Анатомия человека : учебник для стоматологических факультетов медицинских вузов / Под ред. Л.Л. Колесникова, - 4-е изд., перераб. и доп. - 2010. - 816 с. : ил.
|
|
|
|
УЧЕНИЕ О НЕРВНОЙ СИСТЕМЕ - НЕВРОЛОГИЯ
ОБЩАЯ НЕВРОЛОГИЯ
Совокупность нервной ткани в организме объединяется понятием «нервная система». Нервная система обеспечивает восприятие разнообразных чувствительных, или афферентных, импульсов, возникающих при воздействии на рецепторные зоны, которые присутствуют во всех органах и тканях.
Преобразованные в нервные импульсы различные виды раздражения (механические, световые, звуковые, вкусовые, обонятельные и др.) достигают соответствующих уровней нервной системы, где происходят их анализ и синтез. Результатом этой работы является формирование определенного решения, материальное обеспечение которого достигается проведением двигательных, или эфферентных, импульсов к рабочим органам. Эфферентные импульсы усиливают или ослабляют деятельность различных органов, поддерживают их в адекватном тонусе.
Таким образом, нервная система обеспечивает регуляцию всей деятельности организма человека на основе обработки поступающих в нее потоков информации.
Нервную систему условно разделяют на центральную и периферическую. К центральной нервной системе (ЦНС) относят головной и спинной мозг. Вся остальная нервная ткань, объединенная в нервные стволы, нервные узлы, нервные сплетения и нервные окончания, описывается как периферическая нервная система.
Структурно-функциональной единицей нервной ткани является нервная клетка, или нейрон. В любом нейроне выделяют тело; отросток,
по которому информация приходит в клетку, - дендрит; отросток, по которому информация уходит от клетки, - нейрит, или аксон. Каждый нейрон представляет собой особое звено переработки нервных импульсов, его индивидуальность определяется местоположением в нервной системе и связями с другими нейронами или рабочим органом.
Как правило, нервная клетка имеет несколько ветвистых дендритов, специализирующихся на восприятии определенных нервных импульсов. Контакт, посредством которого нервные импульсы передаются от одного уровня на другой, называется синапсом. Синапсы образуются также на поверхности тела нервной клетки, но их, как правило, значительно больше на ветвлениях дендритов.
В отличие от дендритов аксон всегда один и может распространяться на весьма большое расстояние от тела клетки. По ходу аксона от него могут отходить многочисленные коллатерали, конечные ветви которых передают информацию другим нейронам или рабочим органам. На поверхности аксонов, как и на поверхности дендритов, располагаются синапсы. Таким образом, существует 3 вида передачи информации, или 3 вида синапсов: аксон - денд- рит, аксон - тело нейрона, аксон - аксон. Общим для всех видов синапсов является однонаправленность передачи информации, лежащая в основе структуры разнообразных рефлекторных дуг.
Среди различных типов нейронов можно выделить чувствительные, вставочные, двигательные и нейросекреторные. Чув- ствительные (сенсорные, или афферентные) нейроны всегда распо- лагаются вне ЦНС. Это так называемые псевдоуниполярные нейроны, имеющие некоторые характерные черты строения. Свое название эти нервные клетки получили потому, что оба их отростка в непосредственной близости от тела располагаются вместе и лишь затем делятся. Очень длинный по сравнению с другими нейронами дендрит начинается от периферического рецептора, а аксон вступает в ЦНС, передавая информацию на второй или вставочный нейрон.
Вставочные нейроны - это нервные клетки, на которых происходит переключение нервных импульсов на пути от первого (чувствительного) нейрона к двигательному, или моторному, нейрону. В пределах одной рефлекторной дуги может быть несколько вставочных нейронов, расположенных в различных отделах мозга.
Двигательными (моторными, или эфферентными) нейронами счи- таются нервные клетки, которые определенным образом адаптируют принятое в ЦНС решение и в итоге доводят его до органа-ис-
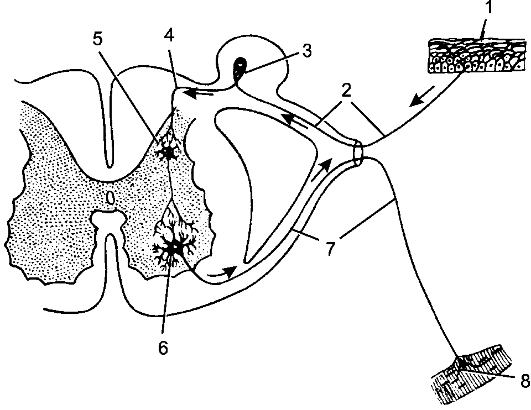
Рис. 193. Рефлекторная дуга.
1 - нервное окончание чувствительного нейрона в коже; 2 - периферический отросток чувствительного нейрона; 3 - спинномозговой узел с псевдоуниполярной клеткой; 4 - центральный отросток чувствительного нейрона; 5 - вставочный нейрон; 6 - двигательная клетка переднего рога; 7 - периферический отросток двигательной клетки; 8 - нервное окончание в мышце.
полнителя или рабочего органа. Импульсы, переданные на эфферентные нейроны, проявляются в сокращении или расслаблении скелетных и гладких мышц, в изменении тонуса мышечной ткани.
В нейросекреторных нейронах образуются, а затем поступают в кровь активные вещества, или нейрогормоны.
Миллиарды нейронов, составляющих основу нервной системы, организованы в различные по сложности рефлекторные дуги (рис. 193). Простым вариантом рефлекторной дуги является звено из двух нейронов: чувствительного и двигательного. Однако большинство рефлекторных дуг многонейронны, т.е. содержат не менее 3 нервных клеток. Такие рефлекторные дуги называются полисинаптическими. Работа рефлекторной дуги начинается от рецептора. Различные виды рецепторов подразделяют на две основные группы: внешние и внутренние. К внешним рецепторам, или экстероцепторам, относят те, которые воспринимают различные воздействия из окружающей организм среды. Внутренние рецепторы, или интероцепторы, адекватны раздражению внутренних органов (висцероцепторы), посылают сигналы от мышц, сухожилий, суставов (проприоцепторы).
Деятельность нервной системы невозможна без постоянного соблюдения принципа обратной связи. Обратная связь обеспечивает саморегуляцию работы нервной системы на основе непрерывного потока информации о соответствии выполняемого действия и поставленной задачи. Таким образом, последующий этап любого акта возможен лишь тогда, когда в ЦНС поступили нервные импульсы, свидетельствующие о выполнении предыдущего этапа.
Все контакты нервной системы, все передачи информативных нервных импульсов обеспечиваются проводящими путями. Проводящие пути нервной системы представляют собой отростки нервных клеток, которые подразделяются на миелиновые (мякотные) и безмиелиновые (безмякотные) нервные волокна. Миелиновые волокна в большей степени свойственны соматической нервной системе. Безмиелиновые волокна встречаются преимущественно в вегетативной нервной системе. Разделение нервной системы на соматическую и вегетативную (автономную) условно. К соматической нервной системе относят восприятие различных раздражений и регуляцию двигательных актов поперечно-полосатой скелетной мускулатурой. Роль вегетативной (автономной) нервной системы сводится к иннервации гладкой мускулатуры всех внутренних органов, потовых желез и сосудов. Кроме того, вегетативная система обеспечивает трофическую иннервацию органов и тканей. Соматическая нервная деятельность в отличие от вегетативной может значительно больше произвольно контролироваться и управляться. Высший отдел ЦНС - кора полушарий большого мозга - осуществляет координирующую регуляцию как соматической, так и вегетативной иннервации.
Промежуточным веществом нервной системы является нейроглия. В ЦНС различают макроглию и микроглию. Макроглия, как и собственно нервные клетки, развивается из эктодермы. Она подразделяется на астроциты, имеющие многочисленные отростки, и малоотростчатые клетки, или олигодендроциты. Функции астроцитов - опорная, репаративная, изоляция рецептивных полей, гематоэнцефалический барьер. Олигодендроглия участвует в образовании миелина и в питании нейронов. Микроглия развивается из мезодермы, и при процессах воспаления и дегенерации клетки ее превращаются в макрофаги.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ
Способностью преобразовывать воспринимаемые раздражения в нервные импульсы (рецепция) и отвечать соответствующей раз- дражителю реакцией обладают уже простейшие одноклеточные организмы (амеба, инфузория). Организм многоклеточных животных может функционировать как единое целое, быстро реагировать на изменения окружающей среды. Эти функции у многоклеточных организмов выполняют нервные клетки и нейроглия, совокупность которых образует их нервную систему.
Наиболее простая нервная система у кишечнополостных представляет собой непрерывную сеть. Вследствие такого строения раздражение любого участка поверхности организма сопровождается возбуждением всей нервной системы и животное отвечает на него движением всего тела.
Эволюция нервной системы у более высокоорганизованных представителей беспозвоночных идет по пути разделения единой нервной сети на отдельные нервные клетки, отростки которых уже не переходят непосредственно друг в друга, а контактируют посредством особых образований - синапсов (synapsis - касание, соединение). Кроме того, у этих животных происходит концентрация нервных клеток с образованием нервных узлов - первичных нервных центров (узловая нервная система).
Централизация нервной системы достигает высшего уровня у позвоночных, у которых четко определяется интегрирующее и ре- гулирующее влияние центральных нервных аппаратов на все нижележащие отделы. Процесс цефализации обусловлен возникновением и концентрацией на переднем конце тела новых рецепторных приборов, особенно дистантных рецепторов (обоняние, зрение), а также органов захватывания пищи и дыхания. Появляется и прогрессивно развивается новая функция нервной системы - накопление индивидуального опыта, что связано с появлением у позвоночных новых нервных структур.
Нервная система позвоночных (трубчатая нервная система) развивается из утолщения эктодермы дорсальной поверхности зародыша - нервной пластинки. В процессе развития нервная пластинка углубляется, образуя нервный желобок. Края желобка растут навстречу друг другу, соединяются и образуют нервную трубку. Нервная трубка полностью обособляется от кожной эктодермы,
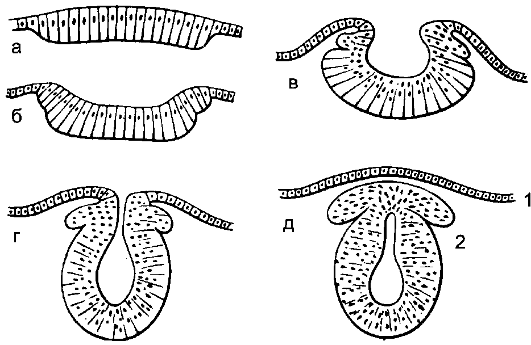
Рис. 194. Стадии эмбриогенеза нервной системы; поперечный разрез.
а - медуллярная пластинка; б, в - медуллярная бороздка; г, д - мозговая трубка;
1 - роговой листок (эпидермис); 2 - ганглиозный валик.
которая, разрастаясь, покрывает ее дорсальную поверхность. От нервной трубки отделяется группа клеток, образующих гангли- озную пластинку - зачаток нервных узлов (рис. 194).
Из нервной трубки развивается вся нервная система человека и позвоночных. Передний (краниальный) отдел нервной трубки разделяется на головной мозг и органы чувств, а из туловищного отдела образуются спинной мозг, спинномозговые и вегетативные узлы.
СПИННОЙ МОЗГ
Основные функции спинного мозга - проводниковая и рефлекторная. Нервные импульсы, содержащие информацию от эк- стероцепторов и проприоцепторов кожи, мышц, сухожилий, суставов, поступают в спинной мозг непрерывными потоками. Сюда же поступают нервные импульсы, возникающие при раздражении висцероцепторов, исключая зоны иннервации блуждающего нерва.
В спинном мозге располагаются центры иннервации поперечно-полосатой мускулатуры шеи, туловища и конечностей. Спин- ной мозг принимает активное участие в регуляции всех сложных реакций организма человека. Одни из этих реакций представляют результаты рефлекторной деятельности собственно спинного мозга, а другие являются следствием сложных процессов, протекающих в головном мозге. Таким образом, в первом случае проявляет-
ся рефлекторная, а во втором - проводниковая деятельность спинного мозга.
Приходящие в спинной мозг сигналы от различных рецепторных зон передаются по его проводящим путям к нервным центрам мозгового ствола, мозжечка и конечного мозга. Эти проводящие пути располагаются в составе его задних и боковых канатиков. Сигналы, идущие в обратном направлении, т.е. из головного мозга, проходят в составе передних и боковых канатиков спинного мозга (см. ниже).
Выходящая из головного мозга эфферентная информация оказывает возбуждающее или тормозящее действие на мотонейроны, или двигательные нейроны спинного мозга, которые в свою очередь регулируют работу поперечно-полосатых и опосредованно гладких мышц.
Следовательно, проводниковая функция спинного мозга проявляется передачей информации в виде нервных импульсов от рецеп- торных зон к отделам головного мозга, а также в обратном направлении - от головного мозга к различным рабочим органам.
РАЗВИТИЕ СПИННОГО МОЗГА
Все отделы ЦНС человека развиваются из нервной трубки, которая в результате митотического деления клеток становится многослойной. В этот период в ней можно выделить 3 слоя: наружный, или краевой, внутренний, или эпендимный, и средний, или мантий- ный (плащевой). Из клеток внутреннего слоя развиваются эпендимные клетки, которые выстилают центральный канал спинного и желудочки головного мозга. Из клеток мантийного слоя образуются нейробласты и спонгиобласты; первые развиваются в нервные клетки, а вторые - в клетки нейроглии.
Деление клеток в различных отделах туловищного конца нервной трубки происходит неодинаково интенсивно. Наиболее резко увеличивается количество клеток в ее боковых отделах, где на внут- ренней поверхности утолщенной боковой стенки появляется пограничная борозда, sulcus limitans, разделяющая боковую стенку на две пластинки: заднелатеральную, lamina dorsolateralis, и передне- латеральную, lamina ventrolateralis. Из клеток переднелатеральной пластинки позже образуются передние столбы серого вещества спинного мозга, а из клеток заднелатеральной - его задние столбы.
Неравномерный рост боковых пластинок по сравнению с передней и задней приводит к изменению формы нервной трубки. Она прогибается в области донной и покровной пластинок с образова-
нием двух продольных борозд, которые делят спинной мозг на две симметричные половины. Утолщение стенок и изменение формы спинного мозга сопровождаются резким сужением его полости - центрального канала.
СТРОЕНИЕ СПИННОГО МОЗГА
Спинной мозг, medulla spinalis, представляет собой неравномерный по толщине, сдавленный спереди назад цилиндрической формы тяж длиной 45 см у мужчин и 41-42 см у женщин. Возле верхнего края I шейного позвонка спинной мозг без резких границ переходит в продолговатый, а на уровне II поясничного позвонка заканчивается мозговым конусом, conus medullaris, тонкая вершина которого (диаметром до 2 мм) продолжается в спинномозговой части терминальной нити, pars spinalis fili terminalis, которая прикрепляется ко II копчиковому позвонку (рис. 195).
Толщина и форма спинного мозга на всем протяжении неодинаковы. В шейном и поясничном отделах имеются 2 веретенообразной формы утолщения: шейное, intumescentia cervicalis, и пояснично-крестцовое, intumescentia lumbosacralis. Поперечный диаметр спинного мозга в области шейного утолщения 13- 14 мм, в области поясничного - 12 мм, а в грудном отделе - 10 мм.
На передней поверхности спинного мозга имеется глубокая передняя срединная щель, fissura mediana anterior, кото-
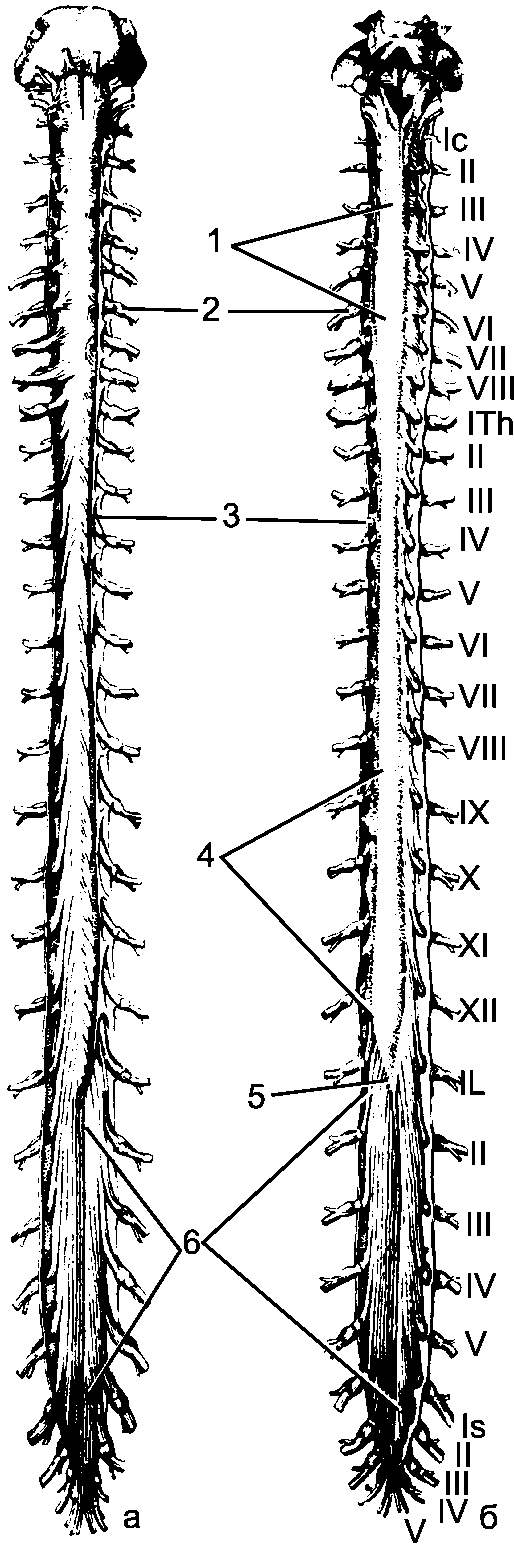
Рис. 195. Спинной мозг. Твердая и паутинная оболочки разрезаны. Сосудистая оболочка снята. Римскими цифрами обозначен порядок расположения шейных, грудных, поясничных и крестцовых спинномозговых нервов. а - передняя поверхность; б - задняя поверхность.
