Ботаника: учебник. Зайчикова С.Г., Барабанов Е.И. 2013. - 288 с. : ил.
|
|
|
|
ГЛАВА III ВЕГЕТАТИВНЫЕ ОРГАНЫ ВЫСШИХ РАСТЕНИЙ
ОРГАН И ЕГО ХАРАКТЕРИСТИКА
Орган это часть растения, выполняющая определенные функции и имеющая специфичное строение. Вегетативные органы, к которым относятся корень и побег, составляют тело высших растений; они обеспечивают индивидуаль- ную жизнь особи (рис. 3.1).
У грибов и низших растений деления тела на органы нет. Их тело представлено системой мицелия или слоевищ.
Образование органов у высших растений в процессе эволюции связано с выходом их на сушу и приспособлением к наземному существованию.
КОРЕНЬ И КОРНЕВАЯ СИСТЕМА
Общая характеристика корня
Корень (от лат. radix) - осевой орган, цилин- дрической формы, обладающий радиальной симметрией и положительным геотропизмом. Он способен к росту до тех пор, пока сохраняется апикальная меристема. Морфологически корень отличается от побега тем, что на нем
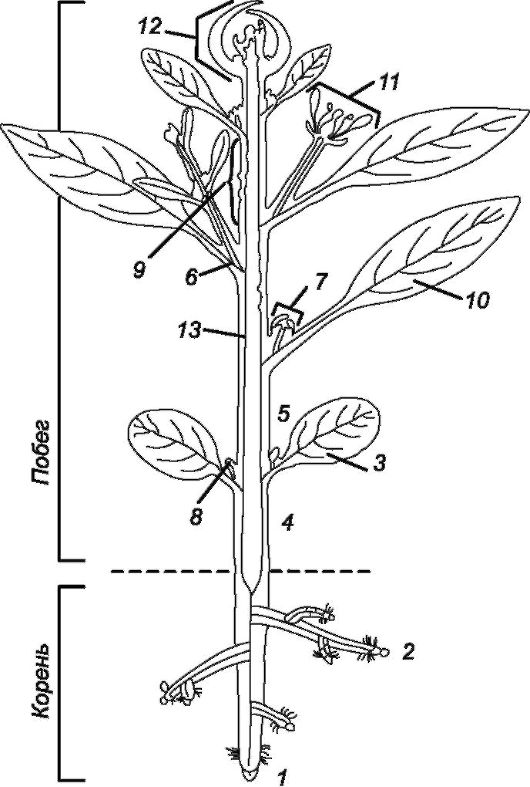
Рис. 3.1. Схема расчленения тела высшего растения на примере строения двудольного растения (показаны также репродуктивные органы):
1 - главный корень; 2 - боковые корни; 3 - семядоли; 4 - гипокотиль; 5 - эпикотиль; 6 - узел; 7 - пазуха листа; 8 - пазушная почка; 9 - междоузлие; 10 - лист;
11 - цветок; 12 - верхушечная почка; 13 - стебель
никогда не возникают листья, а апикальная меристема прикрыта корневым чехлом. Корень, как и побег, может ветвиться, формируя корневую систему.
Функции корня
1. Минеральное и водное питание (поглощение воды и минеральных веществ).
2. Закрепление растения в почве (заякоревание).
3. Синтез продуктов первичного и вторичного метаболизма.
4. Накопление запасных веществ.
5. Вегетативное размножение.
6. Симбиоз с бактериями.
7. Функция дыхательного органа (монстера, филодендрон и др.)
Типы корней и корневых систем
По происхождению корни делятся на главные, боковые и придаточ- ные. Главный корень семенных растений развивается из корешка заро-
дыша семени. Стебель является продолжением корня, и вместе они составляют ось 1-го порядка. Место сочленения оси и семядольных листьев называют семядольным узлом. Участок, расположенный на границе главного корня и стебля, именуется корневой шейкой. Участок стебля от корневой шейки до первых зародышевых листьев (семядолей) называют подсемядольным коленом, или гипокотилем, а от семядолей до первых настоящих листьев - эпикотилем, или надсемядольным коленом. У двудольных и голосеменных растений от главного корня за счет меристематической активности перицикла отходят боковые корни 1-го порядка, которые дают начало боковым корням 2-го и 3-го порядка. Корневая система, образованная системой главного корня, называется стержневой, а с развитой системой боковых корней - ветвистой; таким образом, ветвистая корневая система является разновидностью стержневой. Чем больше боковых корней отходит от главного, тем больше площадь питания растения.
У большинства двудольных растений главный корень сохраняется всю жизнь, у однодольных растений главный корень не развивается, так как зародышевый корешок быстро отмирает, а от базальной части побега берут начало придаточные корни. Придаточные корни могут образовываться от листьев, стеблей, старых корней и даже от цветков
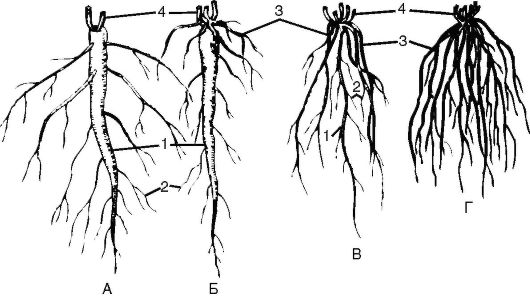
Рис. 3.2. Типы корневых систем: по форме: А, Б - стержневая; В, Г - мочковатая;
по происхождению: А - система главного корня; Б, В - смешанная корневая система; Г - придаточная корневая система; 1 - главный корень; 2 - боковые корни; 3 - придаточные корни; 4 - основания побегов
и иметь ответвления 1-го, 2-го порядка и т.д. Корневая система, образованная придаточными корнями, называется мочковатой (рис. 3.2). У многих двудольных корневищных растений главный корень часто отмирает и преобладает система придаточных корней, отходящих от корневища (лютик ползучий, сныть обыкновенная).
По отношению к субстрату корни бывают следующих типов: земляные - развиваются в почве; водные - находятся в воде (у плавающих водных растений); воздушные, развивающиеся в воздушной среде (у растений, имеющих корни на стволах и листьях).
Зоны корня
В молодом корне различают 4 зоны: деления, растяжения, всасывания, проведения (рис. 3.3).
К зоне деления относят верхушку конуса нарастания (протяженность менее
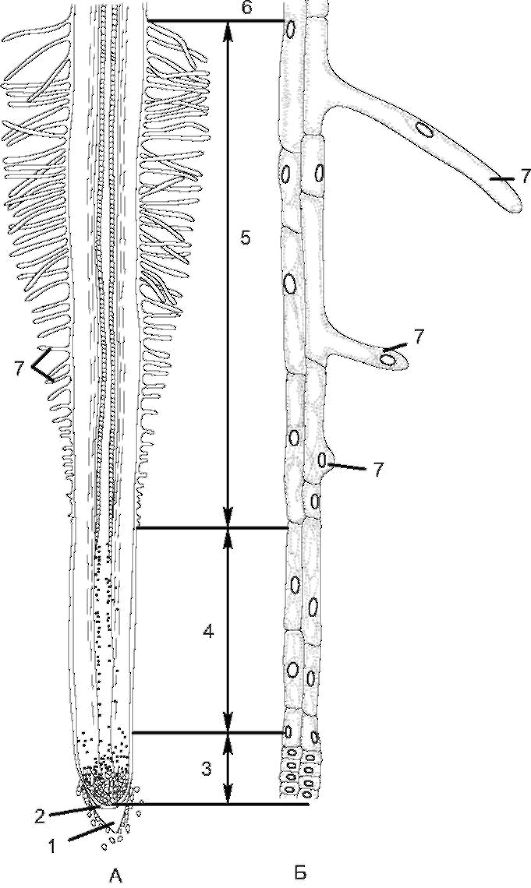
Рис. 3.3. Зоны корня (у проростка пшеницы): А - схема строения корня; Б - периферические клетки отдельных зон при большом увеличении: 1 - корневой чехлик; 2 - калиптроген; 3 - зона деления; 4 - зона растяжения; 5 - зона всасывания; 6 - зона проведения; 7 - корневые волоски
клеток. Верхушечная меристема откладывает наружу клетки корневого чехлика, а внутрь - ткани остальной части корня. Эта зона состоит из тонкостенных паренхимных клеток первичной меристемы, которые прикрыты корневым чехликом, выполняющим защитную функцию при продвижении корня между частичками почвы. От соприкосновения с почвой клетки чехлика постоянно разрушаются, образуя слизь, которая предохраняет зону деления при трении о по- чву и продвижении корня вглубь. У большинства растений корневой чехлик восстанавливается за счет первичной меристемы, а у злаков - за счет особой меристемы калиптрогена.
По теории гистогенов (Ганштейн, 1868), у большинства покрытосеменных растений апикальные меристемы состоят из 3 гистогенных слоев, различающихся направлением деления клеток и имеющих по 1-4 инициальные клетки. Самый наружный слой - дерматоген - формирует протодерму, из которой образуются клетки корневого чехлика и ризодерма - первичная покровно-всасывающая ткань в зоне всасывания. Средний слой - периблема - дает начало всем тканям первичной коры. Третий слой инициалей формирует плером, из которого развиваются ткани центрального осевого цилиндра.
В зоне растяжения клетки меристемы увеличиваются в размерах (вследствие оводнения), вытягиваются в длину, и деление клеток постепенно прекращается. Вследствие вытягивания клеток в продольном направлении происходят рост корня в длину и его продвижение в почве. Зону деления и зону растяжения с учетом сохранения в них меристематической активности можно объединить в одну - зону роста. Ее протяженность составляет несколько миллиметров. В зоне всасывания происходит становление первичной структуры корня.
Протяженность зоны всасывания - от нескольких миллиметров до нескольких сантиметров; она характеризуется наличием корневых волосков, являющихся выростами клеток ризодермы. При их формировании ядро перемещается в переднюю часть корневого волоска. Последние увеличивают всасывающую поверхность корня и обеспечивают активное всасывание воды и растворов солей, но они недолговечны (живут 10-20 дней). Новые корневые волоски образуются под зоной всасывания, а отмирают - над этой зоной. По мере роста растения зона всасывания постепенно перемещается, и растение имеет возможность поглощать минеральные вещества из разных слоев почвы.
Постепенно зона всасывания переходит в зону проведения (укрепления). Она тянется вплоть до корневой шейки и составляет по протяжен-
ности большую часть корня. В этой зоне идет интенсивное ветвление главного корня и появляются боковые корни. У двудольных растений в зоне проведения формируется вторичная структура корня.
Анатомия корня
Первичное строение корня (рис.
