Ботаника: учебник. Зайчикова С.Г., Барабанов Е.И. 2013. - 288 с. : ил.
|
|
|
|
ГЛАВА I ЦИТОЛОГИЯ
ИСТОРИЯ РАЗВИТИЯ КЛЕТКИ
Клетка - основная структурно-функциональная единица животных, растений и грибов. Понятие о клетке и ее строении возникло в связи с изобретением микроскопа в 1590 г. голландскими мастерами братьями Янсен. Впервые увидел и описал клетку английский естествоиспытатель Роберт Гук в 1665 г. Рассматривая в микроскоп тонкий срез бутылочной пробки, он обнаружил, что пробка состоит из многочисленных камер, и назвал их клетками. М. Мальпиги (1671) и Н. Грю (1682) первыми описали микроскопическое строение органов растений, подтвердив их клеточное строение. В 1676 г. А. Левенгук открыл мир микроскопических растений и описал окрашенные включения в клетках высших растений и водорослей. До XIX века существовало представление, что основные функции клетки связаны с ее стенкой, а содержимому клетки отводилась второстепенная роль. С усовершенствованием микротехники расширялись и познания о внутреннем содержимом клетки. Так, в 1831 г. Р. Браун обнаружил в клетке ядро и описал его как важнейшее образование. В 1839 г. Я. Пуркинье ввел новый термин «протоплазма», т.е. живое содержимое клетки.
Обобщив все накопленные знания в области клетки, ботаник М. Шлейден (1838) и зоолог Т. Шванн (1839) сформулировали кле- точную теорию, основным утверждением которой было: клетка есть единая элементарная и функциональная структура всех живых организмов.
В 1858 г. Р. Вирхов добавил новое положение к клеточной теории, обосновав принцип преемственности клеток путем деления (каждая клетка образуется из клетки).
В 1946 г. с применением электронного микроскопа была установлена тонкая структура клетки, получившая название ультраструктуры.
СТРОЕНИЕ РАСТИТЕЛЬНОЙ КЛЕТКИ
Все растения являются эукариотами, так как имеют оформленное ядро (греч. karyon - ядро). Более примитивные организмы - бактерии и, в частности, цианобактерии (сине-зеленые водоросли), являющиеся прокариотами (доядерными организмами), отличаются по ряду признаков от эукариот (табл. 1.1).
Таблица 1.1. Основные особенности прокариот и эукариот
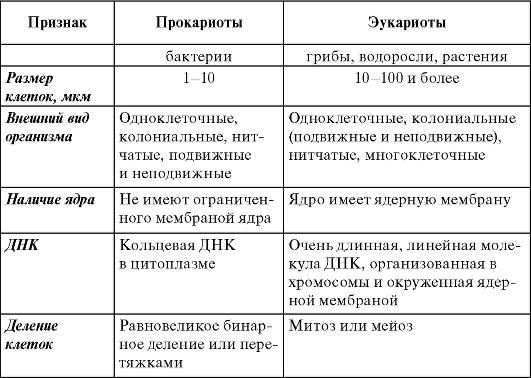
Окончание таблицы
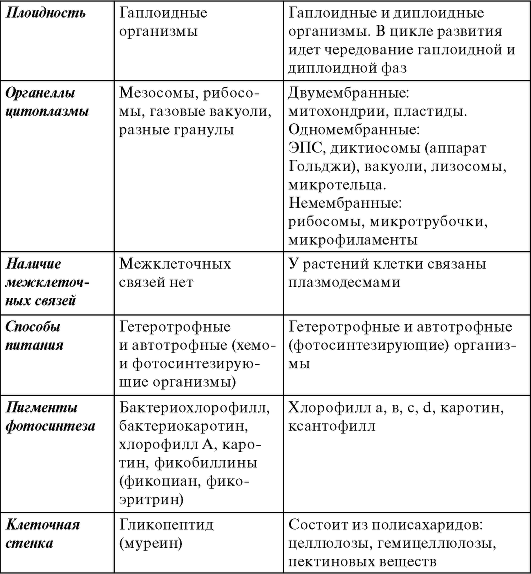
Примечание. ДНК - дезоксирибонуклеиновая кислота, ЭПС - эндоплазматическая сеть.
ПРОТОПЛАСТ
Во взрослой растительной клетке (рис. 1.1) выделяют: протопласт - живое содержимое клетки и производные протопласта. Протопласт представляет собой цитоплазму и ядро; к производным протопласта относят целлюлозную клеточную стенку (оболочку) и вакуоль.
В состав цитоплазмы входит гиалоплазма - внутренняя жидкая среда клетки, в которую погружены клеточные органеллы. Для живых растительных клеток характерно движение цитоплазмы вме-
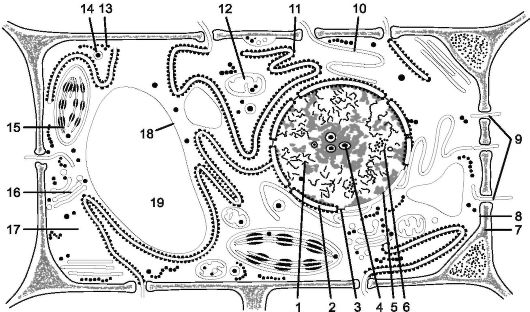
Рис. 1.1. Схема строения растительной клетки (электронная микроскопия):
1 - ядро; 2 - ядерная оболочка (две мембраны - внутренняя и внешняя и перинуклеарное пространство); 3 - ядерная пора; 4 - ядрышко; 5 - конденсированный хроматин; 6 - диффузный хроматин; 7 - клеточная стенка; 8 - плазмалемма; 9 - плазмодесмы; 10 - эндоплазматическая агранулярная сеть; 11 - гранулярная эндоплазматическая сеть; 12 - митохондрии; 13 - свободные рибосомы; 14 - лизосомы; 15 - хлоропласт; 16 - диктиосома аппарата Гольджи; 17- гиалоплазма; 18 - тонопласт; 19 - вакуоль с клеточным соком
сте с погруженными в нее органеллами и ядром, называемое током цитоплазмы, или циклозом. Гиалоплазма представляет собой сложный бесцветный коллоидный раствор слизистой консистенции. Одним из проявлений живого состояния гиалоплазмы является переход коллоидной системы с преобладанием дисперсионной среды - воды (золя) в гель, находящийся в более твердом состоянии, и обратно. Гиалоплазма содержит воду (70-90%), в которой растворены ионы минеральных солей, играющие важную роль в создании осмотического давления в клетке. В состав гиалоплазмы входят также растворимые белки, рибонуклеиновая кислота (РНК), полисахариды, липиды.
Вещества, входящие в состав живой клетки, объединяют в понятие конституционные, т.е. участвующие в обмене веществ. Основны-
ми классами конституционных органических веществ являются белки, нуклеиновые кислоты, липиды и углеводы.
Белки - вещества, определяющие строение и свойства живой материи. На их долю приходится основная масса органических веществ клетки. Они участвуют в построении структуры и функциях всех клеточных органелл. Белки выполняют важную ферментативную функцию, постоянно участвуя в процессах синтеза и распада конституционных веществ. Белки могут быть и эргастическими веществами клетки, т.е. откладываться в запас, а также выполнять сократительную, транспортную функции и могут служить источником энергии.
Белки - это биополимеры, состоящие из аминокислот, соединенных пептидными связями. Из известных 40 аминокислот в состав белков входят 20. Простые белки - протеины - состоят только из аминокислот и откладываются в клетке в качестве запасных веществ. Простые белки могут соединяться с углеводами (гликопротеиды), нуклеиновыми кислотами (нуклеопротеиды), жирными кислотами (липопротеиды), и тогда они называются сложными белками - протеидами. Протеиды являются конституционными белками, так как входят в состав цитоплазмы и ядра.
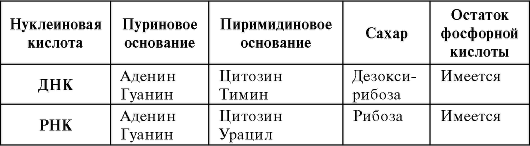
Нуклеиновые кислоты (ДНК и РНК) - важная группа фосфорсодержащих биополимеров, обеспечивающая хранение и передачу наследственной информации. Мономером нуклеиновых кислот является нуклеотид, включающий в себя азотистое основание, сахар и остаток фосфорной кислоты. Для каждого вида характерен свой ну- клеотидный состав нуклеиновых кислот.
