи автотрофными организмами, могут синтезировать все вещества клетки за счет энергии света. Однако они способны и к смешанному типу питания (как гетеротрофы).
Для цианобактерий, как и для бактерий, характерно наличие нуклеоида в центральной части клетки. В качестве запасных включений в цитоплазме имеются: гликоген, волютин, белок цианофицин.
Размножение
1. Вегетативное размножение одноклеточных и колониальных форм происходит за счет деления клетки пополам (см. Бактерии); у нитчатых форм разделение происходит по гетероцистам.
2. В отличие от настоящих бактерий цианобактерии жгутиков не имеют. Половой процесс у них отсутствует.
Распространение и значение
За счет своей миксотрофности (авто- и гетеротрофный способ питания), а также фиксации азота сине-зеленые водоросли встречаются в воде и на суше. Их массовое развитие в воде вызывает ее «цветение». При гниении клеток водорослей в воду выделяются токсические вещества, поэтому она и приобретает неприятный запах. Благодаря газовым вакуолям цианеи всплывают на поверхность воды, на которой образуется маслянистая пленка, не пропускающая воздух; вода становится непригодной для питья и вызывает массо- вую гибель рыб (замор). На суше они образуют налеты на камнях и коре деревьев, входя в симбиоз с грибами и образуя лишайники. Среди относительно немногих случаев полезного использования человеком цианобактерий - искусственное разведение видов анабена (Anabаena oryza) на рисовых полях в тропиках с целью обогащения почвы соединениями азота. Внесение в почву перед посевом анабены повышает урожайность риса на 50% за счет фиксации азота из атмосферы. Некоторые виды спирулины (Spirulina maxima) содержат физиологически активные вещества, йодсодержащие гормоны и простагландины и рекомендуются для использования в качестве пищевых добавок.
Подимперия Ядерные организмы, или Эукариоты (Eucariota) Царство Протоктисты (Protoctista)
Еще в
определенных стадиях развития. Питание - путем заглатывания, всасывания, если автотрофное - с помощью своеобразных пластид. Размножение вегетативное, бесполое и половое.
Грибоподобные протоктисты
Это - гетеротрофы, их репродуктивные клетки подвижны и снабжены 1 или 2 ундулиподиями (в отличие от грибов, у которых ундулиподии отсутствуют), реже подвижны сами амебоидные организмы. В связи с тем что на каком-то отрезке жизни они образуют клеточную стенку, большинство из них в прошлом относили к грибам.
Все представители рассматриваются в ранге отделов, что примерно соответствует типу в зоологической номенклатуре.
ОТДЕЛ ООМИКОТЫ (OOMYCOTA)
Сюда входят водные грибы, которые обитают на растительных остатках, трупах насекомых, или паразиты беспозвоночных, рыб, амфибий. Наиболее высокоорганизованные - облигатные паразиты наземных растений. Это могут быть примитивные одноклеточные грибы и грибы с хорошо развитым неклеточным мицелием. Они имеют двужгутиковые зооспоры. Клеточные стенки состоят из целлюлозы и пектинов, хитин отсутствует.
Очень важное практическое значение имеет род Фитофтора (Phytophtora), насчитывающий около 70 видов и имеющий облигатных и факультативных паразитов. К факультативным паразитам принадлежат виды, поражающие яблоню, цитрусовые, эвкалипты, а также картофельный гриб (Ph. infestans). Этот паразит поражает ботву и клубни картофеля, листья и плоды томатов и других пасленовых. В клетки листа внедряются гаустории, и мицелий проходит по межклетникам. На листьях появляются бурые пятна, пораженные участки быстро отмирают. На нижней стороне листа выходят спорангиеносцы с лимоновидными зооспорангиями. В воде из зооспорангия выходят двужгутиковые зооспоры, которые после периода подвижности развиваются в гифы, проникающие в лист или клубень. Зооспоры с водой могут заражать и молодые клубни. Гриб может жить как сапрофит на растительных остатках, а при благоприятных усло- виях перейти к паразитизму. Оогамное половое размножение встречается достаточно редко. Потери урожая от фитофторы значительны. К мерам борьбы с облигатными и факультативными паразитами относят соблюдение агротехники и тщательную дезинфекцию хранилищ.
ОТДЕЛ ХИТРИДИОМИКОТЫ (CHYTRIDIOMYCOTA)
Насчитывает около 500 видов грибов, тесно связанных с водой. Вегетативное тело представлено голой плазменной массой или зачаточным мицелием. В основном являются паразитами водорослей, водных высших растений, грибов и беспозвоночных животных. Многие представители вызывают различные болезни сельскохозяйственных растений.
Синхитрии. Наиболее важным из 200 представителей этого рода является Synchytrium endobioticum - возбудитель рака картофеля. Тело паразита представлено плазмодием. Бесполое размножение происходит при помощи зооспор, половой процесс - изогамный. Из зиготы образуется циста, сохраняющая жизнеспособность в течение многих лет. На пораженных этим грибом клубнях картофеля образуются напоминающие губку бугристые опухоли, которые разрастаются, чернеют и разрушаются. Это может повторяться несколько раз в течение года. К осени в клубнях образуются цисты. Потери урожая составляют 40-60%. Основные меры борьбы - выведение устойчивых к грибу сортов и обеззараживание почвы.
Протоктисты водоросли
Тело водорослей не расчленено на органы, а представлено слоевищем (талломом). Слоевище может быть одноклеточным, колониальным или многоклеточным. Водоросли - древние представители ор- ганического мира, возникшие в протерозое (800-900 млн лет назад). Считается, что все отделы водорослей ведут свое происхождение от различных групп одноклеточных организмов, т.е. не имеют родственных связей друг с другом. Их отделы различаются набором пигментов, строением пластид, продуктами фотосинтеза, накапливающимися в клетке, числом и строением ундулиподиев, а также особенностями строения митохондрий. Большинство водорослей живет в пресновод- ных водоемах и морях и делится на 2 большие экологические группы: планктонные и бентосные. Водоросли, входящие в планктон, составляют его фототрофную часть, так называемый фитопланктон. Последний производит основную массу органических веществ, за счет которых через цепи питания существует весь остальной живой мир воды. К бентосным водорослям относят макроорганизмы, прикрепленные к донному субстрату. Они служат в основном кормом для рыб и млекопитающих. Однако существуют экологические группы наземных (на камнях, скалах), почвенных водорослей, водорослей снега, льда и т.п. К микроскопическим водорослям льда и снега относится одно-
клеточная водоросль Хламидомонада снежная (Chlamydomonas nivalis), вызывающая покраснение снега - «красный снег».
Клеточная организация водорослей
Клеточная стенка водорослей состоит из целлюлозы, гемицеллюлозы и пектиновых веществ. У некоторых представителей в состав клеточной стенки входят такие вещества, как кремний, карбонат кальция, альговая кислота и др. У большинства водорослей цитоплазма расположена тонким постенным слоем, окружая крупную вакуоль. Кроме характерных для всех растений органоидов (ЭПС, рибосомы, аппарат Гольджи, пластиды, митохондрии и ядро), у многих водорослей присутствуют центриоли. Интересно отметить, что пластиды чрезвычайно разнообразны по форме и составу пигмента. Многие пластиды имеют небольшие включения - пиреноиды, на которых откладываются крахмальные зерна. Как правило, пластиды водорослей называют хроматофорами.
Размножение:
• вегетативное - например фрагментация (разрыв на отдельные участки таллома);
• бесполое - с помощью спор: подвижных, снабженных ундулиподиями (зооспоры), или с помощью апланоспор - неподвижных клеток, лишенных ундулиподиев;
• половое - слияние или копуляция 2 половых клеток-гамет, образующихся в специальных материнских клетках - гаметангиях. У водорослей встречаются хологамия, гетерогамия, изогамия и оогамия. У некоторых зеленых водорослей наблюдается половой процесс, называемый конъюгацией. При конъюгации сливается содержимое 2 недифференцированных вегетативных клеток, выполняющих функции гамет.
У водорослей впервые в цикле развития возникло чередование полового гаплоидного и бесполого диплоидного поколения.
ОТДЕЛ ЭВГЛЕНОВЫЕ (EUGLENOPHYCOTA)
Среди примерно 1 тыс. представителей этого отдела - эвгленовые водоросли - одноклеточные микроскопические организмы (рис. 5.5). Эвгленовые водоросли являются миксотрофами, так как для них характерно автотрофное питание, а для некоторых видов - голозойное (путем заглатывания с помощью ротового аппарата).