Покровно-всасывающая ткань - ризодерма (эпиблема) выполняет как покровную функцию, так и функцию интенсивного всасывания воды и минеральных веществ из почвы. Клетки ризодермы живые, с тонкой целлюлозной стенкой. Из некоторых клеток ризодермы формируются корневые волоски; каждый из них представляет собой длинный вырост одной из клеток ризодермы, ядро же клетки обычно находится в кончике выроста. Корневой волосок содержит тонкий пристенный слой цитоплазмы, более плотный на верхушке волоска, а в центре - крупную вакуоль. Корневые волоски недолговечны и в зоне укрепления отмирают. Физиологически зона всасывания представляет собой очень важную часть корня. Клетки ризодермы поглощают водные растворы всей поверхностью наружных стенок. Развитие корневых волосков во много раз увеличивает поверхность поглощения. Протяженность зоны всасывания - от 1 до
Со временем эпиблема может слущиваться, и тогда покровную функцию выполняет экзодерма, а после ее разрушения - слой кле- ток мезодермы и иногда мезодермы и перицикла, стенки которых опробковевают и одревесневают. Поэтому диаметр старых корней однодольных растений меньше, чем молодых.
Первичная кора корня развита более мощно, чем центральный осевой цилиндр. Она состоит из 3 слоев: экзодермы, мезодермы (рис.
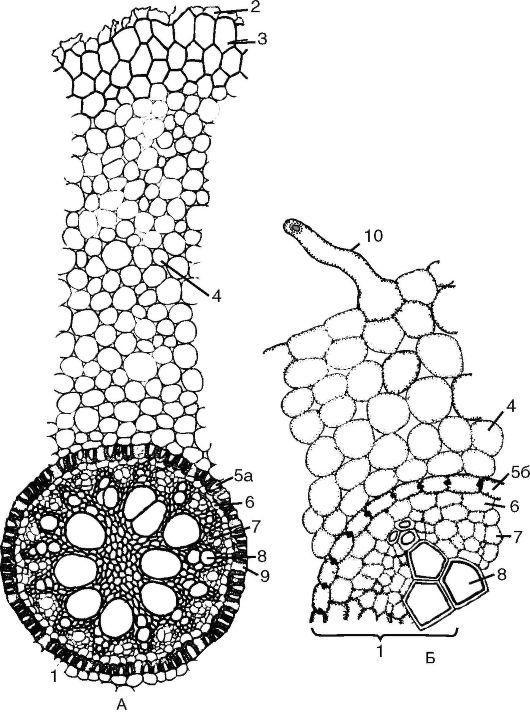
Рис. 3.4. Поперечный срез корня первичного строения: А - первичное строение корня однодольного;
Б - первичное строение корня двудольного: 1 - центральный (осевой) цилиндр; 2 - остатки эпиблемы; 3 - экзодерма; 4 - мезодерма; 5а - эндодерма с подковообразными утолщениями; 5б - эндодерма с поясками Каспари; 6 - перицикл; 7 - первичная флоэма; 8 - сосуды первичной ксилемы; 9 - пропускные клетки эндодермы; 10 - корневой волосок
воды и газов. В экзодерме, обычно под корневыми волосками, сохраняются клетки с тонкими целлюлозными стенками - пропускные клетки, через которые проходят вода и минеральные вещества, по- глощенные ризодермой. Обычно они располагаются напротив лучей ксилемы радиального пучка.
Под экзодермой находятся живые паренхимные клетки мезодер- мы. Это наиболее широкая часть первичной коры. Клетки мезодермы выполняют запасающую функцию, а также функцию проведения воды и растворенных в ней солей от корневых волосков в центральный осевой цилиндр.
Внутренний однорядный слой первичной коры представлен эндодермой. Клетки эндодермы плотно сомкнуты и почти квадратные в поперечном сечении. В зависимости от степени утолщения клеточной стенки различают 2 типа эндодермы - с поясками Каспари (на поперечном срезе они выглядят как пятна Каспари) и с подковообразными утолщениями стенок.
Эндодерма с поясками Каспари - это начальный этап формирования эндодермы, при котором утолщению подвергаются лишь ее радиальные стенки за счет отложения веществ, сходных по химическому составу с суберином и лигнином. У многих двудольных и голосеменных растений процесс дифференциации эндодермы поясками Каспари заканчивается. У эндодермы с подковообразными утолщениями образуется толстая вторичная клеточная стенка, пропитанная суберином, в дальнейшем она одревесневает. Неутолщенной остается только наружная клеточная стенка (рис. 3.5). Эндодерма с подковообразными утолщениями развивается чаще у однодольных растений (рис.
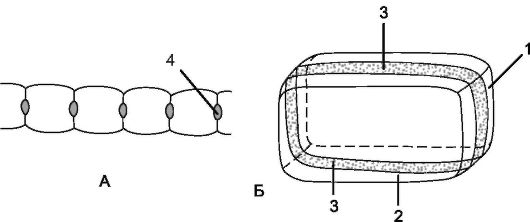
Рис. 3.5. Схема строения клетки эндодермы: А - общий вид; Б - поперечный разрез клеток: 1 - поперечная стенка клетки; 2 - продольная радиальная стенка; 3 - поясок Каспари; 4 - пятна Каспари
Считается, что эндодерма выполняет функцию гидравлического барьера, способствуя продвижению минеральных веществ и воды из первичной коры в центральный осевой цилиндр и препятствуя их выходу обратно.
Центральный осевой цилиндр начинается с клеток перицикла, который обычно в молодых корнях состоит из живых тонкостенных паренхимных клеток, расположенных в один ряд (но может быть и многослойным - например, у грецкого ореха). Клетки перицикла дольше других тканей корня сохраняют свойства меристемы и спо- собность к новообразованиям. Из перицикла образуются боковые корни, поэтому его называют корнеродным слоем. Проводящая система корня представлена одним радиальным сосудисто-волокнистым пучком, в котором группы элементов первичной ксилемы чередуются с участками первичной флоэмы. У однодольных растений количество лучей первичной ксилемы - 6 и более, у двудольных - от 1 до 5. Корни в отличие от стеблей не имеют сердцевины, так как в центре корня располагаются лучи первичной ксилемы.
Таблица 3.1. Формирование тканей корня первичного и вторичного строения
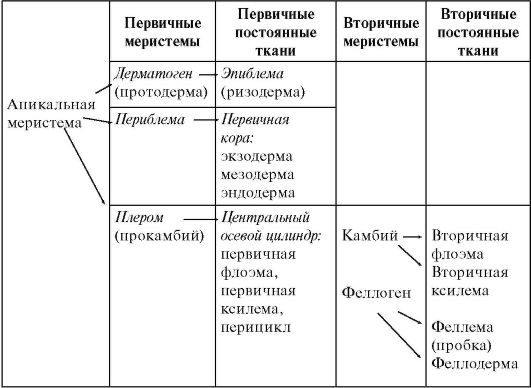
У однодольных и споровых архегониальных растений строение корня не претерпевает значительных изменений в течение всей жизни растения. У голосеменных и двудольных растений на границе зон всасывания и проведения происходит переход от первичного строения корня ко вторичному (табл. 3.1).
Вторичное строение корня. В корнях голосеменных и двудольных растений камбий возникает из прокамбия (камбиальных дуг) за счет тангентального деления тонкостенных клеток, расположенных с внутренней стороны от флоэмных тяжей. На поперечном срезе клетки камбия представлены вогнутыми внутрь дугами (рис.
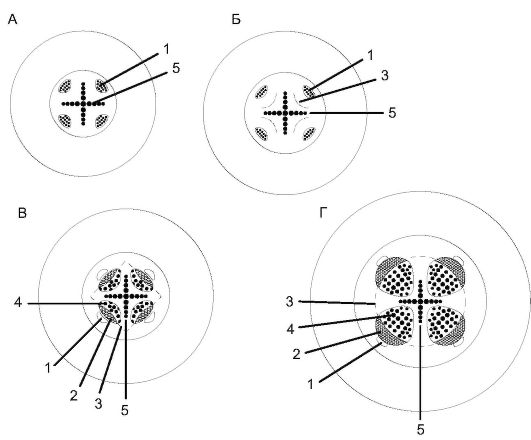
Рис. 3.6. Схема развития вторичного строения в корне: А - первичное строение; Б - заложение камбия; В - начало образования вторичных коллатеральных пучков; Г - вторичное строение корня: 1 - первичная флоэма; 2 - вторичная флоэма; 3 - камбий; 4 - вторичная ксилема; 5 - первичная ксилема
При этом дуги камбия сначала выпрямляются, а затем принимают выпуклую форму.
Когда дуги камбия достигают перицикла, его клетки тоже начинают делиться и образуют камбий межпучковый, а тот, в свою очередь, - сердцевинные лучи, представленные паренхимными клетка- ми, отходящими от лучей первичной ксилемы. Сердцевинные лучи, образованные межпучковым камбием, - это изначально «первичные лучи».
Таким образом, в результате деятельности камбия в корне между лучами первичной ксилемы формируются открытые коллатеральные сосудисто-волокнистые пучки, число которых равно числу лучей первичной ксилемы. Первичная флоэма при этом оттесняется вторичными тканями к периферии и сплющивается (рис. 3.6 и 3.7).
В перицикле, кроме межпучкового камбия, может закладываться феллоген, дающий начало перидерме - вторичной покровной ткани. При тангентальном делении клеток феллогена наружу отделяются клетки пробки, а внутрь - клетки феллодермы. Непроницаемость клеток пробки, пропитанных суберином, является причиной изоляции первичной коры от центрального осевого цилиндра. Первичная кора при этом постепенно отмирает и сбрасывается. Все ткани, располагающиеся от периферии до камбия, входят в понятие «вторич- ная кора» (рис.
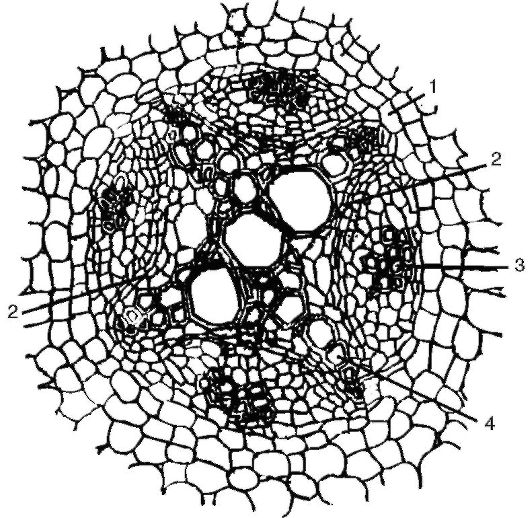
Рис. 3.7. Переход к вторичному строению корня (заложение камбиального кольца): 1 - перицикл; 2 - камбий; 3 - первичная флоэма; 4 - первичная ксилема
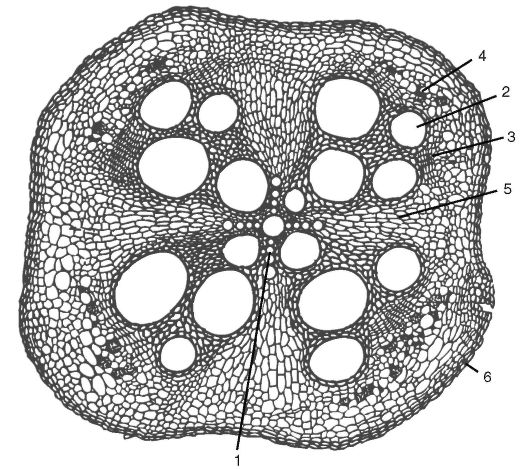
Рис. 3.8. Вторичное строение корня тыквы. Первичная кора слущилась: 1 - остаток первичной ксилемы (четыре луча); 2 - сосуды вторичной ксилемы; 3 - камбий; 4 - вторичная флоэма; 5 - сердцевинный луч; 6 - пробка
между которыми располагаются открытые коллатеральные пучки в количестве, соответствующем лучам первичной ксилемы (рис. 3.8).
