Ботаника: учебник. Зайчикова С.Г., Барабанов Е.И. 2013. - 288 с. : ил.
|
|
|
|
ГЛАВА IV ГЕНЕРАТИВНЫЕ ОРГАНЫ ВЫСШИХ РАСТЕНИЙ
ТЕОРИИ ПРОИСХОЖДЕНИЯ ЦВЕТКА
Древнейшие ископаемые цветки достоверно известны, начиная с середины мелового периода. Относительно происхождения обоеполого цветка существуют различные гипотезы. Наиболее распространена и хорошо обоснована стробилярная (от греч. strobilos - шишка хвой- ная) гипотеза, согласно которой цветок - это метаморфизированный укороченный спороносный побег, первоначально напоминавший обоеполую шишку вымерших голосеменных. Мегаспорофиллы в процессе метаморфоза превратились в плодолистики, а микроспорофиллы - в тычинки. Многие исследователи связывают эти видоизменения с приспособлением покрытосеменных к насекомоопылению (энтомофилии).
ЦВЕТОК
Цветок - орган семенного размножения покрытосеменных растений. Он представляет собой видоизмененный укороченный спороносный побег ограниченного роста, приспособленный для образования микро- и мегаспор, гамет и для перекрестного опыления. От
шишки голосеменных растений цветок отличается тем, что у него в результате опыления пыльца попадает на рыльце пестика, а не непосредственно на семязачаток, а при последующем половом процессе семязачатки у цветковых развиваются в семена внутри завязи.
По месторасположению цветок может быть верхушечным или боковым, т.е. выходит из пазухи прицветного листа (прицветника). Цветок состоит из стеблевой (цветоножка, цветоложе), листовой (чашелистики, лепестки) и генеративной (тычинки, пестики) частей. Цветок прикрепляется к стеблю посредством цветоножки. Если цветоножка укорочена или отсутствует, цветок называется сидячим. Верхняя расширенная часть цветоножки, к которой прикрепляются все части цветка, называется цветоложем. Форма его бывает различной: плоское цветоложе (пион), выпуклое полушаровидное (лютик, ветреница), удлиненное коническое (магнолия, горицвет, малина, земляника, гравилат), вогнутое (миндаль) (рис. 4.1.). У некоторых рас- тений в результате срастания цветоложа и нижних частей покрова и тычинок образуется гипантий. Его форма может быть блюдцевидной (смородина альпийская); шаровидной (роза морщинистая), кувшинчатой (роза коричная), бокаловидной (мушмула японская, таволга дубровколистная), воронковидной (вишня мелкоплодная). Гипантий
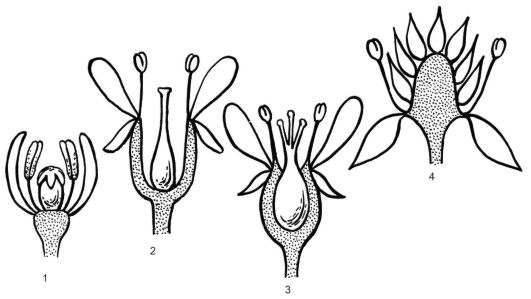
Рис. 4.1. Формы цветоложа и различное положение завязи в цветке: 1 - плоское; 2 - вогнутое; 3 - стенки завязи срастаются со стенками вогнутого цветоложа; 4 - выпуклое; 1, 2, 4 - верхнее положение завязи в цветке; 3 - нижнее положение завязи в цветке
иногда участвует в образовании плода (шиповник) и характерен для представителей семейства Розовые.
Одни части цветка считаются стерильными - это чашелистики, лепестки, а другие репродуктивными - это пестики и тычинки. Стерильные части выполняют защитную функцию и иногда могут (частично или полностью) отсутствовать, а репродуктивные приспособились к спороношению. Цветок, содержащий тычинки и пестики, называют обоеполым. Большинству покрытосеменных (более 70%) свойственны обоеполые цветки. Но цветки могут быть однополыми - содержать только тычинки или только пестик. У однодомных растений однополые цветки могут находиться на одном экземпляре (кукуруза, дуб, огурец), у двудомных (крапива двудомная, тополь, осина) тычиноч- ные или пестичные цветки - на разных экземплярах.
Околоцветник
Околоцветник - стерильная часть цветка, является его покровом, защищающим более нежные части - тычинки и пестики. Околоцветник бывает двойным, тогда он состоит из чашечки (Са - calyx) и венчика (Со - corolla), и простым (Р - perigonium), тогда он может быть чашечковидным или венчиковидным, состоящим и в том, и в другом случае из листочков простого околоцветника (рис. 4. 2).
Чашечка образует наружный круг околоцветника и состоит из совокупности чашелистиков, стоящих в формуле цветка на первом месте. Чашелистики возникли в процессе эволюции из верховых листьев эпидермы. Они покрыты эпидермой с кутином, для которой характерно наличие устьиц и трихом. Чашечка может быть раздельно- листная, состоящая из свободных чашелистиков (лютик, вишня), и сростнолистная - из сросшихся между собой на большем или меньшем протяжении чашелистиков (картофель, наперстянка, горох). В сростнолистной чашечке выделяют трубку чашечки с зубцами, лопастями и долями - в зависимости от степени срастания чашелистиков. Если любые части цветка срастаются, то в формуле цветка количество срастающихся элементов берется в скобки. Чашечка называется дву- губой, если она расчленена на 2 неравные части, каждая из которых именуется губой (шалфей, шлемник). Иногда чашечка имеет 2 круга чашелистиков (земляника, лапчатка), в этом случае наружный круг называется подчашием.
Главная функция чашечки - защита внутренних частей цветка до раскрытия бутона. Как правило, чашечка сохраняется и во вре-
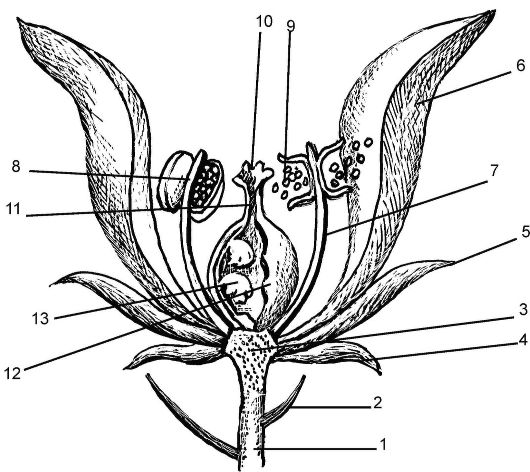
Рис. 4.2. Схема строения цветка: 1 - цветоножка; 2 - прицветник; 3 - цветоложе; 4 - подчашка; 5 - чашечка; 6 - венчик; 7 - тычиночные нити; 8 - пыльник тычинки; 9 - пыльца; 10 - рыльце пестика; 11 - столбик пестика; 12 - завязь пестика; 13 - семязачаток
мя цветения, но иногда опадает при распускании цветка (маковые). У многих растений она остается при плодах и после цветения. Например, у яснотковых служит вместилищем для дробного плода, а у астровых превращена в хохолок (паппус), способствующий переносу плодов ветром. Чашечка, как правило, зеленого цвета, но иногда ярко окрашена и выполняет роль венчика, который в этом случае нередко редуцирован до нектарников (живокость, аконит, морозник). В некоторых случаях чашечка слабо развита (например, в семействах Зонтичные, Сложноцветные, Валериановые).
Венчик, состоящий из совокупности лепестков, образует следующий круг в цветке за чашечкой и в формуле цветка обозначается Со. Венчик может быть свободнолепестным и спайнолепестным. Обычно он ярко окрашен, что привлекает насекомых-опылителей. Именно венчик создает облик цветка. В процессе эволюции лепестки
произошли из потерявших пыльники тычинок. У лепестков могут развиваться различные трихомы, простые и железистые волоски, чешуйки и др.
Простой околоцветник (Perigonium) состоит из совокупности однородных листочков. Околоцветник, состоящий из зеленых листочков, называют чашечковидным (семейство Злаковые), из окрашенных в любой другой цвет - венчиковидным (тюльпан). Он может быть раздельнолистным (тюльпан, лук, ветреница) и сростнолист- ным (ландыш, купена). Околоцветник может быть редуцирован и представлен в виде щетинок (камыш) или волосков (пушица), а может отсутствовать (ива, тополь). Цветок, лишенный покрова, называется беспокровным, или голым. Редукция околоцветника, как полагают, связана с приспособлением к ветроопылению. Для насекомоопыляемых растений характерно наличие нектарников, содержащих главным образом сахара; они разнообразны по происхождению и форме.
У большинства растений части цветка образуют хорошо заметные круги (циклы). Наиболее распространены 5- и 4-, т.е. пента- и тетрациклические цветки. Число частей цветка в каждом круге может быть различным. В зависимости от этого цветки бывают: 5-членные (пятимерные) - у большинства двудольных; реже - 2- или 4-членные (маковые, капустные); 3-членные (трехмерные) - у большинства однодольных. Иногда число кругов и членов в них увеличивается (у садовых форм). У большинства покрытосеменных все части цветка расположены на цветоложе в виде концентрических кругов (цветок круговой, циклический). В других случаях (магнолия, купальница, ветреница) они расположены по спирали (цветок спиральный, ациклический). Иногда одни части цветка расположены в кругах, другие - по спирали (цветок полукруговой, гемициклический). В последних околоцветник имеет циклическое, а тычинки и пестики - спираль- ное расположение (лютик) или чашечка - спиральное, а остальные части цветка - циклическое (шиповник).
По особенностям симметрии цветки делят на актиноморфные, или правильные, через которые можно провести много плоскостей симме- трии (зонтичные, капустные); их обозначают в формуле цветка как *; и зигоморфные (бобовые, злаковые), через которые можно провести только одну вертикальную плоскость симметрии (↑) (рис. 4.3). Если через цветок нельзя провести ни одной плоскости симметрии, его называют несимметричным, или асимметричным (валериана лекарственная, канновые).