1 - шейное утолщение; 2 - спинномозговой узел; 3 - твердая оболочка спинного мозга; 4 - поясничное утолщение; 5 - мозговой конус; 6 - конский хвост.
рая вместе с задней срединной бороздой, sulcus medianus posterior, идущей продольно по его задней поверхности, делит мозг на две симметричные половины. На поверхности каждой половины спинного мозга отмечают слабо выраженные переднюю и заднюю латеральные борозды, sulci anterolateralis et posterolateralis, представляющие собой соответственно места выхода передних и входа задних корешков. Кроме этих борозд, в шейном и верхнегрудном отделах спинного мозга, между передней и задней латеральными бороздами, имеется задняя промежуточная борозда, sulcus intermedius posterior. Все указанные борозды делят белое вещество спинного мозга на обособленные пучки нервных волокон - канатики, funiculi medullae spinalis, которые хорошо видны на поперечных срезах. Самый узкий передний канатик, funiculus anterior, расположен между передней срединной щелью и передней латеральной бороздой; боковой канатик, funiculus lateralis, - между латеральными бороздами, а задний канатик, funiculus posterior, - между задними латеральной и срединной бороздами. В этих же отделах задний канатик подразделяется задней промежуточной бороздой на 2 пучка: медиально распо- ложенный тонкий пучок, fasciculus gracilis, и латеральный клиновидный, fasciculus cuneatus.
По бокам спинного мозга видны 2 вертикальных ряда (передний и задний) пучков нервных волокон, которые называют корешками спинномозговых нервов. Передний корешок, r. anterior, образован преимущественно аксонами двигательных клеток передних рогов спинного мозга. Задний корешок, r. posterior, составляют аксоны чувствительных клеток спинномозговых узлов. Передний корешок выходит из передней боковой борозды спинного мозга и выполняет двигательные функции. Задний корешок вступает в боковую заднюю борозду и является чувствительным. Соединяясь, передний и задний корешки образуют ствол спинномозгового нерва, который по составу волокон является смешанным. Возле места образования спинномозгового нерва на заднем корешке имеется утолщение - спинномозговой узел, ganglion spinale, представляющий собой скопление псевдоуниполярных чувствительных нервных клеток (рис. 196). Дендриты этих клеток оканчиваются на периферии рецепторными аппаратами, а аксоны составляют задние корешки, проникающие в спинной мозг. Часть образующих их волокон вступает в контакт с клетками спинного мозга, а другая часть волокон идет к ядрам продолговатого мозга. Соответственно числу пар корешков в спин- ном мозге 31 сегмент: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и копчиковый. Длина позвоночного канала превышает
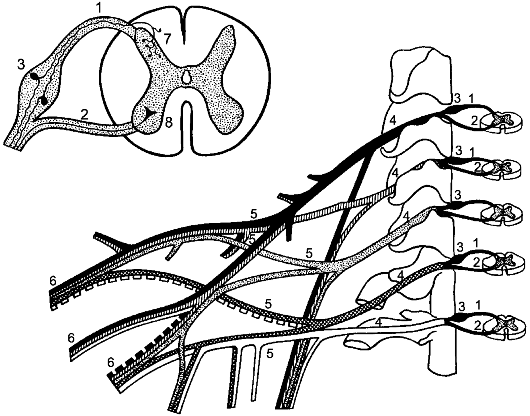
Рис. 196. Элементы периферической нервной системы.
1 - задний корешок; 2 - передний корешок; 3 - спинномозговой узел; 4 - первичный ствол нервного сплетения; 5 - нервное сплетение; 6 - вторичный ствол сплетения; 7 - задний рог; 8 - передний рог.
длину спинного мозга, и у взрослого человека сегменты спинного мозга не соответствуют уровню позвонков, их проекции не совпадают.
Поясничные (начиная с LII), крестцовые и копчиковые корешки на пути к соответствующим межпозвоночным отверстиям образуют крупный пучок, окружающий снаружи конечную нить, кото- рый называется конским хвостом, cauda equina.
Внутреннее строение спинного мозга, как и других отделов ЦНС, изучают на сериях поперечных и продольных срезов. На поперечных срезах видно, что спинной мозг образован серым и белым ве- ществом, причем серое вещество занимает центральное положение. Передняя срединная щель и соединительнотканная задняя срединная перегородка, septum medianum posterior, проникают глубоко в мозговое вещество, так что между двумя симметричными половинами спинного мозга остается узкий участок мозговой ткани, в центре которой расположен центральный канал, canalis centralis.
Являясь остатком полости первичной нервной трубки, центральный канал прослеживается на всем протяжении спинного мозга. В верхних шейных сегментах он принимает форму сагиттальной щели, которая сообщается с IV желудочком мозга. В области мозгового конуса канал смещается ближе к задней срединной борозде и заканчивается небольшим расширением - терминальным желудочком, ventriculus terminalis.
Серое вещество, substantia grisea, спинного мозга на поперечных срезах напоминает по форме бабочку или букву Н, на всем протяжении имеет вид двух неправильной формы колонн - серых столбов, columnae griseae, соединенных между собой узкой перемычкой - серой спайкой, commissura grisea. Серые столбы подразделяются на пе- редний и задний, columnae anterior et posterior. Кроме того, между VIII шейным и II или III поясничным сегментами имеется боковой выступ серого вещества, который образует боковой столб, columnae intermedia.
Выступы серого вещества на поперечных срезах называют рогами. Наиболее массивны передние рога, соти anterior. Их передние концы расширены в шейном и пояснично-крестцовых отделах спинного мозга. Задние рога, cornu posterior, более узкие и длинные.
Между передними и задними рогами расположено промежуточное вещество, центральная часть которого окружает центральный канал, а ее наружное продолжение оканчивается боковым рогом на протяжении между VIII шейным и II или III поясничными сегментами.
Идентичные по строению и функции нервные клетки серого вещества образуют различные по форме и величине скопления - ядра спинного мозга. В задних столбах расположены соматическичувствительные ядра, в передних - соматически-двигательные, а в боковых столбах - ядра вегетативной нервной системы.
Белое вещество, substantia alba, спинного мозга состоит преимущественно из продольно расположенных нервных волокон, составляющих передние, задние и боковые канатики. В образовании этих волокон участвуют отростки чувствительных клеток спинальных ганглиев, ядер спинного мозга и клеток различных отделов головного мозга. В свою очередь каждый канатик спинного мозга образует несколько различных по функциональному значению пучков нервных волокон - проводящих путей спинного мозга (рис. 197). Проводящие пути спинного мозга подразделяются на короткие и длинные. Короткие проводящие пути ассоциативные. Это пути собственного аппарата спинного мозга, посредством которых осуществ-
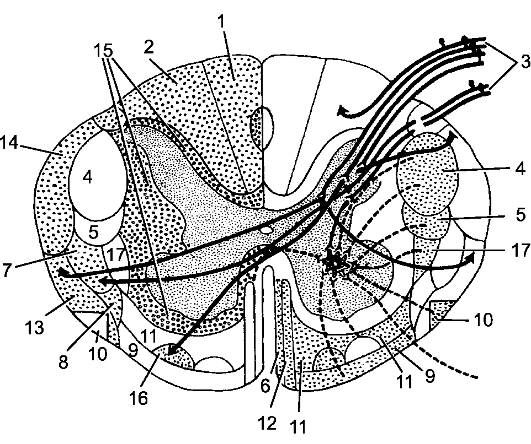
Рис. 197. Проводящие пути спинного мозга (поперечный срез). Слева показаны восходящие системы волокон, справа - топография нисходящих систем волокон. 1 - тонкий пучок; 2 - клиновидный пучок; 3 - задний корешок; 4 - латеральный корково-спинномозговой (пирамидный) путь; 5 - красноядерно-спинномозговой путь; 6 - покрышечно-спинномозговой путь; 7 - латеральный спиноталамический путь; 8 - спинопокрышечный путь; 9 - преддверно-спинномозговой путь; 10 - оливо-спинномозговой путь; 11 - ретикуло-спинномозговой путь; 12 - передний корково-спинномозговой (пирамидный) путь; 13 - передний спиномозжечковый путь; 14 - задний спиномозжечковый путь; 15 - собственные пучки; 16 - передний спиноталамический путь; 17 - таламо-спинномозговой путь.
ляется одна из основных его функций - рефлекторная. С длинными проводящими путями связана другая основная функция спинного мозга - проводниковая. Длинные проводящие пути спинного мозга разделяются на восходящие - чувствительные, которые проводят нервные импульсы в головной мозг, и нисходящие - двигательные, следующие от различных центров головного мозга к спинному. Проводящие пути имеют строго определенную локализацию.
ОБОЛОЧКИ СПИННОГО МОЗГА
Спинной мозг покрыт 3 соединительнотканными оболочками, которые развиваются из окружающей мозговую трубку мезодермы. Снаружи расположена твердая оболочка спинного мозга, dura mater spinalis. Глубже ее находится паутинная оболочка, arachnoidea mater spinalis, которая представляет собой тонкий бессосудистый листок
19
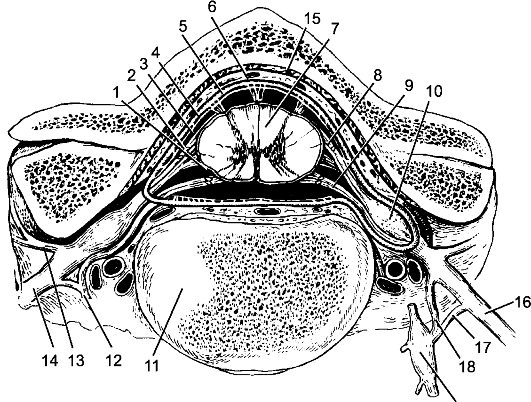
Рис. 198. Оболочки спинного мозга.
1 - мягкая оболочка; 2 - паутинная оболочка; 3 - подпаутинное пространство; 4 - субдуральное пространство; 5 - твердая оболочка; 6 - эпидуральное пространство; 7 - спинной мозг; 8 - задний корешок; 9 - передний корешок; 10 - спинномозговой узел; 11 - позвонок; 12, 18 - белые соединительные ветви; 13 - задняя ветвь спинномозгового нерва; 14 - передняя ветвь спинномозгового нерва; 15 - надкостница; 16 - спинномозговой нерв; 17 - серые соединительные ветви; 19 - узел симпатического ствола.
рыхлой волокнистой соединительной ткани. Непосредственно к веществу мозга прилегает мягкая оболочка, pia mater spinalis, которая образована также волокнистой соединительной тканью, но в отличие от паутинной оболочки содержит сети кровеносных сосудов. Все 3 оболочки в виде единого непрерывного футляра покрывают спинной мозг (рис. 198).
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
1. Каковы основные компоненты соматической рефлекторной дуги?
2. Какие основные типы нейронов вы знаете?
3. Какие виды рецепторов вам известны?
4. Какие основные функции выполняет соматическая и вегетативная (автономная) нервная система?
5. Какие основные этапы развития нервной системы у позвоночных и человека вы знаете?
6. Опишите функциональную приоритетность локализации функций в стволах спинного мозга.
7. Как распределены приводящие пути в канатике спинного мозга?
8. Какие оболочки спинного мозга вам известны? ГОЛОВНОЙ МОЗГ
ОБЩИЙ ОБЗОР СТРОЕНИЯ ГОЛОВНОГО МОЗГА
Головной мозг, encephalon, состоит из 4 основных частей: конечного мозга, telencephalon; промежуточного мозга, diencephalon; мозгового ствола, truncus encephali, включающего в себя продолговатый мозг, мост и средний мозг; мозжечка, cerebellum. Конечный, или большой, мозг разделен на 2 полушария продольной щелью большого мозга, fissura longitudinalis cerebri. В каждом полушарии различают верхнелатеральную поверхность, facies superolateralis hemispherii, медиальную поверхность, facies medialis hemispherii, направленную к продольной щели мозга, и соединяющую полушария спайку - мозолистое тело, corpus callosum. Более сложная по рельефу нижняя поверхность, facies inferior hemispherii, представлена не только самими полушариями, но и некоторыми отделами мозгового ствола и мозжечка (рис. 199).
РАЗВИТИЕ ГОЛОВНОГО МОЗГА
Головной мозг образуется из переднего отдела нервной трубки, который уже на самых ранних стадиях развития отличается от туловищного отдела своей шириной. Неравномерный рост различных участков стенки этого отдела приводит к образованию 3 расположенных друг за другом выпячиваний - первичных мозговых пузырей: переднего, prosencephalon; среднего, mesencephalon, и заднего, rhombencephalon (рис. 200). Далее передний и задний мозговые пузыри делятся на два вторичных мозговых пузыря, в результате чего возникает 5 сообщающихся между собой мозговых пузырей. Из них развиваются все отделы головного мозга: конечный, telencephalon, про- межуточный, diencephalon, средний, mesencephalon, задний, metencephalon, и продолговатый мозг, myelencephalon [medulla oblongata]. Процесс образования 5 мозговых пузырей происходит одновременно с появле-
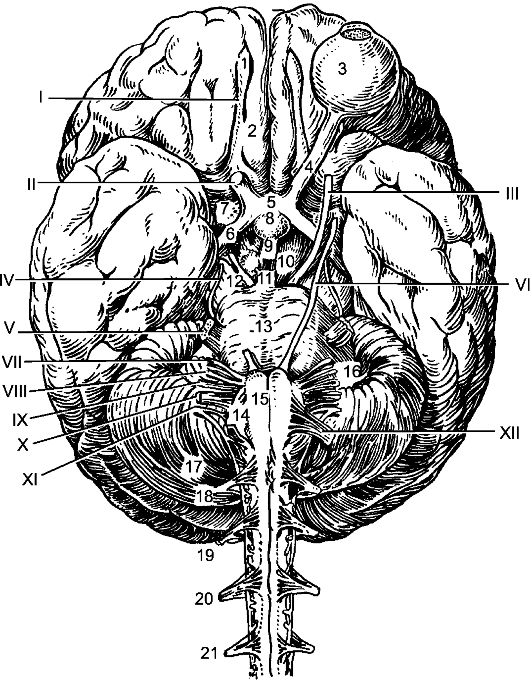
Рис. 199. Нижняя поверхность головного мозга.
I - обонятельный путь; II - зрительный нерв; III - глазодвигательный нерв; IV - блоковый нерв; V - тройничный нерв; VI - отводящий нерв; VII - лицевой нерв; VIII - преддверно-улитковый нерв; IX - языкоглоточный нерв; Х - блуждающий нерв; XI - добавочный нерв; XII - подъязычный нерв.
1 - обонятельная луковица; 2 - обонятельный путь; 3 - левое глазное яблоко; 4 - зрительный нерв; 5 - зрительный перекрест; 6 - зрительный путь; 7 - переднее продырявленное вещество; 8 - гипофиз; 9 - серый бугор; 10 - сосцевидное тело;
11 - заднее продырявленное вещество; 12 - ножка большого мозга; 13 - мост; 14 - олива; 15 - пирамида; 16 - клочок; 17 - мозжечок; 18-21 - корешки 4 верхних спинномозговых нервов.
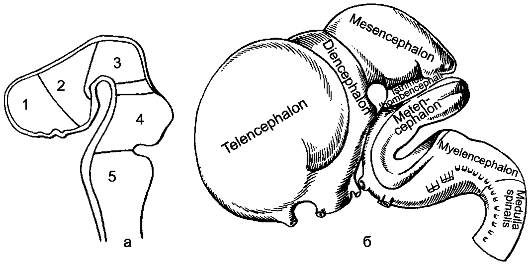
Рис. 200. Развитие головного мозга (по Р.Д. Синельникову).
а - 5 мозговых пузырей: 1 - конечный мозг; 2 - промежуточный мозг; 3 - средний мозг; 4 - собственно задний мозг; 5 - продолговатый мозг; между 3-м и 4-м пузырями - перешеек.
б - модель развивающегося мозга на стадии 5 пузырей.
нием изгибов головного отдела мозговой трубки в сагиттальном направлении. Сначала появляется дорсальный теменной изгиб в области metencephalon, затем в том же направлении - затылочный изгиб между myelencephalon и спинным мозгом и, наконец, третий вент- ральный мостовой изгиб в области metencephalon.
Этот процесс сопровождается усиленным ростом боковых отделов нервной трубки и отставанием в росте дорсальной и вентраль- ной стенок. Утолщенные боковые отделы разделяются пограничной бороздой на основную и крыльную пластинки, из которых нейробласты основной пластинки образуют двигательные, а нейробласты крыльной - чувствительные центры. Между обеими пластинками, в промежуточной зоне, располагаются вегетативные центры.
Наиболее значительные дифференцировка и изменения в форме наблюдаются при развитии производных переднего мозгового пузыря - telencephalon и diencephalon.