Эвгленовые обитают в пресных водоемах, богатых органикой. Они имеют плотную эластичную оболочку пелликулу и 2 ундулиподия.
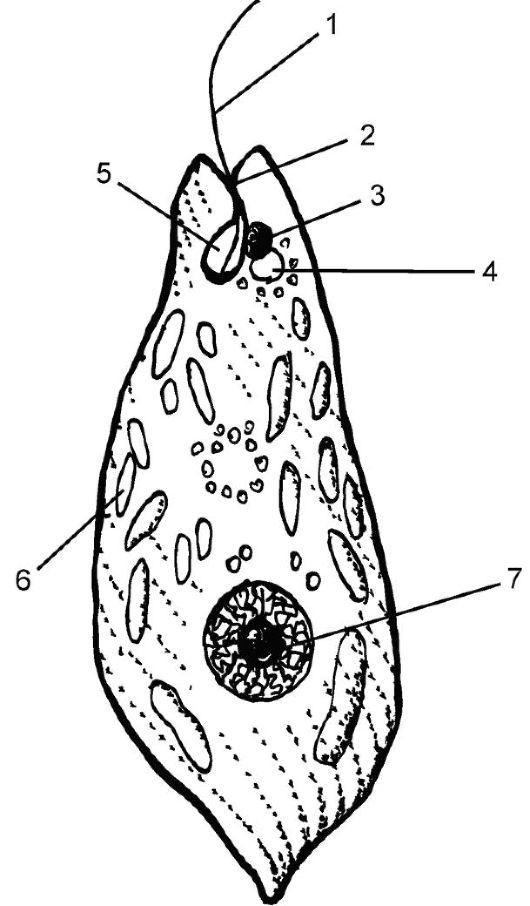
Рис. 5.5. Эвглена зеленая:
1, 2 - жгутик; 3 - стигма (светочув- ствительное пятнышко); 4 - выделительная (сократительная) вакуоль; 5 - резервуар сократительной вакуоли; 6 - хлоропласты; 7 - ядро
Один из них (длинный) участвует в движении и обычно направлен вперед, а короткий в движении не участвует. Пелликула достаточно гибкая, что позволяет клетке принимать разную форму. Это проис- ходит за счет сокращения крошечных фибрилл в цитоплазме, называемых мионемами; сокращение влечет за собой скольжение полосок пелликулы, и клетка изменяет форму тела. Это явление называют эвгленоидным движением.
Клетка водоросли имеет одно ядро, пиреноиды, пластиды, форма которых разная (зернистая, лентовидная, блюдцевидная). Пластиды содержат пигменты: хлорофиллы а и в, β-каротин, ксантофиллы и окружены прилегающей к ним мембраной ЭПС. Основной продукт ассимиляции - парамилон - полимер глюкозы, близкий по составу к ламинарину. У некоторых эвгленовых имеется фоточувствительный красный глазок (стигма), содержащий β-каротин.
Бесполое размножение происходит за счет продольного деления надвое. Половое размножение не выявлено.
ОТДЕЛ КРАСНЫЕ ВОДОРОСЛИ (БАГРЯНКИ) - Rhodophycota (Rhodophyta)
Красные водоросли представляют собой древнюю группу многоклеточных организмов, во многих учебниках выделяемых в особое подцарство. Древнейшие багрянки были обнаружены в кембрии (возраст 550 млн лет). Описано около 41 тыс. видов, относящихся к 650 родам.
Таллом (вегетативное тело водоросли) у багрянок имеет вид разветвленных многоклеточных нитей ярко-красного цвета, прикрепленных к субстрату с помощью ризоидов (рис. 5.6). Нередко эти нити склеены слизистым веществом, поэтому они скользкие на ощупь. В состав слизи входят сульфатированные полимеры галактозы, например агар. Клеточная стенка двуслойная. Наружный слой состоит из пектиновых соединений, внутренний - из гемицеллюлоз, которые могут сильно набухать и образовывать слизистую массу. Пектиновые вещества способны растворяться в кипящей воде, образуя коллоидный раствор.
В хроматофорах красных водорослей содержатся пигменты: хлорофиллаид, каротиноиды и фикобиллины (красные фикоэритрины и синие фикоцианины, дающие различную окраску в зависимости от преобладания тех или иных пигментов).
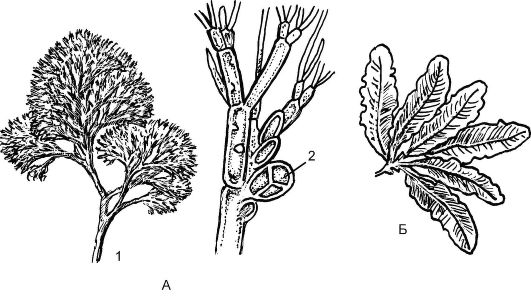
Рис. 5.6. Багрянки:
А - каллитамнион: 1 - внешний вид; 2 - тетраспорангии; Б - делоосприя
Продуктом ассимиляции является «багрянковый крахмал», откладывающийся вне связи с хлоропластами. Этот полисахарид более близок к амилопектину и гликогену, чем к крахмалу.
Размножение может быть вегетативным (частями таллома), бесполым (с помощью неподвижных тетраспор) и половым (оогамия с чередованием ядерных фаз). Женский половой орган - карпогон - состоит из расширенной части: брюшка с яйцеклеткой и выроста - трихогины. Мужские органы - антеридии содержат мелкие, голые, лишенные ундулиподиев, спермации. Последние с током воды пассивно движутся и прилипают к трихогине. После оплодотворения яйцеклетки образуется зигота, а карпогон отделяется перегородкой от трихогины. Из зиготы формируются диплоидные карпоспоры, а из них - диплоидный таллом. На талломе в результате мейоза образуются гаплоидные тетраспоры. Они прорастают в гаплоидный таллом, на котором вновь образуются половые органы красных водорослей.
Среди багрянок ценны виды родов Родимения (Rodimenia) и Порфира (Porphyra). Некоторые багрянки из рода Анфельция служат сырьем для получения агар-агара, основой которого являются фикоколлоиды, используемые в микробиологии (для приготовления сред при культивировании микроорганизмов), а также в пищевой промышленности.
ОТДЕЛ БУРЫЕ ВОДОРОСЛИ (PHAEOPHYTA)
Бурые водоросли (около 1500 видов) - многоклеточные организмы, обитающие во всех морях. Это самые крупные из известных водорослей. В хозяйственном отношении наиболее важен род Ламинария (Laminaria). Ламинария сахарная - представитель северных морей и Дальнего Востока. Отдельные особи достигают 10-60 м в длину (рис. 5.7).
Общая характеристика
Тело ламинарии разделено на части: листовая пластина, стволик и ризоиды. Листовая пластинка каждый год обновляется, а стволик и ризоиды зимуют. Клетки содержат одно ядро, несколько вакуолей и имеют сильно ослизняющиеся стенки. Хроматофор округлый, в виде зерен, содержит пигменты: хлорофилл, каротин, ксантофилл и фукоксантин. Избыток фукоксантина придает водоросли бурую окраску. Запасной полисахарид - ламинарин, откладывающийся вне хлоропласта (в цитоплазме).
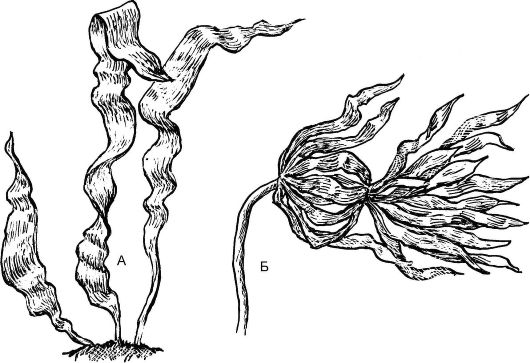
Рис. 5.7. Бурые водоросли:
А - ламинария сахарная (Laminaria saccharina); Б - ламинария северная (L. hyperborea)
Размножение: вегетативное (частями таллома); бесполое (зооспорами); половое (оогомия с чередованием диплоидной и гаплоидной ядерных фаз).