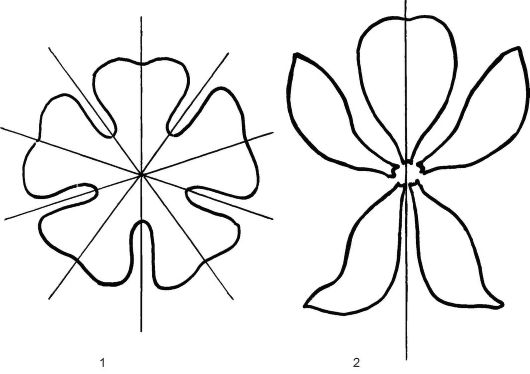
Рис. 4.3. Актиноморфные (1) и зигоморфные (2) цветки
Андроцей (Androeceum)
Совокупность тычинок одного цветка называют андроцеем (А). Они обычно располагаются в 1-2 круга. Как правило, число тычи- нок постоянно для всего вида. Тычинки могут быть свободными или сросшимися. Различают разные типы андроцея: однобратственный - все тычинки в цветке срастаются в одну группу (люпин, камелия); двубратственный - тычинки срастаются в 2 группы (у многих бобовых 9 тычинок срастаются, а 1 остается свободной); многобратственный - многочисленные тычинки цветка срастаются в несколько групп (зверобой, магнолия); братственный - тычинки остаются несросшимися. По длине тычинки бывают равные, если они одной длины (тюльпан), неравные - разной длины (водосбор олимпийский), двусильные, если из 4 тычинок 2 длинные, а 2 короткие (яснотковые); трехсильные, если из 6 тычинок 3 более длинные (нарцисс гибридный); четырехсильные, если из 6 тычинок 4 более длин ные (капустные).
В некоторых цветках андроцей вообще отсутствует - это так называемые женские однополые цветки, а в однополых мужских цветках есть только тычинки. Каждая тычинка состоит из суженной части - тычиночной нити, посредством которой она нижним концом прикреплена к цветоложу, и расширенной части - пыльни-
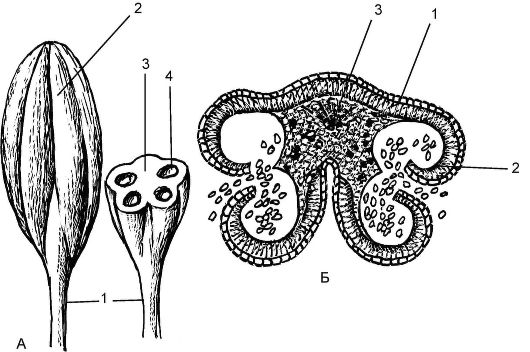
Рис. 4.4. Тычинка и строение пыльника: А - тычинка: 1 - нить тычинки; 2 - пыльник; 3 - связник; 4 - гнезда пыльника; Б - вскрывающийся пыльник: 1 - эпидерма; 2 - фиброзный слой; 3 - проводящий пучок связника
ка (рис. 4.4). Пыльник покрыт эпидермой, под которой располагается слой клеток - эндотеций (фиброгзный слой), способствующий вскрытию гнезд пыльника. Он состоит из 2 половинок, соединенных связником, т.е. продолжением тычиночной нити. Каждая половинка несет 2 пыльцевидных гнезда, называемые пыльцевыми мешками (микроспрорангиями). Гнезда пыльников называют пыльцевыми мешками. В зрелом пыльнике перегородки между гнездами часто не выявляются. Самый внутренний слой, выстилающий полость гнезда, называется тапетумом. Содержимое клеток тапетума идет на питание микроспор. Многие ученые считают, что тычинки эволюционно представляют собой редуцированные микроспорофиллы вымерших голосеменных растений.
Гинецей (Gynoeceum)
Совокупность плодолистиков одного цветка, образующих один или несколько пестиков, называют гинецеем (от греч. gyne - женщина).
Плодолистики представляют собой мегаспорофиллы, несущие семязачатки.
Строение пестика. Пестик возникает из плодолистика или плодолистиков вследствие срастания их краев и представляет собой элемент генеративной части цветка, несущий один или несколько семязачатков. С современной точки зрения плодолистики произошли из мегаспорофиллов древних голосеменных. В целом пестик по конструкции представляет собой новый орган, которого не было у голосеменных растений. Каждый пестик состоит из рыльца, столбика и разросшейся завязи. Пестик может быть простым, состоящим из одного плодолистика, и сложным - из нескольких сросшихся плодолистиков (рис. 4.5).
Рыльце - расширенная часть на верхушке столбика, предназначенная для принятия пыльцы. На поверхности рыльца имеются различные приспособления для восприятия пыльцы: выросты в виде сосочков, короткие, густо переплетенные волоски. На рыльце и внутри пестика вырабатывается железистая ткань, которая стимулирует прорастание пыльцы. Она облегчает передвижение пыльцевых трубок через столбик и снабжает их питательными веществами, необходимыми для развития.
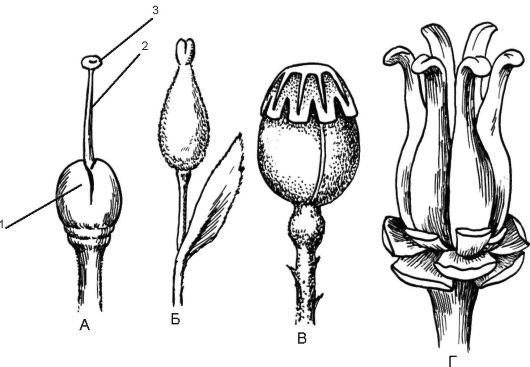
Рис. 4.5. Гинецей:
А-В - ценокарпий (А - махорка, Б - ива, В - мак); Г - апокарпий (сусак); 1 - завязь; 2 - столбик; 3 - рыльце
Столбик - цилиндрическая часть пестика, соединяющая рыльце и верхушку завязи. У многих растений столбик неразвит, а рыльце, находящееся на завязи, называют сидячим (мак). Завязь - самая важная часть пестика, поэтому число пестиков в цветке определяется числом завязей, а не столбиков. О числе плодолистиков, образующих пестик, определяют (но не всегда безошибочно) по несросшимся столбикам, рыльцам или лопастям рыльца. В случае их полного срастания число плодолистиков определяют по количеству средних жилок в стенке завязи. Завязь пестика выполняет функцию влажной камеры, предохраняющей семязачатки от высыхания, поедания их насекомыми и от резких колебаний температуры. Этим покрытосеменные существенно отличаются от голосеменных, у которых семязачатки лежат открыто на мегаспорофиллах. Место прикрепления семязачатков в завязи называется плацентой. Семязачатки сообщаются с плацентой с помощью семяножки. Плацента обычно имеет вид вздутия, выроста или выступа, образованного тканями завязи. В зависимости от формы цветоложа различают положение завязи в цветке - нижнее и верхнее (рис. 4.6). Верхняя завязь располагается на цветоложе любой формы (плоское, вогну- тое, выпуклое) свободно, не срастаясь с другими частями цветка
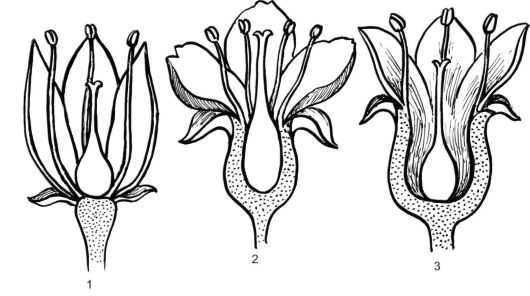
Рис. 4.6. Типы завязи в зависимости от ее положения относительно места прикрепления других частей цветка: 1 - верхняя; 2 - нижняя; 3 - верхняя, окруженная стенками гипантия
(тюльпан, вишня, малина). Нижняя завязь формируется за счет срастания с другими частями цветка так, что ее нельзя выделить, не нарушая целостности цветка (яблоня, рябина, тыквенные). Полунижняя завязь в отличие от нижней частично срастается с цветоложем.
Типы гинецея (G). Структурная единица гинецея - плодолистик. Если плодолистик образует единственный пестик в цветке, то такой гинецей называется монокарпным (бобовые). Края единственного плодолистика свертываются и срастаются, образуя на месте сращения так называемый брюшной шов. На стороне, противоположной брюшному шву, находится средняя жилка плодолистика, образующая спинной шов. В результате формируется одногнездная завязь с семязачатками. Если 2 плодолистика и более образуют множество самостоятельных простых пестиков, такой гинецей называется апокарпным (земляника, малина). Гинецей, формирующийся из нескольких сросшихся плодолистиков, образующих единый пестик, называется ценокарпным (яблоня, мак, помидор).

При простом околоцветнике знаки чашечки и венчика не применяются, и, например, формула цветка тюльпана (простого околоцветника) -*Р3+3А3+3G(3).
Для пространственного изображения всех частей цветка составляют схематическую проекцию цветка на плоскости, называемую диа- граммой.
Принят единый способ ориентации диаграммы: ось соцветия вверху, а кроющий лист внизу. На диаграмме части цветка имеют условные обозначения (рис. 4.7). Части околоцветника на диаграмме обозначаются дугами: чашелистики - с выступом на середине дуги; лепестки - без выступа. Тычинки обозначаются в форме поперечного разреза пыльника, гинецей - в виде поперечного разреза завязи. В случае срастания между собой отдельных членов цветка фигуры, обозначающие их на диаграмме, соединяются дугами.