Метаморфозы корней Микориза
Микориза (от греч. mykes - гриб и rhiza - корень) - это симбиотическое взаимодействие гиф гриба и корневых окончаний растения. Грибы, живущие на корнях растений, используют органиче- ские вещества, синтезируемые зеленым растением, и поставляют растению из почвы воду и минеральные вещества. Благодаря сильно
разветвленным гифам гриба у растения увеличивается всасывающая поверхность. Но иногда симбиоз может переходить и в паразитизм, например, когда гифы гриба наносят растению вред, угнетая его рост, или когда растение начинает использовать клеточное содержимое гиф гриба, проникших в кóровую паренхиму растения.
Клубеньки
Наличие клубеньков характерно для предствителей семейства Бобовые (люпин, клевер и др.). Клубеньки образуются в результате проникновения через корневые волоски в кору корня бактерий рода Rizobium. Бактерии вызывают усиленное деление паренхимы, которая образует выросты бактероидной ткани на корне - клубеньки. Бактерии фиксируют атмосферный молекулярный азот и переводят его в связанное состояние в виде азотистых соединений, усваиваемых растением. Бактерии, в свою очередь, используют вещества, находящиеся в корнях растения. Такой симбиоз очень важен для почвы и используется в сельском хозяйстве при обогащении почв азотистыми веществами.
Воздушные корни
У ряда тропических травянистых растений, живущих на деревьях, для поднятия вверх, к свету, образуются воздушные корни, которые свободно свисают вниз. Воздушные корни способны усваивать влагу, выпадающую в виде дождя и росы. На поверхности этих корней образуется своеобразная покровная ткань - веламен - в виде многослойной мертвой ткани, клетки которой имеют спиральные или сетчатые утолщения.
Корневые клубни
У многих двудольных и однодольных растений в результате метаморфоза боковых и придаточных корней образуются корнеклубни (чистяк весенний и т.д.). Корневые клубни имеют ограниченный рост и приобретают овальную или веретеновидную форму. Такие клубни выполняют запасающую функцию, а поглощение почвенных растворов за них осуществляют хорошо ветвящиеся всасывающие корни. У некоторых растений (таких, как георгин) корнеклубни выполняют запасающую функцию только в определенной части (базальной, срединной), а остальная часть клубня имеет типичное строение корня. Такие корнеклубни могут выполнять и запасающую, и всасывающую функции.
Корнеплоды
В образовании корнеплода могут участвовать различные части растения: разросшаяся базальная часть главного корня, утолщенный гипокотиль и др. Короткокорнеплодные сорта представителей семейства Капустные (редис, репа) имеют плоский или округлый клубень, большая часть которого представлена разросшимся гипокотилем. Такие корнеплоды имеют вторичное анатомическое строение при диархной (двулучевой) первичной ксилеме и хорошо развитой вторичной, выполняющей запасающую функцию (рис.
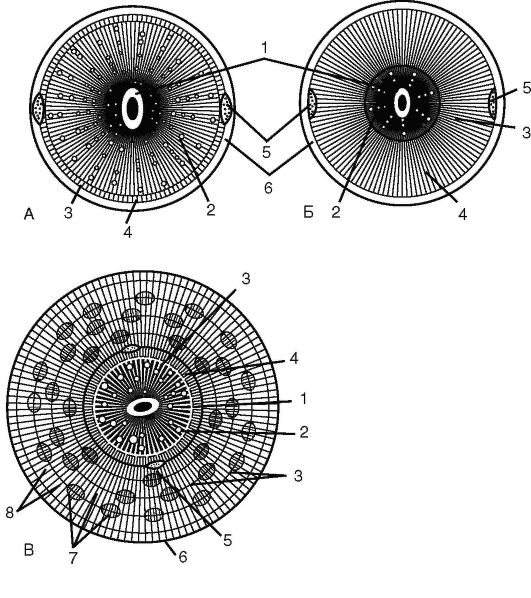
Рис. 3.9. Схема строения корнеплодов: А - тип редьки; Б - тип моркови; В - тип свеклы;
1 - первичная ксилема;
2 - вторичная ксилема; 3 - камбий; 4 - вторичная флоэма; 5 - первичная флоэма; 6 - перидерма; 7 - проводящие пучки; 8 - запасающая паренхима
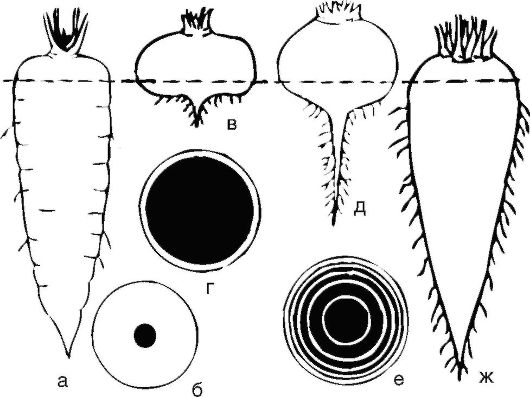
Рис. 3.10. Корнеплоды: моркови (а, б); репы (в, г); свеклы (д, е, ж). На поперечных срезах ксилема показана черным; пунктирной линией обозначена граница стебля и корня
вторичная флоэма (рис.
ПОБЕГ И СИСТЕМА ПОБЕГОВ
Общая характеристика побегов и почки
Побег состоит из оси стебля и отходящих от его листьев и почек. В более конкретном смысле побегом можно назвать однолетний неразветвленный стебель с листьями и почками, развившейся из почки или семени. Побег развивается из почечки зародыша или пазушной почки и представляет собой один из основных органов высших растений. Таким образом, почка является зачаточным побегом. Функция побега состоит в воздушном питании растения. Видоизмененный побег - в виде цветка или спороносного побега - выполняет функцию размножения.
Основные органы побега - стебель и листья, формирующиеся из меристемы конуса нарастания и обладающие единой проводящей системой (рис. 3.11). Участок стебля, от которого отходит лист (или листья), называют узлом, а расстояние между узлами - междоузлием. В зависимости от длины междоузлия каждый повторяемый узел с междоузлием называется метамером. Как правило, метамеров вдоль оси побега бывает много, т.е. побег состоит из серии метамеров. В зависимости от длины междоузлий побеги бывают удлиненными (у большинства древесных растений) и укороченными (например, плодушки у яблони). У таких травянистых растений, как одуванчик, земляника, подорожник, укроченные побеги представлены в виде прикорневой розетки.
Стеблем называют растительный орган, представляющий собой ось побега и несущий на себе листья, почки и цветки.
Основные функции стебля. Стебель выполняет опорную, проводящую и запасающую функции; кроме того, он является органом вегетативного размножения. Через стебель осуществляется связь между корнями и листьями. У некоторых растений только стебель выполняет функцию фотосинтеза (хвощ, кактус). Главная внешняя черта, отличающая побег от корня, - это наличие листьев.
Лист является плоским боковым органом, отходящим от стебля и обладающим ограниченным ростом. Основные функции листа: фотосинтез, газообмен, транспирация. Листовой пазухой является угол между листом и вышележащим участком стебля.
Почка - это зачаточный, еще не развившийся побег. В классификацию почек положены различные признаки: по
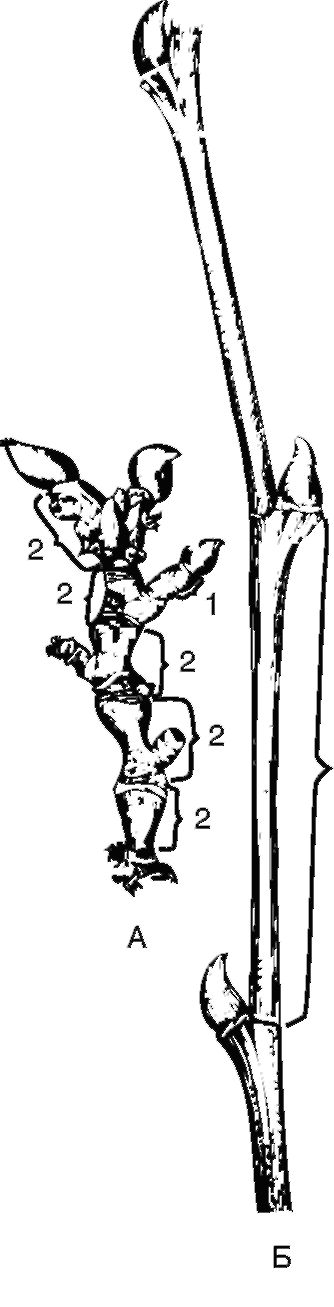
Рис. 3.11. Основные части побега: А - укороченный побег платана восточного: 1 - междоузлие; 2 - годичные приросты; Б - удлиненный побег
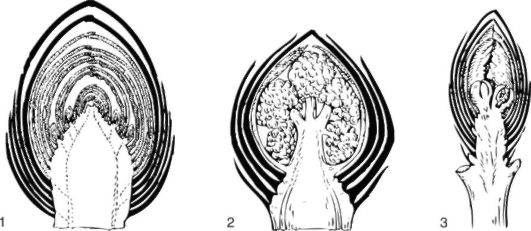
Рис. 3.12. Различные типы закрытых почек: 1 - вегетативная почка (дуб); 2 - вегетативно-генеративная почка (бузина); 3 - генеративная почка (вишня)

Рис. 3.13. Строение открытых почек: 1 - зимующие почки калины-гордовины; 2 - береза; кончик растущего побега (2а) и его верхушечная почка (2б); 3 - почка настурции; 4 - почка клевера; общий вид (4а) и схема внутреннего строения (4б); 5 - побег злака; 6 - схема продольного разреза его верхушечной почки; вегетативной (6а) и вегетативно-генеративной (6б); 7 - черемуха; кончик растущего побега
составу и функциям почки бывают вегетативными, вегетативногенеративными и генеративными.
Вегетативная почка состоит из конуса нарастания стебля, зачатков листьев, зачатков почек и почечных чешуй.
В вегетативно-генеративных почках заложен ряд метамеров, а конус нарастания трансформируется в зачаточный цветок или соцветие.
Генеративные, или цветочные, почки заключают в себе только зачаток соцветия (вишня) или одиночный цветок.
По наличию защитных чешуй почки бывают закрытые (рис. 3.12) и открытые (рис. 3.13). Закрытые почки имеют кроющие чешуи, защищающие их от иссушения и колебания температур (у большинства растений наших широт). Закрытые почки могут впадать на зиму в состояние покоя, поэтому их еще называют зимующими. Открытые почки - голые, без защитных чешуй. У них конус нарастания защи- щают зачатки срединных листьев (у крушины ломкой; древесных пород тропиков и субтропиков; водных цветковых растений). Почки, из которых весной происходит рост побегов, называются почками возобновления.