Молекула ДНК состоит из 2 полинуклеотидных антипараллельных цепей, закрученных в двойную спираль вокруг центральной оси. Цепи выстраиваются по типу комплементарности между азотистыми основаниями, образующими между собой водородные связи. Структура ДНК описана в 1953 г. Д. Уотсоном и Ф. Криком. РНК, в отличие от ДНК, представляет собой одноцепочечную молекулу (табл. 1.2). ДНК и РНК могут находиться как в ядре, так и в цито- плазме, а также в митохондриях и хлоропластах.
Липиды (фосфолипиды) - жироподобные вещества, которые являются структурными компонентами клетки, так как входят в состав клеточной мембраны (плазмолемма, тонопласт). Протопласт растительной клетки содержит: простые липиды (жирные масла), полимерные липиды (воск, кутин, суберин) и сложные липиды (липоиды, или жироподобные вещества). Простые липиды состоят из остатков
Таблица 1.2. Состав нуклеиновых кислот (ДНК, РНК)
жирных кислот и спиртов (жиры, воски). Сложные липиды - это комплексы липидов, их соединения с белками (липопротеиды), фосфорной кислотой (фосфолипиды), сахарами (гликолипиды). Некоторые пигменты (каротиноиды) также относят к сложным липидам. Липиды являются одним из основных компонентов биологических мембран, а также составляют их энергетический резерв.
Углеводы входят в состав гиалоплазмы в виде моносахаридов (глю- коза, фруктоза), дисахаридов (сахароза, мальтоза и др.) и полисахаридов (крахмал, гликоген).
У растений моносахариды являются первичным продуктом фотосинтеза и используются для биосинтеза полисахаридов, аминокис- лот, жирных кислот и др. Углеводы запасаются в виде крахмала как энергетический резерв растений. Некоторые углеводные полимеры служат опорным материалом жестких клеточных стенок (целлюлоза) или выполняют функцию цементирующего материала в межклеточном пространстве (пектины).
ОРГАНЕЛЛЫ ЦИТОПЛАЗМЫ
В гиалоплазме находятся различные по своим функциям клеточные органеллы: митохондрии, пластиды, диктиосомы, ЭПС, микро- тельца, лизосомы, рибосомы, сферосомы, микротрубочки, микрофиламенты (табл. 1.3).
Таблица 1.3. Строение растительной клетки
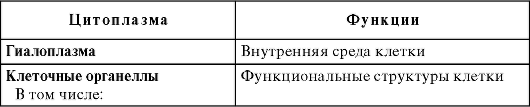
Окончание таблицы
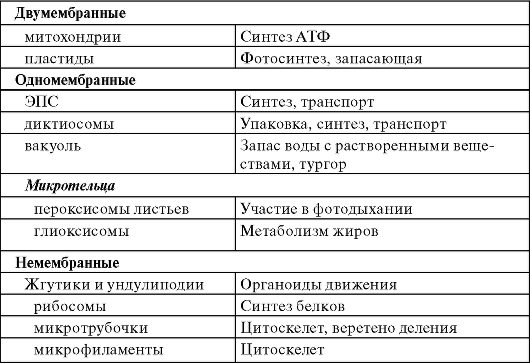
ДВУМЕМБРАННЫЕ ОРГАНЕЛЛЫ
Митохондрия (рис. 1.2) - двумембранный органоид. Внутренняя мембрана образует выросты - кристы. При аэробном дыхании на кристах происходят окислительное фосфорилирование и перенос электронов. Внутреннее содержимое митохондрий - матрикс - включает рибосомы, кольцевую молекулу ДНК и фосфатные гранулы. В матриксе находятся ферменты, участвующие в цикле Кребса и окислении жирных кислот.
Хлоропласт (рис. 1.3) - крупная двумембранная пластида, в кото- рой протекает фотосинтез за счет наличия пигментов: хлорофиллов, каротиноидов и ксантофиллов. Внутренняя среда хлоропласта - студенистообразный матрикс - строма. Строма содержит рибосомы, кольцевую молекулу ДНК и капельки масла. В строме протекает темновая фаза фотосинтеза, в которой непосредственно происходит синтез органических соединений с использованием энергии, синте- зированной в световую фазу. В строме на ламеллах находится система мембран-тилакоидов, собранных в стопки-граны, в которых может откладываться крахмал. В тилакоидах протекает световая фаза фотосинтеза, в ходе которой осуществляются процессы циклического и нециклического фосфорилирования и фотолиза воды под действи-
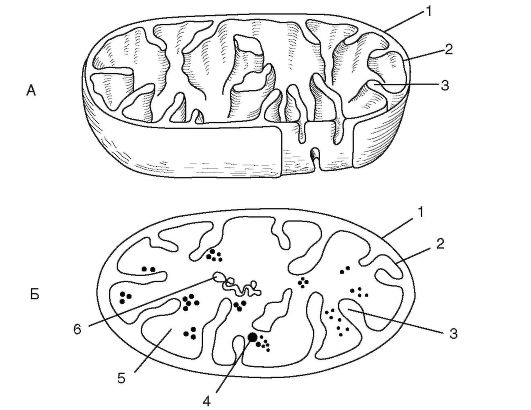
Рис. 1.2. Схемы строения митохондрии в трехмерном изображении (А)
и на срезе (Б):
1 - наружная мембрана митохондрии; 2 - внутренняя мембрана; 3 - криста; 4 - рибосома; 5 - матрикс; 6 - кольцевая нить ДНК
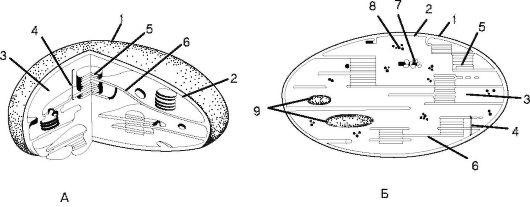
Рис. 1.3.Строениехлоропласта:
А - объемная схема строения хлоропласта; Б - схема среза через хлоропласт:
1 - наружная мембрана; 2 - внутренняя мембрана; 3 - строма; 4 - грана; 5 - тилакоид граны; 6 - ламелла; 7 - ДНК; 8 - рибосомы хлоропласта (отличаются от цитоплазматических рибосом); 9 - крахмальные зерна
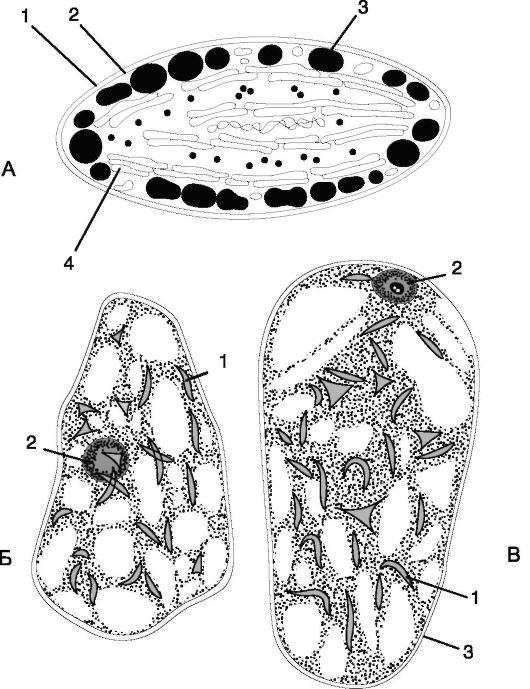
Рис. 1.4. Строение хромопласта:
А. Внешний вид: 1 - наружная мембрана; 2 - внутренняя мембрана; 3 - жировые капли; 4 - ламеллы;
Б, В: хромопласты в клетках мякоти зрелых плодов рябины (Sorbus aucuparia) и боярышника (Crataegus sangunea) соответственно: 1 - хромопласты; 2 - ядро; 3 - стенка клетки
ем квантов света. Хлоропласты могут превратиться в хромопласты (пожелтение листьев) или в лейкопласты (если поместить растение в темноту).