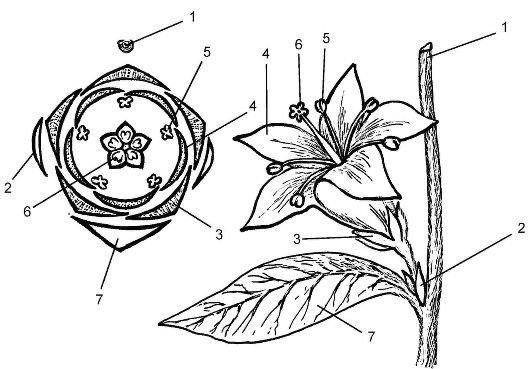
Рис. 4.7. Построение диаграммы цветка: 1 - ось соцветия; 2 - прицветник; 3 - чашелистик; 4 - лепесток; 5 - тычинка; 6 - гинецей; 7 - кроющий лист
СЕМЯЗАЧАТОК
Семязачатки - это небольшие образования, располагающиеся в завязи пестика и развивающиеся в семя. Семязачаток формируется
из меристематического бугорка, возникающего на плодолистике. Место прикрепления семязачатка к плодолистику называется пла- центой. Число семязачатков может колебаться от одного (пшеница, вишня, подсолнечник) до нескольких тысяч (мак) и миллионов (у орхидных).
Развившийся семязачаток имеет семяножку, посредством которой он прикрепляется к плаценте; центральную часть - нуцеллус (гомолог мегаспорангия); 1 или 2 интегумента (покрова) семязачатка, которые на верхушке не срастаются и образуют микропиле, или пыльцевход; противоположную микропиле базальную часть семязачатка - халазу, где нуцеллус и интегументы сливаются. В семязачатке происходят следующие процессы: мегаспорогенез - формирование мегаспор; мегагаметогенез - формирование женского гаметофита и оплодотворение.
Микроспорогенез и микрогаметогенез
Микроспорангиями являются гнезда пыльников. Процесс образования микроспор в микроспорангиях называется микроспорогене- зом. Клетки спорогенной ткани, в гнездах пыльников, в результате митотических делений формируют материнские клетки микроспор (микроспороциты). Из каждого диплоидного микроспороцита в результате мейоза образуются 4 гаплоидные микроспоры. Сформировавшаяся микроспора имеет оболочку и единственное гаплоидное ядро. Такая микроспора дает начало пыльцевому зерну. Процесс образования из микроспоры мужского гаметофита - пыльцы - называется микрогаметогенезом.
Микрогаметогенез. При делении митозом из микроспоры формируется пыльца, состоящая из 2 новых клеток и покрытая оболочкой - спородермой. Маленькая клетка - генеративная (спермагенная), а более крупная - вегетативная (клетка пыльцевой трубки). Таким образом, пыльца, или мужской гаметофит, формируется из микроспоры внутри пыльника.
Спородерма пыльцы имеет 2 слоя - экзину и интину. Наружная слоистая экзина пропитана очень стойкими углеводами спорополленинами и имеет на поверхности различные выросты. Внутренняя оболочка - интина. Она менее стойка, чем экзина, состоит из целлюлозы и пектина и сохраняет эластичность (рис. 4.8). Особенности строения спородермы и пыльцы являются видоспецифичными. После созревания пыльцы пыльники лопаются, и пыльца рассеивается.
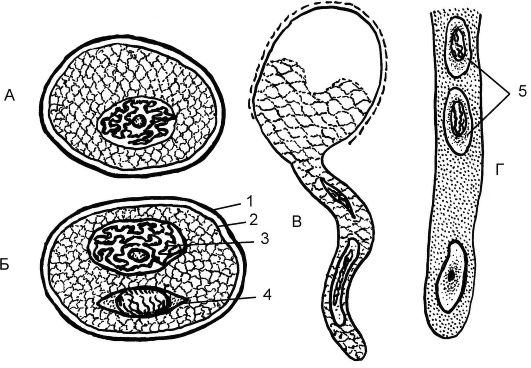
Рис. 4.8. Микроспора и ее прорастание: А - микроспора; Б - пыльцевое зерно; В - формирование пыльцевой трубки; Г - часть пыльцевой трубки; 1 - экзина; 2 - интина; 3 - вегетативная клетка; 4 - репродуктивная клетка; 5 - спермии
Жизнеспособность пыльцы может сохраняться от нескольких часов (у ржи - 12 ч) до нескольких лет (у яблони 4,5 года).
Мегаспорогенез и мегагаметогенез
Центральной частью семязачатка является нуцеллус, который гомологичен мегаспорангию. У цветковых растений в нуцеллусе, вблизи верхушки (в области микропиле), из клеток археспория обособляется одна материнская клетка (мегаспора), имеющая, как и все растение, диплоидный набор хромосом. После мейотического деления этой клетки образуются 4 гаплоидные мегаспоры, располагающиеся, как правило, линейно - от микропиле к халазе. Таким образом, происходит мегаспорогенез.
После этого начинается мегагаметогенез, т.е. формирование женского гаметофита - зародышевого мешка. Из 4 образовавшихся мегаспор 3 дегенерируют и рассасываются, а 1 мегаспора, обращен- ная ближе к халазе, начинает делиться, и из нее развивается женский
гаметофит - зародышевый мешок. При этом ядро мегаспоры претерпевает 3 последовательных митотических деления (эндомитоз), в результате чего образуется 8 моноплоидных ядер. После 1-го деления мегаспоры образуются 2 ядра, которые расходятся к полюсам удлиняющейся мегаспоры. Далее каждое из этих ядер синхронно делится еще 2 раза; в результате у каждого полюса возникают по 4 ядра. На этой 8-ядерной стадии процесс деления ядра женского гаметофита заканчивается. При этом мегаспора разрастается, оттесняя ткань ну- целлуса к интегументам. От каждой из 2 четверок ядер в центральную часть зародышевого мешка отходит по 1 ядру. Эти так называемые полярные ядра сближаются в центральной части и, сливаясь, образуют диплоидное вторичное ядро. В дальнейшем 7 ядер зародышевого мешка претерпевают фазу цитоплазматического деления, т.е. каждое ядро с участком цитоплазмы окружается плазматической мембраной; следовательно, образуется 7 отдельных клеток (рис. 4.9, 4.10).
У микропилярного полюса гаметофита три клетки формируют яйцевой аппарат, одна из которых, наиболее крупная, называется яйце-
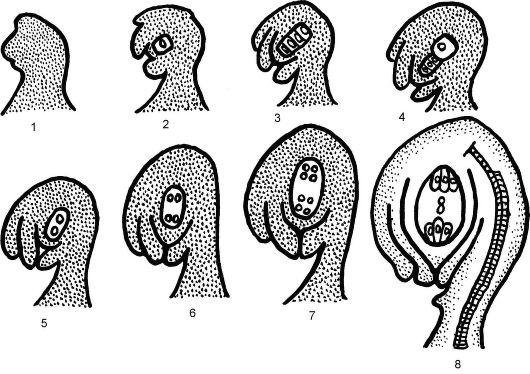
Рис. 4.9. Схема формирования зародышевого мешка: 1-4 - развитие нуцеллуса, обособление и мейоз клетки археспория, отмирание 3 мегаспор; 5-8 - развитие женского гаметофита - зародышевого мешка из оставшейся мегаспоры; 9 - 8-ядерный зародышевый мешок
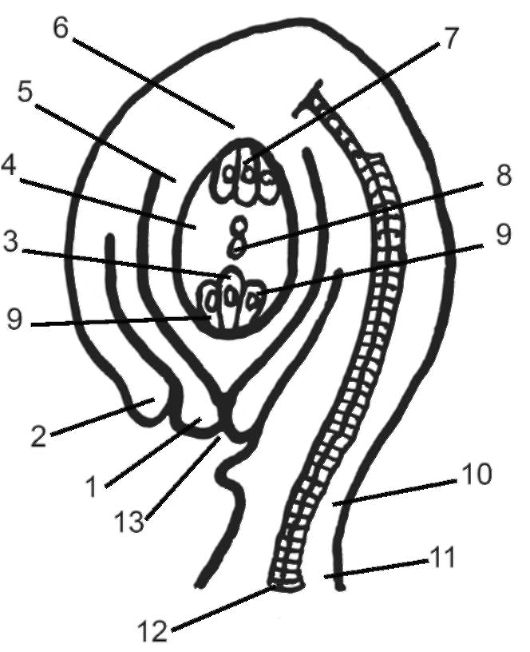
Рис. 4.10. Строение семязачатка: 1 - внутренний; 2 - наружный интегументы; 3 - яйцеклетка; 4 - зародышевый мешок; 5 - нуцеллус; 6 - халаза; 7 - антиподы; 8 - два полярных ядра; 9 - синергиды; 10 - фуникулюс; 11 - пла- цента; 12 - проводящий пучок; 13 - пыльцевход (микропиле)
клеткой, а две другие - вспомогательными клетками-синергидами. 3 клетки халазального полюса получили название антипод. В централь- ной части зародышевого мешка располагается клетка с диплоидным ядром. Так возникает женский гаметофит, который сильно редуцирован даже по сравнению с гаметофитом голосеменных и полностью лишен архегониев.
Опыление
Для образования зародыша должны произойти опыление и оплодотворение. Опыление - это процесс переноса пыльцы с тычинки на рыльце пестика. Опыление впервые появляется у голосеменных растений, но наибольшего совершенства оно достигает у покрытосеменных.
Различают 2 типа опыления: самоопыление и перекрестное опыление. При самоопылении на пестик цветка попадает пыльца того же растения; если перенос пыльцы осуществляется между цветками разных особей, происходит перекрестное опыление. Считается, что оно свойственно 90% растений. Перекрестное опыление обусловливает высокий уровень гетерозиготности популяций. Это создает большие возможности для естественного отбора. Строгое самоопыление встречается относительно редко (например, у гороха) и может вести к расщеплению вида на ряд чистых линий, т.е. делает популяции гомозиготными.