Половое размножение - оогамия
Взрослое растение ламинарии является спорофитом (бесполая фаза). В зооспорангиях, находящихся на листовой пластине, в результате мейоза образуются подвижные зооспоры. Подвижность им придают 2 неравных жгутика, прикрепленных к зооспоре сбоку. Зооспоры попадают на дно и прорастают в заростки (гаметофиты) - микроскопические образования, несущие гаметы. На одних гаплоидных заростках образуются женские половые органы - оогоний с яйцеклеткой, на других - мужские - антеридий со сперматозоидами. Вышедшая яйцеклетка из оогония оплодотворяется сперматозоидом с образованием диплоидной зиготы, которая сразу же, без периода покоя, прорастает во взрослую особь (рис. 5.8).
В северных морях широко распространен род Фукус (Fucus). Представители этого рода в основном обитают в береговой зоне и имеют дихотомически разветвленное слоевище темно-бурого цвета, достигающее
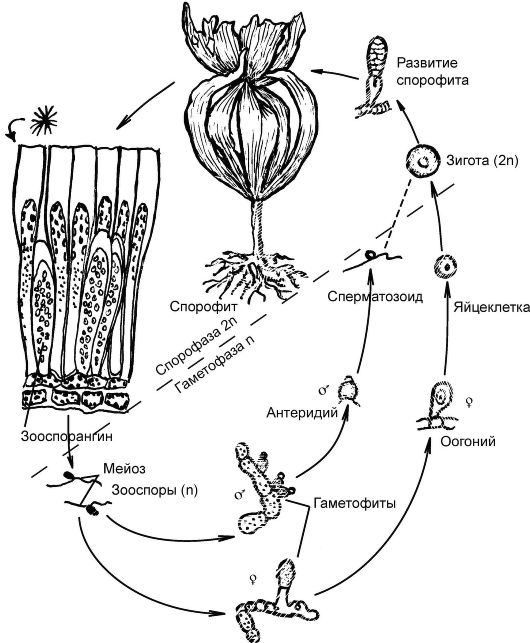
Рис. 5.8. Чередование поколений и смена ядерных фаз у бурой водоросли ламинарии
Бурые водоросли находят различное применение в хозяйственной деятельности человека. Так, из них получают агар, альгинаты (вещества, широко используемые при приготовлении консервов, красящих и клеящих веществ), а также кровезаменитель и маннит. Используют
как добавку к кормам сельскохозяйственных животных. Кроме того, они служат сырьем для получения йода и брома.
ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ (CHLOROPHYTA)
Это самый обширный среди водорослей отдел, насчитывающий свыше 13 тыс. видов. Включает в себя несколько классов, наиболее распространенные из которых - равножгутиковые, сцеплянки и харовые. Зеленые водоросли разнообразны по внешнему виду: одноклеточные, колониальные, многоклеточные, нитчатые и т.д. Представители отдела зеленых водорослей большей частью обитают в пресных водоемах; у них зеленая окраска из-за преобладания хлорофилла.
Класс Равножгутиковые. Порядок вольвоксовые. Род Хламидомонада (Chlamydomonas)
Род Хламидомонада включает около 320 видов одноклеточных организмов, обитающих в пресных водоемах, лужах, канавах.
Общая характеристика. Одноклеточная хламидомонада (рис. 5.9) совсем не похожа на растение, так как она активно движется, и у нее имеются пульсирующие вакуоли. Для нее характерны типичные для эукариот органеллы: аппарат Гольджи, митохондрии, рибосомы, хроматофоры и мелкие вакуоли. В хроматофорах многих водорослей выявлена особая структура - пиреноид. Это белковое образование, осуществляющее фиксацию двуокиси углерода. Пиреноид участвует в запасании углеводов (например крахмала). Красный глазок (стигма)
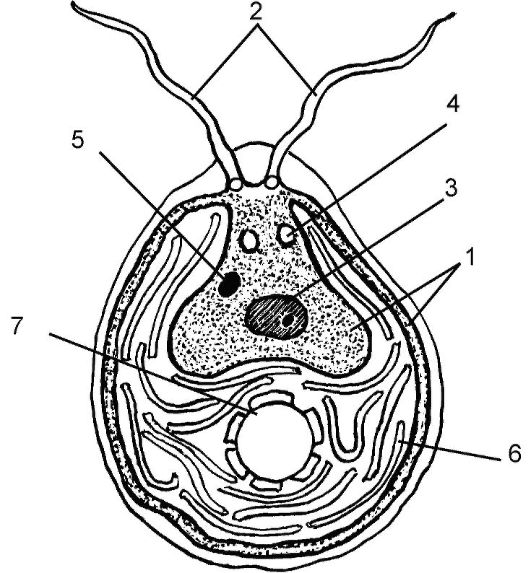
Рис. 5.9. Строение зеленой водоросли (на примере хламидомонады - Chlamydomonas sp.):
1 - цитоплазма; 2 - ундулиподии; 3 - ядро; 4 - пульсирующие вакуоли; 5- стигма «красный глазок»; 6 - хроматофор; 7- пиреноид
воспринимает изменения в интенсивности освещения, и клетка либо перемещается туда, где интенсивность света оптимальна для фото- синтеза, либо остается на месте, если освещенность достаточная. Такая ответная реакция называется фототаксисом.
Размножение. Взрослая особь хламидомонады гаплоидна и имеет 2 типа размножения - бесполое и половое.
Бесполое размножение осуществляется с помощью зооспор (рис. 5.10). Родительская клетка теряет жгутики, стенки ее ослизняются (обычно это происходит при подсыхании водоема), и протопласт клетки делится на 2 дочерних протопласта. В это время происходит митотическое деление ядра; кроме того, делится и хроматофор. У дочерних протопластов образуются новые клеточные стенки, новые глазки и жгутики. В образовании жгутиков участвуют базальные тельца. Клеточная стенка родительской клетки ослизняется, и дочерние клетки, которые называются зооспорами, выходят наружу. Из каждой зооспоры формируется полноценная взрослая клетка.
Половое размножение. Одни виды хламидомонады гомоталличны, другие гетероталличны. При этом разные виды могут быть изогамными, анизогамными или оогамными.
Рассмотрим размножение хламидомонады на примере ее изогамных видов.
Гаплоидные гетероталличные родительские клетки хламидомонады сближаются, сбрасывают свои оболочки и взаимодействуют в дальнейшем как гаметы. Далее происходит изогамия с образованием диплоидной зиготы с 4 жгутиками. Позднее зигота отбрасывает жгутики, образуя прочную клеточную стенку и накапливая запасные питательные вещества. Далее в зиготе происходит мейоз ядра и высвобождаются 4 протопласта; которые тотчас же образуют по паре жгутиков. Таким образом формируются новые гаплоидные особи хламидомонады. Цикл развития хламидомонады происходит в гаплоидной фазе, диплоидна только зигота (см. рис. 5.10, Б).
Класс Равножгутиковые. Порядок вольвоксовые. Род Вольвокс (Volvox) Колониальная водоросль, имеющая шаровидную форму (2-3 мм). Колонии вольвокса состоят из многих (500-600 тыс.) клеток, расположенных по периферии шара в один слой. Каждая клетка имеет 2 жгутика, обращенных к наружи шара, и по строению напоминает клетку хламидомонады. Внутренняя полость шара занята слизью. Все вегетативные клетки связаны между собой тяжами цитоплазмы и выполняют функции движения и питания (рис. 5.11). Функцию поло-
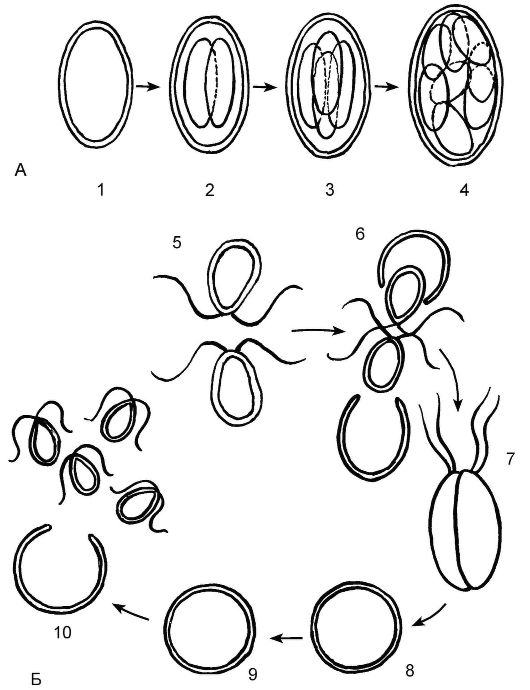
Рис. 5.10. Размножение хламидомонады:
А - бесполое размножение: 1 - родительская клетка; 2 - 1-е деление (два протопласта); 3 - 2-е деление (четыре протопласта); 4 - 3-е деление (восемь протопластов);
Б - половое размножение: 5 - сближение родительских гетероталличных «+» и «-» клеток; 6 - сбрасывание клеточных оболочек и взаимодействие родительских клеток как гамет; 7 - изогамия с образованием зиготы с четырьмя жгутиками; 8 - зигота теряет жгутики; 9 - мейоз зиготы; 10 - образование дочерних клеток (n) в результате мейоза
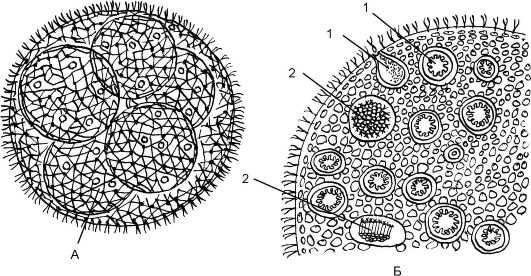
Рис. 5.11. Строение вольвокса:
А - дочерние шары внутри материнского; Б - часть колонии с яйцеклетками (1) и сперматозоидами (2)
вого размножения выполняют другие специализированные клетки, отличающиеся более крупными размерами и отсутствием жгутиков.