ПРОДОЛГОВАТЫЙ МОЗГ
Продолговатый мозг, medulla oblongata, представляет собой отдел мозгового ствола длиной в среднем 25 мм, который является непосредственным продолжением спинного мозга и по форме напоминает усеченный конус (рис. 201). Утолщенный верхний отдел продолговатого мозга переходит в мост. Нижней границей продолговатого мозга считают место выхода корешков I пары шейных нер-
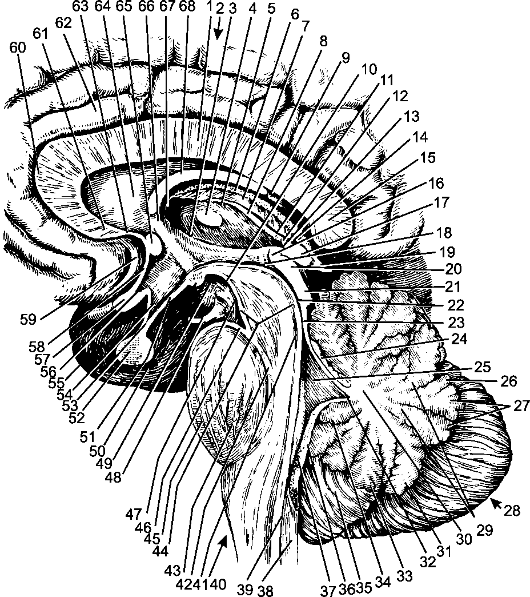
Рис. 201. Мозговой ствол; сагиттальный разрез.
1 - ствол мозолистого тела; 2 - кора большого мозга (плащ); 3 - гипоталамическая борозда; 4 - межталамическое сращение; 5 - таламус; 6 - сосудистое сплетение III желудочка; 7 - сосудистая основа III желудочка; 8 - мозговая полоска таламуса; 9 - ножка свода; 10 - заднее продырявленное вещество; 11 - поводок; 12 - спайка поводков; 13 - шишковидное углубление; 14 - надшишковидное углубление; 15 - валик мозолистого тела; 16 - задняя спайка; 17 - шишковидное тело; 18 - водопровод мозга; 19 - большая мозговая вена; 20 - верхний холмик; 21 - нижний холмик; 22 - водопровод мозга; 23 - верхний мозговой парус; 24 - язычок мозжечка; 25 - IV желудочек; 26 - листок червя; 27 - бугор червя; 28 - мозжечок; 29 - белые полоски; 30 - мозговое тело; 31 - пирамида червя; 32 - шатер; 33 - язычок червя; 34 - узелок червя; 35 - миндалина червя; 36 - сосудистая основа IV желудочка; 37 - сосудистое сплетение IV желудочка; 38 - спинной мозг; 39 - центральный канал; 40 - продолговатый мозг; 41 - ядро блокового нерва; 42 - медиальный продольный пучок; 43 - продольный пирамидный пучок; 44 - ножки мозга; 45 - мост; 46 - межножковое заднее углубление; 47 - межножковая ямка; 48 - глазодвигательный нерв; 49 - межножковое переднее углубление; 50 - сосцевидное тело; 51 - серый бугор; 52 - гипофиз; 53 - воронка; 54 - углубление воронки; 55 - височная доля; 56 - зрительное углубление; 57 - зрительный перекрест; 58 - терминальная пластинка; 59 - борозда подмозолистого тела; 60 - колено подмозолистого тела; 61 - клюв мозолистого тела; 62 - поясная извилина; 63 - пластинка клюва; 64 - прозрачная перегородка; 65 - передняя спайка мозга; 66 - столб свода; 67 - межжелудочковое отверстие; 68 - тело свода.
вов, что соответствует уровню большого затылочного отверстия. Верхней границей на передней поверхности является нижний край моста, а на задней - мозговые полоски ромбовидной ямки.
На всем протяжении продолговатого мозга проходят продольные борозды, продолжающиеся в соответствующие борозды спинного мозга: fissura mediana anterior, sulcus medianus posterior, sulcus anterolateralis, sulcus posterolateralis. На передней поверхности про- долговатого мозга, между передней срединной щелью и передней латеральной бороздой, находится парное возвышение, которое называют пирамидой, pyramis (рис. 202). Пирамиды состоят из волокон двигательных произвольных путей. Большая часть волокон этих путей на расстоянии 6-7 мм от нижней границы продолговатого мозга переходит на противоположную сторону бокового канатика спинного мозга, образуя латеральный корково-спинномозговой путь, tractus corticospinalis lateralis, а оставшиеся неперекрещенными волокна, находясь в переднем канатике, составляют передний корково-спинномозговой путь, tractus corticospinalis anterior. Перекрест двигательных нервных волокон, или перекрест пирамид, decussatio pyramidum, прерывает переднюю срединную щель. Кнаружи от каждой пирамиды, между передней и задней латеральными бороздами, расположено овальной формы возвышение - олива, oliva.
Из передней латеральной борозды между пирамидой и оливой выходят корешки подъязычного нерва. Позади оливы из задней латеральной борозды выходят корешки языкоглоточного, блуждающего и добавочного черепных нервов.
Нижний отдел задней поверхности продолговатого мозга напоминает по рельефу спинной мозг (рис. 203). Здесь, кнаружи от задней срединной борозды, находятся задние канатики, разделенные промежуточной бороздой, sulcus intermedius, на медиальный тонкий пучок, fasciculus gracilis, и латеральный клиновидный, fasciculus cuneatus. В верхнем отделе задней поверхности задние канатики заканчиваются утолщениями - бугорками тонкого и клиновидного ядер, tuberculi gracile et cuneatum. Клетки этих ядер являются вторыми нейронами проводящих путей задних канатиков.
Кнаружи от клиновидного канатика имеется возвышение, называемое тройничным бугорком, tuberculum trigeminale. Он представляет собой ядро спинномозгового пути тройничного нерва, nucleus spinalis nervi trigemini. Это ядро обособлено тонким слоем волокон - аксо- нов нейронов чувствительного узла тройничного нерва, ganglion trigeminale, которые составляют спинномозговой путь тройничного нерва, tractus spinalis nervi trigemini.
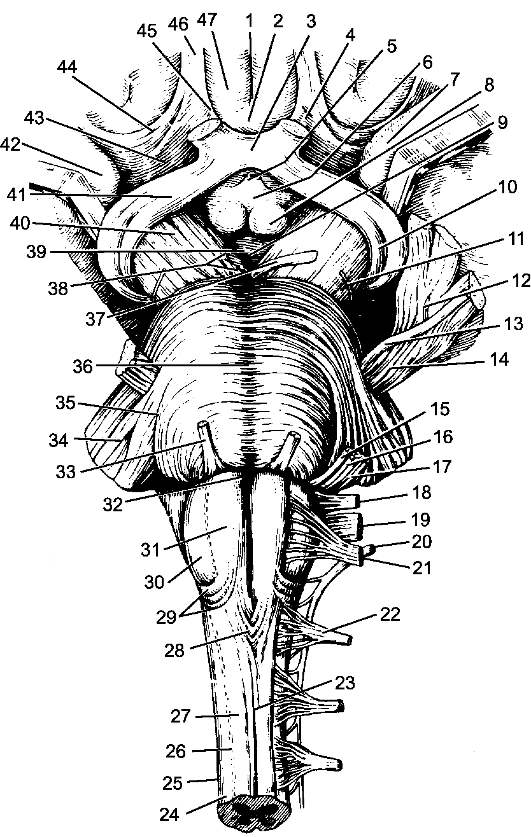
Рис. 202. Передняя поверхность продолговатого мозга, моста и среднего мозга. 1 - продольная щель большого мозга; 2 - подмозолистая борозда; 3 - зрительный перекрест; 4 - обонятельный треугольник; 5 - воронка; 6 - серый бугор; 7 - крючок (парагиппокампальная извилина); 8 - сосцевидное тело; 9 - межножковая ямка; 10, 41 - зрительный тракт; 11 - блоковый нерв; 12 - тройничный узел; 13 - двигательный корешок тройничного нерва; 14 - чувствительный корешок тройничного нерва; 15 - лицевой нерв; 16 - промежуточный нерв; 17 - преддверно-улитковый нерв; 18 - языкоглоточный нерв; 19 - блуждающий нерв; 20 - добавочный нерв; 21 - подъязычный нерв; 22 - I шейный нерв (передний корешок); 23 - передняя срединная щель; 24 - боковой канатик; 25 - спинной мозг; 26 - передняя латеральная борозда; 27 - передний канатик; 28 - перекрест пирамид; 29 - передние наружные дугообразные волокна; 30 - олива; 31 - пирамида (продолговатого мозга); 32 - слепое отверстие; 33 - отводящий нерв; 34 - средняя мозжечковая ножка; 35 - косой пучок моста; 36 - базилярная борозда; 37 - глазодвигательный нерв; 38 - корешок глазодвигательного нерва; 39 - заднее продырявленное вещество; 40 - ножка мозга; 42 - крючок; 43 - переднее продырявленное вещество; 44, 45 - латеральная и медиальная обонятельные полоски; 46 - обонятельный тракт; 47 - прямая извилина.
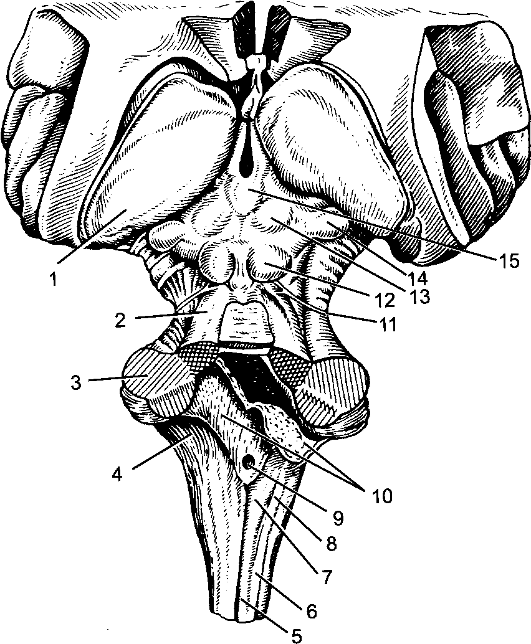
Рис. 203. Мозговой ствол.
1 - подушка таламуса; 2 - верхняя мозжечковая ножка; 3 - средняя мозжечковая ножка; 4 - нижняя мозжечковая ножка; 5 - тонкий пучок; 6 - клиновидный пучок; 7 - бугорок тонкого пучка; 8 - бугорок клиновидного пучка; 9 - срединная апертура IV желудочка; 10 - сосудистая основа IV желудочка; 11 - блоковый нерв; 12 - нижний холмик; 13 - верхний холмик; 14 - медиальное коленчатое тело; 15 - шишковидное тело.
Бугорки тонких ядер и 2 расходящихся в стороны довольно массивных пучка волокон - нижние мозжечковые ножки, pedunculi cerebellares inferiores, ограничивают с боков часть ромбовидной ямки - дна IV желудочка мозга. Нижние мозжечковые ножки образованы волокнами, которые соединяют мозжечок со спинным и продолговатым мозгом. Наружный отдел нижних мозжечковых ножек состоит из волокон заднего спиномозжечкового пути, небольшой части волокон переднего спиномозжечкового пути, волокон от олив про- долговатого мозга, преимущественно противоположной стороны, - оливомозжечкового пути, tractus olivocerebellaris, и небольшой части волокон от ядер задних канатиков. Внутренний отдел нижних ножек мозжечка образован в основном эфферентными нервными во-
локнами к вестибулярным ядрам. Кроме того, в их состав входят афферентные волокна от вестибулярных ядер к мозжечку.
Продолговатый мозг состоит из серого и белого вещества. Серое вещество располагается в виде отдельных, различных по форме и величине скоплений тел нервных клеток - ядер продолговатого мозга. Они подразделяются на ядра черепных нервов и так называемые переключательные ядра.
В заднем отделе продолговатого мозга расположены ядра подъязычного, добавочного, блуждающего, языкоглоточного, преддверно-улиткового черепных нервов и ядро спинномозгового пути тройничного нерва.
Переключательные ядра продолговатого мозга представляют собой массивные скопления клеток. Наиболее крупные из этих ядер оливы - эллипсовидные образования длиной до 10 мм, состоящие из серого и белого вещества.
Кнутри от оливного ядра находится медиальное добавочное оливное ядро, а кзади расположено заднее добавочное оливное ядро. Весь этот комплекс ядер является промежуточным центром равновесия.
Ядра задних канатиков - тонкое и клиновидное, nuclei gracilis et cuneatus, представляют собой скопления клеток длиной до 13 мм. В этих ядрах заканчиваются центральные отростки первых чувстви- тельных нейронов, которые образуют тонкий и клиновидный пучки спинного мозга и проводят импульсы глубокой и отчасти тактильной чувствительности. Отростки клеток этих ядер - вторые нейроны этого пути выше места перекреста пирамид в виде внутренних дугообразных волокон, fibrae arcuatae internae, обходят спереди центральный канал, образуя перекрест медиальных петель, decussatio lemnisci medialis. После перекреста эти волокна образуют медиальную петлю, lemniscus medialis, которая проходит, увеличиваясь в объеме, через все отделы ствола к таламусу. Увеличение объема медиальной петли происходит в результате присоединения к ней пучков нервных волокон болевой и температурной чувствительности (спиноталамический путь), а также волокон от ядра спинномозгового пути тройничного нерва. Кроме ядер черепных нервов и переключательных ядер, к серому веществу продолговатого мозга относится центрально расположенная ретикулярная формация. Ретикулярная формация имеет большое значение в регуляции возбудимости и тонуса различных отделов ЦНС, обеспечивает готовность различных центров к деятельности, усиливает или тормозит рефлекторную деятельность спинного мозга (tractus reticulospinalis), поддерживает тонус коры полушарий большого мозга.
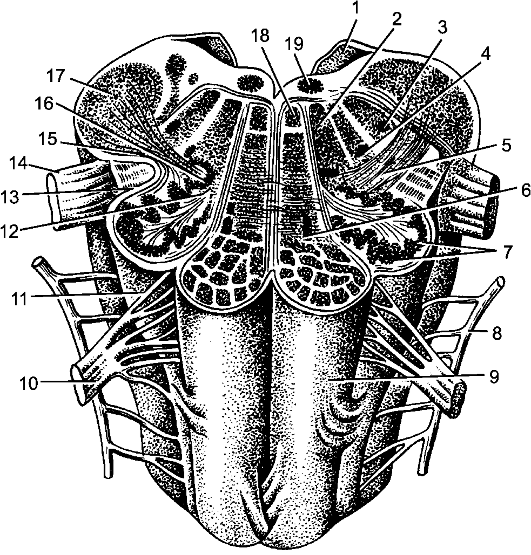
Рис. 204. Продолговатый мозг; горизонтальный разрез на уровне оливы. 1 - нижний мозговой парус; 2 - ретикулярная формация; 3 - ядро спинномозгового пути тройничного нерва; 4 - двойное ядро; 5 - оливо-спинномозговой путь; 6 - нисходящие пути пирамиды; 7 - нижнее оливное ядро; 8 - добавочный нерв; 9 - пирамида; 10 - подъязычный нерв; 11 - олива; 12 - ворота нижнего оливного ядра; 13 - покрышечно-спинномозговой путь; 14 - блуждающий нерв; 15 - красноядерно-спинномозговой путь; 16 - оливо-мозжечковый путь; 17 - нижняя мозжечковая ножка; 18 - медиальный продольный пучок; 19 - ядро подъязычного нерва.
Белое вещество продолговатого мозга состоит из пучков собственных и проходящих через него нервных волокон. Собственные, или эндогенные, волокна подразделяются на короткие и длинные. Короткие волокна соединяют ядра, расположенные в пределах продолговатого мозга. Длинные эндогенные волокна являются отростками клеток ядер продолговатого мозга, которые заканчиваются в других отделах нервной системы. К этой группе относятся волокна медиальной петли, оливо-спинномозговые, оливомозжечковые и ретикулярно-спинномозговые пути (рис. 204).
Проходящие через продолговатый мозг пучки волокон, которые не образуются и не оканчиваются в его ядрах, носят название эк-
зогенных. Среди них различают эфферентные (корково-спинномозговые, красноядерно-спинномозговые, покрышечно-спинно- мозговые) и афферентные (передний и боковой спиноталамический пути, передний и задний спиномозжечковые, спинопокрышечный пути).
МОСТ
Мост, pons, находится между продолговатым и средним мозгом. Его передняя поверхность образована выпуклостью белого цвета, которая снаружи состоит из поперечно расположенных волокон. Задняя поверхность моста участвует в образовании дна IV желудочка - ромбовидной ямки, образуя ее верхний треугольник. Эта часть ромбовидной ямки ограничена верхними мозжечковыми ножками. Поперечные волокна передней поверхности образуют средние мозжечковые ножки, которые погружаются в толщу полушарий мозжечка. Границей между мостом и средней мозжечковой ножкой служит linea trigeminofacialis, проходящая между корешками тройничного и лицевого нервов. Посередине передней поверхности моста расположена базилярная борозда, sulcus basilaris. В боковом отделе передней поверхности моста, ближе к его переднему краю, находятся корешки тройничного нерва. В области мостомозжечкового угла, образованного продолговатым мозгом, мостом и мозжечком, расположены корешки лицевого, промежуточного и преддверно- улиткового нервов, а ближе к средней линии, между задним краем моста и пирамидами, - корешки отводящих нервов.
На поперечных срезах моста различают крупную покрышку моста, tegmentum pontis, и меньшую базилярную часть моста, pars basilaris pontis, границей между которыми служит пучок поперечных волокон - трапециевидное тело, corpus trapezoideum. Передняя и задняя части моста образованы серым и белым веществом, но передняя часть состоит преимущественно из белого вещества.
Серое вещество покрышки моста - это многочисленные собственные ядра моста, nuclei pontis. В этих ядрах заканчиваются корково-мостовые пути и коллатерали от корково-спинномозговых путей. Волокна клеток ядер моста образуют его поперечные волокна, которые почти все переходят на противоположную сторону, составляют средние мозжечковые ножки и заканчиваются в клетках коры полушарий мозжечка. Поперечные волокна передней части моста образуют поверхностный и глубокий слои, между которыми проходят пучки корково-спинномозговых путей.