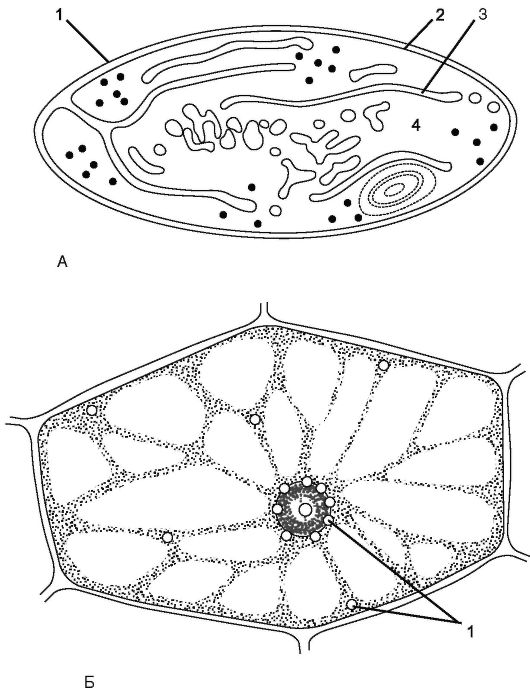
Рис. 1.5. Строение лейкопласта: А. Внешний вид лейкопласта:
1 - наружная мембрана; 2 - внутренняя мембрана; 3 - ламелла; 4 - строма.
Б. Лейкопласты (1) в клетках листа традесканции
Хромопласт - окрашенная пластида, содержащая пигменты: каротиноиды (оранжевые) и ксантофиллы (желтые). Хромопласты являются конечным этапом в развитии пластид, поэтому у них, как правило, отсутствует внутренняя мембранная система. От хлоропластов они отличаются меньшими размерами и разнообразной формой (рис. 1.4).
Больше всего хромопластов в плодах томата, красного перца, в цветках, где их яркая окраска служит для привлечения насекомых и птиц, участвующих в опылении растений и распространении семян.
Лейкопласт - бесцветная пластида, не содержащая пигментов. В отличие от хлоропластов у лейкопластов слабо развитая мембранная система и редко расположенные одиночные тилакоиды. Лейкопласты могут превращаться в хлоропласты и хромопласты.
Лейкопласты приспособлены для хранения запасных питательных веществ, поэтому их особенно много в запасающих органах: корнях, семенах, молодых листьях (рис. 1.5). В амилопластах откладывается запасной крахмал; в олеопластах - липиды; в протеинопластах - белки.
ОДНОМЕМБРАННЫЕ ОРГАНЕЛЛЫ
ЭПС - система уплощенных одномембранных мешочков (ци- стерн) в виде трубочек и пластинок, образующих единое целое с наружной мембраной ядерной оболочки (рис. 1.6). Различают 2 типа ЭПС: при наличии на поверхности ЭПС рибосом - она называется шероховатой, или гранулярной, а если рибосом нет - гладкой. На мембранах шероховатой ЭПС происходит синтез белков, а на мембранах гладкой ЭПС - синтез веществ небелковой природы (углеводы, липиды).
Диктиосома (аппарат Гольджи) - стопка уплощенных одномембранных мешочков (цистерн; рис. 1.7). На одном конце стопки мешочки образуются непрерывно, а на другом - отшнуровываются в виде пузырьков Гольджи. Стопки могут существовать и в виде дискретных (отдельных) диктиосом.
Диктиосомы участвуют в процессе секреции и синтеза углеводов (растительная клеточная стенка); в них образуются первичные лизосомы. Многие ферменты, синтезируемые на ЭПС, претерпевают модификацию в цистернах и траспортируются пузырьками Гольджи.
Микротельца - органеллы, не совсем правильной сферической формы с зернистой структурой, но иногда в них попадаются кристаллоиды или скопление нитей.
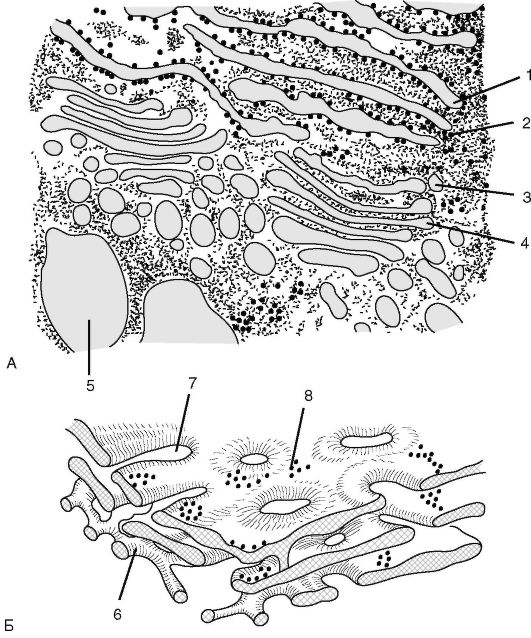
Рис. 1.6. Строение эндоплазматической сети: А - цистерны гранулярной ЭПС; Б - система цистерн гранулярной ЭПС и трубок агранулярной ЭПС; 1 - цистерна гранулярной ЭПС; 2 - рибосома; 3 - пузырек Гольджи; 4 - цистерны диктиосомы; 5 - вакуоль; 6 - трубка агранулярной ЭПС; 7 - окна в ретикулярной цистерне; 8 - прикрепленная полисома
Глиоксисомы имеют отношение к метаболизму глиоксилата и уча- ствуют в превращении липидов в сахарозу в некоторых богатых маслами семенах (например, в эндосперме семени клещевины).
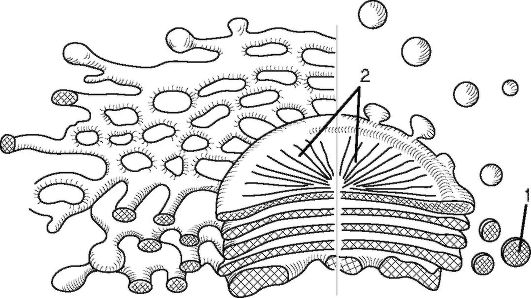
Рис. 1.7. Строение диктиосомы (аппарат Гольджи): 1 - пузырек Гольджи; 2 - цистерна диктиосомы в плане
Пероксисомы содержат фермент каталазу, ускоряющую разложение перекиси водорода на воду и кислород. Перекись водорода является побочным продуктом некоторых окислительных процессов, протекающих в клетке. Она очень токсична и должна немедленно удаляться из клетки. Пероксисомы листьев тесно связаны с процессом фотодыхания при участии хлоропластов и митохондрий.
НЕМЕМБРАННЫЕ ОРГАНЕЛЛЫ. ЖГУТИКИ И УНДУЛИПОДИИ
Жгутики, реснички, фимбрии - двигательные приспособления прокариот. Базальная расширенная часть жгутиков называется ба- зальным тельцем.