Размножение:
• вегетативное размножение осуществляется с помощью дочерних колоний, образующихся в материнских колониях путем последовательных продольных делений их протопластов. Затем дочерние колонии выпадают внутрь материнского шара и освобождаются лишь после ее разрушения;
• при половом размножении часть специализированных клеток преобразуется в оогонии с яйцеклетками. В других специализированных клетках развиваются двужгутиковые сперматозоиды. Сперматозоиды одного материнского шара оплодотворяют яйцеклетки другого с образованием зиготы. Диплоидная зигота называется ооспорой; обычно она прорастает весной. При этом сначала происходит мейоз, а затем многократное деление. В результате возникает пластинка гаплоидных клеток, после чего постепенно формируется новое шаровидное тело вольвокса. Основная форма существования простейших в колонии происходит в гаплоидной фазе, диплоидна только зигота (рис. 5.12).
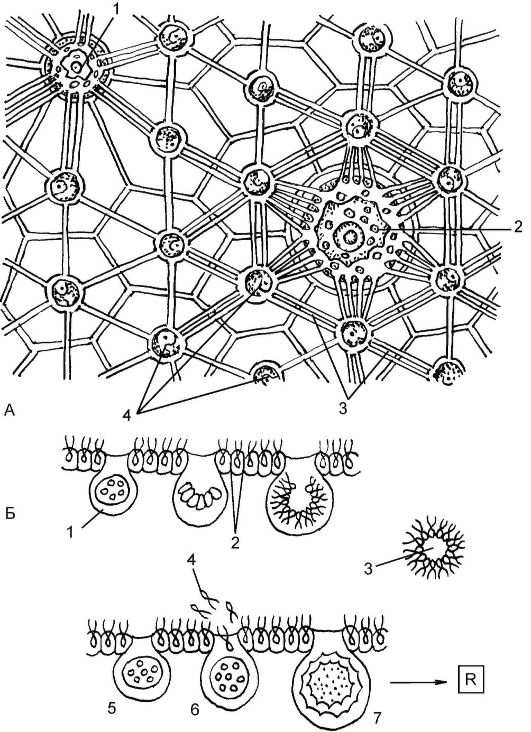
Рис. 5.12. Половое размножение вольвокса:
А - вид колонии с поверхности: 1 - клетка, образующая спермии; 2 - клетка, образующая яйца; 3 - цитоплазматические связи; 4 - соматические клетки; Б - образование половых клеток и зиготы: 1 - клетка, образующая спермии; 2 - соматические клетки; 3 - шаровидная структура, образованная спермиями; 4 - спермии; 5 - клетка, образующая яйца; 6 - оплодотворенное яйцо; 7 - диплоидная зигота; R - мейоз
Класс Равножгутиковые. Порядок хлорококковые или протококковые водоросли. Род Хлорелла (Chlorella)
Род Хлорелла объединяет одноклеточные водоросли, широко распространенные в пресных водоемах, почве. Клетки покрыты обо-
лочкой, содержат чашевидный хроматофор, ядро - одно. Половой процесс отсутствует. При бесполом размножении материнская клет- ка делится на 4-10 частей. Образуются неподвижные, т.е. без жгутиков, автоспоры, которые еще в материнской клетке покрываются собственной оболочкой и выходят из нее после разрыва материнской оболочки (рис. 5.13).
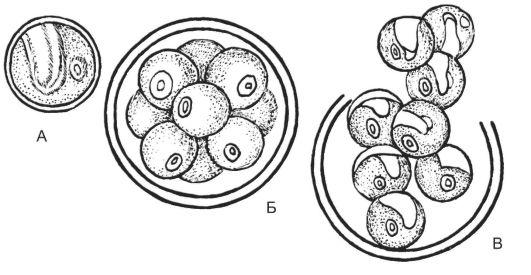
Рис. 5.13. Хлорококковые - хлорелла: А - вегетативная клетка; Б - образование автоспор; В - выход автоспор наружу
Хлорелла характеризуется очень быстрыми темпами размножения. Каждая клетка в течение суток может образовывать до 10 дочерних. Хлорелла способна использовать от 10 до 12% световой энергии (у наземных растений - 1-2%). В сухом веществе содержится: 50% белков, 22% жиров, 12% углеводов, 10% минеральных солей, витамины В, К, С (больше, чем в лимонном соке). Свойство хлореллы быстро накапливать биомассу используется для получения дешевого корма. Она получила применение при полетах космонавтов как источник пищи и кислорода.
Класс Сцеплянки. Род Спирогира (Spirogyra)
У представителей этого класса наблюдается особый половой процесс (конъюгация), а образование зооспор отсутствует.
Одним из представителей класса является спирогира - многоклеточная нитчатая водоросль, свободно плавающая на поверхности пресных водоемов. В центре каждой клетки находится ядро, «подве-
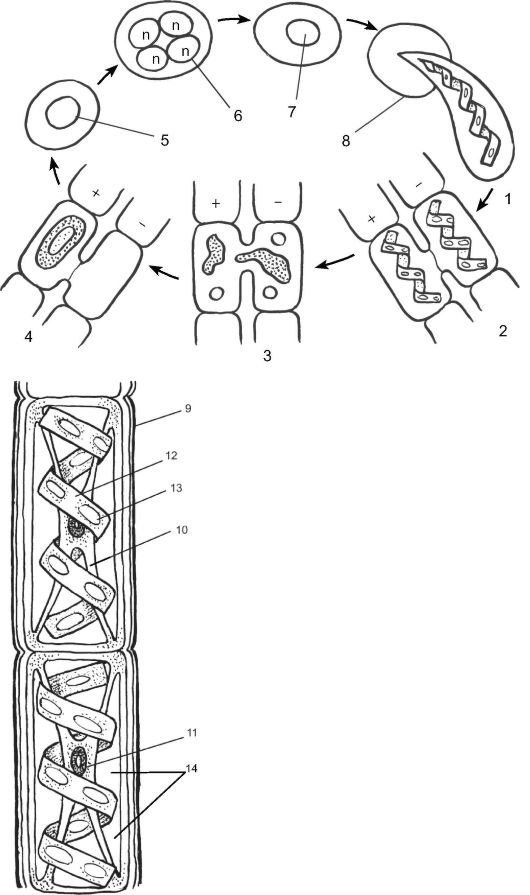
Рис. 5.14. Чередование поколений и смена ядерных фаз у зеленой водоросли
спирогиры (Spirogyra sp.): 1 - часть таллома, 2-4 - последовательные стадии конъюгации; 4,5 - зигота; 6 - зигота с 4 гаплоидными ядрами, возникающими в результате мейоза; 7 - зигота с 1 функционирующим гаплоидным ядром (прочие отмерли); 8 - прорастание зиготы; 9 - клеточная стенка; 10 - цитоплазма; 11 - ядро; 12 - хроматофор; 13 - пиреноиды; 14 - вакуоли
шенное» на тяжах цитоплазмы (рис. 5.14). Имеется спирально закрученный хроматофор с пиреноидами. Размножение:
• вегетативное размножение происходит частями таллома, т.е. путем разрыва нитей на отдельные участки или даже отдельные клетки;
• половой процесс - конъюгация. При конъюгации у 2 гетероталличных параллельно располагающихся нитей водоросли у супротивных клеток образуются выросты. В месте соприкосновения этих 2 выростов оболочки растворяются и образуется копуляционный канал. По этому каналу происходит слияние содержимого 2 клеток с образованием зиготы. Зигота округляется, покрывается толстой оболочкой и переходит в состояние покоя. После периода покоя зигота редукционно делится и образуются 4 новые клетки, из которых 3 отмирают, а 1 прорастает в новую особь. Таким образом, спирогира проходит свой жизненный цикл в гаплоидной фазе, а диплоидна только зигота.