Серое вещество базилярной части моста состоит из центрально расположенной ретикулярной формации, ядер черепных нервов и переключательных ядер. Ретикулярная формация моста является непосредственным продолжением одноименного образования продолговатого мозга.
В мосту расположены ядра отводящего, лицевого, тройничного, преддверно-улиткового черепных нервов.
Белое вещество моста состоит из эндогенных и экзогенных волокон. Короткие эндогенные волокна соединяют между собой отдельные ядра моста и не выходят за его пределы. Так, например, установлены нервные связи между ядрами лицевого и тройничного нервов, посредством которых замыкаются рефлекторные дуги при различных раздражениях кожи лица с реакцией мимических мышц. Длинные эндогенные волокна являются отростками клеток ядер моста и заканчиваются в других отделах ЦНС. К этой группе относятся волокна, идущие от собственных ядер моста к мозжечку, поперечные волокна моста, волокна латеральной петли, пучки волокон от некоторых черепных нервов. Поперечные волокна моста образуют среднюю мозжечковую ножку, посредством которой передается влияние коры полушарий большого мозга на деятельность мозжечка.
Экзогенные волокна моста представлены эфферентными и афферентными путями. Эти пути участвуют в образовании белого вещества продолговатого мозга. Кроме них, в белое вещество моста входят корково-ядерные волокна, корково-мостовые, теменно-височно-мостовые и лобно-мостовые.
МОЗЖЕЧОК
Мозжечок, cerebellum, тесно связан с продолговатым мозгом, мостом и средним мозгом, он располагается кзади от названных образований, заполняет большую часть задней черепной ямки. Масса мозжечка около 150 г. Его наибольший поперечный размер равен 10-12 см, продольный в области червя - до 4 см, а в области полушарий - до 6 см.
Тело мозжечка, corpus cerebelli, составляют центральная узкая часть - червь, vermis cerebelli, и две крупные выпуклые боковые части - по- лушария, hemispheri cerebelli. Червь от полушарий отделяют правая и левая продольные щели, которые лучше выражены на нижней поверхности мозжечка. На верхней поверхности щели неглубокие и прерывистые, в связи с чем граница между червем и полушарием здесь выражена слабо. Верхняя поверхность мозжечка более ровная,
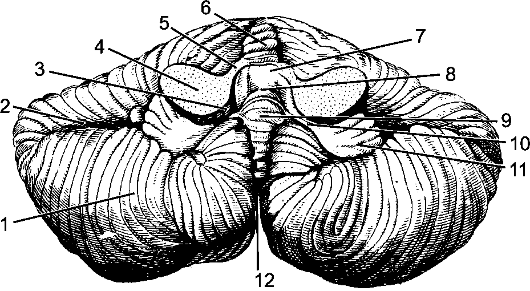
Рис. 205. Мозжечок; вид снизу.
1 - полушарие мозжечка; 2 - горизонтальная щель; 3 - нижний мозговой парус; 4 - средняя мозжечковая ножка; 5 - верхняя мозжечковая ножка; 6 - червь; 7 - верхний мозговой парус; 8 - IV желудочек; 9 - узелок; 10 - ножка клочка; 11 - клочок; 12 - задняя вырезка мозжечка.
чем нижняя, т.е. полушария выступают книзу больше, чем червь, поэтому между их внутренними поверхностями и червем образуется широкое и довольно глубокое пространство - долинка мозжечка, vallecula cerebelli. Посередине долинки имеется глубокая, направленная кверху и кзади выемка, соответствующая крыше IV желудочка. В области переднего, наиболее широкого отдела долинки расположен задний отдел среднего мозга, а книзу - мост и продолговатый мозг.
Различной глубины многочисленные щели, fissurae cerebelli, разделяют поверхность полушарий и червя мозжечка на доли, lobi cerebelli, дольки, lobuli cerebelli, и листки, folia cerebelli. Множество щелей значительно увеличивает поверхность мозжечка. Глубокие щели разделяют тело мозжечка на 3 доли: переднюю, lobus cerebelli anterior; заднюю, lobus cerebelli posterior; и клочково-узелковую, lobus flocculonodularis. В свою очередь каждая из долей щелями, проходящими через червь и полушария, разделяется на дольки; каждая долька червя переходит в правую и левую дольки полушарий (рис. 205).
Серое вещество мозжечка сосредоточено преимущественно на его поверхности в виде трехслойной коры, cortex cerebelli. Под корой находится белое вещество, которое носит название мозгового тела, corpus medullare. Внутри мозгового тела расположены парные подкорковые ядра серого вещества.
Кора покрывает как свободную поверхность извилин мозжечка, так и поверхность, расположенную в глубине борозд. Она состоит из 3 слоев: светлого наружного - молекулярного, stratum moleculare,
слоя грушевидных нейронов, stratum neuronorum piriformium, и темного внутреннего - зернистого, stratum granulosum. Грушевидные нейроны являются эфферентными клетками коры, в то время как клетки молекулярного и зернистого слоев - вставочными и ассо- циативными нейронами.
Подкорковые ядра мозжечка представляют собой различной формы и величины скопления серого вещества. Из них наиболее крупным является зубчатое ядро, nucleus dentatus, которое расположено в нижнем отделе мозгового тела. Зубчатое ядро представляет собой складчатую пластинку, которая не замкнута с внутренней стороны и окаймляет центрально расположенное белое вещество. К зубчатому ядру подходят аксоны грушевидных клеток полушарий мозжечка и сравнительно небольшое количество аксонов от грушевидных клеток червя. Отростки клеток зубчатого ядра образуют его центрально расположенное белое вещество, выходят через ворота и далее составляют основную часть волокон верхней мозжечковой ножки, pedunculus cerebellaris superior.
Пробковидное ядро, nucleus emboliformis, расположено в белом веществе полушария, медиальное верхнего края зубчатого ядра.
Самое медиальное положение среди подкорковых ядер мозжечка, над шатром IV желудочка, занимает ядро шатра, nucleus fastigii.
Между пробковидным ядром и ядром шатра расположено шаровидное ядро, nucleus globosus.
Белое вещество мозжечка состоит из внутри- и внемозжечковых волокон, которые составляют мозговое тело мозжечка и центральную часть его листков.
Группу внутримозжечковых волокон образуют отростки клеток коры мозжечка. Среди них различают ассоциативные волокна, ко- торые соединяют между собой различные участки коры мозжечка; комиссуральные волокна, соединяющие участки коры противоположных полушарий; короткие проекционные волокна - отростки грушевидных клеток к подкорковым ядрам мозжечка.
К внемозжечковым волокнам относятся длинные проекционные эфферентные и афферентные волокна, посредством которых мозжечок соединен с другими отделами головного мозга. Эти волокна формируют 3 пары мозжечковых ножек, из которых нижние и средние состоят преимущественно из афферентных, а верхние - из эфферентных волокон, образующихся в подкорковых ядрах мозжечка. В составе нижних ножек в мозжечок вступают задний спиномозжеч- ковый путь, наружные дугообразные волокна, fibrae arcuatae externae, волокна от вестибулярных ядер к ядру шатра, волокна от оливы -
оливомозжечковый путь, tractus olivocerebellaris. Кроме того, сюда входит путь от ядра шатра к латеральному вестибулярному ядру.
Средние мозжечковые ножки образуют поперечные волокна моста, которые соединяют ядра моста с корой мозжечка.
В составе верхних мозжечковых ножек в мозжечок вступает передний спиномозжечковый путь, отростки клеток зубчатого ядра, которые после перекреста заканчиваются в красном ядре среднего мозга и таламусе.
IV ЖЕЛУДОЧЕК
IV желудочек, ventriculus quartus, образован продолговатым мозгом, мостом и мозжечком. В нем различают дно, боковые стенки и крышу. Дно IV желудочка составляет ромбовидная ямка, fossa rhomboidea, которая представляет собой ограниченный верхними и нижними мозжечковыми ножками ромбовидной формы участок задней поверхности моста и продолговатого мозга (рис. 206). Верхний угол ромбовидной ямки переходит в водопровод среднего мозга. Нижний угол прикрыт небольшой пластинкой - задвижкой, obex, и продолжается в центральный канал спинного мозга. Боковые углы ромбовидной ямки переходят в латеральные карманы, recessus lateralis, IV желудочка. Срединная борозда, sulcus medianus, делит ромбовидную ямку на две симметричные половины.
Боковые стенки IV желудочка представлены 3 парами мозжечковых ножек. Крышу IV желудочка, tegmen ventriculi quarti, образуют верхний мозговой парус, вещество мозжечка и нижний мозговой парус.
Верхний мозговой парус, velum medullare superius, представляет собой тонкую треугольную пластинку белого вещества, которая расположена между обеими верхними мозжечковыми ножками и образует верхнюю часть крыши IV желудочка.
Нижний мозговой парус, velum medullare inferius, образован тонкой эпителиальной пластинкой - остатком задней стенки первичного мозгового пузыря. Эта пластинка дополняется сосудистой основой IV желудочка, tela choroidea ventriculi quarti. Нижний мозговой парус хорошо виден после удаления миндалины мозжечка.
По средней линии нижнего мозгового паруса расположено отверстие - срединная апертура IV желудочка, apertura mediana ventriculi quarti, апертуры латеральные IV желудочка, aperturae laterales ventriculi quarti. Указанные отверстия соединяют между собой полость IV желудочка и субарахноидальное пространство головного мозга. Кроме tela choroidea ventriculi quarti, в полости IV желудочка имеет-
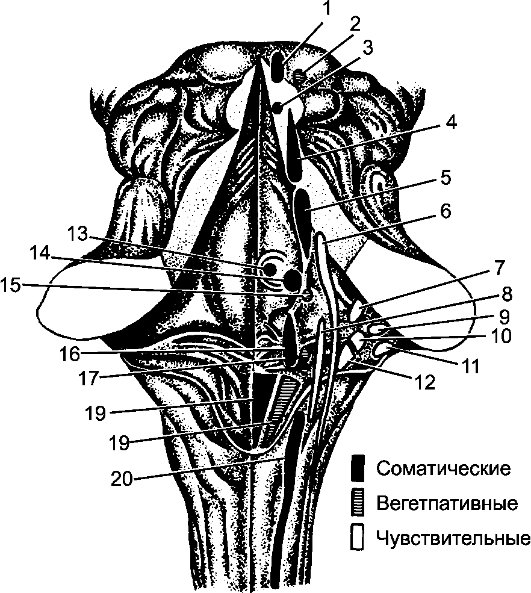
Рис. 206. Ромбовидная ямка и ядра черепных нервов.
1 - ядро глазодвигательного нерва; 2 - добавочное ядро глазодвигательного нерва; 3 - ядро блокового нерва; 4 - ядро среднемозгового пути тройничного нерва; 5 - двигательное ядро тройничного нерва; 6 - мостовое ядро тройничного нерва; 7 - верхнее вестибулярное ядро; 8 - ядро одиночного пути; 9 - переднее улитковое ядро; 10 - латеральное вестибулярное ядро; 11 - заднее улитковое ядро; 12 - медиальное вестибулярное ядро; 13 - ядро отводящего нерва; 14 - ядро лицевого нерва; 15 - верхнее слюноотделительное ядро; 16 - двойное ядро; 17 - нижнее слюноотделительное ядро; 18 - ядро подъязычного нерва; 19 - заднее ядро блуждающего нерва; 20 - ядро добавочного нерва.
ся сосудистое сплетение, plexus choroideus ventriculi quarti, которое в виде сильно извитых сосудов располагается вдоль средней линии, проходит в боковые карманы и выходит наружу через латеральные апертуры IV желудочка в области мостомозжечкового угла.
СРЕДНИЙ МОЗГ
Средний мозг, mesencephalon, расположен между мостом и промежуточным мозгом. В его состав входят ножки мозга и крыша среднего мозга.
Ножки мозга, pedunculi cerebri, представляют собой 2 расходящихся под острым углом массивных валика, которые образованы продольно ориентированными нервными волокнами. Между ножками мозга находится межножковая ямка, fossa iterpeduncularis. Она закрыта тонкой пластинкой, пронизанной множеством отверстий для кровеносных сосудов, - задним продырявленным веществом, substantia perforata posterior.
Крыша среднего мозга, tectum mesencephali, составляет его задний отдел, который скрыт под полушариями большого мозга. Пластинка крыши, lamina tecti, продольной и поперечной бороздами делится на два верхних и два нижних холмика, colliculi superiores et inferiores. В переднем отделе продольной борозды находится шишковидная железа, а от ее заднего конца отходят волокна, образующие уздечку верхнего мозгового паруса. Наружная поверхность каждого холмика переходит в пучок волокон, который называют ручкой холмика, brachium colliculi. Ручка верхнего холмика проходит в область промежуточного мозга к латеральному коленчатому телу, corpus geniculatum laterale, а часть ее волокон - в зрительный тракт. Ручка нижнего холмика вступает в медиальное коленчатое тело.
Полостью среднего мозга является узкий канал длиной около 2 см - водопровод среднего мозга, aqueductus mesencephali. Этот канал выстлан эпендимой и соединяет между собой IV и III желудочки мозга.
На поперечных срезах среднего мозга выделяют 3 отдела: крышу среднего мозга, заднюю часть ножки мозга - покрышку, tegmentum mesencephali, и переднюю часть - основание ножки мозга, basis pedunculi cerebralis. Границей между покрышкой и основанием ножки мозга является черное вещество, substantia nigra. Основание ножки мозга образовано белым веществом, которое состоит из продольных эфферентных путей корково-спинномозговых, корково-мостовых, корково-ядерных, теменно-височно-мостовых и лобно-мостовых волокон. Покрышку и крышу среднего мозга наряду с белым веществом образуют ядра серого вещества, причем белое вещество покрышки состоит как из эфферентных (красноядерно-спинномозговой путь), так и афферентных (медиальная и латеральная петли) проводящих путей.
Серое вещество крыши среднего мозга составляют ядра верхних и нижних холмиков.
Ядро нижнего холмика, nucleus colliculi inferioris, является первичным слуховым рефлекторным центром. В нем заканчивается часть волокон латеральной петли. Отростки клеток этого ядра образуют
ручку нижнего холмика, которая вступает в медиальное коленчатое тело, а часть волокон входит в состав покрышечно-спинномозгового и покрышечно-бульбарного путей, заканчивающихся в двигательных ядрах мозгового ствола и спинного мозга. При участии ядер нижних холмиков осуществляются двигательные, ориентировочные и оборонительные рефлексы на звуковые раздражения.
Ядра верхних холмиков - слои (серый и белый) верхнего холмика, strata (grisea et alba) colliculi superioris, являются первичными зрительными рефлекторными центрами. В них заканчивается часть волокон зрительного тракта, а также волокна от спинного мозга, идущие в составе спинопокрышечного пути, и ответвления латеральной и медиальной петель. Клетки этих ядер образуют основную массу волокон покрышечно-спинномозгового и покрышечнобульбарного путей, которые, как известно, заканчиваются в двигательных ядрах мозгового ствола и спинного мозга. Они осуществляют двигательные, ориентировочные и оборонительные рефлексы на световые раздражения.
Серое вещество покрышки среднего мозга представлено несколькими ядрами и ретикулярной формацией, которая является продолжением кпереди ретикулярной формации моста. В окружающем водопровод среднего мозга центральном сером веществе выделяют значительные по длине (5-6 мм) ядра глазодвигательных нервов. Эти парные ядра расположены кпереди от водопровода среднего мозга, на уровне его верхних холмиков. Верхний конец этих ядер заходит в область промежуточного мозга.
На уровне верхней части нижних холмиков крыши среднего мозга расположены парные ядра блокового нерва.
Самым длинным из ядер черепных нервов среднего мозга является ядро среднемозгового пути тройничного нерва, nucleus tractus mesencephalici nervi trigemini.
От клеток парного красного ядра, nucleus ruber, расположенного в покрышке среднего мозга, начинается красноядерно-спинномозго- вой путь, tractus rubrospinalis, который после перекреста в среднем мозге заканчивается в двигательных ядрах спинного мозга. Вместе с ретикулярной формацией мозгового ствола красные ядра осуществляют регуляцию мышечного тонуса, в которой принимает участие также черное вещество, расположенное в ножках мозга. Это вещество образовано клетками, содержащими черный пигмент - меланин. Черное вещество прослеживается на всем протяжении среднего мозга между основанием ножки мозга и покрышкой. Наряду с регуляцией мышечного тонуса черное вещество участвует
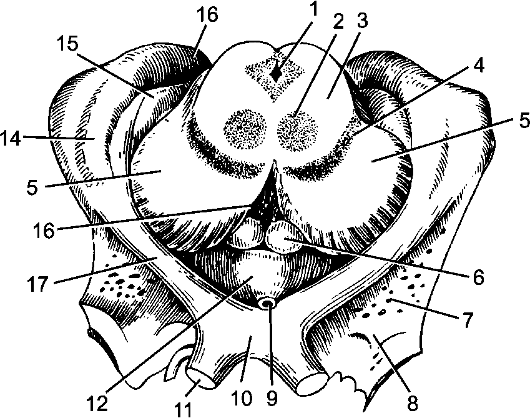
Рис. 207. Метаталамус и гипоталамус, средний мозг.
1 - водопровод среднего мозга; 2 - красное ядро; 3 - покрышка; 4 - черное вещество; 5 - ножки мозга; 6 - сосцевидное тело; 7 - переднее продырявленное вещество; 8 - обонятельный треугольник; 9 - воронка; 10 - зрительный перекрест; 11 - зрительный нерв; 12 - серый бугор; 13 - заднее продырявленное вещество; 14 - латеральное коленчатое тело; 15 - медиальное коленчатое тело; 16 - подушка; 17 - зрительный тракт.
также в координации сложных двигательных актов, таких, как жевание, глотание (рис. 207).