У определенной части эукариотических клеток двигательными приспособлениями являются ундулиподии, отличающиеся по строению от жгутиковидных образований. Ундулиподии имеются у многих протоктистов - водорослей и грибоподобных (см. главу 5), особенно на одноклеточных стадиях их жизненного цикла. У таких растений, как мхи, папоротники и часть голосеменных, ундулиподиями снабжены только мужские половые клетки. Ундулиподии снаружи покрыты мембраной, которая представляет собой единое целое с плазматической мембраной клетки. На поперечном срезе 9 пар микротрубочек образуют кольцо, а 2 дополнительные микротрубочки располагаются в центре кольца (организация: 9+2). Ундулиподии отходят от цилиндрических структур, называемых кинетосомами. У кинетосом на по-
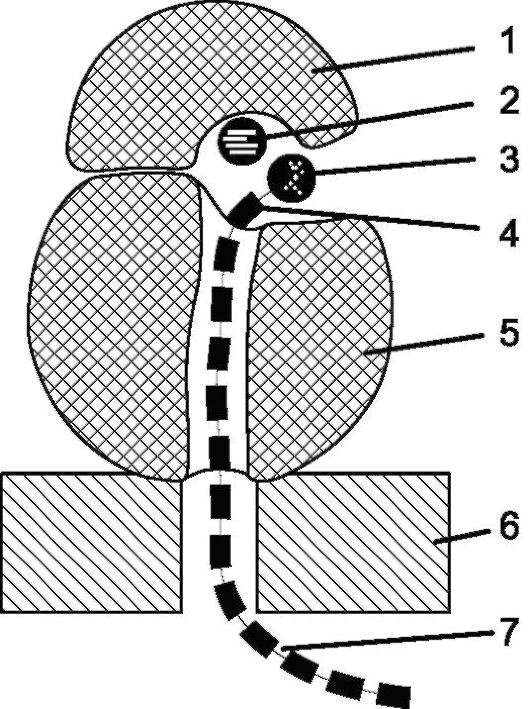
Рис. 1.8. Схема строения рибосомы, сидящей на мембране ЭПС: 1 - малая субъединица; 2 - иРНК; 3 - аминоацил тРНК; 4 - аминокислота; 5 - большая субъединица; 6 - мембрана ЭПС; 7 - синтезируемая полипептидная цепь
перечном срезе заметно лишь периферическое кольцо микротрубочек, собранных по 3 (организация: 9+0). Движение ундулиподиев может также осуществляться автономно, они способны двигаться и после отделения от клетки.
Рибосомы - очень мелкие немембранные органеллы, состоящие из 2 субчастиц (единиц): большой и малой. Они состоят из РНК и белка. Рибосомы свободно располагаются в цитоплазме или прикрепляются к мембранам ЭПС (рис. 1.8). Рибосомы способны образовывать полисому, в которой они нанизаны на единую нить информационной РНК. В рибосомах происходит синтез белка.
Микротрубочки - тонкие цилиндрические структуры, состоящие из субъединиц белка, называемого тубулином. Микротрубочки контролируют упаковку целлюлозных микрофибрилл при формировании клеточной стенки; участвуют в формировании веретена деления.
Микрофиламенты - это длинные нити, состоящие из сократительного белка актина. Пучки микрофиламентов играют определяющую роль в токах цитоплазмы. Микрофиламенты вместе с микротрубочками образуют гибкую сеть, называемую цитоскелетом.
ЯДРО
Ядро - самая крупная органелла, имеющая двумембранную обо- лочку, пронизанную ядерными порами. В матриксе ядра находится хроматин: гетерохроматин (компактный, менее активный) и эухроматин (диффузный, активный) - в такой форме раскрученные
хромосомы находятся в интерфазе. Хромосомы содержат нуклеиновую кислоту ДНК, отвечающую за наследственность. Внутри ядра содержится хорошо заметная округлая структура - ядрышко, в котором происходит синтез рибосомальной РНК (рРНК); ядрышек может быть несколько. Деление ядра лежит в основе размножения клеток.
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
Цитоплазматическая мембрана, отделяющая цитоплазму от клеточной стенки, называется плазмалеммой (плазматической мембраной), а отделяющая ее от вакуоли - тонопластом (элементарной мембраной).
В настоящее время пользуются жидкостно-мозаичной моделью мембраны (рис. 1.9), в соответствии с которой мембрана состоит из бислоя липидных молекул (фосфолипидов) с гидрофильными головками и 2 гидрофобными хвостами, обращенными внутрь слоя. Помимо липидов в состав мембран входят белки.
Различают 3 типа мембранных белков, «плавающих» в билипидном слое: интегральные белки, пронизывающие всю толщу бислоя; полуинтегральные, пронизывающие бислой неполностью; перифери- ческие, прикрепляющиеся с внешней или внутренней стороны мембраны к другим мембранным белкам. Мембранные белки выполняют различные функции: одни из них являются ферментами, другие выполняют роль переносчиков специфических молекул через мембрану или образуют гидрофильные поры, через которые могут проходить полярные молекулы.
Одним из основных свойств клеточных мембран является их полупроницаемость: они пропускают воду, но не пропускают растворенные в ней вещества, т. е. обладают избирательной проницаемостью.
Транспорт через мембраны
В зависимости от затрат энергии транспорт веществ и ионов через мембрану делится на пассивный, не требующий затрат энергии, и активный, связанный с потреблением энергии. К пассивному транс- порту относятся такие процессы, как диффузия, облегченная диффузия, осмос.
Диффузия - это процесс проникновения молекул через липидный бислой по градиенту концентраций (из области большей концентра-
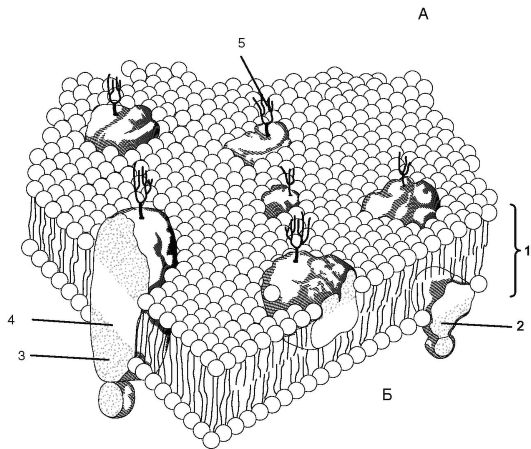
Рис. 1.9. Схема строения биологической мембраны: А - внеклеточное пространство; Б - цитоплазма; 1 - бимолекулярный слой липидов; 2 - периферический белок; 3 - гидрофильная область интегрального белка; 4 - гидрофобная область интегрального белка; 5 - углеводная цепь
ции в область меньшей). Чем меньше молекула и чем более неполярная, тем быстрее она диффундирует через мембрану.
При облегченной диффузии прохождению вещества через мембрану помогает какой-либо транспортный белок. Таким образом, в клетку поступают различные полярные молекулы, такие, как сахара, амино- кислоты, нуклеотиды и др.
Осмос - это диффузия воды через полупроницаемые мембраны. Осмос вызывает передвижение воды из раствора с высоким водным потенциалом в раствор - с низким водным потенциалом.
Активный транспорт - это перенос молекул и ионов через мембрану, сопровождаемый энергетическими затратами. Активный транспорт идет против градиента концентрации и электрохимического градиента и использует энергию АТФ. В основе механизма ак-
тивного транспорта веществ лежит работа протонного насоса (Н+ и К+) у растений и грибов, которые сохраняют внутри клетки высокую концентрацию К+ и низкую - Н+ (Na+ и К+ - у животных). Энергия, необходимая для работы этого насоса, поставляется в виде АТФ, синтезируемой в процессе клеточного дыхания.
Известна еще одна разновидность активного транспорта - эндо- и экзоцитоз. Это 2 активных процесса, с помощью которых различные молекулы транспортируются через мембрану в клетку (эндоцитоз) либо из нее (экзоцитоз).
При эндоцитозе вещества попадают в клетку в результате инвагинации (впячивания) плазматической мембраны. Образующиеся при этом пузырьки, или вакуоли, переносятся в цитоплазму вместе с заключенными в них веществами. Поглощение больших частиц, таких, как микроорганизмы или обломки клеток, называется фагоцитозом. В этом случае образуются крупные пузырьки, называемые вакуолями. Поглощение жидкостей (суспензий, коллоидных растворов) или растворенных веществ с помощью небольших пузырьков носит на- звание пиноцитоз.
Обратный эндоцитозу процесс называется экзоцитозом. Многие вещества выводятся из клетки в специальных пузырьках или вакуолях. Примером может служить вывод из секреторных клеток их жидких секретов; другой пример - это участие пузырьков диктиосом в формировании клеточной оболочки.
ПРОИЗВОДНЫЕ ПРОТОПЛАСТА
Вакуоль
Вакуоль - это резервуар, ограниченный одинарной мембраной - тонопластом. В вакуоли содержится клеточный сок - концентрированный раствор различных веществ, таких, как минеральные соли, сахара, пигменты, органические кислоты, ферменты. В зрелых клетках вакуоли сливаются в одну, центральную.
В вакуолях хранятся различные вещества, в том числе конечные продукты обмена. От содержимого вакуоли в сильной степени зависят осмотические свойства клетки.