Для эволюционного процесса оптимальным считается сочетание самоопыления и перекрестного опыления, что чаще и происходит в природе. Одна из форм, ограничивающих самоопыление, - дву- домность, т.е. на одних растениях развиваются только мужские (тычиночные) цветки, а на других - женские (пестичные), имеющие только гинецей. Однодомные растения имеют цветки, содержащие и андроцей, и гинецей. Другая форма, ограничивающая самоопыление, - полная и физиологическая несовместимость. Она выражается в подавлении при самоопылении прорастания пыльцы на рыльце пестика той же особи.
Выделяют 2 типа перекрестного опыления - биотическое и абиотическое. Биотическое опыление осуществляется животными, абиотическое - с помощью неживых факторов внешней среды (ветер, вода у водных растений).
Двойное оплодотворение
Пыльца, попав на рыльце пестика, начинает прорастать. Из вегетативной клетки образуется пыльцевая трубка, а из генеративной - 2 спермия. Пыльцевая трубка проникает в зародышевый мешок и, достигнув яйцеклетки, лопается, что обеспечивает проникновение в него спермиев. Один спермий копулирует с яйцеклеткой, образуя зиготу, дающую начало зародышу. Второй спермий сливается с вторичным диплоидным ядром, располагающимся в центре зародышевого мешка, что приводит к образованию триплоидного ядра. В результате формируется триплоидная клетка, развивающаяся в специальную питательную ткань - эндосперм (3n) (от греч. endon - внутри и sperma - семя). Таким образом, происходит двойное оплодотворение, свойственное только покрытосеменным растениям. Оно впервые описано в
Биологический смысл двойного оплодотворения состоит в том, что триплоидный эндосперм развивается лишь в случае оплодотворе- ния, чем достигается существенная экономия энергетических и пластических ресурсов в отличие от голосеменных растений, у которых образование эндосперма не связано с оплодотворением.
У покрытосеменных растений эндосперм называется «вторичным», или «белком». Только у покрытосеменных растений зародыш (спорофаза) начинает свое развитие самостоятельно за счет трипло-
идной фазы. У всех предшествующих групп (голосеменные и др.) зародыш развивается за счет гаметофазы.
Таким образом, из составных частей цветка образуются:
• из оплодотворенной яйцеклетки - зародыш (2n);
• из диплоидного ядра - эндосперм (3n);
• из интегументов семязачатка - семенная кожура семени (2n);
• из нуцеллуса - перисперм семени (2n);
• из стенок завязи, часто с участием других элементов цветка (чашечки, цветоложа), - стенка плода (перикарпий). Перикарпий имеет 3 слоя: наружный - экзокарпий, средний - мезокарпий и внутренний - эндокарпий.
У многих цветковых (около 10% видов) в процессе эволюции половое размножение замещается различными формами бесполого. Из них наиболее известен апомиксис, при котором семена у апомиктичных растений образуются без оплодотворения из половой (партено- генез) или вегетативной (аногамия) клетки. При этом не происходит обмена генетической информацией, и поэтому все апомиктические особи имеют одинаковую генетическую и соматическую конституцию. Ярким примером растения с апомиктическим образованием семян по типу партеногенеза является одуванчик, отличающийся высокой жизнеспособностью. Нередко, особенно у многолетних корневищных растений, преобладает вегетативное размножение, а семенное - подавлено.
Соцветия
Соцветия - это побег или система специализированных побегов, несущих цветки. Соцветия свойственны громадному большинству цветковых растений. Соцветие имеет главную ось (ось соцветия) и боковые. Последние могут ветвиться или быть неразветвленными и нести цветки. На осях соцветий имеются узлы и междоузлия. На узлах соцветия располагаются прицветники, а на узлах цветоножки - прицветнички.
Биологический смысл возникновения соцветия - в возрастающей вероятности опыления цветков. Несомненно, что соцветия более за- метны среди зелени листьев, нежели единичные цветки, и насекомое за единицу времени посетит гораздо больше цветков, если они собраны в соцветия. Обычно соцветия группируются близ верхней части растения на концах ветвей.
Классификация соцветий достаточно сложна, но в справочниках и определителях по систематике растений учитывают 2 признака: характер ветвления и способ нарастания.
В зависимости от степени разветвления осей соцветия делят на простые и сложные. У простых соцветий на главной оси располагаются одиночные цветки (черемуха, подорожник, подсолнечник); у сложных - не одиночные цветки, а боковые оси (донник лекарственный, сирень, пшеница, мятлик). У одних растений апикальные меристемы расходуются на формирование верхушечного цветка, и в этом случае соцветия относят к закрытым, или определенным. У закрытых соцветий (симподиальных) верхушечные цветки обычно раскрываются раньше нижележащих боковых, поэтому их называют верхо цветными. У других растений апикальные меристемы остаются в вегетативном состоянии, и такие соцветия называют открытыми, или неопределен ными. У открытых (моноподиальных) соцветий цветки распускаются последовательно снизу вверх, поэтому их называют бокоцветными.
Существует несколько типов ботрических (от греч. botris - кисть) соцветий (рис. 4.11).
Простые ботрические соцветия - это моноподиальные соцветия, характеризующиеся длиной и формой главной оси, а также наличием или отсутствием цветоножек и прицветников.
Кисть (основной вариант простых соцветий) - ось тонкая, с цветками на цветоножках более или менее одинаковой длины, которые поочередно прикрепляются к главней оси соцветия. Кисти бывают односторонние (ландыш) и двусторонние (черемуха).
Щиток - соцветие, сходное с кистью, но у щитка нижние цвето- ножки длиннее верхних, поэтому его цветки расположены на одном уровне и в одной плоскости (рябина, боярышник).
Зонтик - производное кисти с сильно укороченной тонкой осью и цветками на цветоножках одинаковой длины (вишня, чистотел, примула).
Головка - видоизмененный зонтик, у которого главная ось раз- растается в виде головки, а на ней располагаются цветки на укороченных цветоножках (клевер, люцерна).
Колос - производное кисти, отличающееся от нее отсутствием цветоножек (подорожник).
Сережка - повислый колос, т.е. колос с мягкой осью, несущий однополые цветки; после цветения соцветие обычно целиком опадает (ива, тополь).
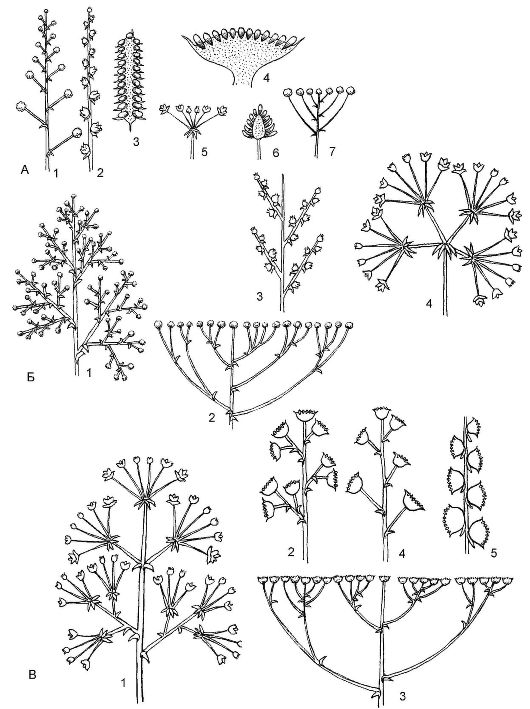
Рис. 4.11. Типы ботрических соцветий: А - простые: 1 - кисть; 2 - колос; 3 - початок; 4 - корзинка; 5 - простой зонтик; 6 - головка; 7 - щиток;
Б - сложные: 1 - метелка; 2 - сложный щиток; 3 - двойной колос; 4 - сложный зонтик;
В - типы агрегатных соцветий: 1 - метелка зонтиков; 2 - метелка корзинок; 3 - щиток корзинок; 4 - кисть корзинок; 5 - колос корзинок
Початок - разновидность колоса, но с толстой мясистой осью (кукуруза). Часто початок окружен листом разной формы и окраски, который называют покрывалом или крылом.
Корзинка - главная ось соцветия разрастается в виде чаши или ложа, на котором располагаются сидячие цветки, окруженные со всех сторон (часто в 2 ряда) видоизмененными присоцветными листьями - листовой оберткой (подсолнечник, ромашка). Поскольку корзинка - соцветие моноподиальное, распускание цветков в ней идет центростремительно - от периферии к центру.
Сложные ботрические соцветия могут быть как открытыми, так и закрытыми.
Сложная кисть (метелка) - соцветие, у которого на удлиненной моноподиальной главной оси располагаются ботрические парциальные (или частные) соцветия, являющиеся простыми кистями. В зависимости от степени ветвления кисть может быть двойная - соцветие, у которого на главной оси располагаются простые кисти (донник, вероника простертая); тройная - имеющая оси 3-го порядка (хрен, алоэ крупноплодное). К сложным метелкам относятся составные (агрегатные) соцветия. Они характеризуются тем, что тип нарастания главной оси отличается от такового у парциальных соцветий - например, метелка корзинок (полынь обыкновенная), кисть зонтиков (плющ обыкновенный), колос корзинок (сушеница лесная).
Сложный колос - соцветие, у которого на удлиненной главной оси располагаются простые колосья, называемые колосками (пшеница, рожь, ячмень); морфологически близко к двойной кисти.
Сложный зонтик - соцветие с укороченной главной осью, на которой расположена розетка верховых листьев (прицветников), называемая общей оберткой. Из пазух листьев выходят оси, завершающиеся парциальными соцветиями - простыми зонтиками, которые называют зонтичками. Цветки в зонтичках часто имеют выраженные прицветники, составляющие оберточки. Сложные зонтики свойственны многим растениям семейства Сельдерейные (морковь, пе- трушка, укроп, сельдерей). Это открытые соцветия, поэтому цветки в них распускаются центростремительно.