Белое вещество среднего мозга образовано короткими и длинными эндогенными волокнами и длинными экзогенными.
Ножку мозга образуют длинные экзогенные эфферентные волокна, из которых корково-спинномозговые и корково-ядерные пути занимают центральную часть этого образования. Кнаружи и кнутри от них расположены корково-мостовые пути. В покрышке среднего мозга проходят различные восходящие и нисходящие пучки волокон, среди которых длинными афферентными экзогенными являются волокна латеральной и медиальной петель. Длинные эн- догенные нервные волокна возникают в ядрах среднего мозга: красном ядре (красноядерно-спинномозговой путь), ядрах крыши среднего мозга (покрышечно-спинномозговой путь) и других, а короткие эндогенные волокна соединяют ядра среднего мозга между собой.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг, diencephalon, анатомически представляет собой следующую за мозговым стволом часть головного мозга.
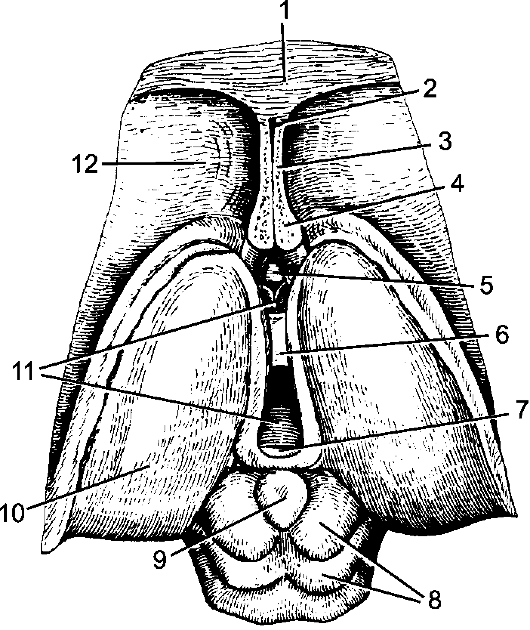
Рис. 208. Промежуточный и средний мозг; вид сверху.
1 - мозолистое тело; 2 - полость прозрачной перегородки; 3 - прозрачная перегородка; 4 - свод (поперечный разрез столби- ков); 5 - передняя спайка; 6 - межталамическое сращение; 7 - задняя спайка; 8 - крыша среднего мозга; 9 - шишковидное тело; 10 - таламус; 11 - III желудочек; 12 - хвостатое ядро (головка).
В процессе эмбриогенеза промежуточный мозг образуется из переднего мозгового пузыря. Филогенетически различают более «молодые» отделы промежуточного мозга: передний таламус, задний таламус, метаталамус, эпиталамус и относительно «старый» гипоталамус. «Молодые» отделы промежуточного мозга часто объединяют термином «таламический мозг», или «таламус».
Таламус, thalamus, представ- ляет собой парное массивное образование яйцевидной формы, на переднем конце которого имеется бугорок, а задний утолщенный конец носит название подушки, pulvinar (рис. 208). Латеральная поверхность таламуса прилегает к внутренней капсуле, capsula interna, нижняя соединяется с гипоталамусом, а верхняя и медиальная свободны. Верхняя поверхность таламуса выпуклая, покрыта тонким слоем белого вещества, ограничена снаружи пограничной полоской, stria terminalis, которая отделяет ее от хвостатого ядра, nucleus caudatus. Медиальная поверхность таламуса серого цвета. Обе поверхности соединены между собой межталамическим сращением, adhesio interthalamica. Таламус представляет собой наиболее крупное скопление серого вещества мозгового ствола. На его разрезах видно, что вся масса серого вещества разделена полосками белого вещества, которые называют мозговыми пластинками таламуса, laminae medullares thalami, на множество ядер разной величины.
Таламус является коллектором чувствительных путей, по которым к коре полушарий большого мозга следуют импульсы от эксте- ро- и интероцепторов.
На ранних стадиях развития основными ядрами таламуса являются те, которые уже ограничены мозговыми пластинками. В даль-
нейшем происходят развитие и разделение этих ядерных образований. Они обособляются на дорсальные и вентральные, передние и задние, латеральные и медиальные.
У взрослого человека все ядра таламуса топографически подразделяются на переднюю, внутреннюю и боковую группы. На ядрах таламуса происходит переключение 4 крупных пучков нервных волокон, или чувствительных петель: lemniscus medialis, lemniscus spinalis, lemniscus trigeminalis, lemniscus lateralis.
Пучки нервных волокон, связывающие таламус с корой полушарий большого мозга, образуют лучистый венец, corona radiata. В лучистом венце выделяют зрительную и слуховую лучистости; переднюю, центральную и заднюю таламические лучистости.
Ядра таламуса в функциональном аспекте можно разделить на две основные группы: специфические и неспецифические. В свою очередь в группе специфических ядер можно выделить переключательные и ассоциативные. Каждое переключательное ядро получает нервные импульсы по конкретному чувствительному тракту, т.е. от второго нейрона чувствительного тракта, или вставочного нейрона. Ассоциативные ядра получают информацию от нейронов переключательных ядер.
Основными переключательными ядрами являются переднее дорсальное, переднее вентральное, переднее медиальное, вентролате- ральное, заднее вентральное латеральное, заднее вентральное медиальное, латеральное и медиальное коленчатые тела. Передние ядра таламуса связаны с передачей в лимбическую область коры конечного мозга информации, воспринимаемой на обонятельных и лимбических рецепторах.
Импульсы общей кожной и проприоцептивной чувствительности, приходящие в таламус в составе нервных волокон медиальной, спинномозговой и тройничной чувствительных петель, претерпевают синаптическое переключение на нейронах задних вентральных латерального и медиального ядер.
В вентролатеральные ядра таламуса приходит информация от базальных ядер и мозжечка. Сигналы от этих ядер по прямым свя- зям поступают в моторную зону коры. Вентролатеральные ядра участвуют в регуляции непроизвольной двигательной активности.
Переключательными таламическими ядрами зрительной и слуховой информации являются соответственно латеральное и меди- альное коленчатые тела - метаталамус. Большая часть нейронов переключательных ядер таламуса передает информацию в соответствующие специфические сенсорные зоны коры полушарий боль-
шого мозга, а другая часть - в адекватные ассоциативные ядра таламуса.
Ассоциативные ядра таламуса передают адаптированную информацию, полученную от переключательных ядер, в соответствующие ассоциативные зоны коры конечного мозга, расположенные рядом с сенсорными зонами.
Основными ассоциативными ядрами таламуса являются латеральные, медиодорсальное и подушка. При этом, например, лате- ральная часть подушки получает информацию от латерального коленчатого тела, передает ее в зрительную ассоциативную зону коры конечного мозга, а медиальная часть подушки - из медиального коленчатого тела в слуховую ассоциативную зону коры.
Деятельность неспецифических ядер таламуса в известной степени похожа на функцию ретикулярной формации. Различие состоит в том, что ретикулярная формация мозгового ствола обеспечивает медленную и длительную активацию коры конечного мозга, а неспецифические ядра таламуса создают быструю и кратковременную настройку для разрешения какой-либо конкретной ситуации.
Эпиталамус, epithalamus, состоит из нескольких образований, расположенных над таламусом. К ним относятся мозговые полоски, striae medullares, которые разделяют верхнюю и медиальную поверхности таламуса. Треугольной формы расширение этих полосок образует треугольник поводка, trigonum habenulae. Правый и левый поводки соединены между собой спайкой поводков, commissura habenularum. Каждый поводок переходит в мозговую пластинку, которая соединяется с шишковидной железой, grandula pinealis, - железой внутренней секреции. От основания шишковидного тела к верхним бугоркам крыши среднего мозга проходит мозговая пластинка, в месте перегиба которой находится эпиталамическая спайка, commissura epithalamica.
Гипоталамус, hypothalamus, топографически располагается под полостью промежуточного мозга, III желудочком. К гипоталамусу относят следующие образования: серый бугор, воронка, предзрительное поле, зрительный перекрест, зрительный тракт, нейрогипофиз, сосцевидные тела. Более чем 30 пар ядер гипоталамуса разделяются на 3 группы: переднюю, среднюю и заднюю. Ядра гипоталамуса имеют многочисленные связи со структурами таламуса, лимбической системой и ретикулярной формацией. Обширные нервные и сосудистые связи с гипофизом лежат в основе гипоталамо-гипофизарной системы. В ядрах гипоталамуса располагаются
подкорковые центры, контролирующие деятельность вегетативной нервной системы.
Серый бугор, tuber cinereum, представляет собой серого цвета возвышение, расположенное между зрительным перекрестом спе- реди и сосцевидными телами сзади. Вершина возвышения, суживаясь, образует воронку, infundibulum, которая переходит в нейрогипофиз, neurohypophysis. Области воронки со стороны дна III желудочка соответствует углубление, recessus infundibuli. Серый бугор и воронка образованы тонкой пластинкой серого вещества, которая кпереди постепенно истончается и переходит в конечную пластинку, lamina terminalis.
Зрительный перекрест, chiasma opticum, состоит главным образом из волокон зрительных нервов, при неполном перекресте кото- рых образуется пластинка примерно четырехугольной формы. Задние углы этой пластинки продолжаются в зрительные тракты, tractus optici.
Сосцевидные тела, corpora mamillaria, расположены между серым бугром и задним продырявленным веществом в виде двух белого цвета округлых образований величиной с горошину. Внутри каждого из сосцевидных тел под слоем белого вещества находятся два ядра - латеральное и медиальное. В сосцевидных телах заканчиваются волокна столбов свода, а также волокна из покрышки моста. От медиальных ядер сосцевидных тел возникают пучки волокон к передним ядрам таламуса - сосцевидно-таламический пучок, fasciculus mamillothalamicus, и к ядрам покрышки - сосцевидно-покрышечный пучок, fasciculus mamillotegmentalis.
III ЖЕЛУДОЧЕК
Третий (III) желудочек, ventriculus tertius, является полостью промежуточного мозга и представляет собой узкую вертикальную щель между медиальными поверхностями таламуса. Боковыми стенками III желудочка служат медиальные поверхности таламуса и образования гипоталамуса. Нижнюю стенку, или дно, III желудочка образуют передние участки ножек мозга, заднее продырявленное вещество, сосцевидные тела, серый бугор с воронкой и зрительный перекрест. В области дна III желудочка имеются 2 кармана: углубление воронки, recessus infundibuli, и супраоптическое углубление, recessus supraopticus.
Переднюю стенку III желудочка образуют конечная пластинка, столбы свода и передняя спайка. Между изгибом столбов свода и
таламусом находится овальной формы межжелудочковое отверстие, foramen interventriculare, которое соединяет полость бокового и III желудочков. Задняя стенка желудочка образована спайками поводков, основанием шишковидного тела и эпиталамической спайкой. В области основания шишковидного тела находится шишковидное углубление, recessus pinealis. Верхняя стенка, или крыша, III желудочка расположена под сводом и мозолистым телом. Она имеет вид тонкой пластинки, называемой сосудистой основой III желудочка, tela choroidea ventriculi tertii; прикрепляется к внутреннему краю мозговых полосок таламуса, а сзади - к верхней поверхности поводков и шишковидному телу, образуя здесь надшишковидное углубление, recessus suprapinealis.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
1. Каковы этапы развития головного мозга человека?
2. Назовите основные жизнеобеспечивающие центры продолговатого мозга.
3. Какие собственные ядра мозжечка вам известны?
4. Назовите закономерности проекции ядер черепных нервов на ромбовидную ямку.
5. Назовите ядра среднего мозга.
6. Какие анатомические структуры относят к гипоталамусу?
7. Чем ограничены стенки III желудочка?
КОНЕЧНЫЙ МОЗГ
Конечный мозг, telencephalon, - самый крупный отдел ЦНС (рис. 209). В образованиях конечного, или большого мозга, cerebrum, сосредоточены центры, управляющие деятельностью различных отделов мозгового ствола и спинного мозга. Кора большого мозга осуществляет высшую нервную деятельность, определяет поведение организма в зависимости от условий внешней и состояния внутренней среды. Если в низших отделах нервной системы рефлекторные реакции протекают по безусловным вариантам, то в коре большого мозга нервные связи создаются под воздействием приобретенного индивидуального опыта.
Большой мозг составляют правое и левое полушария, hemispheria cerebralis dextrum et sinistrum, которые соединены между собой мозолистым телом, corpus callosum, и другими спайками или комиссурами (передняя спайка, спайка свода и др.).
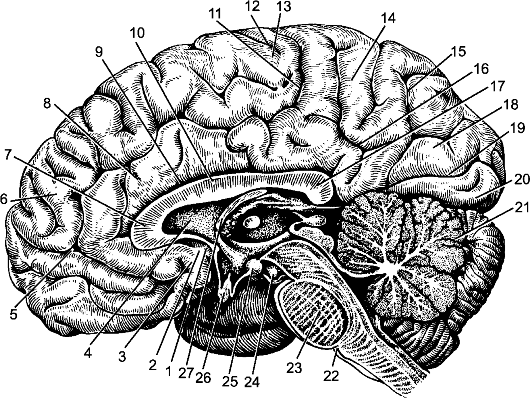
Рис. 209. Медиальная поверхность правого полушария большого мозга, мозгового ствола и мозжечка. Водопровод среднего мозга, IV желудочек. 1 - подмозолистое тело; 2 - задняя обонятельная борозда; 3 - обонятельная пло- щадка; 4 - прозрачная перегородка; 5, 11 - поясная борозда; 6 - верхняя лобная извилина; 7 - колено мозолистого тела; 8 - поясная извилина; 9 - борозда мозо- листого тела; 10 - ствол мозолистого тела; 12 - центральная борозда; 13 - околоцентральная долька; 14 - предклинье; 15 - теменно-затылочная борозда; 16 - подтеменная борозда; 17 - валик мозолистого тела; 18 - клин; 19 - шпорная борозда; 20 - медиальная затылочно-височная извилина; 21 - долька нижнего червя; 22 - задняя граница моста; 23 - мост; 24 - глазодвигательный нерв; 25 - сосцевидное тело; 26 - зрительный перекрест; 27 - свод.
В каждом полушарии различают кору большого мозга, cortex cerebri; белое вещество; обонятельный мозг, rhinencephalon; базальные ядра, nuclei basales.
КОРА БОЛЬШОГО МОЗГА
Полушария большого мозга соответствуют форме черепа. В каждом из полушарий выделяют 3 поверхности: сферическую верхнелатеральную, facies superolateralis, сравнительно плоскую медиальную, facies medialis, и довольно сложной формы нижнюю поверхность, facies inferior. Передний выступающий участок полушария называ-
ют лобным полюсом, polus frontalis, задний - затылочным полюсом,
polus occipitalis, а нижнюю заостренную часть бокового отдела - височным полюсом, polus temporalis.
Поверхность полушарий разделяют различной глубины и длины борозды большого мозга, sulci cerebri, на множество извилин. Извилиной большого мозга, gyrus cerebri, называют участок мозгового вещества в форме валика, который ограничен двумя бороздами. Все борозды полушарий можно объединить в 3 группы: первичные, вторичные и третичные. Первичные борозды глубокие, постоянно встречаются и рано появляются в онтогенезе. Вторичные также довольно постоянны, но появляются позже первичных, а третичные борозды непостоянны и крайне изменчивы. Глубокие первичные борозды делят каждое полушарие на большие участки, называемые долями большого мозга. В каждом полушарии имеется 5 долей: лобная, lobus frontalis, теменная, lobus parietalis, височная, lobus temporalis, затылочная, lobus occipitalis, и островковая, lobus insularis, которая скрыта в глубине латеральной борозды. Поверхность полушарий как в углублениях борозд, так и на вершинах извилин представляет собой значительный по толщине слой серого вещества, который называют корой большого мозга, cortex cerebri. Вследствие многочисленных борозд кора полушарий большого мозга имеет складчатый вид.
БОРОЗДЫ И ИЗВИЛИНЫ ВЕРХНЕЛАТЕРАЛЬНОЙ ПОВЕРХНОСТИ ПОЛУШАРИЯ
Самой глубокой бороздой верхнелатеральной поверхности полушария является латеральная борозда, sulcus lateralis. Она прослеживается от основания мозга в виде глубокой щели, которая разделяется на 3 ветви: две короткие - переднюю, r. anterior, и восходящую, r. ascendens, и длинную заднюю, r. posterior. Задняя ветвь заканчивается примерно на границе средней и задней третей верхнелатеральной поверхности полушария. Несколько кзади от середины полушария с медиальной поверхности сверху вниз и сзади наперед проходит центральная борозда, sulcus centralis, которая, как правило, не доходит до боковой борозды. Наиболее глубоким участком центральной борозды является ее средняя часть. Участок полушария, расположенный кпереди от центральной борозды и кверху от латеральной борозды, относится к лобной доле (рис. 210).
Позади центральной борозды находится теменная доля. Ее ограничивает спереди центральная борозда, снизу - задняя ветвь лате-
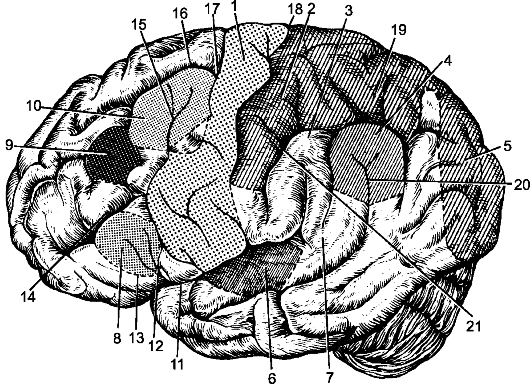
Рис. 210. Верхнелатеральная поверхность левого полушария большого мозга. 1 - ядро двигательного анализатора; 2 - ядро кожного анализатора; 3 - ядро двигательного анализатора, посредством которого синтезируются привычные целенаправленные движения; 4 - ядро зрительного анализатора письменной речи; 5 - ядро зрительного анализатора (зрительная память); 6 - ядро слухового анализатора; 7 - ядро слухового анализатора речи; 8 - ядро двигательного анализатора артикуляции речи; 9 - ядро двигательного анализатора, имеющего отношение к сочетанному повороту головы и глаз; 10 - ядро двигательного анализатора письменной речи; 11 - латеральная борозда мозга; 12 - восходящая ветвь латеральной борозды мозга; 13 - передняя ветвь боковой борозды мозга; 14 - нижняя лобная борозда; 15 - нижняя предцентральная борозда; 16 - верхняя лобная борозда; 17 - верхняя предцентральная борозда; 18 - центральная борозда; 19 - внутритеменная борозда; 20 - верхняя височная борозда; 21 - постцентральная борозда.