В связи с тем что вакуоли содержат крепкие растворы солей и других веществ, клетки растений постоянно осмотически поглощают воду и создают гидростатическое давление на клеточную стенку, на-
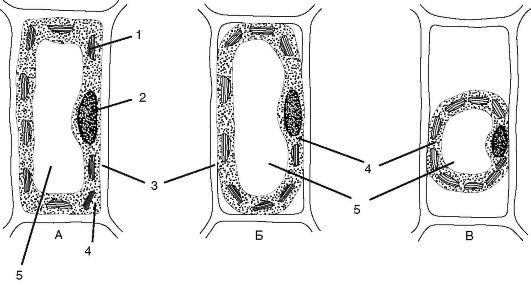
Рис. 1.10. Схема плазмолиза: А - клетка в состоянии тургора (в изотоническом растворе); Б - начало плазмолиза (клетка, помещенная в 6% раствор КNО3); В - полный плазмолиз (клетка, помещенная в 10% раствор КNО3); 1 - хлоропласт; 2 - ядро; 3 - клеточная стенка; 4 - протопласт; 5 - центральная вакуоль
зываемое тургорным. Тургорному давлению противостоит равное ему по величине давление клеточной стенки, направленное внутрь клетки. Большинство растительных клеток существуют в гипотонической среде. Но если такую клетку поместить в гипертонический раствор, вода по законам осмоса начнет выходить из клетки (для выравнивания водного потенциала по обе стороны мембраны). Вакуоль при этом сократится в объеме, ее давление на протопласт уменьшится, и мембрана начнет отходить от клеточной стенки. Явление отхождения протопласта от клеточной стенки называется плазмолизом. В природных условиях такая потеря тургора в клетках приведет к увяданию растения, опусканию листьев и стеблей. Однако этот процесс обратим: если клетку поместить в воду (например, при поливе растения), возникает явление, обратное плазмолизу - деплазмолиз (рис. 1.10).
Включения
Клеточными включениями являются запасные и экскреторные вещества.
Запасные вещества (временно выключенные из обмена) и вместе с ними отбросы (экскреторные вещества) часто называют эргастиче-
скими веществами клетки. К запасным веществам относят запасные белки, жиры и углеводы. Эти вещества накапливаются в течение вегетационного периода в семенах, плодах, подземных органах растения и в сердцевине стебля.
Запасные вещества
Запасные белки, относящиеся к простым белкам - протеинам, чаще откладываются в семенах. Осаждающиеся белки в вакуолях образуют зерна округлой или эллиптической формы, называемые алейроновыми. Если алейроновые зерна не имеют заметной внутренней структуры и состоят из аморфного белка, их называют простыми. Если в алейроновых зернах среди аморфного белка встречаются кристаллоподобная структура (кристаллоид) и блестящие бесцветные тельца округлой формы (глобоиды), такие алейроновые зерна называют сложными (рис. 1.11). Аморфный белок алейронового зерна представлен гомогенным непрозрачным белком желтоватого цвета, набухающим в воде. Кристаллоиды имеют характерную для кристаллов ромбоэдрическую форму, но в отличие от истинных кристал-
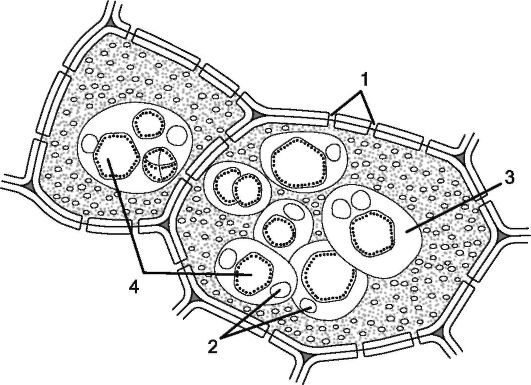
Рис. 1.11. Сложные алейроновые зерна: 1 - поры в оболочке; 2 - глобоиды; 3 - аморфная белковая масса; 4 - кристаллоиды, погруженные в амфорную белковую массу
лов составляющий их белок набухает в воде. Глобоиды состоят из кальциево-магниевой соли, содержат фосфор, нерастворимы в воде и не дают реакцию на белки.
Запасные липиды обычно располагаются в гиалоплазме в виде ка- пель и встречаются почти во всех растительных клетках. Это основной тип запасных питательных веществ большинства растений: наиболее богаты ими семена и плоды. Жиры (липиды) - наиболее калорийное запасное вещество. Реактивом на жироподобные вещества является судан III, окрашивающий их в оранжевый цвет.
Углеводы входят в состав каждой клетки в виде растворимых в воде сахаров (глюкозы, фруктозы, сахарозы) и нерастворимых в воде по- лисахаридов (целлюлозы, крахмала). В клетке углеводы играют роль источника энергии для реакций обмена веществ. Сахара, связываясь с другими биологическими веществами клетки, образуют гликозиды, а полисахариды с белками - гликопротеины. Состав углеводов растительной клетки значительно более разнообразен, чем у животных клеток, за счет разнообразного состава полисахаридов клеточной оболочки и сахаров клеточного сока вакуолей.
Главнейшим и наиболее распространенным запасным углеводом является полисахарид крахмал. Первичный ассимиляционный крахмал образуется в хлоропластах. Ночью, при прекращении фотосинтеза, крахмал гидролизуется до сахаров и транспортируется в запасающие ткани - клубни, луковицы, корневища. Там в особых типах лейкопластов - амилопластах - часть сахаров откладывается в виде зерен вторичного крахмала. Для крахмальных зерен характерна слоистость, что объясняется различным содержанием воды из-за неравномерного поступления крахмала в течение суток. В темных слоях воды больше, чем в светлых. Зерно с одним центром крахмалообразования в центре амилопласта называют простым концентрическим, если центр смещен - простым эксцентрическим. Зерно с несколькими крахмалообразующими центрами - сложное. У полусложных зерен новые слои откладываются вокруг нескольких крахмалообразующих центров, а затем формируются общие слои и покрывают крахмалообразующие центры (рис. 1.12). Реактивом на крахмал является раствор йода, дающий синее окрашивание.
Экскреторные вещества (продукты вторичного обмена)
К клеточным включениям относятся и экскреторные вещества, например кристаллы оксалата кальция (одиночные кристаллы, рафи-
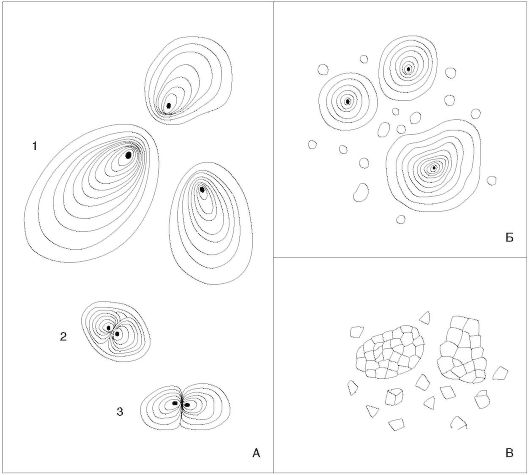
Рис. 1.12. Крахмальные зерна картофеля (А): 1- простое зерно; 2 - полусложное; 3 - сложное; пшеницы (Б), овса (В)
ды - игольчатые кристаллы, друзы - сростки кристаллов, кристалли- ческий песок - скопление множества мелких кристаллов) (рис. 1.13). Реже кристаллы состоят из карбоната кальция или кремнезема (цистолиты; рис. 1.14). Цистолиты откладываются на клеточной стенке, вдающейся внутрь клетки в виде гроздьев винограда, и характерны, например, для представителей семейства крапивных, листьев фикуса.
В отличие от животных, выводящих избыток солей вместе с мочой, растения не имеют развитых органов выделения. Поэтому считается, что кристаллы оксалата кальция являются конечным продуктом ме- таболизма протопласта, образующимся как приспособление для выведения из обмена излишков кальция. Как правило, эти кристаллы накапливаются в органах, которые растение периодически сбрасывает (листья, кора).