Цимозные (симподиальные соцветия) (от греч. kyma - волна) получили свое название по особому способу зацветания. Их соцветия закрытые, главная ось у них не выражена, а парциальные соцветия имеют симподиальное ветвление. Цветки возникают в пазухах прицветных листьев на осях предыдущих порядков, т.е. распускание цветков происходит центробежно. В зависимости от числа боковых
осей цимоиды разделяют на три основных типа: монохазий, дихазий, плейохазий (рис. 4.12).
Монохазий (однолучевик) - такое цимозное соцветие, у которого каждая материнская ось несет 1 дочернюю, формирующуюся под цветком, завершающим ось предыдущего порядка. В зависимости от направления осей различают 2 типа монохазиев: завиток и извилина.
Завиток (улитка) - от оси 1-го порядка отходит ось 2-го порядка, от нее - 3-го порядка и т.д., но при этом оси отходят только в одном направлении, как бы закручиваясь в улитку или завиток (незабудка и другие представители семейства Бурачниковые).
Извилина - от оси первого порядка отходит ось второго порядка, от второго - третьего и т.д., то в одну, то в другую сторону (лютик едкий).
Дихазий (двулучевик) - соцветие, у которого под верхушечным цветком главной оси развиваются 2 парциальных соцветия, при этом
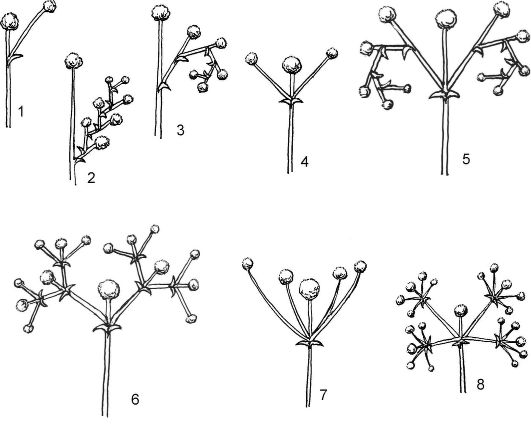
Рис. 4.12. Типы цимоидных соцветий: монохазии: 1 - «элементарный» монохазий; 2 - извилина; 3 - завиток; дихазии: 4 - простой; 5 - двойной, или двойной завиток; 6 - тройной; плейохазии: 7 - простой; 8 - двойной
каждая ось предыдущего порядка несет 2 оси следующего порядка. К дихазиям относятся развилина и двойной завиток.
Развилина - под цветком главной оси образуются супротивно рас- положенные ветви (оси), заканчивающиеся цветками. В дальнейшем каждая из этих осей также образует 2 супротивно расположенные оси следующего порядка (гвоздика).
Двойной завиток - под цветком главной оси супротивно образуются 2 простых завитка (картофель).
Плейохазий (многолучевик) - соцветие, у которого каждую ось предыдущего порядка сменяют более 2 осей следующего порядка (молочай, родиола).
Цветение. Однолетние растения зацветают не ранее чем через 20- 30 дней после посева. Все однолетние (рожь, пшеница) и двулетние (редька, капуста, морковь) растения цветут и плодоносят один раз в жизни, после чего погибают, поэтому их называют монокарпическими (или монокарпиками). Многолетние травы зацветают на 2-й год жизни, а многие деревья - через десятки лет. У многолетних растений, как правило, цветение и плодоношение многократно повторяются в течение жизни, поэтому их называют поликарпическими (или поли- карпиками). Лишь немногие многолетние растения являются монокарпиками, т.е. после первого цветения и плодоношения отмирают (некоторые агавы, бамбуки, пальмы, ферулы).
ПЛОД
Плод - орган размножения покрытосеменных растений, раз- вивающийся из одного цветка и заключающий в себе семена. Плод (или соплодия) у покрытосеменных растений развивается из завязи после цветения. Из стенки завязи формируется стенка плода - околоплодник (перикарп, или перикарпий: от греч. peri - около, carpos - плод). В нем выделяют 3 зоны: наружную (экзокарпий), среднюю (мезокарпий) и внутреннюю (эндокарпий). Все 3 зоны хорошо различимы, например, в плоде персика: тонкий кожистый наружный слой - экзокарпий, съедобная сочная мякоть плода - мезокарпий, твердая косточка из каменистой ткани, окружающая единственное семя, - эндокарпий (рис. 4.13). Нередко эти зоны околоплодника различаются слабо. Стенка плода может быть сухой и сочной (мясистой).
Функции плода: формирование, защита и распространение семян. Максимальное число семян в плоде равно числу семязачатков в за-
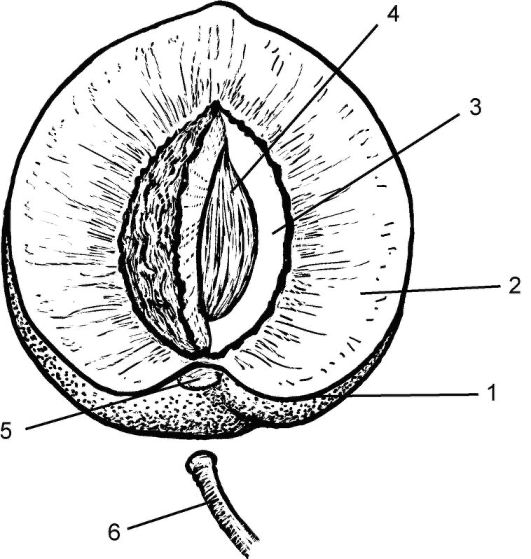
Рис. 4.13. Строение плода (однокостянки) персика обыкновенного (Persica vulgaris): 1-3 - околоплодник, или перикарпий (1 - акзокарпий, 2 - мезокарпий, 3 - эндокарпий); 4 - семя; 5 - след плодоножки; 6 - плодоножка
вязи, но обычно оно меньше, так как не все семязачатки достигают зрелости. Плоды в зависимости от структуры стенки делят на 2 группы: сухие и сочные. Сухие плоды бывают вскрывающимися или невскрывающимися - в зависимости от того, вскрывается (растрескивается) стенка плода после созревания или нет. Эволюционно сочные плоды появились как производные сухих. Стенка сочного плода может развиваться из стенки завязи (перикарпа) либо из стенки завязи, сросшейся с цветочной трубкой или вогнутым цветоложем.
Соплодия - это образования, возникшие путем срастания завязей отдельных цветков одного соцветия. Например, у ананаса ось соцветия срастается с многочисленными завязями и основаниями прицветников в мясистую сочную ткань. У шелковицы «тутовая ягода» образована сближенными, но не срастающимися друг с другом семянками, которые заключены в сочные, съедобные, окрашенные околоцветники. У инжира (винной ягоды) плоды семянки также не срастаются и находятся в полом вместилище, мясистая стенка которого образована осями соцветия. У каштана настоящего соплодие формируется из нижней синкарпной завязи женского цветка (G(6-9)), и состоит из 2-3 желудей луковицеобразной формы.
Морфогенетическая классификация плодов основана на типе гинецея. По этому признаку их делят на 4 главных типа. Апокарпии
образуются из цветков с апокарпным гинецеем; из каждого свободного пестика одного цветка формируется отдельный плодик. Моно- карпии возникают из цветков, имеющих монокарпный гинецей. Они генетически родственны апокарпиям и образовались в результате редукции плодолистиков до 1. Ценокарпии (синкарпий, лизикарпий и паракарпий) формируются из цветков с ценокарпным гинецеем. Псевдомонокарпии внешне похожи на монокарпии, но образуются из гинецея, в котором первоначально закладываются 2 или более плодо- листиков, но потом чаще развивается только 1; в результате возникает одногнездная завязь с 1 семязачатком.
Плоды монокарпии возникают из цветков, имеющих монокарпный гинецей, образованный одним плодолистиком, и всегда - верхнюю завязь (рис. 4.14).
Однолистовка - одногнездный многосеменной плод, вскрывающийся одной щелью только по брюшному шву, вдоль которого прикрепляются семена (живокость полевая).
Боб - одно-, дву-, многосеменный монокарпий, раскрывающий- ся по 2 швам - спинному и брюшному: от вершины к основанию. Семена располагаются непосредственно на створках вдоль брюшного шва (донник, арахис, горошек, акация). Бобы могут быть спирально закрученные (люцерна посевная), членистые (софора, вязель), односеменные невскрывающиеся (донник, люцерна, клевер) и даже сочные невскрывающиеся (софора японская).
Однокостянка - односеменный невскрывающийся монокарпий, эндокарпий которого (косточка) - твердый, склерефицированный. Мезокарпий может быть сочным (как в плодах вишни, черемухи, сливы, абрикоса) или сухим, кожистым и несъедобным (как в плодах миндаля, облепихи). У миндаля съедобно только семя, в 2 крупных семядолях которого находятся запасные вещества. Несмотря на то что плод у облепихи сочный, это по существу сухая костянка, так как экзо- и мезокарп - пленчатые и сросшиеся; эндокарп - кожистый, толстый, темно-коричневый. Сухую костянку окружают сочные ткани разросшейся трубки чашечковидного околоцветника, формирующие мякоть плода.
Плоды апокарпии - наиболее архаичные плоды, возникающие из цветков с верхней завязью, имеющие гинецей (G) со свободными плодолистиками (от 2 до ∞) (см. рис. 4.14).
Многолистовка - многосеменной сборный плод, состоящий из простых листовок (2 и более), каждая из которых возникает из отдельного пестика апокарпного гинецея, например у лютиковых (купальница, калужница болотная).