ральной борозды, сзади - две борозды: теменно-затылочная борозда, sulcus parietooccipitalis, и поперечная затылочная борозда, sulcus occipitalis transversus. Книзу от латеральной борозды расположена височная доля, которая кзади переходит в затылочную. Границей этих долей считают линию, проведенную от предзатылочной вырезки к нижнему краю полушария. Затылочная доля расположена позади предзатылочной вырезки и поперечной затылочной борозды. Ее границей с височной долей служит линия, соответствующая продолжению книзу поперечной затылочной борозды.
Лобная доля
Параллельно центральной борозде в задней части лобной доли проходит предцентральная борозда, sulcus precentralis. От этой борозды почти под прямым углом в продольном направлении идут верхняя и нижняя лобные борозды, sulci frontales superior et inferior. Указанные борозды делят лобную долю на 4 извилины: предцентральную, gyrus precentralis; верхнюю лобную, gyrus frontalis superior; среднюю лобную, gyrus frontalis medius; нижнюю лобную, gyrus frontalis inferior.
Теменная доля
Позади центральной борозды и параллельно ей проходит постцентральная борозда, sulcus postcentralis. Верхний конец этой борозды находится у края полушария, а нижний часто переходит во внутрите- менную борозду, sulcus intraparietalis, которая проходит параллельно верхнему краю полушария. Центральная и постцентральная борозды ограничивают вертикальную постцентральную извилину, gyrus postcentralis. Остальная часть теменной доли внутритеменной бороздой делится на верхнюю и нижнюю теменные дольки, lobuli parietales superior et inferior. Нижняя теменная долька огибает концы задней ветви латеральной и верхней височной борозд. Часть дольки, огибающей конец задней ветви латеральной борозды, называют надкрае- вой извилиной, gyrus supramarginalis, а участок, огибающий конец верхней височной борозды, - угловой извилиной, gyrus angularis.
Височная доля
На латеральной поверхности височной доли имеются продольные верхняя и нижняя височные борозды, sulci temporales superior et inferior. Между латеральной и верхней височными бороздами расположена верхняя височная извилина, gyrus temporalis superior, на верхней поверхности которой имеются две или три поперечные височные извилины, gyri temporales transversi. Верхняя и нижняя височные борозды ограничивают среднюю височную извилину, gyrus temporalis medius, которая соединяется с расположенной ниже нижней височной извилиной, gyrus temporalis inferior, несколькими переходными извилинами.
Затылочная доля
Борозды латеральной поверхности затылочной доли непостоянны по числу и направлению. Поперечные затылочные борозды, sulci
occipitales transversi, разделяют латеральную поверхность этой доли на одноименные извилины.
Островковая доля
Островковая доля, lobus insularis, расположена в глубине латеральной борозды и образует ее дно. Она представляет собой треугольной формы выступ, вершина которого направлена кнаружи и вниз. Островковая доля окружена круговой бороздой островка, sulcus circularis insulae. Покрывающие островковую долю края латеральной борозды называют покрышкой, ореrculum, которая разделяется на лобную, operculum frontale, теменную, operculum parietale, и височную, operculum temporale.
БОРОЗДЫ И ИЗВИЛИНЫ НИЖНЕЙ ПОВЕРХНОСТИ ПОЛУШАРИЯ
Участок нижней поверхности полушария, facies inferior hemispherii, расположенный кпереди от боковой борозды, относится к лобной доле. Здесь в сагиттальном направлении параллельно продольной щели мозга проходит обонятельная борозда, sulcus olfactorius. Она прикрыта обонятельной луковицей, bulbus olfactorius, переходящей в обонятельный тракт, tractus olfactorius. Между обонятельной бороздой и продольной щелью мозга расположена прямая извилина, gyrus rectus. Кнаружи от обонятельной борозды находится несколько глазничных борозд, sulci orbitales, ограничивающих глазничные извилины, gyri orbitales. Последние занимают остальную часть нижней поверхности лобной доли.
Участок нижней поверхности, который расположен позади латеральной борозды, относится к височной и затылочной долям. Наружной бороздой нижней поверхности височной доли является за- тылочно-височная, sulcus occipitotemporalis, которая вместе с нижней височной бороздой ограничивает нижнюю височную извилину, gyrus temporalis inferior. Медиальнее затылочно-височной борозды и почти параллельно ей проходит глубокая коллатеральная борозда, sulcus collateralis. Между этими бороздами расположена латеральная заты- лочно-височная извилина, gyrus occipitotemporalis lateralis. Кнутри от коллатеральной борозды находится медиальная затылочно-височная извилина, gyrus occipitotemporalis medialis. Эту извилину ограничивают коллатеральная борозда и шпорная борозда, sulcus calcarinus. Латеральная и медиальная затылочно-височные извилины соединены
переходными извилинами с парагиппокампальной извилиной, gyrus parahippocampalis.
БОРОЗДЫ И ИЗВИЛИНЫ
МЕДИАЛЬНОЙ ПОВЕРХНОСТИ ПОЛУШАРИЯ
На медиальной поверхности полушария, facies medialis hemispherii, непосредственно под мозолистым телом имеется борозда мозолистого тела, sulcus corporis callosi, которая огибает все отделы мозолистого тела и переходит в глубокую борозду гиппокампа, sulcus hippocampi. Выше борозды мозолистого тела проходит вторая дугообразная борозда, которую называют поясной бороздой, sulcus cinguli. Эта борозда начинается под клювом мозолистого тела, идет назад и заканчивается на верхнем краю полушария, несколько кзади от верхнего конца центральной борозды. Непосредственным продолжением поясной борозды кзади является подтеменная борозда, sulcus subparietalis. Над поясной бороздой в передней половине полушария расположен медиальный отдел верхней лобной извилины, задней границей которой на этой поверхности считают околоцентральную борозду, sulcus paracentralis. Между околоцентральной бороздой и задним концом поясной борозды расположена парацентральная долька, lobulus paracentralis. Кзади от этой дольки находится четырехугольной формы образование, называемое предклиньем, precuneus, которое ограничено спереди концом поясной борозды, снизу - подтеменной бороздой, sulcus subparietalis, сзади - теменно-затылочной бороздой. Позади предклинья расположен клин, cuneus, кото- рый относится к затылочной доле. Клин ограничен спереди теменно-затылочной, а сзади - шпорной бороздой. Поясная борозда и борозда мозолистого тела ограничивают поясную извилину, gyrus cinguli, которая посредством узкой полоски - перешейка, istmus gyri cinguli, переходит в парагиппокампальную извилину. Передний отдел парагиппокампальной извилины образует направленный кзади изгиб, который называют крючком, uncus. Парагиппокампальную извилину ограничивает с внутренней стороны борозда гиппокампа, sulcus hippocampi, а с наружной - коллатеральная борозда. Суженное место перехода поясной извилины в парагиппокампальную извилину расположено позади валика (мозолистого тела), splenium (corporis callosi). В борозде гиппокампа находится зубчатая извилина, gyrus dentatus, которая представляет собой серого цвета зазубренную полоску.
СТРОЕНИЕ КОРЫ ПОЛУШАРИЙ БОЛЬШОГО МОЗГА
Структура коры полушарий большого мозга, или коры конечного мозга, представлена скоплением нейронов и клеток про- межуточного вещества - нейроглии. Толщина коры, покрывающей всю поверхность полушарий большого мозга, варьирует от 1,5 до 4,5 мм. Общая площадь поверхности двух полушарий составляет от 1450 до 2200 см2. Борозды обеспечивают относительно максимальную площадь коры конечного мозга при относительно ми- нимальном объеме полушарий, при этом более 60% площади поверхности коры конечного мозга скрыто между извилинами. По приблизительным данным, в коре от 12 до 17 млрд нервных клеток, каждая из которых в среднем имеет тысячи, а иногда и десятки тысяч прямых и опосредованных контактов.
Нервные клетки, составляющие основу коры конечного мозга, располагаются слоями, или пластинками, друг над другом. Различают 6 таких слоев. Слой I - молекулярная пластинка, она содержит немного нейронов и множество переплетений нервных волокон. Слой II - наружная зернистая пластинка. В ней многочисленные мелкие нейроны, напоминающие по форме зерна. Слой III - наружная пирамидная пластинка, содержащая средние и мелкие пирамидные нейроны. Слой IV - внутренняя зернистая пластинка, по клеточному составу она напоминает II слой. Слой V - внутренняя пирамидная пластинка, содержит гигантские пирамидные нейроны. Слой VI - мультиформная пластинка, состоит из веретенообразных и треугольных нервных клеток.
В функциональном отношении все нервные клетки коры полушарий большого мозга можно разделить на 3 основных типа. К I типу - сенсорным корковым нейронам - относятся нервные клетки, на которых переключаются аксоны третьих нейронов специфических чувствительных трактов. Ко II типу относятся эфферентные нейроны, которые передают нервные импульсы из коры к нижележащим структурам: в базальные ядра, ядра мозгового ствола, спинной мозг. Такими двигательными, моторными, или эфферентными, нейронами являются гигантские пирамидные клетки V слоя коры конечного мозга. Наконец, к III типу относятся нервные клетки, обеспечивающие взаимные контакты между нейронами в пределах одной или нескольких структурно-функциональных зон коры. Эти нейроны называются контактными, промежуточными, или вставочными, и представлены мелкими, средними пирамидными и веретенообразными нервными клетками.
Наряду с шестислойным расположением тел нервных клеток в коре конечного мозга направленность проводящих путей или отростков нейронов упорядочена определенным образом.
Шесть слоев горизонтально направленных нервных волокон пересекаются вертикально идущими пучками. Такая конфигурация проводящих путей обеспечивает как внутрикорковые связи, так и взаимосвязь коры с другими структурами нервной системы.
Учение о цитоархитектонике коры полушарий головного мозга соответствует учению И.П. Павлова о коре как о системе корковых концов анализаторов. Анализатор, по И.П. Павлову, «есть сложный нервный механизм, начинающийся наружным воспринимающим аппаратом и кончающийся в мозгу». Корковый конец анализатора, по И.П. Павлову, состоит из «ядра» и «рассеянных элементов». В коре большого мозга описано множество цитоархитектонических полей, в которых расположены ядра основных анализаторов (рис. 211).
1. Предцентральная извилина и передний отдел околоцентральной дольки входят в состав прецентральной области - дви- гательной, или моторной, зоны коры. Афферентные клетки этих полей воспринимают проприоцептивные импульсы от мышц, сухожилий, суставов, а двигательные клетки V и VI пластинок связывают кору полей 4 и 6 с двигательными ядрами черепных нервов и ядрами передних рогов спинного мозга. Группы нейронов осуществляют иннервацию определенных мышц или групп мышц; в верхних отделах находятся центры для мышц верхней конечности, в нижнем - для мышц головы. Наибольшую площадь всей зоны занимают центры иннервации кисти руки, лица, губ, языка, а меньшую площадь - центры иннервации мышц туловища и нижних конечностей. Центры двигательной зоны осуществляют иннервацию противоположной части тела.
2. В задней части средней лобной извилины (поле 8) и на границе теменной и затылочной долей (поле 19) находятся центральные отделы двигательного анализатора, которые осуществляют координацию поворота головы и глаз в противоположную сторону. Участки коры (поля 6 и 8), расположенные кпереди от моторной зоны, называют премоторной зоной. Отростки клеток этой зоны связаны как с ядрами передних рогов спинного мозга, так и с подкорковыми ядрами, красным ядром, черной субстанцией.
3. В постцентральной извилине (поля 1, 2, 3, 5) находится ядро кожного анализатора (центры температурной, болевой, осязательной чувствительности). Последовательность расположения центров и их территория соответствуют моторной зоне коры.
Рис. 211. Карта цитоархитектонических полей мозга человека. Объяснение в тексте. а - наружная поверхность; б - внутренняя поверхность.
4. Корковые концы интероцептивного анализатора лежат в премоторной зоне (поля 6 и 8) и лимбической области коры. Эфферентные пути из этих участков коры идут к ядрам гипоталамуса и другим центрам вегетативной системы.
5. В области верхней височной извилины находится ядро слухового анализатора (поле 41). Здесь проецируются волокна, которые
проводят импульсы от рецепторного аппарата, расположенного в улитке внутреннего уха.
6. Корковый конец зрительного анализатора расположен в затылочной доле мозга (поля 17, 18, 19). Ядро зрительного анализатора находится в области шпорной борозды (поле 17). Здесь спроецирована сетчатка латеральной половины одного глаза и медиальной поло- вины другого, поэтому полная слепота возникает при двустороннем поражении ядер зрительного анализатора. В случае поражения поля 18 утрачивается зрительная память. При поражении поля 19 человек утрачивает способность к ориентировке в новой для него обстановке.
7. В области крючка и гиппокампа имеется ядро обонятельного анализатора.
8. В нижнем отделе постцентральной извилины находится ядро вкусового анализатора.
9. Поля лимбической области коры благодаря многочисленным связям с другими ее областями играют важную роль в синтезе афферентных раздражений.
10. В левой нижней теменной дольке, в области надкраевой извилины (поле 40), у правшей находится центр целенаправленных комбинированных движений.
11. В правой и левой верхних теменных дольках (поле 7) расположены центры особого сложного вида кожной чувствительности - стереогностического чувства, т.е. способности узнавать предметы на ощупь.
12. В задней части нижней лобной извилины (поле 44), у правшей - в левом полушарии, находится двигательный анализатор артикуляции речи (речедвигательный анализатор).
13. С речедвигательным анализатором тесно связан слуховой анализатор устной речи, который расположен в задней части верхней височной извилины (поле 42).
14. В заднем отделе средней лобной извилины (поле 8), рядом с ядром двигательного анализатора поворота глаз и головы в противоположную сторону, имеется ядро двигательного анализатора письменной речи - графии.
15. В нижней теменной дольке, в области угловой извилины, расположен зрительный анализатор письменной речи (поле 39). При повреждении этого поля больной утрачивает способность воспринимать написанные буквы, т.е. читать (алексия).
ОБОНЯТЕЛЬНЫЙ МОЗГ
К обонятельному мозгу, rhinencephalon, относятся обонятельная луковица, bulbus olfactorius, обонятельный тракт, tractus olfactorius,
обонятельный треугольник, trigonum olfactorium, переднее продырявленное вещество, substantia perforata anterior, поясная извилина, gyrus cinguli, перешеек поясной извилины, isthmus gyri cinguli, парагиппо- кампальная извилина, gyrus parahippocampalis, зубчатая извилина, gyrus dentatus, и гиппокамп, hippocampus.
Поясная извилина, перешеек, парагиппокампальная извилина, гиппокамп, зубчатая извилина, свод и миндалевидное тело входят в состав лимбической системы. Считают, что эта система выполняет не только обонятельную функцию, но и имеет важное значение в процессах синтеза афферентных раздражений, эмоциональных реакций и реакций так называемого врожденного поведения (половые, поисковые и оборонительные).
БАЗАЛЬНЫЕ ЯДРА
Базальные ядра расположены в белом веществе полушарий большого мозга, ближе к их основанию. Они образуют 3 парных скоп- ления серого вещества: полосатое тело, ограду и миндалевидное тело (рис. 212).
Полосатое тело, corpus striatum, состоит из двух ядер: хвостатого, nucleus caudatus, и чечевицеобразного, nucleus lentiformis.
Хвостатое ядро представляет собой округлой формы образование, которое в виде дуги окружает таламус и чечевицеобразное ядро. Резко утолщенный, булавовидный передний отдел хвостатого ядра называют головкой, caput nuclei caudati. Кзади головка истончается и образует тело хвостатого ядра, corpus nuclei caudati, которое переходит в хвост, cauda nuclei caudati. Головка, тело и хвост участвуют в образовании стенок бокового желудочка мозга.
Чечевицеобразное ядро представляет собой крупное, пирамидальной формы скопление серого вещества, которое расположено кнаружи от хвостатого ядра и таламуса и отделено от них внутренней капсулой. Выпуклая наружная поверхность чечевицеобразного ядра обращена к островковой доле, а суженная вершина - внутрь и вниз. Ядро неоднородно по строению и разделяется пластинками белого вещества на 3 части: темного цвета наружную - скорлупу, putamen, и две светлые - медиальный бледный шар, globus pallidus medialis, и латеральный бледный шар, globus pallidus lateralis.
Хвостатое и чечевицеобразное ядра представляют собой главное звено экстрапирамидной системы, основной функцией которой является осуществление сложных безусловно-рефлекторных двигательных актов.
Рис. 212. Фронтальный разрез полушарий, проходящий через полосатое тело и таламус.