Царство Грибы (Mycota, Fungi)
Существует около 100 тыс. видов грибов, разнообразных по внешнему виду и распространенных во всех географических зонах Земли. Это одно из самых больших царств организмов, появившееся около 450 млн лет назад.
Общая характеристика
Тело гриба представляет собой мицелий, состоящий из тонких нитей - гиф. Мицелий имеет тесную связь с субстратом, что обусловлено осмотическим поглощением питательных веществ. У высших грибов мицелий разделен на отдельные клетки перегородками - септами, т.е. у них септированнный (клеточный) мицелий. Низшие грибы имеют неклеточное строение мицелия, так как его гифы не разделены на перегородки, а представляют собой как бы одну разветвленную клетку со множеством ядер.
Грибы обособлены по своей морфофизиологической организации от остального мира живых существ. Их нельзя отнести ни к растениям, ни к животным. Существуют две теории происхождения грибов: животная и растительная, поскольку клетки грибов имеют признаки как животной, так и растительной клетки (табл. 5.2).
Теория растительного происхождения грибов предполагает их происхождение от зеленых водорослей, из чего следует, что грибы прежде всего явно регрессивная группа растений, утративших хлоропласты.
Теория животного происхождения основывается на том, что грибы изначально являются бесхлорофильными организмами, т.е. происходят от простейших гетеротрофных организмов, а не от водорослей. Эта теория предпочтительнее, поскольку бесхлорофильные водоросли, относимые к зеленым, в качестве запасного продукта накапливают крахмал, в то время как у грибов крахмала нет.
Таблица 5.2. Особенности строения грибной клетки

Грибы - это гетеротрофы. Как и бактериям, им свойственно внеклеточное переваривание, осуществляемое за счет выделения во внешнюю среду ферментов. Всасывание расщепленных питательных веществ происходит осмотически, всей поверхностью тела. Клетки мицелия в качестве запасных питательных веществ откладывают углеводы в виде гликогена, жиры в виде капель липидов, а в вакуолях - белки.
Грибы могут быть паразитами, сапрофитами и симбионтами. Среди паразитов выделяют облигатные и факультативные. Облигатные паразиты узко специализированы и имеют небольшой круг хозяев (мучнеросные, ржавчинные и головневые). Циклы развития гриба строго согласованы с продолжительностью жизни хозяина. При этом на гифах гриба образуются присоски-гаустории, которые внедряются в клетку для поглощения из нее пищи. Факультативные паразиты лишь временно способны к паразитизму. Они не образуют гаусторий, а с помощью ферментов разрушают ткань растения, поглощают содержимое клеток и вызывают гибель хозяина. В дальнейшем они питаются сапрофитно мертвыми остатками (например, картофельный гриб фитофтора, поражающий многие растения семейства Пасленовые).
Грибы способны вступать в симбиоз с высшими растениями, образуя микоризу (грибокорень). Грибы используют углеводы, синтезируемые растением, и добывают для него (за счет минерализации органических соединений) различные соединения с азотом, фосфором, вырабатывают активаторы роста и витаминоподобные вещества.
Размножаться грибы могут вегетативно, бесполым и половым путем.
Вегетативное размножение может происходить частями мицелия (почти у всех грибов), почкованием (дрожжи). Бесполое размножение происходит за счет образования зооспор, спорангиоспор и конидий.
Зооспоры образуются у грибов, ведущих водный образ жизни (хи- тридиомицеты, оомицеты). Их подвижность обеспечивают жгутики (их 1 или 2). Образуются внутри одноклеточных зооспорангиев и при созревании выходят в воду. Покрываются оболочкой и прорастают в новую особь.
Спорангиеспоры образуются эндогенно - внутри одноклеточных спорангиев, возникающих на гифах-спорангиеносцах. В одном спорангии может быть до 10 тыс. спор, которые при созревании выходят из спорангия и распространяются ветром на значительные расстояния. Попав в благоприятные условия, спора прорастает в новый мицелий (например, у мукора).
Конидии образуются экзогенно на особых гифах - конидиеносцах. Конидии образуют цепочки, отчленяются и в благоприятной среде прорастают в новый мицелий (например, у пеницилла).
Половое размножение у низших грибов происходит:
• при слиянии гамет - гаметогамия (изогамия, гетерогамия и оогамия);
• при слиянии двух многоядерных специализированных половых органов (гаметангиев) - зигогамия.
Половое размножение у высших грибов:
• гаметангиогамия; архикарп - женский гаметангий, антеридий - мужской (у сумчатых грибов);
• соматогамия - слияние гаплоидных соматических клеток гетероталличных гиф (+ и - физиологически различных гиф), например у высших базидиальных грибов.
Половой процесс всегда завершается образованием диплоидной зиготы, ее мейотическим делением и спороношением.
К низшим грибам относится отдел зигомикоты, к высшим - отделы: сумчатые, базидиомикоты, несовершенные.
ОТДЕЛ ЗИГОМИКОТЫ (ZYGOMYCOTA)
Почти все представители этого класса ведут наземный образ жизни. Среди них есть как сапрофиты, так и паразиты высших грибов, насекомых, животных и человека. Мицелий в основном неклеточный, клеточные стенки содержат хитин. Наиболее известен род мукор (головчатая плесень) - Mucor.
Мукор широко распространен в природе как белая плесень (рис. 5.15). Сапрофит по способу питания; развивается на почве, пищевых продуктах. Гифы мицелия представляют собой вытянутую разросшуюся гигантскую клетку с множеством ядер (неклеточное строение). Ядра - с гаплоидным набором хромосом (n). На мицелии развиваются многочисленные вертикальные спорангиеносцы с буро-черными спорангиями. В результате митоза содержимое спорангия распадается на множество спор (до 10 тыс.). После созревания оболочка спорангия лопается, и споры рассеиваются, прорастая в новые особи. Размножение может быть бесполым (спорами), вегетативным (частями мицелия), редко - половым (зигогамия).
Рис. 5.15. Строение мукора (Мuсоr mucedo): 1 - гифа; 2 - мицелий; 3 - спорангиеносец; 4 - спорангий со спорами
При зигогамии (рис. 5.16) физиологически различные гифы - гетероталличные, условно обозначающиеся как + и -, начинают расти навстречу друг к другу. На концах гиф образуются гаметангии, от- деляющиеся перегородками от остальной гифы. Далее происходит гаметангиогамия, состоящая в слиянии 2 специализированных половых структур (гаметангиев), не дифференцированных на гаметы, и образуется зигота с множеством диплоидных ядер. Зигота покрывается толстой бурой оболочкой. После периода покоя ядра претер- певают мейоз, а зигота прорастает в зародышевый спорангий. В него переходят гаплоидные ядра + и-, образовавшиеся после мейоза. В спорангии формируются споры, после их созревания спорангий вскрывается, споры рассеиваются и прорастают в новые мицелии (+
и -).