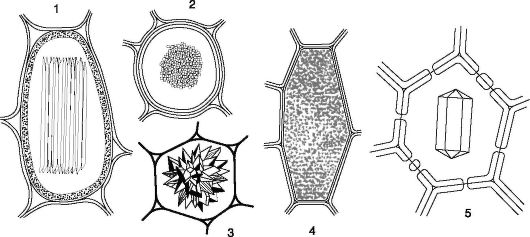
Рис. 1.13. Формы кристаллов оксалата кальция в клетках: 1, 2 - рафида (недотрога; 1- вид сбоку, 2 - на поперечном срезе); 3 - друза (опунция); 4 - кристаллический песок (картофель); 5 - одиночный кристалл (ваниль)
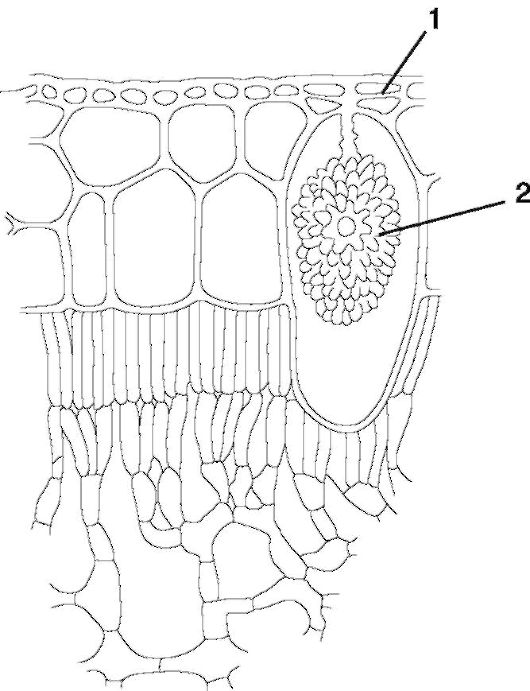
Рис. 1.14. Цистолит (на поперечном срезе листа фикуса): 1 - кожица листа; 2 - цистолит
Эфирные масла скапливаются в листьях (мята, лаванда, шалфей), цветках (шиповник), плодах (цитрусовые) и семенах растений (укроп, анис). Эфирные масла не принимают участия в обмене веществ, но их широко используют в парфюмерии (розовое, жасминное масла), пищевой промышленности (анисовое, укропное масла), медицине
(мятное, эвкалиптовое масла). Резервуарами для скопления эфирных масел могут быть желёзки (мята), лизигенные вместилища (цитрусовые), железистые волоски (герань).
Смолы - это комплексные соединения, образующиеся в процессе нормальной жизнедеятельности или в результате разрушения тканей. Они образуются эпителиальными клетками, выстилающими смоляные ходы, как побочный продукт обмена веществ, часто с эфирными маслами. Могут накапливаться в клеточном соке, цитоплазме в виде капель или во вместилищах. Они нерастворимы в воде, непроницаемы для микроорганизмов и благодаря своим антисептическим свойствам повышают сопротивляемость растений болезням. Применяются смолы в медицине, а также при изготовлении красок, лаков и смазочных масел. В современной промышленности заменяются синтетическими материалами.
Клеточная стенка
Жесткая клеточная стенка, окружающая клетку, состоит из целлюлозных микрофибрилл, погруженных в матрикс, в состав которого входят гемицеллюлозы и пектиновые вещества. Клеточная стенка обеспечивает механическую опору клетке, защиту протопласта и сохранение формы клетки. При этом клеточная стенка способна к растяжению. Являясь продуктом жизнедеятельности протопласта, стенка может расти только в контакте с ним. Через клеточную стенку происходит передвижение воды и минеральных солей, но для высокомолекулярных веществ она полностью или частично непроницаема. При отмирании протопласта стенка может продолжать выполнять функцию проведения воды. Наличие клеточной стенки более чем все другие признаки отличает растительные клетки от животных. Архитектуру клеточной стенки в значительной степени определяет целлюлоза. Мономером целлюлозы является глюкоза. Пучки молекулы целлюлозы формируют мицеллы, которые объединяются в более крупные пучки - микрофибриллы. Реактивом на целлюлозу является хлор-цинк-йод (Cl-Zn-I), дающий сине-фиолетовое окрашивание.
Целлюлозный каркас клеточной стенки заполнен нецеллюлозными молекулами матрикса. В состав матрикса входят полисахариды, называемые гемицеллюлозами; пектиновые вещества (пектин), очень близкие к гемицеллюлозам, и гликопротеиды. Пектиновые вещества, сливаясь между соседними клетками, образуют срединную пластинку, которая располагается между первичными оболочками соседних
клеток. При растворении или разрушении срединной пластинки (что происходит в мякоти созревших плодов) возникает мацерация (от лат. maceratio - размягчение). Естественную мацерацию можно наблюдать у многих перезрелых плодов (арбуз, дыня, персик). Искусственную мацерацию (при обработке тканей щелочью или кислотой) используют для приготовления различных анатомических и гистологических препаратов.
Клеточная стенка в процессе жизнедеятельности может подвергаться различным видоизменениям - одревеснению, опробковению, ослизнению, кутинизации, минерализации (табл. l.4).
Таблица 1.4. Характерные изменения вторичной клеточной стенки
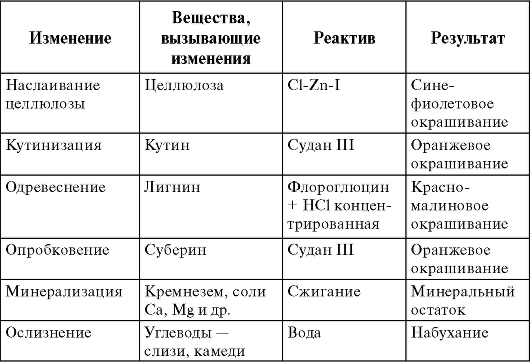
Одревеснение клеточной стенки связано с внедрением между молекулами целлюлозы лигнина, который является самым распространенным (после целлюлозы) полимером растительных клеток. Он увеличивает жесткость оболочки, вызывая одревеснение клеточных стенок; лигнин обычно содержится в клетках, выполняющих опорную или механическую функцию. Реактив на лигнин HCl (концентрированная) + флороглюцин дает красно-малиновое окрашивание.
Кутин, суберин и воск - жироподобные вещества. Кутин и воск обычно откладываются на поверхности клеток эпидермы. Кутиновая пленка образует кутикулу. Суберин пропитывает клеточные стенки вторичной покровной ткани, вызывая опробковение. В момент завершения опробковения протопласт клетки разрушается, а клеточная стенка пробки становится непроницаемой для воды и газов. Кутин и суберин встречаются в комбинации с восками и предотвращают чрезмерную потерю воды растением и проникновение в его клетки различных бактерий и грибов.
Клеточные стенки могут пропитываться оксалатами и кремнеземом, что придает им твердость и хрупкость и приводит к их минерализации. Отложение кремнезема характерно для стеблей и боковых побегов хвощей, стеблей злаков и осок.
Некоторые клеточные стенки кожуры семян (льна) способны к ослизнению. Это происходит за счет превращения целлюлозы и пектина в слизи и камеди, которые, являясь полимерами, сильно набухают при соприкосновении с водой. Слизи удерживают влагу, защищая семена от высыхания, и закрепляют их на определенном месте, с частицами почвы. Например, у стеблей вишни выделение слизей и камедей на- блюдается из пораненных участков. Камедь при этом выделяется в виде наплывов (вишневый клей); этот процесс называют гуммузом.
При образовании клетки в процессе митоза формируется первичная клеточная стенка. На всем своем протяжении первичные клеточные стенки не одинаковы по толщине, а имеют тонкие участки, которые называются первичными поровыми полями (рис. 1.15). У некоторых клеток протопласт откладывает на внутреннюю поверхность первичной оболочки еще один слой - вторичную клеточную стенку (см. рис. 1.15). На первичных поровых полях вторичная оболочка не откладывается. Таким образом, во вторичной оболочке образуется пора. Поры двух контактирующих клеток лежат друг против друга. В клетках, имеющих вторичные оболочки, существуют 2 типа пор - простые и окаймленные. Вторичная оболочка в окаймленных порах нависает над порой, а внутри имеется торус (утолщение), регулирующий движение жидкости в соседние клетки (рис. 1.16). В простых порах диаметр порового канала одинаковый по всей длине.