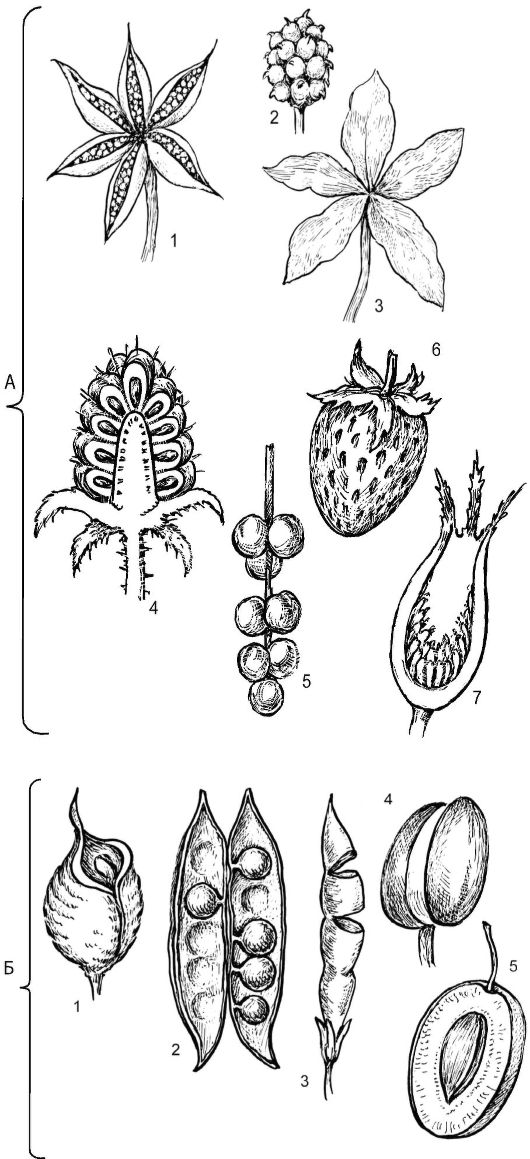
Рис. 4.14. Типы апокарпных и монокарпных плодов:
А - сухие и сочные апокарпии: 1, 3 - многолистовка (многие лютиковые и пион); 2 - многоорешек (некоторые лютиковые); 4 - многокостянка (розоцветные из рода Rubus); 5 - сочная многолистовка, отдельные плодики сидят на удлиненном цветоложе (лимонник); 6 - земляни- чина, особый тип сочного многоорешка с разросшимся цветоложем (земляника); 7 - цинародий, особый тип сочного многоорешка с мясистым разросшимся гипантием (шиповник); Б - сухие и сочные монокарпии: 1 - однолистовка (род Consolida из сем. лютиковых); 2 - боб (большинство представителей бобовых и некоторых других семейств); 3 - членистый боб; 4 - сухая однокостянка (миндаль); 5 - сочная однокостянка (род Prunus из розоцветных)
Многоорешек - сборный многосеменной плод, возникающий из апокарпного гинецея и состоящий из отдельных односеменных невскрывающихся плодиков - орешков. Многоорешки характерны для видов лютиков, лапчаток, горицветов, шиповников. Многоорешек шиповника, плодики которого сидят внутри сильновогнутого кувшинчатого сочного гипантия, называют цинородием. У многоорешка земляники и клубники мелкие орешки сидят на выпуклой, сильно разросшейся поверхности мясистого сочного цветоложа. У этих растений такое видоизменение многоорешка получило название «земляничина» или «фрага».
Многокостянка - сборный плод, возникающий из апокарпного гинецея и состоящий из 2 или множества плодиков - костянок. Мезокарпий каждого такого плодика сочный, а эндокарпий - твердый, склерефицированный (малина, ежевика, морошка, костяника).
Плоды ценокарпии - самая многочисленная группа плодов. Возникают они из цветков как с верхней, так и с нижней завязью; при этом хотя завязь в цветке одна, она образована двумя сросшимися плодолистиками и более (рис. 4.15).
Ягода - многосеменной невскрывающийся плод с сочным мясис- тым эндо- и мезокарпом и тонким пленчатым или кожистым экзокарпом, в который погружены семена. Например, плод помидора, картофеля, возникающий из гинецея, образованного 2 сросшимися плодолистиками.
Гесперидий, или померанец, формируется из синкарпного гинецея, образованного 8-12 плодолистиками с верхней завязью. Это плод цитрусовых из семейства Рутовые (мандарин, апельсин, грейпфрут, лимон). Экзокарп - желтая ткань с масляными желёзками с различным набором эфирных масел, покрытая кутикулой и слоем воска. Мезокарп - рыхлая белая безвкусная ткань. Эндокарп пленчатый. Субэпидермальные клетки эндокарпа формируют соковые мешочки на длинной ножке; мешочки заполняются клеточным соком и образуют съедобную мякоть плода.
Тыквина - плод, близкий к ягоде, образуется из паракарпного гинецея, состоящего из 3 плодолистиков с нижней завязью; отличается кожистым жестким экзокарпием (представители семейства Тыквенные). Мякоть плода образована мезо- и эндокарпом (тыква, дыня) или разросшимися плацентами (арбуз).
Коробочка - многосеменной плод с сухим вскрывающимся пе- рикарпием. Возникает из гинецея, образованного 2 плодолистиками и более. Коробочки варьируют по форме и способам вскрывания.
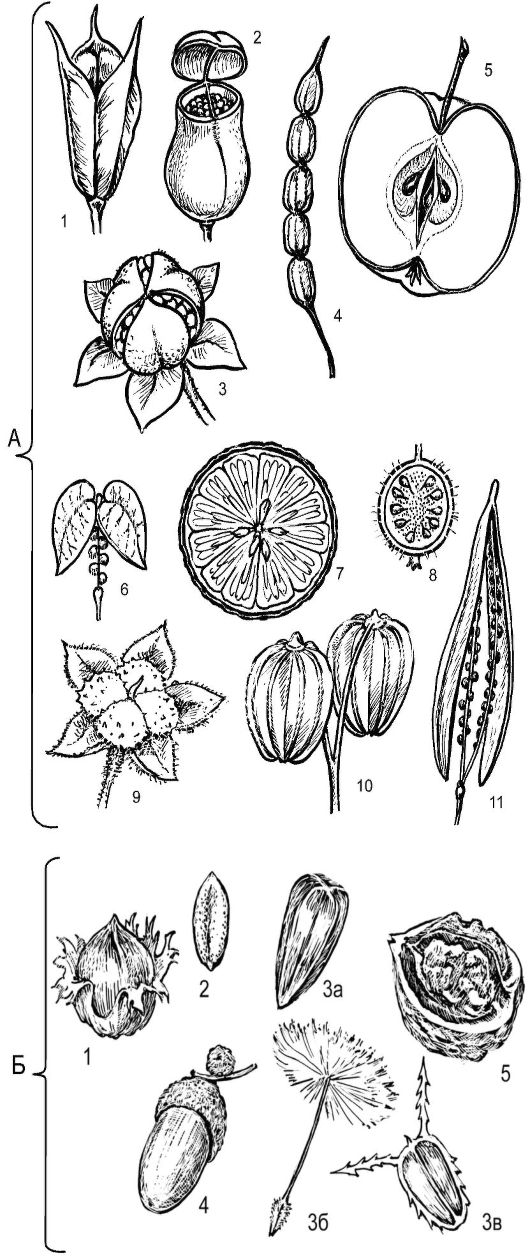
Рис. 4.15. Типы ценокарпных и псевдомонокарпных плодов: А - сухие и сочные ценокарпии: 1 - ценокарпная многолистовка (водосбор из семейства лютиковых); 2 - коробочка, вскрывающаяся крышечкой (белена); 3 - коробочка, вскрывающаяся по створкам (представители многих семейств); 4 - членистый стручок (редька дикая из крестоцветных); 5 - яблоко (все представители подсемейства Яблоневых, сем. Розоцветных); 6 - стручочек (многие крестоцветные); 7 - гесперидий, или померанец (плоды цитрусовых); 8 - ягода (представители многих семейств, типичные ягоды у черники, брусники, винограда и т. д.); 9 - ценобий, видны
4 ярема (плоды бурачниковых и губоцветных); 10 - вислоплодник, разделившийся на 2 мерикарпия, - пример дробного ценокарпия (плоды зонтичных); 11 - стручок (большинство крестоцвет- ных);
Б - сочные и сухие псевдомонокарпии: 1 - орех (лещина); 2 - зерновка (злаки); 3 - семянки различного типа: 3а - под- солнечника, 3б - одуванчика, 3в - череды; 4 - желудь (плоды буковых);
5 - псевдомонокарпная костянка (грецкий орех)
Они могут вскрываться зубчиками на верхушке (примулы), дырочками (мак), крышечкой (подорожник, белена), а могут - створками, расходящимися по месту срастания плодолистиков (хлопчатник), по средней жилке плодолистиков (тюльпан, ирис, лилия). У чистотела большого коробочка удлиненная, стручковидная, вскрывается 2 створками.
Схизокарпий. Многие ценокарпии не вскрываются, а распадаются на отдельные доли, отсюда и название: схизокарпий (греч. schiso - раскалываю). Схизокарпий распадается продольно на отдельные замкнутые или вскрывающиеся доли мерикарпии, содержащие одно или несколько семян. Плоды мальвовых (канатник) распадаются на незамкнутые с брюшной стороны мерикарпии, называемые калачиками. Схизокарпий сельдерейных (вислоплодник) состоит из 2 мерикарпиев, висящих на карпофоре, который представляет собой сросшиеся брюшные участки 2 плодолистиков. Схизокарпий многих яснотковых называется ценобием. Он возникает из двугнездной завя- зи, у которой на ранних стадиях развития появляются перегородки, разделяющие ее на 4 гнезда; в каждом гнезде располагается по 1 семязачатку. Зрелый плод состоит из 4 долей.
Стручок - сухой двугнездный многосеменной плод, возникает из гинецея, образованного двумя сросшимися плодолистиками. По краям срастания плодолистиков на рамке располагаются семена. Стручки бывают невскрывающиеся и вскрывающиеся (вскрываются двумя швами по месту срастания краев плодолистиков от основания к верхушке). Стручки могут быть членистыми (редька дикая). У стручка длина превышает ширину в 4 раза и более (горчица, капуста), у стручочков она приблизительно равна ширине (ярутка полевая, пастушья сумка).