Некоторые мукоровые грибы вызывают микоз (мукоромикоз) легких (ложный туберкулез), головного мозга и других органов че-
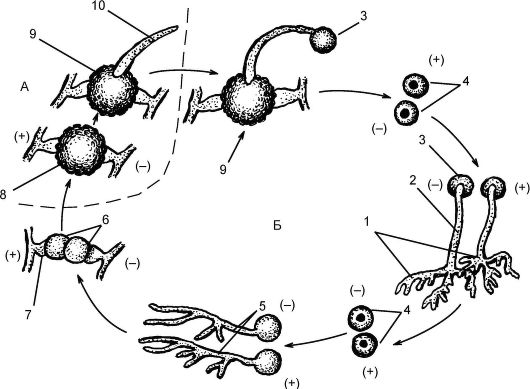
Рис. 5.16. Жизненный цикл мукора (Мuсоr): А - гаплоидная фаза; Б - диплоидная фаза: 1 - два гетероталличных (противоположных по физиологическому знаку) мицелия; 2 - спорангиеносец; 3 - спорангий; 4 - споры; 5 - прорастание спор; 6 - гаметангий; 7 - подвесок; 8 - зигоспора; 9 - прорастающая зигоспора; 10 - прорастающий мицелий
ловека, а также сельскохозяйственных растений. Многие виды рода обладают высокой ферментативной активностью, что используется при получении «соевого сыра» из семян сои, спирта из клубней кар- тофеля и т.д.
ОТДЕЛ СУМЧАТЫЕ ГРИБЫ, ИЛИ АСКОМИКОТЫ (ASCOMYCOTA)
Это один из наиболее обширных классов грибов, включающий более 30 тыс. видов. К этому классу относятся дрожжи, представ- ленные одиночными почкующимися клетками, и грибы с крупными плодовыми телами, например сморчки и строчки. Аскомикоты широко распространены в природе во всех природных зонах. По способу питания это сапрофиты. Мицелий сумчатых грибов является септированным, т.е. разделен на клетки (с гаплоидным набором хромосом). Характерный признак аскомикот - наличие сумок (аск), образующихся в результате полового процесса. Сумки - замкнутые структуры, содержащие определенное количество аскоспор (споры полового размножения) и образующиеся в результате мейоза.
У многих аскомикот сумки образуются в плодовых телах (подкласс Плодосумчатые). Различают 3 типа плодовых тел: клейстотеций, перитеций и апотеций. У других представителей сумки лежат открыто на мицелии (подкласс Голосумчатые).
Большая роль в цикле развития принадлежит и бесполому размножению. Споры бесполого размножения - конидии - образуются в результате митоза на мицелии с гаплоидными ядрами (n) или кони- диеносцах различного строения.
Подкласс Голосумчатые (Hemiascomycetidae)
Наиболее распространенным и имеющим практическое значение является род Дрожжи (Saccharomyces). Дрожжи представлены единичными, овальными клетками (рис. 5.17). Для дрожжей харак- терно вегетативное размножение, осуществляющееся почкованием; для этого им необходимы питательная среда, наличие в ней сахара и определенная температура. При неблагоприятных условиях происходит половой процесс; при слиянии 2 гаплоидных дочерних клеток (хологамия) образуется зигота, превращающаяся в сумку. В результате мейоза в сумке образуются четыре споры (аскоспоры), прорастающие в новые дрожжевые клетки.
Пекарские дрожжи (Saccharomyces cerevisiae) объединяют многие выведенные в культуре дрожжи: спиртовые, пивные, винные, хлебопекарные. Все эти дрожжи разлагают сахар на этиловый спирт и СО2. Так, при добавлении дрожжей в тесто они начинают разлагать имею-
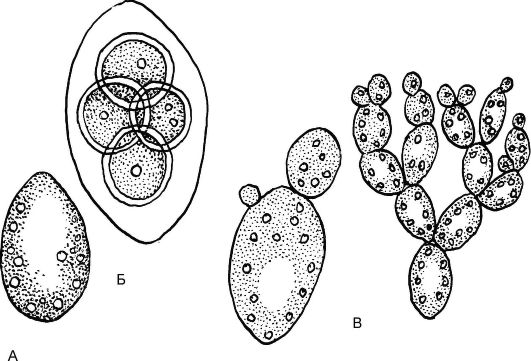
Рис. 5.17. Дрожжи пивные (Saccharomyces cerevisiae): А - одноклеточный таллом; Б - сумка с аскоспорами; В - почкование
щуюся там глюкозу, образующуюся из крахмала. При этом выделяется СО2, обеспечивающий тесту пористость и увеличение объема. При выпечке этанол и СО2 улетучиваются.
Дрожжи являются ценным пищевым и кормовым продуктом. Содержат до 50% белка, а также жиры и углеводы. Синтезируют в больших количествах витамины, особенно В2. Их используют при лечении малокровия, а также как источник белка при добавлении в кормовые продукты в животноводстве и птицеводстве.
Подкласс Плодосумчатые (Carpoascomycetidae)
Представители этого подкласса характеризуются наличием плодовых тел, в которых находятся сумки. Плодовые тела образуются за счет плотного сплетения гаплоидных и дикарионных (двуядерных) гиф, называемых также аскогенными. Плодовые тела (аскокарпы) бывают 3 типов: замкнутые (закрытые) - клейстотеции, полузамкнутые - перитеции, незамкнутые (открытые) - апотеции.
К грибам-паразитам относится род Спорынья (Claviceps). Большинство представителей рода паразитируют на злаках - культивируемых (рожь, пшеница, ячмень) и дикорастущих (тимофеевка, пырей, костер). Самый распространенный вид - Спорынья пурпурная (Claviceps purpurea).
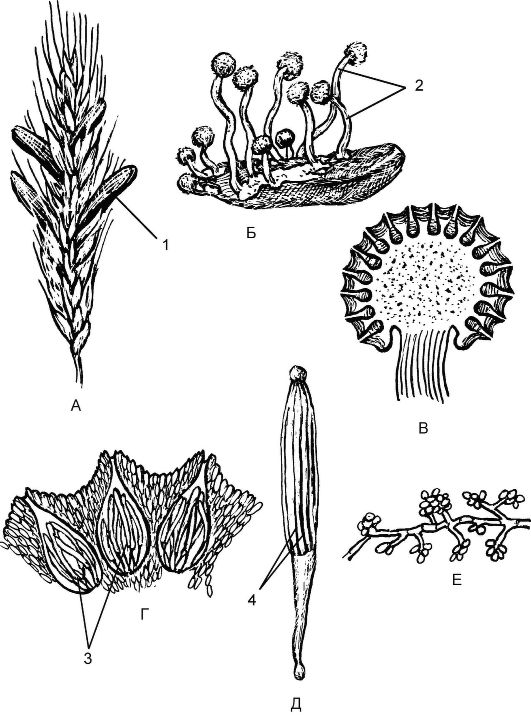
Рис. 5.18. Спорынья пурпурная (Claviceps рurpurеа): А - колос ржи со склероциями (1); Б - стромы (2), выросшие на перезимовавшем склероции; В - продольный срез через строму с перитециями; Г - продольный срез через перитеции (3) с сумками; Д - сумка с нитевидными аскоспорами (4); Е - конидиальное спороношение
Цикл развития спорыньи протекает со сменой ядерных фаз (рис. 5.18). Так, осенью на злаковых растениях образуются склеро- ции - темно-фиолетовые снаружи и белые внутри рожки, представляющие собой мицелий гриба (обезвоженные гифы) в стадии покоя. На зиму склероции выпадают из злаков на почву и зимуют в ней. Весной на почве склероции прорастают, образуя нитевидные выросты, увенчанные головками - стромами. В этих стромах в результате полового процесса образуются плодовые тела - перитеции, заполнен- ные длинными цилиндрическими сумками (асками), содержащими нитевидные аскоспоры - споры полового размножения (рис. 5.19). Созревание спор идет в результате мейоза во время цветения злака. Споры активно выбрасываются с помощью ветра, попадают на рыльце цветущего злака и прорастают. Образующийся мицелий проникает в завязь пестика и разрушает ее. На концах гиф мицелия в результате митоза отшнуровываются конидии - споры бесполого размножения, т.е. наступает конидиальное спороношение. При этом гифы гриба выделяют капельки сладкой жидкости - «медвяную росу». Насекомые переносят конидии на цветки соседних растений и заражают их.