По месту расположения на стебле почки бывают верхушечные и боковые. За счет верхушечной почки осуществляется рост основного побега; за счет боковых почек - его ветвление. Если верхушечная почка отмирает, трогается в рост боковая почка. Генеративная верхушечная почка после развертывания верхушечного цветка или соцветия больше не способна к верхушечному росту.
Пазушные почки закладываются в пазухах листьев и дают боковые побеги следующего порядка. Пазушные почки имеют такое же строение, как и верхушечные. Конус нарастания представлен первичной меристемой, защищенный зачаточными листочками, в пазухах которых находятся пазушные почки. Многие пазушные почки пребывают в состоянии покоя, поэтому их еще называют спящими (или глазками). Придаточные почки развиваются обычно на корнях. У древесных и кустарниковых растений из них возникает корневая поросль.
Развертывание побега из почки. Первый побег растения формируется при прорастании семени из зародышевого побега. Это главный побег, или побег 1-го порядка. Все последующие метамеры главного побега образуются из зародышевой почки. Из боковых пазушных почек главного побега формируются боковые побеги 2-го, а в дальнейшем - и 3-го порядка. Так формируется система побегов (главного и боковых побегов 2-го и последующих порядков).
Превращение почки в побег начинается с открытия почки, появления листьев и роста междоузлий. Почечные чешуи быстро подсыхают и отпадают при начале развертывания почки. От них у основания побега часто остаются рубцы - так называемое почечное кольцо, которое хорошо заметно у многих деревьев и кустарников. По числу почечных колец можно подсчитать возраст ветви. Побеги, вырастающие из почек за один вегетационный период, называют годичными побегами, или годичным приростом.
В нарастании побега в длину и толщину участвует ряд меристем. Рост в длину происходит за счет верхушечной и вставочной меристем, а в толщину - за счет боковых меристем (камбия и феллогена). На начальных этапах развития формируется первичная анатомическая структура стебля, у однодольных растений сохраняющаяся в течение всей их жизни. У двудольных и голосеменных растений в результате деятельности вторичных образовательных тканей довольно быстро из первичной структуры формируется вторичное строение стебля.
Листорасположение - порядок размещения листьев на оси побега (рис. 3.14). Различают несколько вариантов листорасположения:
1) очередное, или спиральное - от каждого узла стебля отходит один лист (береза, дуб, яблоня, горох);
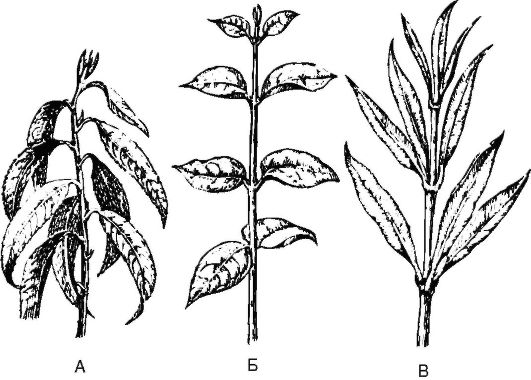
Рис. 3.14. Листорасположение: А - очередное (персик обыкновенный); Б - супротивное (бирючина овальнолистная); В - мутовчатое (олеандр)
2) супротивное - на каждом узле прикреплены друг против друга два листа (клен);
3) накрестсупротивное - разновидность супротивного, когда супротивно расположенные листья одного узла находятся во взаимно перпендикулярной плоскости другого узла (яснотковые, гвоздичные);
4) мутовчатое - от каждого узла отходят 3 листа и более (вороний глаз, ветреница).
Характер ветвления побега (рис. 3.15). Ветвление побега у растений необходимо для увеличения площади соприкосновения со средой -
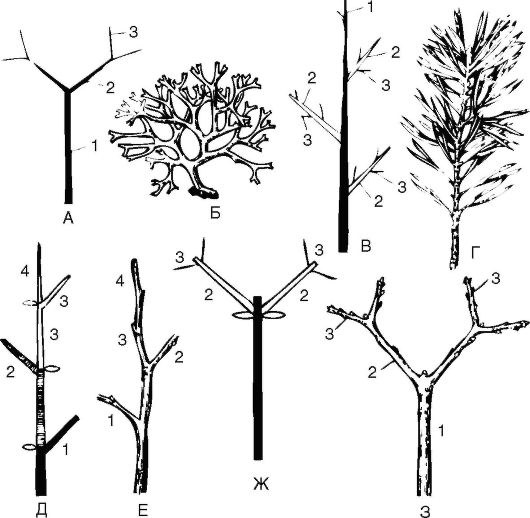
Рис. 3.15. Типы ветвления побегов: верхушечное дихотомическое: А - схема; Б - водоросль (диктиота); боковое моноподиальное: В - схема; Г - ветка сосны; боковое симподиальное по типу монохазия: Д - схема; Е - ветка черемухи; боковое симподиальное по типу дихазия: Ж - схема; З - ветка сирени; 1-4 - оси первого и последующих порядков
водной, воздушной, почвенной. Различают моноподиальное, симподиальное, ложнодихотомическое и дихотомическое ветвления побега.
1. Моноподиальное - длительное время сохраняется рост побега за счет верхушечной меристемы (у ели).
2. Симподиальное - ежегодно верхушечная почка отмирает, а рост побега продолжается за счет ближайшей боковой почки (у березы).
3. Ложнодихотомическое (при супротивном листорасположении, вариант симподиального) - верхушечная почка отмирает, а рост идет за счет 2 ближайших боковых почек, расположенных ниже апекса (у клена).
4. Дихотомическое - конус нарастания верхушечной почки (апекс) делится надвое (плаун, маршанция и т.д.).
По характеру расположения побега в пространстве различают: пря- мостоячий побег; приподнимающийся побег, который в гипокотильной части развивается в горизонтальном направлении, а в дальнейшем растет вверх как прямостоячий; стелющийся побег - растет в горизонтальном направлении, параллельно поверхности земли. Если на стелющемся стебле имеются пазушные почки, которые укореняются, побег называется ползучим (или усами). У ползучих побегов в узлах образуются придаточные корни (традесканция) или усы-столоны, заканчивающиеся прикорневой розеткой и дающие начало дочерним растениям (земляника). Вьющийся побег обвивает дополнительную опору, так как в нем плохо развиты механические ткани (вьюнок); це- пляющийся стебель растет, как и вьющийся, вокруг дополнительной опоры, но с помощью специальных приспособлений-усиков, видоизмененной части сложного листа.
Метаморфозы побегов
Видоизменение побегов произошло в процессе длительной эволюции, в результате приспособления к выполнению специальных функций. Например, корневища, клубни и луковицы, являясь запасающими побегами, часто выполняют функцию вегетативного размножения. Кроме того, видоизменения побега способны служить органом прикрепления (усики) и средством защиты (ко- лючки).
1. Подземные видоизменения побегов (рис. 3.16):
а) корневище (папоротник, ландыш) - многолетний подземный побег, имеющий редуцированные листья в виде бесцветных или бурых мелких чешуек, в пазухе которых лежат почки;
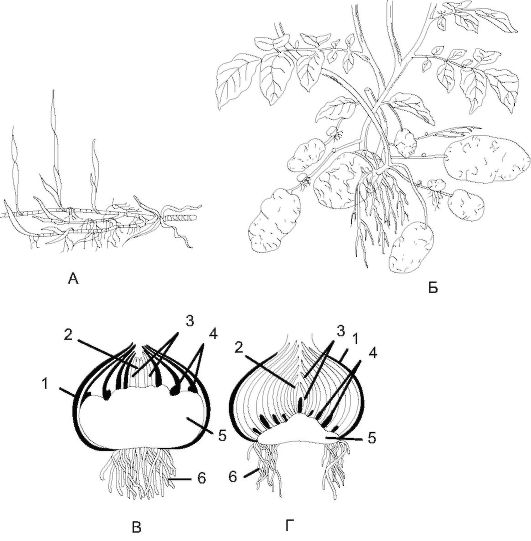
Рис. 3.16. Подземные видоизменения побегов: А - корневище; Б - клубень; В - клубнелуковица (продольный разрез); Г - луковица (продольный разрез): 1 - отмершие чешуи; 2 - зачаток цветоносного побега; 3 - листья будущего вегетативного периода; 4 - почки; 5 - укороченный стебель (у луковиц - донце); 6 - придаточные корни
б) клубень (картофель) - метаморфоз побега с ярко выраженной запасающей функцией стебля, наличием чешуевидных листьев, которые быстро сшелушиваются, и почек, формирующихся в пазухах листьев и называемых глазками. У клубня также имеются столоны - однолетние подземные недолговечные корневища, на которых образуются клубни;
в) луковица - это укороченный побег, стеблевая часть которого называется донцем. В луковице различают 2 типа видоизмененных листьев: с чешуевидными сочными основаниями, запасающими воду с растворенными в ней питательными веществами (главным образом сахарами), и сухие, покрывающие луковицу снаружи, выполняющие
защитную функцию. Из верхушечной и пазушных почек вырастают фотосинтезирующие надземные побеги, а на донце образуются при- даточные корни.
г) клубнелуковица (гладиолус) - это видоизмененная луковица с разросшимся донцем, образующим клубень, покрытый основаниями зеленых листьев. Зеленые листья высыхают и образуют пленчатые чешуи.
2. Надземные видоизменения побега (рис. 3.17).
Колючки побегового происхождения выполняют главным образом защитную функцию. Они могут образовываться за счет превращения верхушки побега в острие - колючку. У таких растений, как дикая яблоня, терн, алыча, концы ветвей оголены, заострены и превраще-
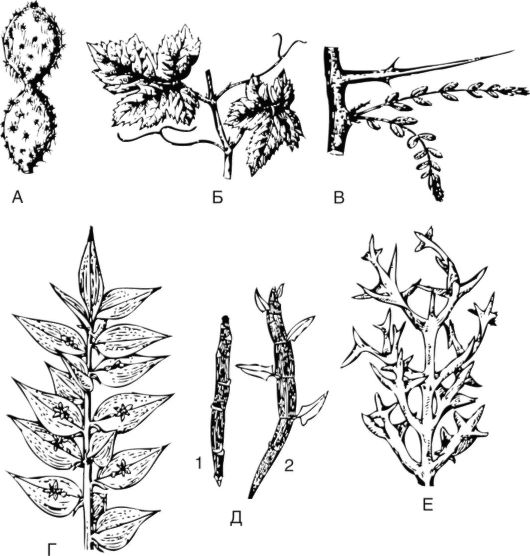
Рис. 3.17. Надземные видоизменения побега: А - мясистый побег кактуса с редуцированными листьями; Б - усыприщепки винограда (видоизмененные соцветия); В - колючка гледичии; Г - филлокладии иглицы; Д - кладодии мюленбекии (1 - нормальные; 2 - в условиях повышенной влажности); Е - кладодии коллеции
ны в колючки, торчащие во все стороны и предохраняющие плоды и листья от поедания животными. У представителей семейства Рутовые - лимона, апельсина, грейпфрута - в колючку полностью превращается специализированный боковой побег. У таких растений в пазухе листа имеется 1 крупная прочная колючка. У многих видов боярышника - многочисленные колючки, представляющие собой видоизмененные укороченные побеги, развивающие из пазушных почек нижней части однолетних побегов.