Яблоко - многосеменной невскрывающийся плод, образованный из цветка с нижней завязью. Завязь в цветке образует от 3 до 5 сросшихся плодолистиков (яблоня, рябина, боярышник). Мякоть плода яблока развивается главным образом из цветочной ткани, в меньшей - из тканей экзо- и мезокарпа; эндокарп - кожистый, образующий стенки гнезд с семенами (представители подсемейства Яблоневые семейства Розовых - яблоня, груша, айва, рябина). Ябло- ко костянковидное - мякоть плода образуется только из тканей гипантия; экзо- и мезокарп - пленчатые, эндокарп - деревянистый, окружающий каждое семя (боярышник).
Плоды псевдомонокарпии - образуются из цветков с псевдомонокарпным гинецеем, в котором закладываются 2 плодолистика или
более, но развивается только 1, а остальные редуцируются (см. рис.
4.15).
Семянка - односеменной плод с кожистым перикарпием, не сра- стающимся с семенем. Характерен для всех представителей семейства Сложноцветных с нижней (подсолнечник, василек, ромашка) и верхней (коноплевые, крапивные) завязью. Семянки многих астровых (одуванчик) снабжены летучками, развившимися из видоизмененной чашечки.
Зерновка - односеменной плод с тонким пленчатым перикарпием, срастающимся с семенем. Характерен для всех злаков (пшеница, рожь, тимофеевка). Зерновка формируется из верхней завязи, состоящей из 3 (бамбук), чаще из 2 (пшеница, рожь) плодолистиков.
Орех - односеменной, редко двусеменной плод с сильно склерефицированным, почти деревянистым перикарпием, не сросшимся с кожурой семени (лещина, ольха, береза).
Крылатка, крылатый орех - это орех без плюски, снабженный крылом. У крылатки крыло образуется из сросшихся с околоплодником чешуевидных прицветников и прицветничков (береза, ольха, граб), у крылатого ореха из прирастающих к околоплоднику сегментов околоцветника (вяз, ревень, щавель).
Желудь - околоплодник менее жесткий, чем у ореха, тонко- кожистый или тонкодеревянистый, не срастающийся с семенем (дуб, бук). У желудя дуба плюска блюдцевидная. Она образована видоизмененными стерильными веточками соцветия.
СЕМЯ
Семя - орган полового размножения и расселения растений, развивающихся в основном из оплодотворенного семязачатка. Развитие зародыша и семян после двойного оплодотворения получило название «амфимиксис» (от греч. amphi - с обеих сторон). Развитие зародыша и семян может происходить и без оплодотворения - апомиксис. В результате апомиксиса при мегаспорогенезе мейоз не происходит, поэтому все клетки зародышевого мешка диплоидны. Зародыш может образоваться из яйцеклетки (партеногенез), из любой другой клетки зародышевого мешка (апогамия), из клеток нуцеллуса и т.д. Апомиксис часто встречается у представителей семейств розовых, рутовых, пасленовых, астровых, мятликовых.
Семя состоит из зародыша, эндосперма, семенной кожуры. Зародыш - это миниатюрный спорофит, являющийся основной частью
семени. В нем различают 3 зародышевых органа: зародышевый корешок, зародышевый стебелек с зародышевой почкой и зародыше- вые листья (семядоли). Зародышевый побег представлен осью (зародышевым стебельком) и семядольными листьями, или семядолями: 2 - у двудольных и 1 - у однодольных растений (у зародыша однодольных намечаются зачатки 2 семядолей, но одна из них не получает дальнейшего развития). Участок стебелька в зародыше выше семядолей называется эпикотилем, или надсемядольным коленом, ниже семядолей - гипокотилем, или подсемядольным коленом.
Семенная кожура обычно многослойна и присутствует у семени всегда. Ее главная функция - защита зародыша от чрезмерного высыхания; она также предохраняет зародыш от преждевременного прорастания. При прорастании первые порции воды проникают внутрь семени через отверстие в семенной кожуре - микропиле.
Эндосперм обычно состоит из округлых клеток запасающей ткани. Это могут быть зерна крахмала или капли жирного масла, нередко в сочетании с запасными белками. Вещества эндосперма гидролизуются при набухании семян под действием ферментов и поглощаются зародышем в процессе прорастания; после этого его клетки разрушаются.
Виды семян
Различают 4 типа семян: 1) с эндоспермом; 2) с эндоспермом и периспермом; 3) с периспермом; 4) без эндосперма и перисперма.
Семена двудольных без эндосперма. К этой категории относят семена бобовых, тыквенных, сложноцветных, крестоцветных, дуба, березы, клена и др. На рисунке 4.15 представлено строение семени и зародыша тыквы (Cucurbita реро). Под плотной кожурой находится плоский зародыш с крупными семядолями, в тканях которых сосредоточены запасы питательных веществ. Эндосперм отсутствует - он «съеден» в процессе созревания семени. На семядолях заметно зачаточное жилкование. Ось зародыша небольшая, обращенная корневым полюсом к микропиле; на этом же конце семени находится рубчик. Зародышевая почечка выражена слабо: на конусе нарастания побега еле заметны листовые бугорки - зачатки следующих за семядолями листьев.
Обычный учебный объект - семена бобовых. На рисунке 4.16 изображены детали строения зрелого семени фасоли, где эндосперма нет и запасающими органами являются крупные, сильно утолщен-
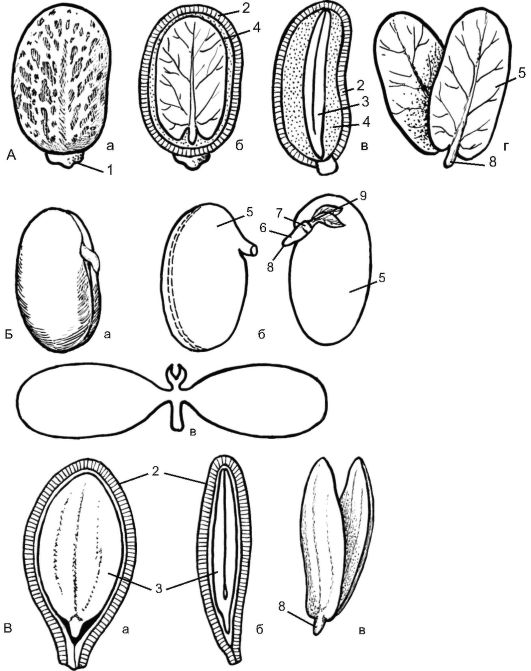
Рис. 4.16. Семена двудольных: клещевины (c эндоспермом); фасоли, тыквы (без эндосперма): А - клещевина: а - внешний вид семе- ни; б - продольный разрез в плоскости семядолей зародыша; в - продольный разрез, перпендикулярный плоскости семядолей; г - изолированный и полуразвернутый зародыш;
Б - фасоль: а - вычлененный зародыш; б - расчлененный зародыш; в - схема зародыша в выпрямленном виде);
В - тыква: а, б - продольный разрез семени во взаимно перпендикулярных плоскостях; в - вычлененный зародыш;
1 - присеменник; 2 - семенная кожура; 3 - зародыш; 4 - эндосперм; 5 - семядоли; 6 - гипокотиль; 7 - эпикотиль; 8 - зародышевый корешок; 9 - почечка
ные семядольные листья зародыша. Хорошо развита зародышевая почечка с эпикотилем - 1-м междоузлием побега. Зародыш фасоли (и других бобовых) вследствие неравномерного разрастания оси сильно согнут. Если мысленно выпрямить его ось и семядоли, получится схема, ничем не отличающаяся от схемы прямого зародыша тыквы и др. Согнутый или спирально закрученный зародыш иногда с разнообразно сложенными в складки семядолями имеется у многих двудольных, в том числе у крестоцветных (капуста, редис и др.).
Семена двудольных с эндоспермом. Между семядолями находится конус нарастания побега; почечка еще не сформирована (у семени клещевины) (см. рис. 4.16).
Семена двудольных с периспермом и эндоспермом. Иногда, помимо эндосперма, в семенах развивается запасающая ткань иного происхождения - перисперм, возникающая из нуцеллуса семязачатка и лежащая под кожурой. Функционально эндосперм и перисперм равноценны, хотя морфологически имеют разное происхождение: они аналоги, но не гомологи (у семени свеклы) (рис. 4.17).
Например, в семени перца черного (Piper nigrum) маленький двусемядольный зародыш погружен в небольшой эндосперм, а кнаружи от него располагается мощный перисперм (см. рис. 4.17). Иногда эндосперм в зрелом семени поглощается полностью, а перисперм остается и разрастается, как у гвоздичных, лебедовых (например, у звездчатки, куколя, свеклы).