1 - свод; 2 - сосудистое сплетение III желудочка; 3 - сосудистое сплетение бокового желудочка; 4 - боковой желудочек; 5 - хвостатое ядро; 6 - задняя ножка внутренней капсулы; 7 - скорлупа; 8 - латеральный и медиальный бледные шары; 9 - наружная капсула; 10 - ограда; 11 - боковая борозда; 12, 14, 15 - соответственно верхняя, средняя и нижняя височные извилины; 13 - островковая доля; 16 - парагиппокампальная извилина; 17 - глазодвигательный нерв; 18 - мост; 19 - ядро сосцевидного тела; 20 - зрительный тракт; 21 - III желудочек; 22 - межталамическое сращение; 23-25 - ядра таламуса; 26, 28, 31 - соответственно нижняя, средняя и верхняя лобные извилины; 27 - мозолистое тело; 29 - извилина пояса; 30 - борозда пояса.
Ограда, claustrum, - сравнительно тонкая пластинка серого вещества. Расположена в белом веществе, разделяющем наружную поверхность, скорлупу и кору островка. Белое вещество, которое разделяет ограду и скорлупу, называют наружной капсулой, capsula externa, а участок белого вещества между оградой и корой островка - самой наружной капсулой, capsula extrema. Ограда является сложным образованием, связи которого до настоящего времени изучены мало, а функциональное значение неясно.
Миндалевидное тело, corpus amygdaloideum, - крупное ядро, которое расположено под скорлупой в переднем отделе височной доли. Это ядро имеет сложное строение. Оно состоит из нескольких ядер, различающихся по клеточному составу. Миндалевидное тело, яв-
ляющееся подкорковым обонятельным центром, входит в состав лимбической системы.
БОКОВЫЕ ЖЕЛУДОЧКИ
Полостями конечного мозга являются боковые желудочки, ventriculi laterales. Они расположены в толще полушарий. В каждом боковом желудочке различают центральную часть, pars centralis, и 3 рога: передний, соrnи frontale, задний, соти occipitale, и нижний, соти temporale.
Центральная часть в виде узкой горизонтальной щели расположена в теменной доле (рис. 213).
На внутренней поверхности нижнего рога имеется возвышение - гиппокамп, hippocampus, который образуется вследствие вдавления в желудочек борозды гиппокампа. Гиппокамп представляет собой сместившуюся в полость желудочка часть старой коры. Межжелудочковые отверстия, foramina interventriculares, соединяют полость боковых желудочков с III желудочком мозга.
БЕЛОЕ ВЕЩЕСТВО ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
Белое вещество полушарий образовано волокнами проводящих путей конечного мозга, которые группируются в 3 системы: 1) ас- социативные, или сочетательные, 2) комиссуральные, или спаечные, 3) проекционные.
Ассоциативные волокна соединяют между собой различные участки коры в пределах одного полушария. Эти волокна делятся на короткие и длинные. К коротким относятся дугообразные волокна, которые соединяют кору двух соседних извилин. Они образуют самый поверхностный слой белого вещества, расположенный непосредственно под корой. Длинные волокна находятся глубже коротких, но по отношению ко всей массе белого вещества полушария являются поверхностными. Пучки длинных ассоциативных волокон соединяют отделенные друг от друга участки коры. Известны следующие пучки длинных ассоциативных волокон (рис. 214).
1. Пояс, cingulum, прослеживается от переднего продырявленного вещества до парагиппокампальной извилины. Он соединяет кору извилин медиальной поверхности полушария и относится к обонятельному мозгу.
2. Нижний продольный пучок, fasciculus longitudinalis inferior, соединяет затылочную долю с височной. Крупные, различной длины
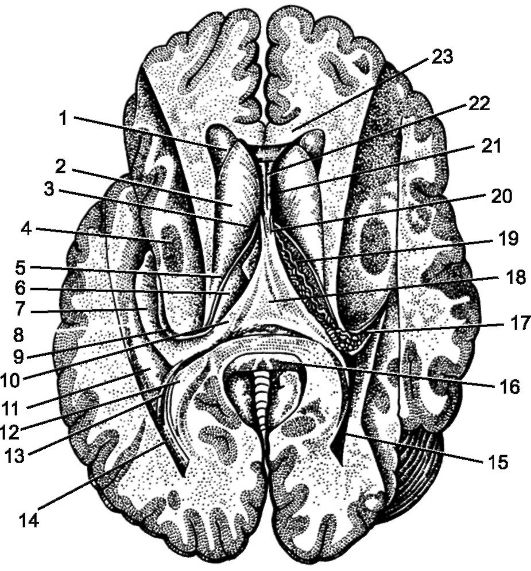
Рис. 213. Боковые желудочки, вскрытые сверху; удалена часть полушарий вместе с мозолистым телом.
1 - передний рог; 2 - хвостатое ядро (головка); 3 - межжелудочковое отверстие; 4 - чечевицеобразное ядро (в разрезе); 5 - конечная полоска; 6 - верхняя поверхность таламуса; 7 - гиппокамп; 8 - боковое возвышение; 9 - бахромка гиппокампа; 10 - ножка свода; 11 - задний рог бокового желудочка; 12 - медиальная стенка заднего рога; 13 - птичья шпора; 14, 15 - задние рога; 16 - валик мозолистого тела; 17, 19 - сосудистое сплетение в центральной части бокового желудочка и его продолжение в нижний рог; 18 - спайка свода; 20 - столб свода; 21 - прозрачная перегородка; 22 - полость прозрачной перегородки; 23 - мозолистое тело.
волокна, образующие пучок, расположены вдоль наружной стенки заднего и нижнего рогов бокового желудочка.
3. Верхний продольный пучок, fasciculus longitudinalis superior, соединяет лобную долю с теменной и височной.
4. Крючковидный пучок, fasciculus uncinatus, соединяет прямую и глазничные извилины лобной доли с височной долей.
Комиссуральные пути соединяют области коры обоих полушарий. Они образуют следующие комиссуры, или спайки.
Мозолистое тело, corpus callosum, образовано волокнами, соединяющими между собой различные участки новой коры, в связи с чем его величина у человека значительно больше, чем у животных.
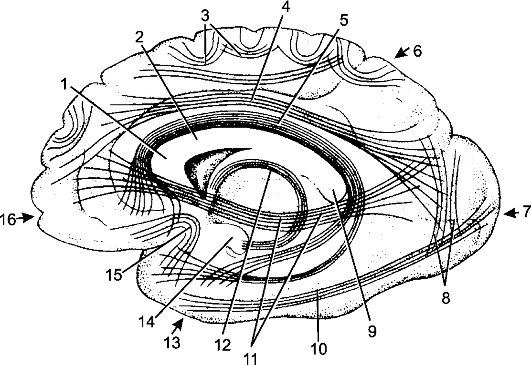
Рис. 214. Ассоциативные пути полушарий большого мозга.
1 - колено мозолистого тела; 2 - ствол мозолистого тела; 3 - дугообразные волокна большого мозга; 4 - верхний продольный пучок; 5 - пояс; 6 - теменная доля; 7 - затылочная доля; 8 - вертикальный затылочный пучок; 9 - валик мозолистого тела; 10 - нижний продольный пучок; 11 - нижний лобно-затылочный пучок; 12 - свод; 13 - височная доля; 14 - крючок; 15 - крючковидный пучок; 16 - лобная доля.
Сравнительно тонкая средняя часть мозолистого тела называется стволом, truncus corporis callosi. Кпереди ствол утолщается и изгибается книзу, образуя колено мозолистого тела, genu corporis callosi, нижний конец которого, истончаясь, переходит в клюв, rostrum corporis callosi. Задний утолщенный конец мозолистого тела - валик, splenium, несколько изогнут книзу. Вся верхняя поверхность мозолистого тела по- крыта тонким слоем серого вещества - серым покровом, indusium griseum.
Передняя спайка, commissura anterior, и спайка свода, commissura fornicis, являются самыми крупными спайками мозга после мозолистого тела. Передняя спайка лежит кзади от клюва мозолистого тела, а спайка свода соединяет столбы свода, columnae fornicis. Свод, fornix, расположен под мозолистым телом. В нем различают ножки свода, crurae fornicis, и столбы, которые, изгибаясь кзади, вступают в гипоталамус и заканчиваются в сосцевидных телах (рис. 215).
Проекционные пути соединяют кору полушарий большого мозга с ядрами мозгового ствола и спинного мозга. Различают эфферентные - двигательные пути и афферентные - чувствительные пути. Двигательные пути проводят нервные импульсы от клеток двига-
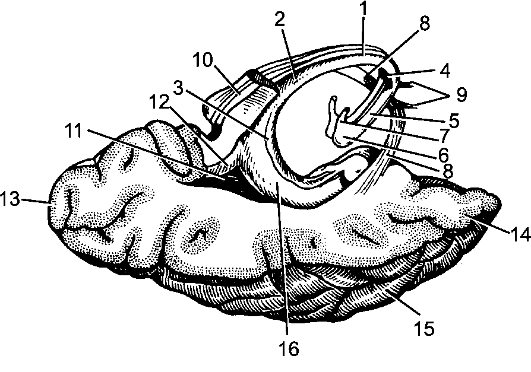
Рис. 215. Свод, гиппокамп и передняя комиссура.
1 - тело свода; 2 - ножка свода; 3 - бахромка гиппокампа; 4, 5 - столб свода; 6 - сосцевидное тело; 7 - сосцевидно-таламический пучок; 8, 9 - передняя спайка; 10 - валик мозолистого тела; 11 - боковой треугольник; 12 - птичья шпора; 13 - затылочный полюс; 14 - височный полюс; 15 - средняя височная извилина; 16 - гиппокамп.
тельных областей коры к двигательным ядрам спинного мозга, ядрам мозгового ствола и подкорковым ядрам. Благодаря этим путям двигательные центры коры головного мозга проецируются на периферию. Посредством чувствительных путей осуществляется проекция тела в чувствительные и двигательные центры коры.
Чувствительные волокна являются отростками клеток спинномозговых узлов и чувствительных узлов черепных нервов. Они образуют первые нейроны чувствительных путей. Отростки клеток спинномоз- говых узлов и узлов черепных нервов заканчиваются на телах или дендритах нейронов переключательных ядер спинного либо продолговатого мозга. От тел клеток переключательных ядер продолжаются чувствительные пути, которые в составе медиальной петли идут к вентральным ядрам таламуса. В этих ядрах заканчиваются вторые вставочные и начинаются третьи нейроны, которые идут в соответствующие ядерные центры коры. Как чувствительные, так и дви- гательные пути проходят между таламусом и подкорковыми ядрами и образуют в веществе полушарий систему лучеобразно расходящихся пучков, которая получила название лучистого венца, corona radiata.
Волокна, которые образованы проводящими путями каждого полушария, собираются в компактный мощный пучок, заполняющий пространство между таламусом, с одной стороны, хвостатым и чечевицеобразным ядрами - с другой. Этот пучок волокон называют внут-
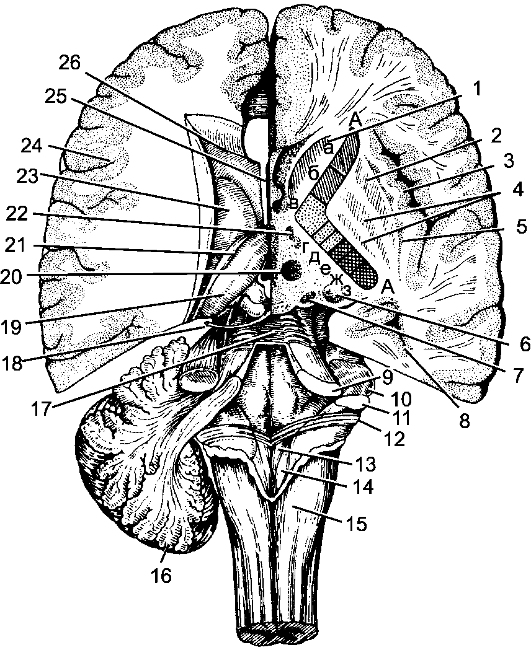
Рис. 216. Полушария большого мозга на разных уровнях горизонтального разреза: справа - ниже уровня дна бокового желудочка, слева - над дном бокового желудочка; IV желудочек мозга вскрыт сверху.
1 - головка хвостатого ядра; 2 - скорлупа; 3 - кора островковой доли полушария; 4 - латеральный и медиальный бледные шары; 5 - ограда; 6 - хвост хвостатого ядра; 7 - ядро медиального коленчатого тела; 8 - нижний рог бокового желудочка; 9 - верхняя мозжечковая ножка; 10 - средняя мозжечковая ножка; 11 - нижняя мозжечковая ножка; 12 - мозговые полоски; 13 - треугольник подъязычного нерва; 14 - треугольник блуждающего нерва; 15 - бугорок тонкого ядра; 16 - мозжечок; 17 - верхний мозговой парус; 18 - блоковый нерв; 19 - таламус; 20 - красное ядро; 21 - конечная полоска; 22 - ядра гипоталамуса; 23 - хвостатое ядро; 24 - кора полушарий; 25 - полость прозрачной перегородки; 26 - передний рог бокового желудочка. АА - внутренняя капсула: а - лобно-таламический путь; б - лобно-мостовой путь; в - корково-ядерный путь; г - корково-спинномозговой путь; д - таламокорковый путь; е - затылочно-височно-мостовой путь; ж - центральный слуховой путь; з - центральный зрительный путь.
ренней капсулой, capsula interna. На срезах полушарий мозга внутренняя капсула имеет вид довольно широкой полосы белого вещества, в которой различают переднюю ножку, заднюю ножку и колено (рис. 216).
Передняя ножка, crus anterior capsula internae, находится между хвостатым и чечевицеобразным ядрами. Она образована волокнами,
идущими от коры лобной доли к таламусу, - tractus frontothalamicus, красному ядру, - tractus frontorubralis, ядрам моста, - tractus frontopontinus. Кроме этих волокон, которые имеют продольное направление, здесь проходят поперечные волокна хвостатого ядра к чечевицеобразному ядру.
Задняя ножка, crus posterior capsulae internae, длиннее передней. Она расположена между таламусом и чечевицеобразным ядром. Передняя часть задней ножки образует волокна от клеток коры предцентральной извилины к двигательным ядрам мозга и составляет корково-спинномозговой путь, tractus corticospinalis. Задняя треть этой ножки образована чувствительными волокнами - отростками клеток ядер таламуса к коре постцентральной извилины - таламокорковый путь, tractus thalamocortical. Здесь же проходят затылочно-височно-мостовой путь, tractus occipitotemporopontinus, а также центральный слуховой и центральный зрительный пути.
Колено, genu capsulae internae, образовано нервными волокнами от нижних отделов коры предцентральной извилины, которые идут к ядрам черепных нервов среднего мозга, моста и продолговатого мозга - корково-ядерный путь, tractus corticonuclearis.
ПРОВОДЯЩИЕ ПУТИ ГОЛОВНОГО И СПИННОГО МОЗГА
Проводящие пути головного и спинного мозга делят на ассоциативные, комиссуральные и проекционные. Ассоциационные и комиссуральные пути были рассмотрены при описании различных отделов ЦНС. Проекционные пути подразделяются на чувствительные - афферентные и двигательные - эфферентные. Наиболее важные из них следующие.
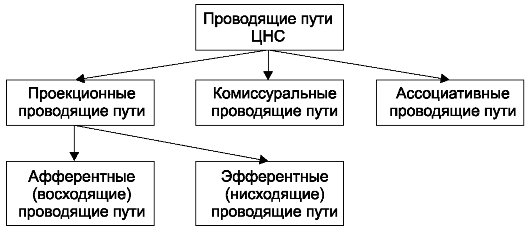
Рис. 217. Классификация проводящих путей ЦНС.
АФФЕРЕНТНЫЕ ПРОВОДЯЩИЕ ПУТИ
1. Проводящие пути проприоцептивной чувствительности (глубокой) передают в область коркового ядра двигательного анализатора импульсы глубокой чувствительности от проприоцепторов мышц сухожилий, суставов, которые являются сигналами изменений напряжений мышц, суставных сумок и других элементов двигательного аппарата, т.е. сигналы кинестетических раздражений. Первые нейроны - это псевдоуниполярные клетки спинномозговых узлов. Дендриты псевдоуниполярных клеток спинномозговых узлов заканчиваются проприоцепторами в мышцах, сухожилиях, суставах, а нейриты в составе задних корешков вступают в корешковую зону задних рогов спинного мозга и, не прерываясь в его ядрах, переходят в задние канатики.
Вторыми нейронами являются клетки переключательных ядер тонкого и клиновидного канатиков, волокна которых изгибаются, обходят спереди центральный канал и в межоливарном слое образуют перекрест с волокнами противоположной стороны. Эти волокна называют внутренними дугообразными, а их перекрест - верхним чувствительным перекрестом, или перекрестом петли, так как они составляют медиальную петлю. Медиальная петля в области моста расположена позади двигательного пути. Уже в верхних от- делах продолговатого мозга к ней начинают присоединяться пути тактильной, болевой и температурной чувствительности - спиноталамические волокна.
Из моста медиальная петля проходит кнаружи от красных ядер среднего мозга и затем вступает в таламус, где заканчивается в од- ном из ядер вентролатеральной группы. Здесь располагается третий нейрон. Отростки клеток этого ядра образуют таламокорковые пучки. Эти пучки заканчиваются в ядрах кожного и двигательного анализаторов коры конечного мозга.
Считают, что проприоцептивные пути от двигательного аппарата головы проходят в составе проводящих путей тройничного, лицевого, языкоглоточного, блуждающего, подъязычного и добавочного нервов.