Усики характерны для растений, которые не могут самостоятельно поддерживать вертикальное (ортотропное) положение и поэтому всегда образуются в пазухе листа. Неветвящаяся, прямая часть усика представляет собой первое междоузлие пазушного побега, а закручивающаяся часть соответствует листу. У представителей семейства Тыквенные (огурец, дыня) усики простые, неветвящиеся; а у арбуза, тыквы - сложные, образующие от 2 до 5 ветвей.
Кладодии и филлокладии - видоизмененные побеги, выполняющие функцию листьев.
Кладодии - это боковые побеги, сохраняющие способность к дли- тельному росту, находящиеся на зеленых плоских длинных стеблях (у опунции).
Филлокладодии - это уплощенные боковые побеги, имеющие ограниченный рост, так как верхушечная меристема быстро дифференцируется в постоянные ткани. Побеги филлокладиев - зеленые, плоские, короткие, внешне часто напоминают листья (иглица). У представителей рода Спаржа филлокладии имеют нитевидную, линейную или игловидную форму.
Анатомия стебля
В 1924-1928 гг. немецкие ученые Дж. Будер и А. Шмидт разработали теорию туники и корпуса, отличающуюся от гистогенной теории Ганштейна (от греч. histos - ткань и genos - род, происхождение). Согласно их теории, в конусе нарастания стебля покрытосеменных выделяются 2 зоны: наружная - туника и внутренняя - корпус. Туника состоит из нескольких слоев клеток, чаще из 2, которые делятся пер- пендикулярно поверхности органа. Самый поверхностный ее слой дает начало протодерме, из которой в дальнейшем развивается эпидерма, покрывающая листья и стебли. Внутренний слой (или слои туники) образуют все ткани первичной коры. Иногда внутренние слои туники могут образовывать только наружную часть первичной коры,
в этом случае происхождение внутренней ее части связано с корпусом. Это свидетельствует об отсутствии резкой границы между туникой и корпусом. Теория туники и корпуса объясняет и формирование органов побега: листьев и пазушных почек. Так, зачатки листьев закладываются во 2-м слое туники, а пазушные почки - в корпусе.
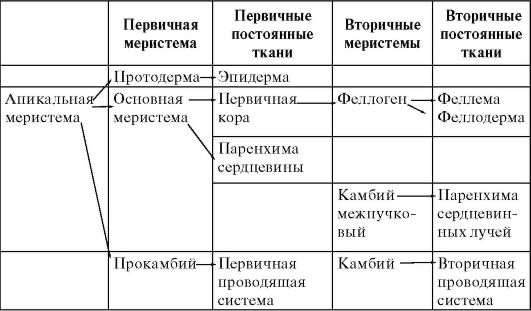
Развитие стебля осуществляется за счет дифференциации клеток туники и корпуса - первичных меристем. Из них образуются первичная покровная ткань - эпидерма, первичная кора и центральный осевой цилиндр (табл. 3.2).
Таблица 3.2. Структура меристем стебля
Формирование тканей стебля первичного строения
Первичное строение стебля формируется за счет деятельности первичных меристем апекса и включает 3 анатомо-топографические зоны: покровную ткань, первичную кору и центральный осевой цилиндр (рис. 3.18-3.20) (рис.
С поверхности стебель покрыт однослойной эпидермой, которая впоследствии покрывается кутикулой. Непосредственно под эпидермой располагается первичная кора.
Первичная кора представлена однородными клетками хлорофиллоносной паренхимы, граничащей со склеренхимой перициклического происхождения центрального осевого цилиндра (рис.
цв. вкл.). Иногда хлорофиллоносная паренхима отсутствует, и тогда перициклическая склеренхима располагается сразу под эпидермой.
Центральный осевой цилиндр начинается с перициклической склеренхимы, придающей прочность растению. Центральный осевой цилиндр пронизан изолированными сосудисто-волокнистыми пучками, которые образуются за счет деятельности прокамбия. У однодольных растений прокамбий полностью дифференцируется в первичные проводящие элементы (у двудольных растений прокамбиальные клетки в центре пучка формируют камбий). Форма пучков на поперечном срезе овальная: ближе к периферии стебля располагаются элементы первичной флоэмы, а к центру - первичной ксилемы. У стеблей однодольных формируются пучки коллатерального типа, всегда закрытые, поэтому стебель к дальнейшему утолщению не способен. Сформировавшиеся сосудисто-волокнистые пучки расположены беспорядочно. Как правило, они окружены склеренхимой, максимальное количество которой сосредоточено возле поверхности стебля. От периферии к центру стебля происходит увеличение размера пучков. Пространство между пучками занято запасающей или основной паренхимой. Клетки основной паренхимы крупные, среди них могут быть межклетники.
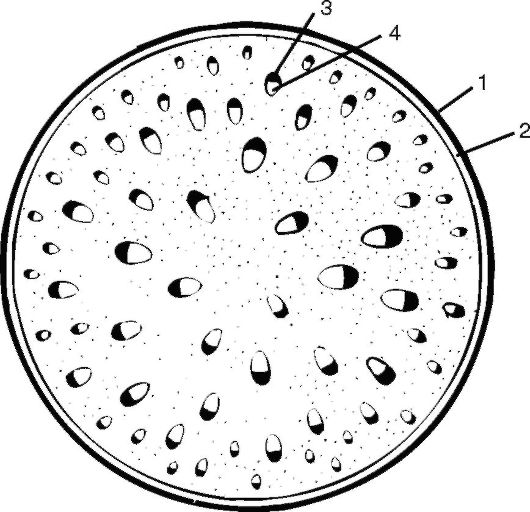
Рис. 3.18. Схема строения стебля однодольного растения (кукурузы): 1 - эпидерма; 2 - механическое кольцо; 3 - флоэма; 4 - ксилема
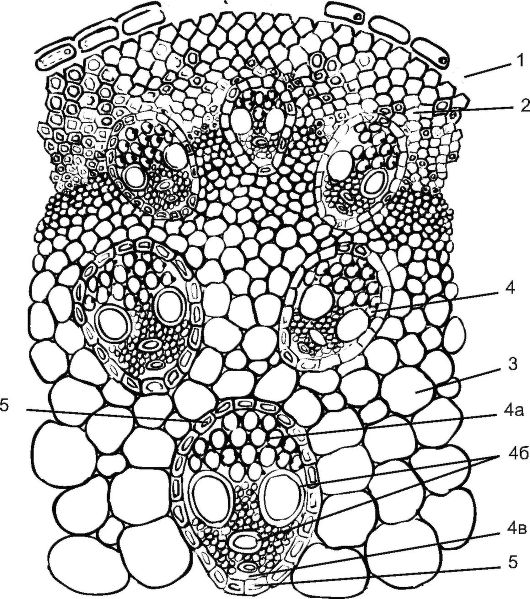
Рис. 3.19. Поперечный срез стебля кукурузы: 1 - эпидерма; 2 - склеренхима; 3 - основная паренхима; 4 - закрытый коллатеральный пучок: 4а - флоэма, 4б - сосуды ксилемы, 4в - воздухоносная полость; 5 - склеренхимная обкладка пучка
Для однодольных растений в отличие от двудольных нехарактерно наличие сердцевины в центре стебля, хотя может быть развита центральная воздушная полость (например, у стеблей злаков - со- ломина). Соломина (рис. 3.21 и 3.22) - это особый тип стебля с полыми междоузлиями и узлами между ними. В зрелой соломине ржи, пшеницы и других злаков эпидерма и хлорофиллоносная паренхима, утратившая хлоропласты, подвергаются одревеснению (рис. 14,
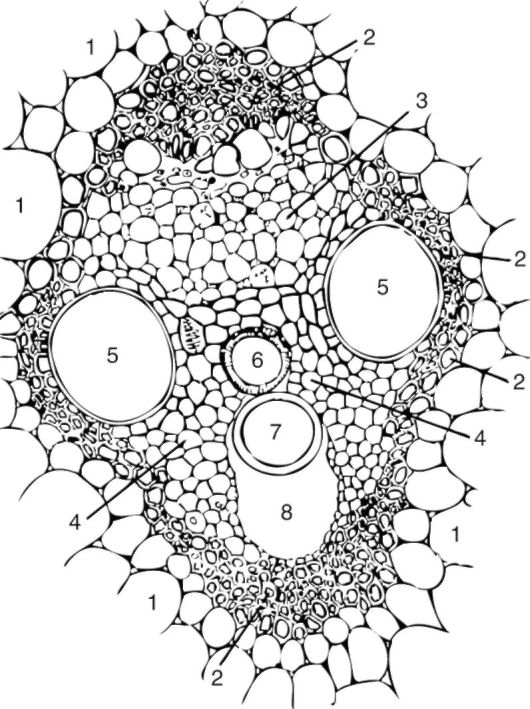
Рис. 3.20. Закрытый сосудисто-волокнистый пучок кукурузы (поперечный срез): 1 - тонкостенная паренхима стебля; 2 - склеренхима; 3 - луб (флоэма); 4 - древесинная паренхима; 5 - сетчатые сосуды; 6 - кольчато- спиральный сосуд; 7 - кольчатый сосуд; 8 - воздушная полость
дания механической прочности стеблю, который приобретает в этот период вместо зеленой окраски желтую. Пучки располагаются в 2 слоя в шахматном порядке и окружены склеренхимой. Внутренние пучки более крупные, наружные - мельче, их склеренхимная обкладка сливается с перициклической склеренхимой, образуя кольцо механической ткани.
Особенности строения стебля однодольных:
1) сохранение первичного строения в течение всей жизни;
2) слабо выраженная первичная кора;
3) разбросанное расположение сосудисто-волокнистых пучков;
4) коллатеральные пучки только закрытого типа (без камбия);
5) наличие во флоэме только проводящих элементов - ситовидных трубок с клетками-спутницами;
6) отсутствие сердцевины;
7) вторичное утолщение стеблей однодольных.