Склероции содержат высокотоксичные алкалоиды, вызывающие отравления. Такие алкалоиды, как эрготамин, эргометрин, ши-
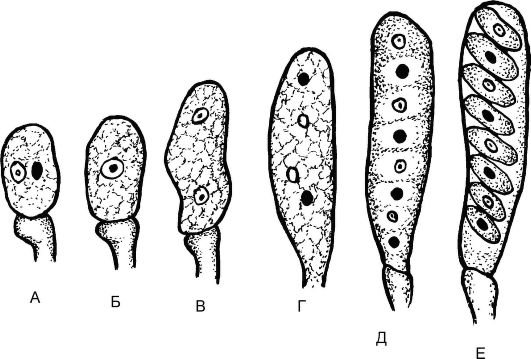
Рис. 5.19. Развитие сумки с аскоспорами: А, Б - образование зиготы на верхушке аскогенной гифы; В-Е - мейоз и развитие сумки с аскоспорами
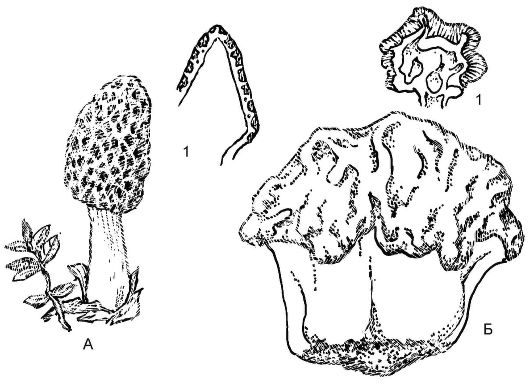
Рис. 5.20. Аскомикоты - внешний вид и плодовые тела сморчков
и строчков:
А - сморчок конический (Моrchella coinca); Б - строчок обыкновенный (Gyromitra exculenta); 1 - разрезы плодовых тел
роко применяют в современной медицине для лечения сердечнососудистых, нервных и других заболеваний. Особенно эффективны они в акушерско-гинекологической практике.
Незамкнутые плодовые тела - апотеции - встречаются у таких представителей, как сморчки (Morchella), строчки (Gyromitra). Это открытое плодовое тело обычно блюдцевидной, бокаловидной формы размером от 0,1 до 10 см, разнообразной окраски - от ярко-оранжевой или красной до коричневой и черной. Верхний слой (гимений) содержит множество сумок. Плодовые тела грибов из этой группы состоят из стерильной ножки и складчатой или лопастной шапки (рис. 5.20).
Сморчки и строчки - съедобные грибы, но при употреблении в пищу строчки предварительно необходимо прокипятить, а воду слить.
ОТДЕЛ БАЗИДИОМИКОТЫ (BASIDIMYCОTА)
В этом классе объединены почти все группы шляпочных грибов, насчитывающих около 30 тыс. видов. Вегетативное тело представлено членистым мицелием, состоящим из членистых гиф.
Размножение: вегетативное (осуществляется частями мицелия), бесполое (конидиями) и половое.
При половом процессе специальных органов полового размножения не образуется. Половой процесс протекает в форме соматогамии (рис. 5.21). Из прорастающей гаплоидной базидиоспоры развивает-
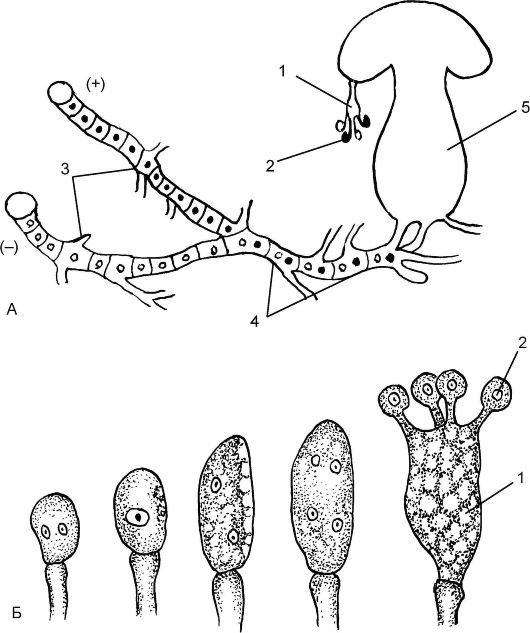
Рис. 5.21. Развитие базидиального гриба. Схема цикла развития: А - схема цикла развития: 1 - базидия; 2 - базидиоспора; 3 - первичный мицелий; 4 - дикарионный мицелий; 5 - плодовое тело из дикарионного мицелия; Б - развитие базидии с базидиальными спорами
ся первичный мицелий, который затем превращается в членистый. Каждый членик одноядерный. Вскоре происходит гологамия - слияние конечных клеток гиф. Однако слияние содержимого члеников не сопровождается слиянием ядер. Образуются дикарионы, которые затем синхронно делятся. Так формируется вторичный дикарионный мицелий.
На дикарионном мицелии образуется плодовое тело, которое состоит из пенька и шляпки. Гимениальный слой шляпки может быть пластинчатым или трубчатым. В гимениальном слое на концах дикарионных гиф из 2 ядерных клеток образуются базидии. По своему развитию базидии гомологичны сумкам. В базидии завершается половой процесс, т.е. сливаются ядра дикариона и образуется диплоидное ядро. Такая одноклеточная базидия называется холобазидией. Образовавшееся диплоидное ядро делится мейозом с образованием 4 гаплоидных ядер (см. рис. 5.19, А). К этому времени в верхней части базидии образуются четыре трубчатых выроста - стеригмы. В стеригмы перетекают образовавшиеся ядра и формируются 4 базидиоспоры: 2 условно со знаком - и 2 со знаком +. Поэтому первичные мицелии, вырастающие из них, будут гетероталличными. Базидии образуются непосредственно на гифах или в плодовых телах различной формы, но чаще состоящих из шляпки и ножки. В цикле развития чередуются 3 фазы: гаплоидная (короткая) - это базидиоспоры, дикарионная (длится основную часть жизни) - дикарионный мицелий и диплоидная (кратковременная) - молодая базидия до образования базидиоспор.
ОТДЕЛ ДЕЙТЕРОМИКОТЫ (DEUTEROMYCOTA), ИЛИ НЕСОВЕРШЕННЫЕ ГРИБЫ (FUNGI IMPERFECT!)
Дейтеромикоты, наряду с бизидиомикотами и аскомикотами, являются крупнейшей группой грибов, объединяющей 25-30 тыс. ви- дов. Эти грибы представляют собой бесполые формы (анаморфы), размножающиеся бесполым путем - конидиями. Жизненный цикл у них проходит в гаплоидной стадии без полового процесса. Вполне возможно, что дейтеромикоты - это наиболее специализированные линии эволюции грибов.
Дейтеромикоты широко распространены в природе и встречаются на различных субстратах, принимая вместе с другими грибами участие в разложении органических остатков и в почвообразовательном процессе. Многие грибы этого отдела паразитируют на высших растениях, вызывая серьезные болезни сельскохозяйственных культур. Аспергилл паразитический (Aspergillus parasiticus) выделяет
аффлатоксины, очень опасные из-за своей канцерогенности. У части представителей этих родов известны также сумчатые стадии, в связи с чем их нередко рассматривают среди аскомикот. Пенициллы и аспергиллы широко используются в микробиологической промышленности для биотехнологического производства ряда органических кислот (лимонной, фумаровой, глюконовой и др.), ферментов (протеиназ, амилаз и др.) и антибиотиков (пенициллина, фумагиллина). Антибиотики подавляют рост других организмов (особенно бакте- рий), поэтому их применяют для лечения многих заболеваний

Большое медицинское значение имеет род Пенициллиум (Penicillium). Пеницилл имеет членистый мицелий зеленоватого цвета, состоящий из одноядерных члеников. Отходящие вверх гифыконидиеносцы ветвятся на верхнем конце на стеригмы. Последние по внешнему виду напоминают кисточку или кисть руки и заканчиваются цепочкой наружных спор - конидий (рис. 5.22). Конидии - это споры бесполого размножения, образующиеся за счет митоза.
Наблюдается и половой процесс, в результате которого непосредственно на мицелии образуются закрытые шаровидные плодовые тела ярко-желтого цвета - клейстотеции. Внутри клейстотециев формируются сумки с 8 аскоспорами. Созревшие аскоспоры выходят из сумок после разрыва клейстотеция.
Пеницилл (Penicillium), сапрофит по способу питания, поселяясь на пищевых продуктах и изделиях (ткани, кожа), вызывает их порчу. Пеницилл используют не только в медицинской практике, но и в пищевой промышленности для приготовления особых сортов сыра («рокфор»).