Такие оболочки нужны специализированным клеткам, выполняющим механическую и проводящую функции. После отложения вторичной оболочки и ее одревеснения протопласт клеток часто разрушается. Обычный компонент вторичных оболочек клеток древесины (ксилемы) и склеренхимы - лигнин.
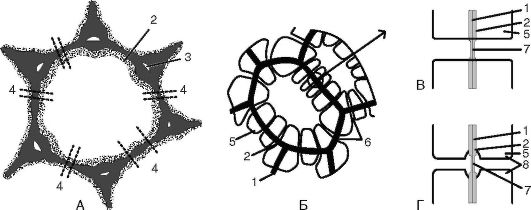
Рис. 1.15. Первичные поровые поля, поры и плазмодесмы: А - паренхимная клетка с первичной оболочкой и первичными поровыми полями; Б - клетки с вторичными оболочками и многочисленными порами; В - пара простых пор; Г - пара окаймленных пор; 1 - срединная пластинка; 2 - первичная оболочка; 3 - межклеточное пространство; 4 - плазмодесмы в первичном поровом поле; 5 - вторичная оболочка; 6 - поры; 7 - поровая мембрана; 8 - окаймление
В отличие от животной клетки для строения растительной клетки характерны:
1) наличие прочной целлюлозной клеточной стенки, представляющей собой видоизмененный гликокаликс животной клетки;
2) наличие пластид и автотрофнос питания (фотосинтез);
3) наличие (как правило) крупной центральной вакуоли;
4) рост клеток - путем растяжения (в основном за счет увеличения объема вакуоли);
5) отсутствие клеточного центра у высших растений, участвующего в делении клетки, роль которого выполняют отдельные микротрубочки;
6) способность к неограниченному или очень продолжительному росту;
7) малая подвижность (как правило, у всех клеток высших растений).
Растениям свойственны медленные движения: настии (уплотнения с изгибом), нутации (круговые движения), тропизмы (изгибы органов растения). Лишь среди зеленых водорослей встречается много подвижных форм.
ДЕЛЕНИЕ ЭУКАРИОТИЧЕСКИХ КЛЕТОК
Жизненный (митотический) цикл клетки включает ее существование от момента деления до следующего деления или смерти. Цикл
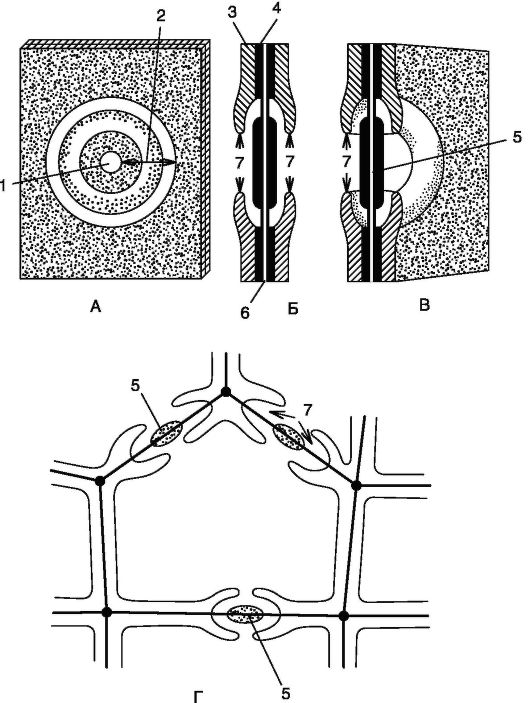
Рис. 1.16. Строение окаймленных пор в трахеидах хвойных: А - схема поры в плане; Б - поперечный срез; В - объемное изображение; Г - поперечный срез трахеиды сосны с окаймленными порами; 1 - внутреннее отверстие поры, упирающееся в полость клетки; 2 - наружное отверстие поры, упирающееся в первичную оболочку; 3 - вторичная оболочка; 4 - первичная оболочка; 5 - торус; 6 - срединная пластинка; 7 - внутреннее отверстие поры
клетки складывается из 3 главных стадий: интерфазы, в которой происходят активный рост и функционирование клеток, а также подготовка их к делению; деления ядра и цитокинеза - процесса разделения цитоплазмы между дочерними клетками.
Деление, при котором сохраняется идентичность хромосомного набора материнской и дочерних клеток, называется митозом. Ми- тоз - основной способ деления всех соматических клеток. Так, из одной материнской клетки с набором 2п4с образуются две дочерние с набором 2п2с*. Но в митоз могут вступать и гаплоидные клетки. Так, у голосеменных и покрытосеменных растений митоз наблюдается при образовании половых клеток.
Деление, при котором происходит редукция хромосомного набора (от диплоидного к гаплоидному), называется мейозом и наблюдается у растений при образовании спор. Так, из 1 диплоидной клетки (2п4с) образуются 4 с гаплоидным набором (пс).
Интерфаза
Интерфаза перед митозом и мейозом складывается из 3 периодов (рис. 1.17): предсинтетического (G1), синтетического (S) и постсинтетического (G2)
G1-период - синтез всех видов РНК, белка, рост клетки.
S-период - редупликация молекул ДНК (число ДНК в каждой хромосоме удваивается, но плоидность хромосомного набора - число хромосом в ядре - не меняется), синтез гистоновых белков.
G2-период - продолжение синтеза белка, накопление энергии, удвоение клеточных органелл.
Продолжительность митотического цикла различна. Так, у одноклеточных эукариот он может варьировать от 0,5 ч до 2-3 сут. У многоклеточных (например, у вики посевной) составляет 15 сут, у бо- бов - около 2 сут.
Деление клетки подразделяется на процессы, происходящие в ядре: кариокинез (деление ядра) и цитокинез (деление цитоплазмы).
Митоз
Митоз подразделяют на 4 фазы: профаза, метафаза, анафаза, телофаза.
* n - гаплоидный набор; 2 n - диплоидный набор; с - количество молекул ДНК в хромосомах.
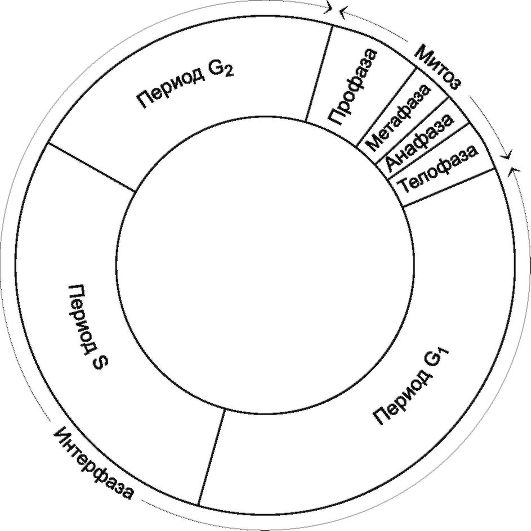
Рис. 1.17. Периоды интерфазы
Профаза - самая длительная фаза митоза. В ней происходят: спирализация и укорочение ДНК хромосом; исчезновение ядрышка и распад ядерной оболочки; формирование ахроматинового веретена, или веретена деления, состоящего из пучков микротрубочек, идущих от полюсов клетки. Таким образом, в конце профазы каждая хромосома состоит из 2 хроматид (2 молекул ДНК - 2с), скрепленных центромерой.
В метафазу хромосомы располагаются на экваторе клетки. Происходит продольное расщепление хромосом на 2 хроматиды. Плечи хроматид продольно расходятся, но они остаются скреплеными центромерой.
В анафазу происходит деление центромер надвое. Хроматиды расходятся к полюсам клетки за счет сокращения ахроматинового веретена. В результате на каждом полюсе сохраняется тот же набор хромосом (2n), но все они теперь состоят из 1 хроматиды (2п2с), а во всей клетке находятся 2 диплодных набора (4п4с).