Вторичное утолщение стеблей древесных однодольных растений осуществляется за счет кольца утолщения (оно представляет собой особый валик вокруг конуса нарастания), дающего дополнительный
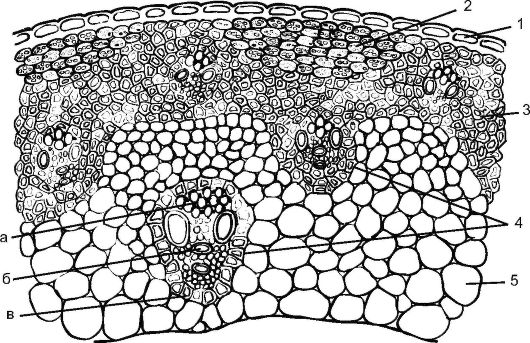
Рис. 3.21. Схема строения соломины ржи: 1 - эпидерма; 2 - хлорофиллоносная ткань; 3 - склеренхима; 4 - закрытые коллатеральные сосудисто-волокнистые пучки; а - флоэрма; б - ксилема; в - склеренхимная обкладка пучка; 5 - основная паренхима
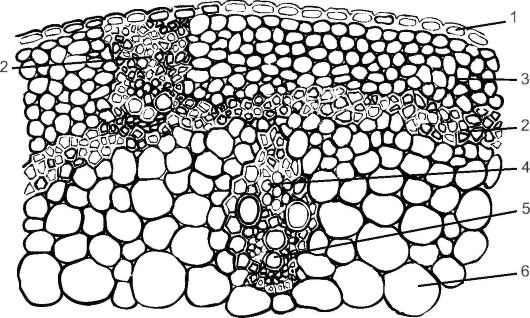
Рис. 3.22. Строение соломины пшеницы: 1 - эпидерма; 2 - склеренхима; 3 - хлоренхима; 4 - флоэма; 5 - ксилема; 6 - основная паренхима
ряд сосудисто-волокнистых пучков. Подобное утолщение наблюдается у таких однодольных, как пальмы, бананы, алоэ.
Особенности строения корневищ однодольных. Корневища, являясь подземным видоизменением побега, в своем анатомическом строении сохраняют характерные черты стеблей и приобретают некоторые особенности, связанные с подземным существованием.
Покровной тканью остается эпидерма, часто одревесневшая. Первичная кора значительно шире и представлена запасающей паренхимой. Во внутреннем слое первичной коры, примыкающем к центральному осевому цилиндру, формируется однослойная эндодерма (подковообразная или с пятнами Каспари). Изредка (например, в корневище ландыша) она бывает двухслойной.
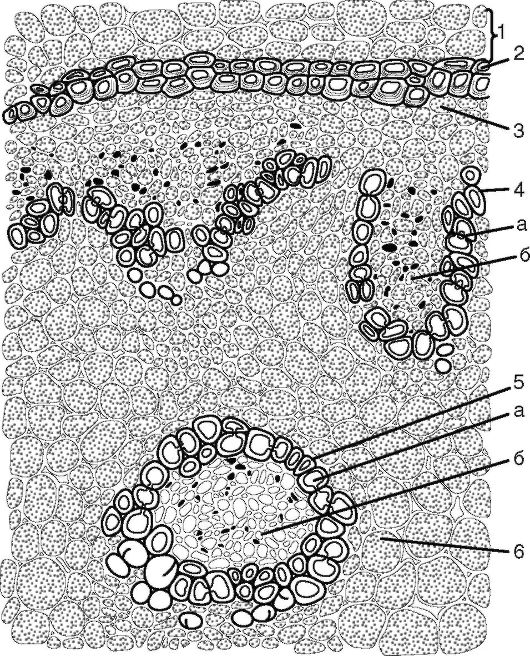
Рис. 3.23. Часть центрального цилиндра корневища ландыша: 1 - паренхима первичной коры; 2 - эндодерма с подковообразными утолщениями; 3 - перицикл; 4 - закрытый коллатеральный пучок; 5 - концентрический пучок; а - ксилема; б - флоэма; 6 - паренхима
Центральный осевой цилиндр начинается с живого перицикла. Его роль в подземных побегах - формирование придаточных кор- ней. Пучки бывают 2 типов: закрытые коллатеральные и концентри- ческие, расположенные также беспорядочно в центральном цилиндре (рис. 3.23) (рис.
Формирование тканей стебля вторичного строения
Вторичное строение стебля характерно для однолетних и многолетних травянистых, древесных двудольных, а также голосеменных растений. У двудольных растений первичное строение очень недолговечно, и с началом деятельности камбия образуется вторичная структура. В зависимости от закладки прокамбия формируется несколько типов вторичного строения стебля. Если тяжи прокамбия разделены широкими рядами паренхимы, то формируется пучковое строение, если они сближены так, что сливаются в цилиндр, - формируется непучковое строение.
Пучковое строение стебля встречается у таких растений, как клевер, горох, лютик, укроп (рис. 3.24). У них прокамбиальные тяжи закладываются в один круг по периферии центрального цилиндра. Каждый
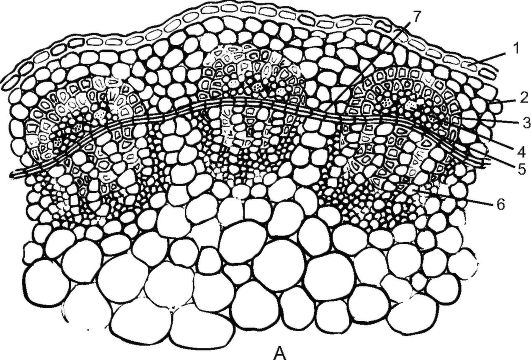
Рис. 3.24. Пучковый тип строения стебля двудольного растения: А - клевер: 1 - эпидерма; 2 - хлоренхима; 3 - склеренхима перициклического происхождения; 4 - флоэма; 5 - пучковый камбий; 6 - ксилема; 7 - межпучковый камбий
прокамбиальный тяж превращается в коллатеральный пучок, состоящий из первичной флоэмы и первичной ксилемы. В дальнейшем между флоэмой и ксилемой из прокамбия закладывается камбий, формирующий элементы вторичной флоэмы и вторичной ксилемы. К периферии органа откладывается флоэма, а к центру - ксилема, причем ксилемы откладывается больше. Первичные флоэма и ксилема остаются на периферии пучка, а вторичные элементы примыкают к камбию. Для стеблей двудольных растений характерно формирование открытых колла- теральных или биколлатеральных пучков (рис.
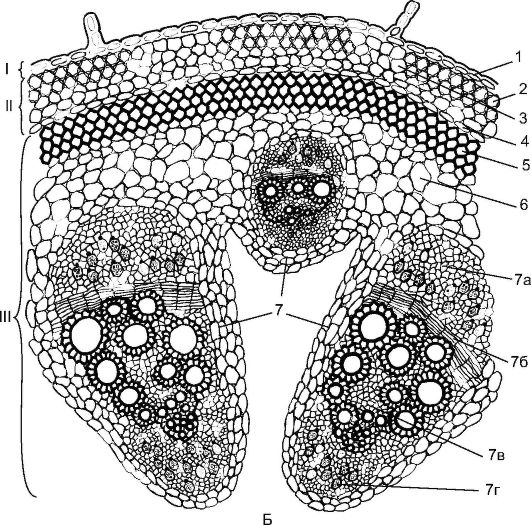
Рис. 3.24. (продолжение) Б - тыква: I - покровная ткань; II - первичная кора; III - центральный осевой цилиндр; 1 - эпидерма; 2 - уголковая колленхима; 3 - хлоренхима; 4 - эндодерма; 5 - склеренхима; 6 - основная паренхима; 7 - биколлатеральный сосудисто-волокнистый пучок: 7а - флоэма; 7б - камбий; 7в - ксилема; 7г - внутренняя флоэма
Также для стеблей двудольных растений характерна дифференциация первичной коры, в состав которой входят: колленхима (уголковая (рис.
Непучковое строение характерно для древесных растений (липа) (рис.
Молодой побег липы, образовавшийся весной из почки, покрыт эпидермой. Все ткани, лежащие до камбия, называют корой. Кора бывает первичная и вторичная. Первичная кора представлена пластинчатой колленхимой, располагающейся сразу под эпидермой сплошным кольцом, хлорофиллоносной паренхимой и однорядным крахмалоносным влагалищем. В этом слое находятся зерна «оберегаемого» крахмала, которые растение не расходует. Считается, что этот крахмал участвует в поддержании растением равновесия.
Центральный осевой цилиндр у липы начинается с перициклической склеренхимы над участками флоэмы. В результате деятельности камбия возникает вторичная кора (от камбия до перидермы), пред- ставленная вторичной флоэмой, сердцевинными лучами и паренхимой вторичной коры. Кору с липы заготавливают, снимая до камбия, особенно легко это делать весной, когда клетки камбия активно делятся. Раньше кору липы (лыко) использовали для плетения лаптей, изготовления коробов, мочалок и др.
Трапециевидная флоэма разделена треугольными первичными сердцевинными лучами, пронизывающими древесину до сердцевины. Состав флоэмы у липы неоднороден. В ней имеются составляющие твердый луб одревесневшие лубяные волокна, а мягкий луб
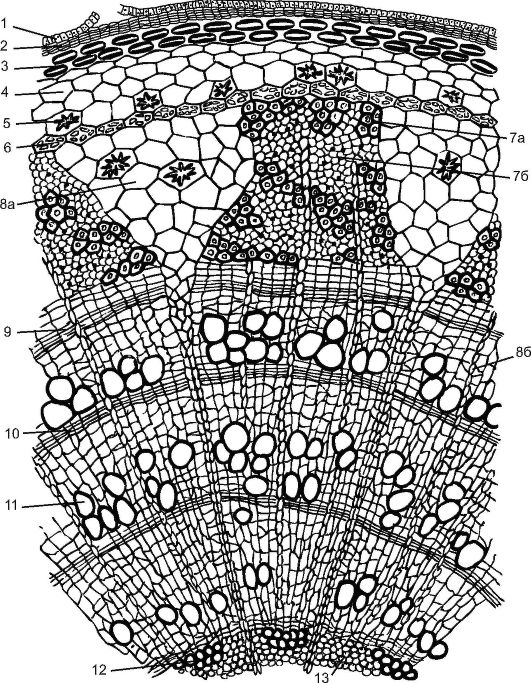
Рис. 3.25. Поперечный разрез трехлетней ветви липы: 1 - остатки эпидермы; 2 - пробка; 3 - пластинчатая колленхима; 4 - хлоренхима; 5 - друзы; 6 - эндодерма; 7 - флоэма: 7а - твердый луб, (лубяные волокна); 7б - мягкий луб - (ситовидные трубки с клетками-спутницами и лубяная паренхима); 8а - первичный сердцевинный луч; 8б - вторичный сердцевинный луч; 9 - камбий; 10 - осенняя древесина; 11 - весенняя древесина; 12 - первичная ксилема; 13 - паренхима сердцевины
представлен ситовидными трубками с клетками-спутницами и лубяной паренхимой. Луб теряет способность проводить органические вещества обычно через год и обновляется новыми слоями за счет деятельности камбия.
Камбий образует и вторичные сердцевинные лучи, но они не доходят до сердцевины, теряясь во вторичной древесине. Сердцевинные лучи служат для продвижения воды и органических веществ в ра- диальном направлении. В паренхимных клетках сердцевинных лучей к осени откладываются запасные питательные вещества (крахмал, масла), расходуемые весной на рост молодых побегов.
Уже летом под эпидермой закладывается феллоген и формируется вторичная покровная ткань - перидерма. К осени, с образованием перидермы, клетки эпидермы отмирают, но их остатки сохраняются в течение 2-3 лет. Наслоение многолетних перидерм формирует корку.