Велико значение грибов в деятельности человека. Они участвуют в круговороте веществ в природе. Грибы, как и бактерии, минерализуют органические вещества и принимают участие в образовании гу- муса. Их используют в пищевой промышленности для производства
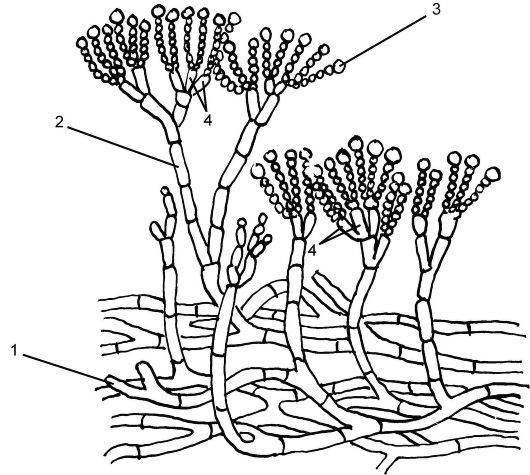
Рис. 5.22. Дейтеромикоты (Deuteromycota) пеницилл: 1 - мицелий; 2 - конидиеносец; 3 - конидии; 4 - стеригмы
спирта, вина, пива, кваса, в хлебопечении, при получении белков и витаминов. Грибы образуют органически активные вещества - антибиотики, ферменты, органические кислоты и др.
Грибы могут вызывать коррозию металлов, разрушать кожу, бумагу, ткани. Многие грибы наносят существенный вред человеку, животным и растениям, вызывая ряд заболеваний (микозы, стригущий лишай, паршу), а также приводить к порче пищевых продуктов и тем самым служить причиной различных отравлений.
ОТДЕЛ ЛИШАЙНИКИ (LICHENES)
Это группа симбиотропных растений, состоящих из 2 компонентов - автотрофных водорослей и гетеротрофных грибов. Грибная основа лишайников формируется в основном сумчатыми грибами. Водорослевый компонент состоит из видов, относимых в большинстве случаев к представителям отделов зеленые и сине-зеленые водоросли. Выделенные из лишайника водоросли не отличаются от свобод-
но живущих форм. Физиологически этот тип симбиоза основан на межклеточном обмене между водорослями и грибами. Гриб питается углеводами водоросли, а водоросли получают от грибов минеральные вещества. Однако симбиоз с грибами приводит к появлению нового биологического качества, которое выражается у лишайника в его способности размножаться как единый организм.
Вегетативное тело лишайников представлено слоевищем, имеющим различную окраску (серую, зеленоватую, буро-коричневую, желтую или почти черную). Морфологически различают 3 основных типа слоевища лишайников: накипной (корковый), листоватый и кустистый (рис. 5.23), однако существуют и переходные формы. Наиболее низкоорганизованные - накипные, или корковые, слоевища; они имеют вид порошковатых, зернистых, бугорчатых налетов, плот-
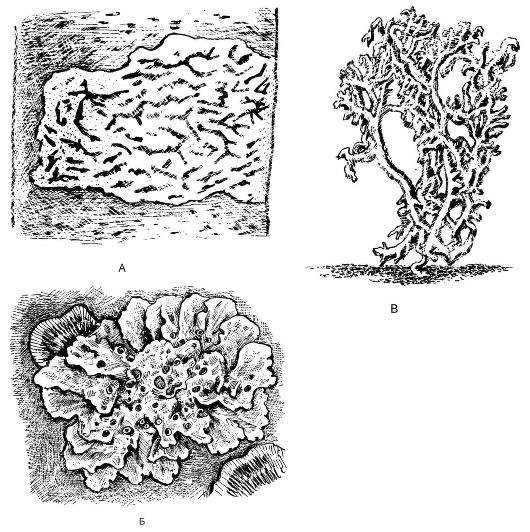
Рис. 5.23. Различные типы слоевищ лишайников: А - корковый (графис - Graphis scripta); Б - листоватый (ксантория - Xanthoria); В - кустистый (кладония - Cladonia)
но срастающихся с субстратом и не отделяющихся от него без значительных повреждений.
Более высокоорганизованные лишайники имеют листоватое слоевище в форме пластинок, чешуек или розеток, прилепленных к по- чве или деревьям с помощью ризин - аналогов ризоидов, состоящих из пучков грибных гиф.
Наивысшей организации в своем строении достигают лишайники с кустистым типом слоевища, имеющие вид разветвленного кустика (12-15 см в высоту) и срастающиеся с субстратом только основанием.
По анатомическому строению лишайники бывают гомеомерными и гетеромерными (рис. 5.24). У более примитивных - гомеомерных - по всей толще слоевища равномерно расположены грибные гифы и водоросли. При гетеромерном строении на поперечном разрезе лишайника сверху можно видеть так называемую верхнюю кору. Она образована переплетающимися и тесно смыкающимися грибными
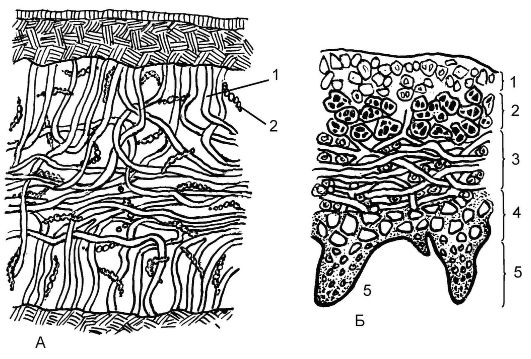
Рис. 5.24. Анатомическое строение слоевищ лишайников: А - разрез гомеомерного слоевища лишайника: 1 - гифы гриба; 2 - водорослевый компонент;
Б - разрез гетеромерного лишайника: 1 - верхний корковый слой; 2 - гонидиальный слой; 3 - средний слой с гифами гриба; 4 - нижний корковый слой; 5 - ризины
гифами. Под корой грибные гифы лежат более рыхло, и между ними находятся клетки водоросли (гонидиальный слой). Внутри слоевища можно выделить сердцевину, состоящую из рыхлых грибных гиф и больших пустот, заполненных воздухом. Под ней размещается нижняя кора, которая по строению сходна с верхней. Через нее из сердцевины проходят отдельные гифы (ризины), закрепляя лишайник в субстрате.
Большинство лишайников легко переносят высыхание. Фотосинтез и питание у них в это время прекращаются, чем и объясняется их незначительный ежегодный прирост.
Размножение лишайников преимущественно вегетативное, осно- вано на способности лишайников регенерировать из отдельных участков. Оно осуществляется путем фрагментации (отделения участков слоевища) или с помощью обособленных групп клеток водорослей, окруженных гифами гриба и различных по своей форме, - соредий, изидий и лобул (рис. 5.25). Соредии - мельчайшие образования округлой формы, включающие одну или несколько клеток водоросли и окруженные грибными гифами. Изидии - бугорчатые палочковидные
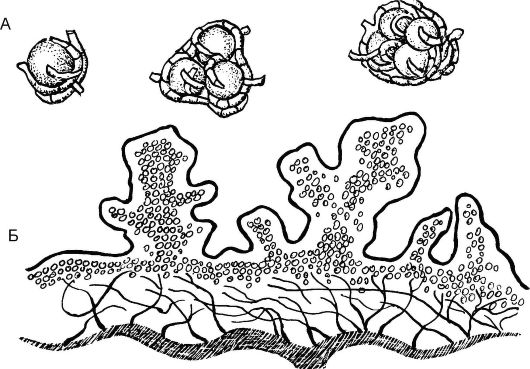
Рис. 5.25. Размножение лишайников: А - соредии; Б - изидии
выросты на верхней поверхности слоевища. Лобулы имеют вид маленьких чешуек, расположенных вертикально на поверхности слоевища или по его краям. Кроме того, наблюдается бесполое размножение с помощью спор, самостоятельно образующихся и у водорослей, и у грибов.
Половое размножение изучено недостаточно, но в общих чертах протекает так же, как у свободно живущих грибов.
Значение лишайников велико. Они разлагают и минерализуют ор- ганические вещества почвы. Являются первопроходцами - одними из первых заселяя скалы, они разрушают их поверхностный слой и, отмирая, образуют гумус, на котором расселяются другие растения. Лишайники являются индикаторами чистоты воздуха, так как не выносят даже малейших примесей сернистых газов. Из некоторых их видов получают краску и особое вещество - лакмус (для химической промышленности). В тундре и лесотундре лишайники (ягель) являются основным кормом для оленей. Встречаются съедобные лишайники также в полупустынных и пустынных районах Киргизии и Туркмении.