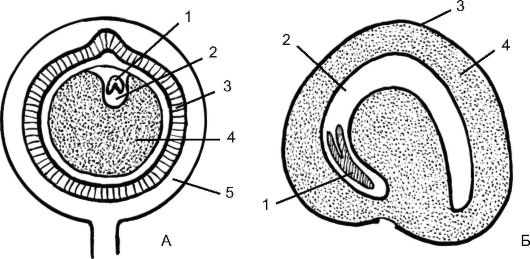
Рис. 4.17. Семена двудольных растений с периспермом: А - плодик перца черного; Б - незрелое семя свеклы (виден эндосперм, который потом исчезает); 1 - зародыш; 2 - эндосперм; 3 - семенная кожура; 4 - перисперм; 5 - околоплодник
Семена однодольных с эндоспермом. К этой категории принадлежит преобладающее большинство семян однодольных. Один из самых наглядных примеров типичного строения семени однодольных - семя ириса, или касатика (любого вида, дикорастущего или культурного). На рисунке 4.18 изображено строение семени желтого водяного ириса (Iris pseudacorus). Крупные уплощенные семена со- зревают в плоде-коробочке и одеты плотной коричневой кожурой. Внутренний слой кожуры обычно отстает от наружного, образуя воздушную камеру. Это способствует увеличению плавучести семян, распространяемых с током воды. Большую часть объема семени занимает эндосперм, богатый маслами и белками. В него погружен палочковидный прямой зародыш. Зачаток корешка обращен кончиком к микропиле; он переходит в прямой гипокотиль, оканчивающийся меристематическим апексом (верхушкой) побега, смещенным вбок. Семядоля цилиндрическая; ее нижняя часть представляет собой влагалище, охватывающее конус нарастания со всех сторон и прикрывающее его. Функция влагалища семядоли - защита точки роста. Очень похожи на зародыш ириса зародыши многих представителей лилейных - центрального семейства однодольных, например лука репчатого (Allium сера).
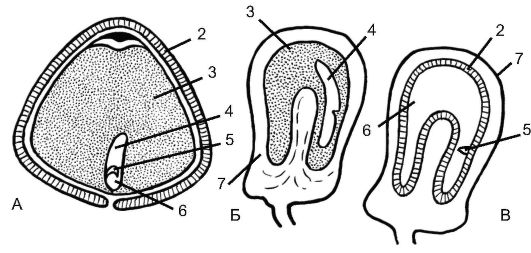
Рис. 4.18. Семена однодольных растений с эндоспермом (А - ирис) и без эндосперма (Б, В - частуха подорожниковая): А - ирис; Б, В - частуха подорожниковая (Б - незрелое семя, виден остаток эндосперма, В - зрелое, без эндосперма);.
У незрелого семени частухи (Б) виден остаток эндосперма, у зрелого семени (В) эндосперм исчезает; 1 - воздушная полость; 2 - семенная кожура; 3 - эндосперм; 4 - зародыш; 5 - почечка; 6 - семядоля; 7 - околоплодник
Семена злаков. Строение семени злаков (семейство Роасеае) до- вольно своеобразно (рис. 4.19). Зародыш в плоде-зерновке соприкасается с эндоспермом одной стороной, а не окружен его тканью, как у большинства других однодольных. Вследствие такого расположения семядоля злаков имеет форму плоского щитка, прижатого к эндосперму. Всасывающая функция щитка обеспечивается сильно специализированными клетками его поверхностного слоя. В отличие от большинства однодольных почечка зародыша злаков обычно довольно сильно развита, имеет несколько листовых зачатков. Наружный колпачковидный листок почечки называется колеоптилем. Гипокотиль у злаков недоразвит; зародышевый корешок окружен специальным многослойным чехлом - колеоризой, которая при прорастании набухает, на ее поверхности развиваются всасывающие во- лоски, корень пробивает ткань колеоризы, чтобы выйти наружу, в почву. Функциональное значение частей зародыша злаков в общем понятно: защита меристематических конусов нарастания колеоптилем и колеоризой, в то же время существуют весьма противоречивые гипотезы о происхождении и морфологической природе большинства зародышевых органов злаков.
Зародыш у злаков имеет структуру гораздо более сложную и специализированную, чем у большинства других однодольных, и поэтому не может считаться эталоном для всего этого класса.
Семена однодольных без эндосперма. Семя имеет форму подковы, под тонкой кожурой находится зародыш, сосредоточивший в семядоле все запасы, поглощенные им в ходе созревания семени; эндосперм им уже «съеден». Примером могут служить семена широко распространенных полуводных растений стрелолиста (Sagiltaria) и частухи подорожниковой (Alisma plantago-aquatica) (см. рис. 4.18), а также полностью погруженных в воду видов рода Рдест (Potamogeton).
Прорастание семян
Цветковые растения размножаются семенами, созревающими внутри плода. Однако во многих случаях (например, если плоды су- хие односеменные) семена распространяются, не отделяясь от околоплодника. В таких случаях посевным материалом оказываются не семена, а плоды или их части. Если плоды срастаются между собой, посевной материал морфологически представляет собой соплодие.
Для прорастания семян (многие плодовые и дикорастущие древесные) обязателен период пониженных температур. Для более быстрого
проращивания в условиях культуры семена таких растений подвергают стратификации - длительному выдерживанию при низкой температуре, во влажной среде и при хорошей аэрации. Иногда покровы семени бывают водонепроницаемыми (твердосеменные бобовые или косточковые плодовые). Такие семена подвергают скарификации (искусственное нарушение целостности покровов семени перетиранием, надрезанием, пропусканием через металлические щетки).
Прорастанию семени предшествует его набухание - процесс, связанный с поглощением большого количества воды и обводнением тканей семени. Одновременно с поглощением воды активизируются ферменты, которые переводят запасные вещества семени в легкоусвояемую, доступную для зародыша форму.
Для прорастания семян необходимы вода (ткани зрелых семян сильно обезвожены), кислород для дыхания, определенная темпера- тура, а иногда и свет. Прорастание семян - это переход их от со- стояния покоя к росту зародыша и формированию проростка.
На первых этапах развития проросток питается органическими веществами, запасенными в семени, т.е. гетеротрофно. С появле- нием 1-го срединного листа проросток превращается в сеянец, который начинает самостоятельно синтезировать органические вещества. Однако некоторое время он еще продолжает пользоваться запасами
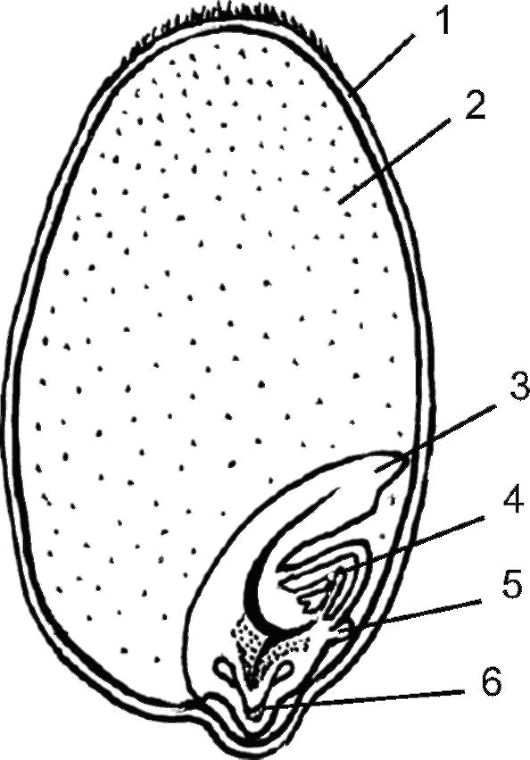
Рис. 4.19. Зерновка пшеницы:
схема продольного разреза зерновки пшеницы; 1 - покровы зерновки; 2 - эндосперм; 3 - щиток; 4 - почечка; 5 - эпибласт; 6 - главный корень
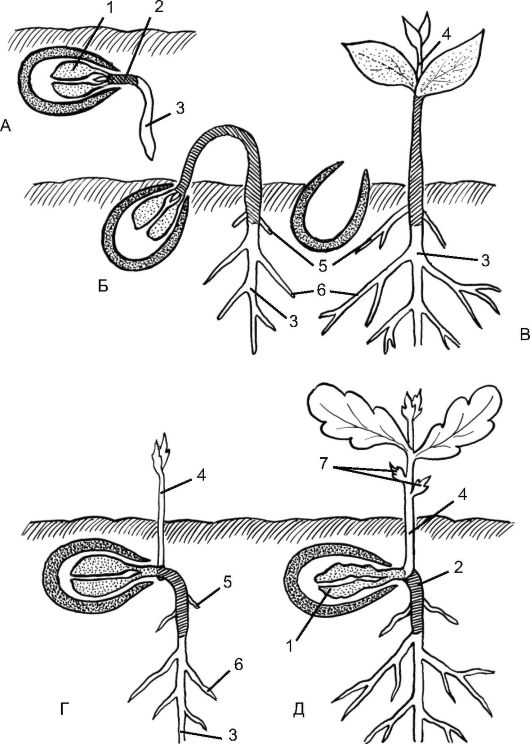
Рис. 4.20. Схема надземного и подземного прорастания двудольных растений:
А - начало прорастания семени; Б, В - этапы надземного прорастания; Г, Д - этапы подземного прорастания; 1 - семядоли; 2 - гипокотиль (выделен черным цветом); 3 - главный корень; 4 - эпикотиль; 5 - при- даточные корни; 6 - боковые корни; 7 - чешуевидные листья
семени, т.е. питание его на этом этапе смешанное. И только позже сеянец полностью переходит к автотрофному питанию.
Прорастание бывает надземным и подземным. При надземном прорастании (рис. 4.20) семядоли выносятся на поверхность, зеле- неют и становятся первыми ассимилирующими листьями. Вынос семядолей над почвой у двудольных чаще происходит за счет удлинения гипокотиля (фасоль, тыква, клен) либо в результате разрастания черешков семядолей (аконит). Гипокотиль, выйдя на поверхность, выпрямляется и вытаскивает семядоли. При надземном прорастании однодольных (лук, вороний глаз) выход семядоли на поверхность иной: за счет вставочного роста основания самой семядоли, которая петлеобразно изгибается, и при отсутствии роста гипокотиля.
При подземном прорастании семядоли, как правило, сморщиваются и отмирают, не выходя на поверхность, остаются в почве и служат вместилищем запасных питательных веществ либо гаусторием, передающим их из запасающих тканей проростку (горох, дуб, настурция, пшеница, кукуруза), а первыми ассимилирующими листьями становятся следующие за семядолями настоящие листья (см. рис. 4.20). При подземном прорастании рост гипокотиля ограничен, и побег сразу начинает расти вверх.