Слой ксилемы, образуемый камбием, у древесных растений значительно шире, чем слой флоэмы. Древесина функционирует в тече- ние нескольких лет. Отмершие же клетки древесины не участвуют в проведении веществ, но способны поддерживать колоссальную тяжесть кроны растения.
Состав древесины неоднороден, в нее входят: трахеиды (рис.
Особенности строения стебля двудольных:
1) рост стебля в толщину (за счет деятельности камбия);
2) хорошо дифференцированная первичная кора (колленхима, хлорофиллоносная паренхима, крахмалоносная эндодерма);
3) биколлатеральные и коллатеральные пучки только открытого типа (с камбием);
4) сосудисто-волокнистые пучки расположены по кольцу или сливаются (непучковое строение);
5) наличие сердцевины;
6) для древесных растений характерно наличие в ксилеме годичных колец.
Особенности строения корневищ двудольных. Покровной тканью корневищ двудольных может быть эпидерма, а у многолетних корневищ эпидерма сменяется перидермой. Первичная кора представлена запасающей паренхимой и эндодермой с пятнами Каспари. Причем ширина первичной коры приближается к ширине центрального цилиндра. Строение центрального осевого цилиндра, сосудисто-волокнистых пучков и их расположение в нем имеют те же особенности, что и для надземных стеблей.
ЛИСТ - БОКОВОЙ ОРГАН ПОБЕГА
Общая характеристика листа
Лист - уплощенный боковой орган побега с билатеральной симметрией; он закладывается в виде листового бугорка, представляющего собой боковой выступ побега. Лист имеет одну плоскость симметрии и характерную плоскую форму.
Зачаток листа увеличивается в длину за счет роста верхушки, в ширину - за счет краевого роста. У семенных растений верхушечный рост быстро прекращается. После развертывания почки происходят многократное деление всех клеток листа (у двудольных) и увеличение их размеров. После дифференциации клеток меристемы в постоянные ткани лист нарастает за счет интеркалярной меристемы основания листа. У большинства растений деятельность этой меристемы быстро заканчивается, и лишь у немногих (таких, как кливия, амариллис) продолжается достаточно долго.
У однолетних травянистых растений продолжительность жизни стебля и листа практически одинаковая - 45-120 дней, у вечнозеленых - 1-5 лет, у хвойных (таких, как пихта) - до 10 лет.
Первыми листьями семенных растений являются семядоли зародыша. Следующие (настоящие) листья формируются в виде меристе- матических бугорков - примордиев, возникающих из верхушечной меристемы побега.
Основными функциями листа являются фотосинтез, транспирация и газообмен.
Основные части листа (рис. 3.26):
• листовая пластинка;
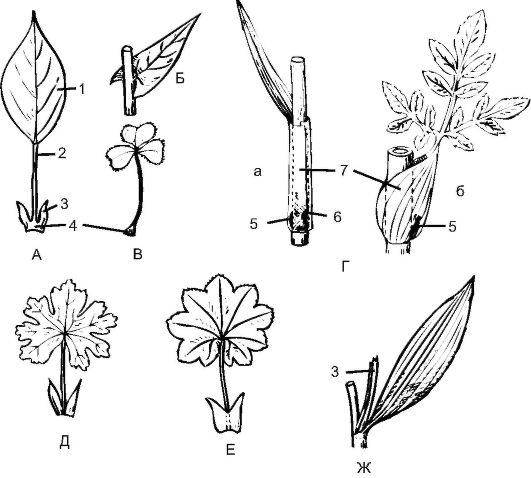
Рис. 3.26. Части листа (схема): А - черешковый; Б - сидячий; В - с подушечкой в основании; Г (а и б) - с влагалищем; Д - со свободными прилистниками; Е - с приросшими прилистниками; Ж - с пазушными прилистниками; 1 - пластинка; 2 - черешок; 3 - прилистники; 4 - основание; 5 - пазушная почка; 6 - интеркалярная меристема; 7 - влагалище
• черешок;
• основание листа;
• прилистники - выросты из основания листа.
Листовая пластинка - основная, наиболее важная фотосинтезирующая часть листа.
Черешки ориентируют листовые пластинки по отношению к источнику света, создавая листовую мозаику, т.е. такое размещение листьев на побеге, при котором они не затеняют друг друга. Это достигается за счет: различной длины и изогнутости черешка; различной величины и формы листовой пластинки; вследствие светочувствительности листьев. Если черешок отсутствует, лист называется сидячим; тогда он прикрепляется к стеблю основанием листовой пластинки.
Основание - это базальная часть листа, сочлененная со стеблем. Если основание листа разрастается, образуется листовое влагалище (семейства Злаковые, Лилейные, Зонтичные). Влагалище защищает пазушные почки и основания междоузлий.
Прилистники - парные боковые выросты основания листа. Они прикрывают боковые почки и предохраняют их от различных повреждений. В почке прилистники обязательно закладываются вместе с листьями, однако у многих растений быстро опадают или пребывают в зачаточном состоянии. Если прилистники срастаются, образуется раструб (например, в семействе гречишные).
Жилкование
Жилка листа представлена сосудисто-волокнистым пучком и выполняет проводящую и механическую функции. Жилки, входящие в лист от стебля через основание и черешок, называют главными. От главных жилок отходят боковые жилки 1-го, 2-го и т.д. порядка. Между собой жилки могут соединяться сетью мелких жилоканастомозов.
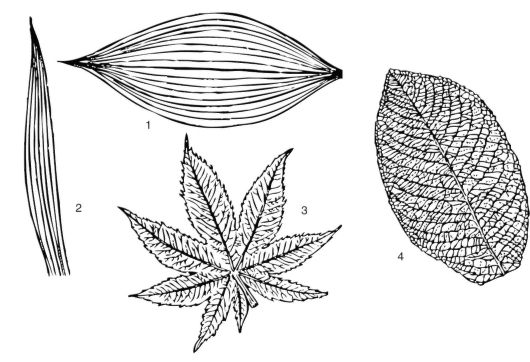
Рис. 3.27. Типы жилкования: 1 - дуговое; 2 - параллельное; 3 - пальчатое; 4 - перистое
Дуговое и параллельное жилкование чаще встречается у однодольных растений. При дуговом жилковании неветвящиеся жилки расположены дугообразно и сходятся на верхушке и к основанию листовой пластинки (ландыш). При параллельном жилковании жилки листовой пластинки проходят паралелльно друг другу (злаки, осоки).
Пальчатое жилкование - из черешка в листовую пластинку входит несколько главных жилок 1-го порядка (в виде пальцев руки). От главных жилок отходят жилки последующих порядков (у двудольных растений - например, клена татарского).
Перистое жилкование - выражена центральная жилка, идущая от черешка и сильно ветвящаяся в листовой пластинке в виде пера (характерно для двудольных растений - например, листа черемухи обыкновенной) (рис. 3.27).
Классификация листьев
Лист, состоящий из одной листовой пластинки, называется простым. Такие листья опадают в месте сочленения стебля с черешком у деревьев и кустарников, где возникает разделительный слой. Лист называется сложным, если на общей оси, называемой рахисом (от греч. rhachis - хребет), располагаются несколько листовых пластинок (листочков), имеющих свои черешочки. При листопаде у сложного листа сначала опадают листочки, а затем рахис (семейства Бобовые, Розоцветные).
Простые листья подразделяются на листья с цельной и расчлененной листовой пластинкой.
Простые листья с цельной листовой пластинкой характеризуются рядом признаков (рис. 3.28):
а) формой листовой пластинки (округлая, яйцевидная, продолговатая и т.д.);
б) формой основания листа (сердцевидное, копьевидное, стреловидное и т.д.);
в) формой края листовой пластинки (зубчатый, пильчатый, выямчатый и т.д.).
Простые листья с расчлененной листовой пластинкой в зависимости от жилкования (пальчатое или перистое) и степени глубины расчленения подразделяют на:
а) пальчатолопастные, или перистолопастные - если расчленение листовой пластинки доходит до 1/3 ширины пластинки или полупластинки;
Рис. 3.28. Простые листья с цельной листовой пластинкой
б) пальчатораздельные, или перистораздельные - если расчленение листовой пластинки доходит до 1/2 ширины пластинки или полупластинки;
в) пальчаторассеченные, или перисторассеченные - если степень расчленения листовой пластинки доходит до ее основания или центральной жилки (рис. 3.29).
Сложные листья бывают тройчатосложные, состоящие из 3 листочков (земляника), и пальчатосложные, состоящие из множества листочков (каштан). У этих типов сложных листьев все листочки прикрепляются к верхушке рахиса.
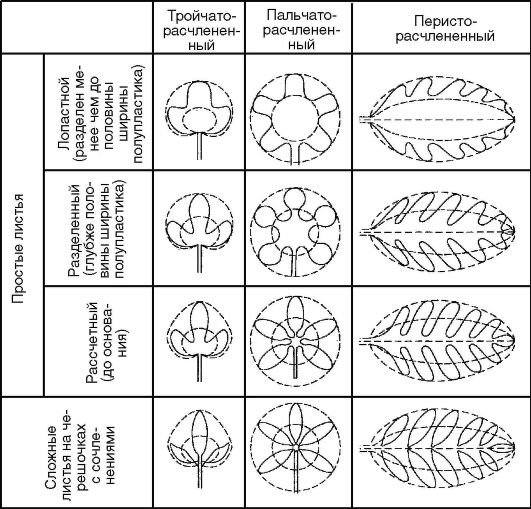
Рис. 3.29. Сложные и простые листья с расчлененной листовой пластинкой
Кроме того, есть сложные листья, листочки которых располагаются по всей длине рахиса. Среди них различают парноперистосложные, если они заканчиваются на верхушке листовой пластины парой листочков (горох посевной), и непарноперистосложные (рябина обыкновенная), заканчивающиеся одним листочком (см. рис. 3.25).
Анатомическое строение листовой пластинки
Клетки меристемы зачатка листа дифференцируются в первичную покровную ткань - эпидерму, основную паренхиму и механические ткани. Слои прокамбия, возникшие из срединного меристе-
матического слоя зачатка листа, дифференцируются в проводящие пучки.
По анатомическому строению выделяют дорсовентральные, изолатеральные и радиальные листья.
При равномерном освещении листа с обеих сторон, когда листовая пластинка расположена почти вертикально (под острым углом к стеблю), лист становится изолатеральным, т.е. равносторонним. При такой структуре листа столбчатая хлоренхима располагается с верхней и нижней стороны (например, у листа гладиолуса, нарцисса, ириса; рис.
У большинства растений вследствие неравномерного освещения листа с верхней и нижней стороны на верхней стороне листовой пластинки происходит развитие столбчатой хлоренхимы, а на нижней - губчатой. Такая структура называется дорсовентральной, т.е. ярко вы- раженная дорсальная и вентральная сторона (свекла сахарная).
У хвои сосны ассимиляционная часть листа представлена складчатой хлоренхимой, расположенной вокруг центрального осевого цилиндра. Структура таких листьев называется радиальной.