Телофаза (обратная профазе) включает деспирализацию хромосом, они становятся плохо видимыми. Формируются ядрышко и ядерная оболочка вокруг хромосом на каждом полюсе. Веретено деления исчезает.
Цитокинез
Цитокинез - это формирование клеточной пластинки в экваториальной плоскости после появления 2 ядер. Между ядрами образуется бочкообразная система волокон, состоящая из микротрубочек, назы- ваемая фрагмопластом. В экваториальной плоскости фрагмопласта появляются пузырьки Гольджи, содержащие пектиновые вещества. Из них формируется срединная пластинка, а мембраны пузырьков идут на построение плазматической мембраны по обеим сторонам клетки. Каждый протопласт откладывает над срединной пластинкой первичную оболочку (рис. 1.18). После цитокинеза наступает состояние покоя - интерфаза.
Биологическое значение митоза заключается в передаче дочерним клеткам идентичной генетической информации родительской клет- ки. Кроме того, митоз обеспечивает рост и регенерацию тканей.
Мейоз
Мейоз был открыт в 1885 г. В.И. Беляевым. В мейоз вступают только диплоидные клетки. При половом процессе происходит слияние ядер 2 клеток, т.е. набор становится диплоидным (2п). Сущностью мейоза является восстановление гаплоидного набора в ядрах половых клеток, так как при повторении полового процесса из поколения в поколение число хромосом все время увеличивалось бы.
Мейоз состоит из 2 последовательных делений ядра; 1-е мейотическое деление называется редукционным, а 2-е, протекающее по схеме митоза, - эквационным (рис. 1.19).
Первое мейотическое (редукционное) деление
Профаза I: самая длительная по времени фаза, протекающая от нескольких часов до нескольких недель. Хромосомы спирализуются и становятся видимыми. Гомологичные хромосомы сближаются (конъюгируют) по всей длине и образуют пару - бивалент. В бивалентах происходит обмен участками гомологичных хромосом - кроссинговер, что приводит к обмену генетической информацией между хромосомами. К концу профазы исчезают ядрышко и ядерная оболочка, формируется ахроматиновое веретено.
Метафаза I: биваленты выстраиваются в экваториальной плоскости. Сестринские хроматиды скреплены центромерами, но их плечи продольно расходятся. Гомологичные хромосомы скреплены в нескольких точках - хиазмах (от греч. chiasma - перекрест).
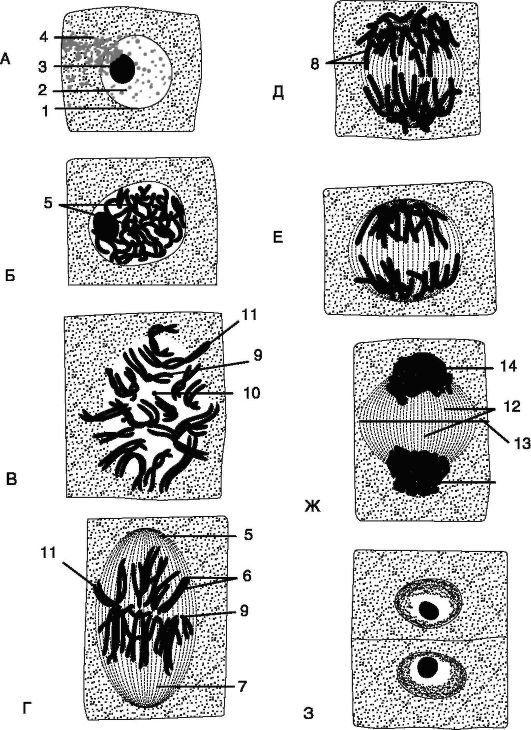
Рис. 1.18. Митоз и цитокинез в растительных клетках (корня лука): А - ядро в интерфазе; Б - ранняя и В - поздняя профазы; Г - метафаза; Д - ранняя анафаза; Е - поздняя анафаза; Ж - телофаза; З - сестринские клетки;
1 - ядерная мембрана; 2 - кариолимфа; 3 - ядрышко; 4 - цитоплазма; 5 - хроматиновая нить; 6 - хромосомы; 7 - ахроматированное веретено; 8 - хроматиды; 9 - центромера; 10 - спутник; 11 - плечи хромосом; 12 - фрагмопласт; 13 - закладывающаяся пектиновая перегородка; 14 - формирующееся ядро
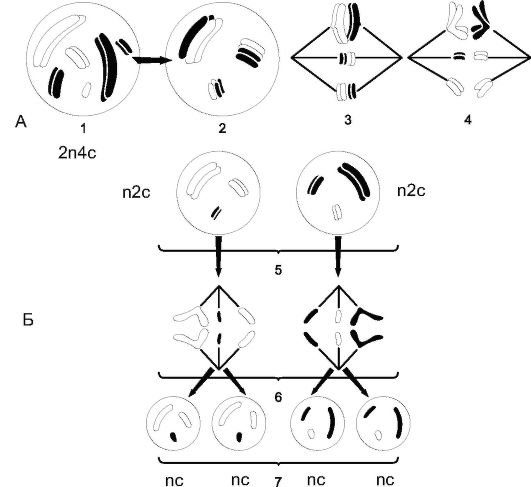
Рис. 1.19. Схема мейоза: А - первое мейотическое (редукционное) деление: 1, 2 - профаза I; 3 - метафаза I; 4 - анафаза I; 5 - телофаза I; Б - второе мейотическое деление: 6 - анафаза II; 7 - телофаза II
Анафаза I: ахроматиновые нити отделяют по одному мономеру и подтягивают по одной хромосоме, состоящей из 2 хроматид к полюсу. На каждом полюсе образуется гаплоидный набор (п2с), т.е. именно в анафазу происходит редукция хромосомного набора.
Телофаза I: кратковременна, в ней происходят деспирализация хромосом и формирование ядра.
Второе мейотическое деление следует за первым, минуя S-период, т.е. удвоение наследственного материала. Профаза II, метафаза II и телофаза II протекают по схеме митоза. В анафазе II в отличие от ми-
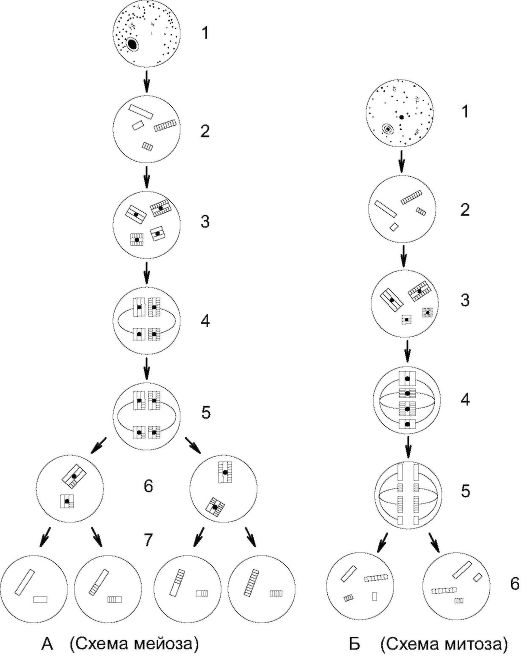
Рис. 1.20. Сравнительная схема мейоза (А) и митоза (Б) А. 1 - материнская клетка; 2 - период G1 интерфазы; 3 - S-период интерфазы; 4 - образование бивалентов; 5 - кроссинговер (обмен участками в бивалентах); 6 - расхождение хромосом в анафазе I; 7 - расхождение хроматид в анафазе II
Б. 1 - материнская клетка; 2 - период G1 и интерфазы; 3 - S-период интерфазы; 4 - метафаза митоза; 5 - расхождение хроматид в анафазе митоза; 6 - дочерние клетки с идентичным набором хромосом, как у материнской клетки
тоза центромеры делятся надвое, и к полюсам расходятся хроматиды. Теперь на каждом полюсе формируются хроматиды, а не хромосомы (набор - пс). Из двух гаплоидных ядер (п2с) формируются четыре ядра с набором (пс).
Значение мейоза велико: редукция хромосомного набора в половых клетках за счет расхождения хромосом в анафазе I и процессам достижения генетического разнообразия вследствие процесса кроссинговера (рис. 1.20).