Рассмотрим анатомическое строение листа дорсовентральной структуры (рис. 3.30 и 3.31).
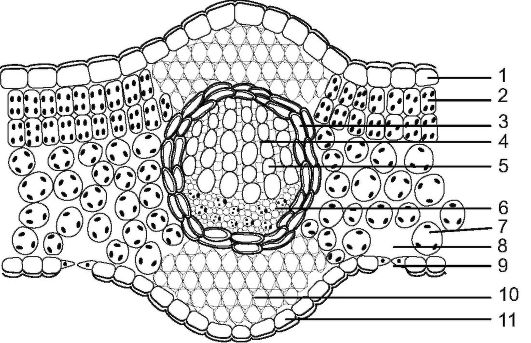
Рис. 3.30. Схема строения дорсовентрального листа: 1 - верхняя эпидерма; 2 - столбчатая хлоренхима; 3 - склеренхима; 4 - сердцевинные лучи ксилемы; 5 - сосуды ксилемы; 6 - флоэма; 7 - губчатая хлоренхима; 8 - воздухоносная полость; 9 - устьице; 10 - колленхима; 11 - нижняя эпидерма
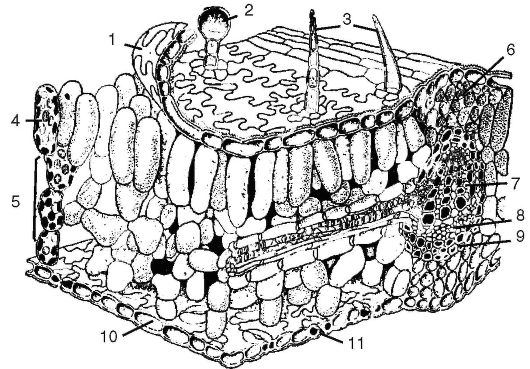
Рис. 3.31. Полусхематическое объемное изображение части листовой
пластинки:
1 - верхняя эпидерма; 2 - железистый волосок; 3 - кроющий волосок; 4 - палисадный (столбчатый) мезофилл; 5 - губчатый мезофилл; 6 - колленхима; 7 - ксилема; 8 - флоэма; 9 - обкладочная склеренхима пучка; 10 - нижняя эпидерма; 11 - устьице
Сверху и снизу лист покрыт живой однослойной эпидермой. Причем верхняя по сравнению с нижней эпидермой представлена более крупными клетками и покрыта кутикулой. Часто верхняя эпидерма пропитывается воском, что усиливает защитную функцию листа от потери воды. Эти клетки плотно сомкнуты, чему способствуют их извилистые очертания. Клетки эпидермы играют роль в образовании трихом. Трихомы могут быть различной формы: одноклеточные, многоклеточные ветвистые, щетинки, звездчатые (см. раздел «Покровные ткани»). В клетках трихом протопласт отмирает, содержимое заполняется воздухом; основная их функция - защитная (от потери воды, перегрева, поедания животными).
В эпидерме располагаются устьица. Чаще они встречаются в нижней эпидерме, но могут быть и с обеих сторон, а у водных рас- тений с плавающими листьями только на верхней эпидерме. Если у двудольных растений устьица расположены достаточно свободно по всей эпидерме, то у однодольных с вытянутыми листьями - ровны-
ми рядами, причем устьичные щели ориентированы вдоль оси листа. Устьицам всегда сопутствуют воздухоносные полости, через которые осуществляются транспирация и газообмен.
Под верхней эпидермой в 1-3 слоя размещается столбчатый мезофилл (столбчатая хлоренхима). Клетки его - цилиндрической формы, узкая их сторона примыкает к эпидерме. Это высокоспециализированная ткань для выполнения фотосинтеза.
Прямоугольная (цилиндрическая) форма клеток обеспечивает сохранность хлорофилла, содержащегося в хлоропластах. Находясь большую часть времени на вытянутых радиальных стенках, чечевицеобразные хлоропласты не подвергаются воздействию прямых солнечных лучей. Последние скользят вдоль них, равномерно освещая хлоропласты и не разрушая при этом хлорофилл. Все это способствует активному протеканию фотосинтеза.
Ниже лежит губчатый мезофилл, характеризующийся рыхло расположенными округлыми клетками с большими межклетниками. Губчатый мезофилл, как и столбчатый, содержит хлоропласты, но их в 2-6 раз меньше, чем в столбчатой хлоренхиме. Основные функции губчатой ткани - транспирация и газообмен, хотя она участвует и в фотосинтезе.
Крупные жилки листа представлены полным сосудисто-волокнистым пучком, а мелкие - неполным. Вверху полного сосудисто- волокнистого пучка располагаются ксилема, а под ней флоэма. Как правило, это закрытые пучки, но у некоторых двудольных видны следы деятельности камбия, которая прекращается рано.
У двудольных также вокруг пучка кольцом лежит склеренхима, оберегающая пучок от давления разрастающихся клеток мезофилла листа. Над пучком и под ним располагается уголковая, реже - пластинчатая колленхима, примыкающая к эпидерме и выполняющая опорную функцию. Мелкие жилки проходят в толще мезофилла под столбчатой хлоренхимой. Склеренхима может располагаться участками или вокруг этих жилок.
Своеобразное строение имеют листья хвойных растений; рассмотрим это строение на примере хвои сосны (рис. 3.32).
Клетки эпидермы толстостенные, одревесневшие, почти квадратной формы, покрыты толстым слоем кутикулы. Под эпидермой располагается гиподерма в один слой, а по углам - несколькими слоями. Клетки гиподермы со временем одревесневают и выполняют водозапасающую и механическую функции. С обеих сторон листа имеются устьица, под которыми лежат большие воздухонос-
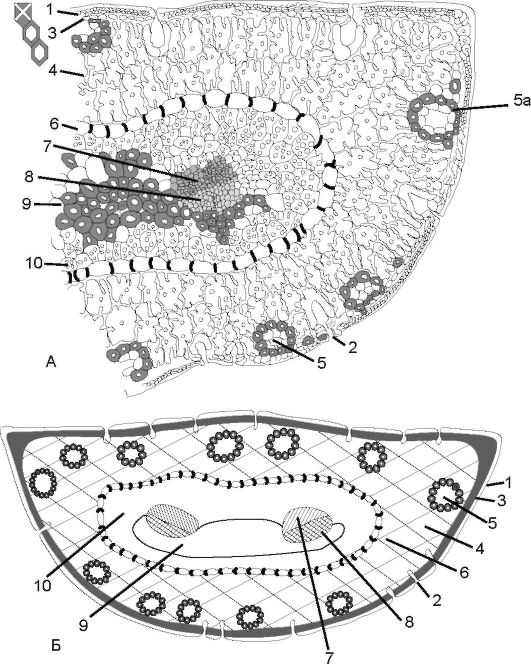
Рис. 3.32. Лист (хвоя) сосны в поперечном разрезе (А) и схематическое
изображение (Б):
1 - эпидерма; 2 - устьичный аппарат; 3 - гиподерма; 4 - складчатая паренхима; 5 - смоляной ход; 5а - склеренхимная обкладка; 6 - эндодерма с пятнами Каспари; 7 - ксилема; 8 - флоэма; 7, 8 - закрытый проводящий пучок; 9 - склеренхима; 10 - паренхима (трансфузионная ткань)
ные полости. Под гиподермой находится мезофилл, представленный клетками, имеющими внутренние складки, увеличивающие их ассимилирующую поверхность. В складчатой хлоренхиме проходят смоляные ходы.
Центральный осевой цилиндр отделяется от складчатой хлоренхимы эндодермой с пятнами Каспари. Проводящая система
представлена 2 пучками, снизу обрамлена тяжами склеренхимы. Остальное пространство занято трансфузионной тканью, осуществляющей связь пучков с мезофиллом. Трансфузионная ткань состоит из мертвых и живых клеток. По рядам живых клеток передаются ассимиляты во флоэму, а по мертвым клеткам - вода из ксилемы к хлоренхиме.
Листопад
Листопад - это биологическое явление, обусловленное жизнедеятельностью растения. Лист, достигший предельных размеров, довольно быстро начинает стареть и отмирать. При старении листа в нем замедляются жизненно важные процессы: дыхание, фотосинтез. Начинают преобладать процессы распада, а не синтеза, и из листа начинают оттекать органические вещества (углеводы, аминокислоты). Лист опустошается от питательных веществ, но в нем начинают накапливаться балластные вещества - такие, как соли оксалата кальция. Видимый признак старения листа - изменение его окраски. С разрушением хлорофилла и накоплением каротиноидов и антоцианов лист приобретает желтый, оранжевый или багряный цвет. Образованию антоцианов способствуют низкая температура, солнечная погода, высокое содержание сахара в клетках мезофилла. Во время дождливой, пасмурной осени листья, как правило, желтые, а не багряные, и дольше остаются на деревьях. У травянистых растений лист разрушается, но остается на стебле, у деревьев и кустарников старые листья опадают - таким образом растения реагируют на уменьшающийся световой день, понижение температуры. Это связано с тем, что в конце лета в месте прикрепления листа к стеблю образуется отделительный пробковый слой, изолирующий лист от стебля. При порывах ветра и под собственной тяжестью лист отделяется от стеб- ля по разделительному (пробковому) слою. На этом месте остается листовой рубец; он покрыт пробкой, которая защищает ткани стебля в месте, где крепился лист.
Опадание листьев может происходить и летом - для предотвращения растением физиологической засухи, так как оставшиеся ли- стья испаряли бы воду, которая не может в это время в достаточном количестве поступить в корни.
Кроме листопадных растений есть вечнозеленые, которые в течение всего года имеют зеленые листья, но они также после истечения жизненного срока (несколько лет) опадают.

Рис. 3.33. Гомологичные органы листового происхождения: А - ловчий аппарат непентеса; Б - колючки акации белой; В - колючки барбариса; Г - усик чины
Метаморфозы листа
Усики. У многих лазающих растений (таких, как диоскорея, настурция) часть листа или весь лист превращаются в усики. У многих представителей Бобовых (горох посевной, чечевица) усиками становятся верхняя часть рахиса и несколько пар листочков.
Колючки - это приспособления, уменьшающие испарение влаги и защищающие от поедания животными. Лист может полностью преобразоваться в колючку (например, у кактусов). У некоторых растений (акации, робинии, молочаи) колючки образуются из прилистников, после опадания листьев.
Филлодий - это метаморфоз черешка (у некоторых видов чин Кав- каза) или основания листа в образование, похожее на плоский лист. Филлодии выполняют функцию фотосинтеза и характерны для растений, обитающих в засушливом климате.
Ловчие аппараты насекомоядных растений - это видоизмененные листья. Эти растения автотрофные, но наряду с этим они способны переваривать животных и добывать готовые органические вещества. Например, росянка, обитающая на торфяных болотах, имеет ловчий аппарат в виде пурпурной ножки - вырост пластинки листа и овальной головки - желёзки, выделяющей секрет с кислотой и ферментом, схожим с пепсином (рис. 3.33).