2. Проприоцептивные пути к мозжечку участвуют в безусловнорефлекторной регуляции мышечного тонуса, равновесия, координации и синергии движений. Различают передний и задний спиномозжечковые пути, первыми нейронами которых являются псевдоуниполярные клетки спинномозговых узлов. Дендриты этих клеток заканчиваются проприоцепторами в мышцах, сухожилиях,
суставах и постоянно воспринимают раздражения, сигнализирующие о состоянии двигательного аппарата. Аксоны этих клеток в составе задних корешков вступают в спинной мозг и заканчиваются в ядрах заднего рога. Часть нейритов этих ядер проходит в боковой канатик спинного мозга той же стороны и образует задний спиномозжечковый путь, который в составе нижних мозжечковых ножек заканчивается в коре червя, где перекрещивается. Другая часть волокон ядра как своей, так и противоположной стороны проходит в боковом канатике спинного мозга и образует перекрест - передний спиномозжечковый путь. Волокна этого пути проходят продолговатый мозг, мост и возле крыши среднего мозга снова образуют перекрест с волокнами противоположной стороны, после чего передний спиномозжечковый путь в составе верхних мозжечковых ножек проходит в червь мозжечка и заканчивается на клетках его коры.
3. Проводящие пути болевой и температурной чувствительности от кожи шеи, туловища и конечностей организованы в соответствии с принципом сегментарности.
Эти пути составляют трехнейронную цепь. Тела первых - рецепторных нейронов расположены в спинномозговых узлах и пред- ставлены псевдоуниполярными клетками. Периферические отростки первых нейронов контактируют с рецепторными комплексами.
Центральные отростки в составе задних корешков входят в спинной мозг и синаптически оканчиваются на клетках собственного ядра, располагающихся в пределах заднего рога и являющихся вто- рыми нейронами тракта.
Далее аксоны вторых нейронов в восходящем направлении переходят на противоположную сторону спинного мозга, формируя латеральный спиноталамический путь, увеличивающийся в объеме по направлению к начальным сегментам.
В продолговатом мозге латеральный спиноталамический путь лежит кзади от ядра оливы. Здесь латеральный спиноталамический путь, являясь средним связующим звеном пути проведения боле- вой и температурной чувствительности, становится основой спинномозговой петли, которая, присоединяясь к нервным волокнам медиальной петли, достигает тела третьего нейрона, которое расположено в таламусе промежуточного мозга.
В нейронах таламуса осуществляется сложная многопрофильная обработка поступившей от вторых нейронов информации и ее адап- тация для следующего уровня. Аксоны третьих нейронов проходят через внутреннюю капсулу и в составе нервных волокон лучистого
венца достигают зоны центрального отдела кожного анализатора в постцентральной извилине коры конечного мозга.
4. Проводящие пути болевой и температурной чувствительности от
головы располагаются на основе тройничного нерва.
В составе пути имеются 3 уровня, обозначенные тремя нейронами. Тела рецепторных - первых нейронов локализованы в трой- ничном узле. Дендриты или периферические отростки первых нейронов имеются в составе каждой из трех главных ветвей тройничного нерва.
Центральные отростки первых нейронов в составе тройничного нерва входят в мост, а потом образуют нисходящий пучок нервных волокон, достигающих вторых нейронов, образующих ядро спинномозгового пути тройничного нерва.
Аксоны вторых нейронов, переходя на контралатеральную половину, формируют основу тройничной петли, располагающейся рядом с медиальной петлей и достигающей соответствующего ядра таламуса, образованного телами третьих нейронов.
После анализа и интегративной обработки информации в пределах уровня третьего нейрона нервные импульсы распространя- ются по аксонам третьих нейронов во внутреннюю капсулу и лучистый венец к центральному отделу кожного анализатора в область постцентральной извилины.
5. Проводящие пути тактильной чувствительности (сознательной) являются трехнейронными. Рецепторные - первые нейроны располагаются в спиномозговых узлах и представляют собой псевдоуниполярные нервные клетки. Каждый нейрон имеет периферический отросток - дендрит, контактирующий с рецепторами кожи, и центральный отросток - аксон, вступающий в составе задних корешков в спинной мозг.
Преобладающая часть аксонов первого нейрона оканчивается на нейронах студенистого вещества, расположенных в заднем роге спинного мозга и являющихся вторыми нейронами пути. Сравнительно небольшая часть вторых нейронов располагается в составе ядер тонкого и клиновидного пучков в продолговатом мозге. Чтобы достичь их, аксоны первых нейронов, минуя серое вещество спинного мозга, поднимаются вверх в составе задних канатиков.
Аксоны вторых нейронов обеих частей пути достигают следующего уровня - таламуса, проходя в составе афферентных пучков, сопровождающих медиальную петлю.
Следует подчеркнуть, что большая часть аксонов вторых нейронов, локализованная в задних рогах спинного мозга, до вступления
в продолговатый мозг переходит на восходящем сегментарном уровне на контралатеральную сторону и формирует передний спиноталамический тракт в составе боковых канатиков.
Тела третьих нейронов пути проведения тактильной чувствительности располагаются в таламусе, их аксоны достигают центрального отдела анализатора в области постцентральной извилины, проходя в составе внутренней капсулы через лучистый венец к коре конечного мозга.
6. Проводящие пути стереогноза или пространственной кожной чувствительности участвуют в обеспечении узнавания предметов ощупыванием и являются трехнейронными.
Тела рецепторных, или первых, нейронов традиционно участвуют в образовании спинномозговых узлов и представлены псевдоуниполярными клетками. Их периферические отростки в составе производных спинномозговых нервов контактируют с рецепторными зонами.
Аксоны первых нейронов входят в спинной мозг как часть задних корешков и, не переключаясь на нейронах задних рогов, всту- пают в состав тонкого и клиновидного пучков задних канатиков. В составе ядер тонкого и клиновидного пучков в продолговатом мозге располагаются тела вторых нейронов проводящего пути стереогноза.
В свою очередь аксоны вторых нейронов присоединяются к нервным волокнам медиальной петли и в ее составе переходят на противоположную сторону.
Следующий уровень проводящих путей пространственной кожной чувствительности, или третий нейрон, располагается в таламусе промежуточного мозга. Отсюда аксоны третьих нейронов в составе внутренней капсулы, а затем лучистого венца достигают центрального отдела анализатора в коре конечного мозга.
7. Обонятельные проводящие пути подразделяют на периферический и центральный отделы. К периферическому отделу относят системы центральных отростков обонятельных клеток, которые через отверстия решетчатой пластинки решетчатой кости проникают из полости носа в полость черепа и заканчиваются на митральных клетках обонятельной луковицы. Обонятельные луковицы и ряд подкорковых и корковых образований составляют центральный отдел обоня- тельного проводящего пути. Нейриты митральных клеток, соединившись, образуют обонятельный тракт. Часть нейритов митральных клеток заканчивается на зернистых клетках обонятельной луковицы, нейриты которых образуют синапсы с митральными клетками.
Считают, что такие связи между клетками в пределах обонятельной луковицы способствуют усилению обонятельных импульсов. В области обонятельного треугольника некоторые волокна обонятельного тракта заканчиваются. Большая часть волокон разделяется на 3 пучка: наружный пучок заканчивается в коре крючка парагиппокампальной извилины, средний - в переднем продырявленном веществе, а медиальный проходит в околообонятельное поле коры лобной доли мозга.
Обонятельные центры коры соединены проекционными волокнами с подкорковыми обонятельными центрами, из которых сосцевидные тела в свою очередь соединены со многими ядрами мозгового ствола (сосцевидно-таламический и сосцевидно-покрышечный пучки).
8. Проводящие пути от органов растительной жизни - внутренних органов, кровеносных и лимфатических сосудов и др. - образуют цепи из 3 нейронов. Первыми нейронами этих путей являются клетки узлов некоторых черепных нервов (тройничного, лицевого, языкоглоточного и блуждающего) и спинномозговых узлов, вторые нейроны - это клетки ядер указанных нервов, а третьи - клетки таламуса, отростки которых заканчиваются в нижнем отделе постцентральной извилины.
ЭФФЕРЕНТНЫЕ ПРОВОДЯЩИЕ ПУТИ
1. Корково-спинномозговой (пирамидный) и корково-ядерный пути
осуществляют сознательную произвольную регуляцию движений. Эти пути составляет цепочка из двух нейронов - центрального и периферического. Центральными нейронами называют клетки кор- кового ядра двигательного анализатора, которые заканчиваются в двигательных ядрах черепных нервов (librae corticonuclearis) и в двигательных ядрах передних рогов спинного мозга (tractus corticospinalis). Периферическими нейронами являются клетки двигательных ядер черепных нервов и двигательных ядер спинного мозга, отростки которых в составе нервных стволов заканчиваются в соответствующих мышцах двигательными окончаниями. Пирамидный путь проводит импульсы от коркового ядра к активной части двигательного аппа- рата - мышцам. Эти импульсы возникают в коре как результат анализа и синтеза раздражений, поступивших в нее по проприоцептивному пути, а также по другим путям (зрительным, вестибулярным, слуховым, кожным). Основную массу волокон составляют аксоны пирамидных клеток предцентральной извилины.
В средних отделах моста от двигательного пути отделяется пучок волокон к двигательному ядру тройничного нерва своей и противоположной сторон. От клеток этих ядер возникают двигательные волокна тройничного нерва, которые в составе нижнечелюстного нерва идут к соответствующим мышцам и заканчиваются в них двигательными окончаниями. В нижнем отделе моста от двигательного пути отделяются волокна к двигательным ядрам лицевого нерва своей и противоположной сторон, после чего пирамидный путь переходит в продолговатый мозг, где снова принимает вид компактного пучка. В верхнем отделе продолговатого мозга от пирамидного пути отходят волокна к двигательным ядрам блуждающего, языкоглоточного и подъязычного нервов. После отхождения этих пучков пирамидный путь состоит только из волокон, соединяющих кору с ядрами передних рогов спинного мозга. Большая часть этих волокон совершает перекрест в нижнем отделе продолговатого мозга (decussatio pyramidum) и переходит в боковой канатик спинного мозга, где образует латеральный (пирамидный) корково-спинальный путь, tractus corticospinalis lateralis. Волокна этого пути заканчиваются в двигательных ядрах передних рогов спинного мозга. Меньшая часть волокон в области продолговатого мозга перекреста не образует. Эти волокна в спинном мозге составляет передний корково-спинальный путь, tractus corticospinalis ventralis, часть волокон которого заканчивается в двигательных ядрах спинного мозга своей стороны, а другая часть волокон через переднюю белую спайку совершает пе- рекрест и заканчивается в таких же ядрах противоположной стороны. Передний пучок прослеживается только в шейных и верхних грудных сегментах спинного мозга.
Таким образом, пирамидный путь осуществляет преимущественно перекрестную иннервацию мускулатуры тела, и при одностороннем поражении центральных нейронов любого отдела наступает паралич мышц на противоположной очагу поражения стороне. Такая односторонняя иннервация характерна не для всех мышечных групп. Большая часть мышц, а именно мышцы глазного яблока, жевательные, мимические мышцы верхней части лица, мышцы глот- ки, гортани, шеи, туловища и промежности, получают так называемую двустороннюю иннервацию, т.е. от центральных нейронов как своей, так и противоположной стороны. Это в известной степени обеспечивает сохранение иннервации этих мышц при одностороннем поражении центральных нейронов.
Односторонняя иннервация у мышц конечностей, языка и мимических мышц нижней половины лица. В связи с этим при одно-
стороннем поражении центральных нейронов этих мышц полностью утрачивается способность к движению, т.е. наступает паралич.
2. Проводящие пути экстрапирамидной системы проходят к передним рогам спинного мозга вне пирамидных путей, через покрышку и ретикулярную формацию мозгового ствола. К экстрапирамидной системе относят хвостатое и чечевицеобразное ядра, таламус, черное вещество, красное ядро, образования гипоталамуса, мозжечок, нижнюю оливу и ядра ретикулярной формации мозгового ствола. Экстрапирамидная система является филогенетически более старым образованием по сравнению с пирамидной. Функция экстрапирамидной системы заключается в осуществлении сложных безусловнорефлекторных двигательных актов, основанных на опыте вида, а двигательная сознательная корковая деятельность основана на условных рефлексах, закрепляющих индивидуальный опыт.
Экстрапирамидная система обеспечивает автоматические, без участия сознания, регуляцию и координацию тонуса мышечной ткани, осуществляет сокращение или расслабление определенных групп мышц, обеспечивает готовность двигательного аппарата к выполнению сложных, точных, целенаправленных действий. Поражения экстрапирамидной системы проявляются главным образом изменениями мышечного тонуса и двигательной активности.
ТИПОВЫЕ ОСОБЕННОСТИ СТРОЕНИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Центральная нервная система у разных людей имеет некоторые специфические структурно-функциональные различия. Эти разли- чия касаются формы мозга и его отдельных образований, величины, массы и других показателей.
Так, масса головного мозга человека, как правило, составляет от 1100 до 2300 г. Следует отметить, что большая масса мозга (в пределах нормы) не свидетельствует о высоком интеллекте, и наоборот. Между формой головного мозга и формой черепа наблюдается определенное соответствие. Долихоцефалической, мезоцефалической и брахицефалической формам черепа соответствует определенное положение его отделов, образований, борозд и т.д. Это обстоятельство имеет важное практическое значение в связи с возможностью проекций на наружные покровы головы различных ядер, борозд и извилин.
ОБОЛОЧКИ ГОЛОВНОГО МОЗГА
Твердая оболочка головного мозга, dura mater encephali, прилегает к внутренней поверхности костей черепа и плотно сращена с ней в области основания и швов. Рыхлое соединение оболочки с костями свода черепа послужило основанием для выделения очень узкого эпидурального пространства, которое пронизано соединительнотканными волокнами, сосудами и нервами. К гладкой внутренней поверхности твердой оболочки прилегает паутинная оболочка головного мозга. Узкую щель между ними условно называют субду- ральным пространством.
В некоторых местах твердая оболочка образует отроги (отростки), которые проникают между различными отделами головного мозга. Так, в продольной щели полушарий расположен серповидной формы отросток - серп большого мозга, falx cerebri. Верхний утолщенный край серпа большого мозга, образованный двумя расходящимися пластинками твердой оболочки, примыкает к сагиттальной борозде черепа от петушиного гребня решетчатой кости до внутреннего затылочного выступа (рис. 218).
Намет мозжечка, tentorium cerebelli, представляет собой широкую складку твердой мозговой оболочки, которая в виде двускатной крыши располагается в поперечной борозде мозга, отделяя содержимое задней черепной ямки от затылочных долей. Заднелатеральный утолщенный край мозжечкового намета сращен с бороздой поперечного синуса и верхней гранью пирамиды височной кости. Свободный край имеет округлую форму. Он натянут между задним концом свободного края мозгового серпа и задним наклонным отростком клиновидной кости. Свободный край ограничивает вырезку намета мозжечка, incisura tentorii, через которую сообщаются задняя и средняя ямки черепа. В области этой вырезки расположены ножки мозга. В задней ямке черепа, между полушариями мозжечка, лежит серп мозжечка, falx cerebelli.
Диафрагма турецкого седла, diaphragma sellae, натянута над турецким седлом. В середине диафрагмы имеется узкое отверстие, через которое проходит воронка к гипофизу. На верхушке пирамиды височной кости отростки твердой оболочки образуют тройничную полость, cavum trigeminale; в ней расположен тройничный узел.
В толще твердой оболочки проходят артерии, вены, нервные волокна и их окончания, а также особые каналы - венозные синусы (пазухи).
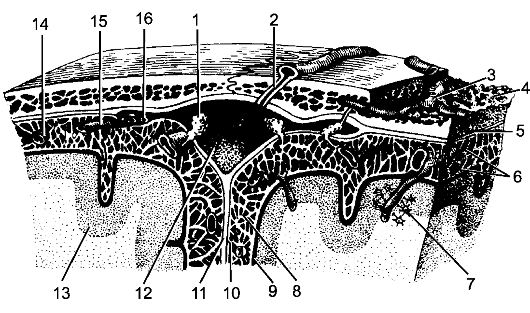
Рис. 218. Оболочки головного мозга.
1 - грануляция паутинной оболочки; 2 - эмиссарная вена; 3 - вена губчатого вещества кости; 4 - губчатое вещество кости; 5 - твердая оболочка; 6 - перекладины паутинной оболочки; 7 - околососудистое пространство; 8 - подпаутинное пространство; 9 - сосудистая оболочка; 10 - паутинная оболочка; 11 - серп большого мозга; 12 - верхний сагиттальный синус; 13 - кора мозга; 14 - ветвь артерии мозга; 15, 16 - вены мозга.
Венозные синусы представляют собой пространства между расщепившимися листками твердой оболочки головного мозга. Большая часть венозных синусов расположена пристеночно. В венозные синусы поступает кровь из вен головного мозга и затем выводится в экстракраниальные венозные образования, главным образом во внутренние яремные вены.
Паутинная оболочка головного мозга, arachnoidea mater encephali, в виде тонкого прозрачного бессосудистого листа покрывает головной мозг и книзу переходит в паутинную оболочку спинного мозга. С твердой оболочкой головного мозга паутинная оболочка прочно сращена посредством особых структур - грануляций паутинной оболочки.
Паутинная и мягкая оболочки ограничивают заполненное спинномозговой (церебральной) жидкостью подпаутинное пространство. Это пространство пронизано многочисленными соединительно- тканными волокнами, которые соединяют обе оболочки и обеспечивают фиксацию расположенных здесь кровеносных сосудов. Иные соотношения этих оболочек в области различных углублений мозга. Мягкая оболочка покрывает мозговую поверхность в глубине борозд, щелей и ямок, а паутинная, не следуя за ней в глубину борозд и щелей, как бы мостиками перекидывается с одной возвышенности извилины на другую, образуя различной величины под-
паутинные пространства. Самыми крупными из этих вместилищ или цистерн являются следующие:
1) мозжечково-мозговая цистерна, cisterna cerebellomedullaris, находится между мозжечком и задней поверхностью продолговатого мозга;
2) межножковая цистерна, cisterna interpeduncularis, расположена между ножками мозга;
3) цистерна перекреста, cisterna chiasmatis, находится впереди зрительного перекреста;
4) цистерна латеральной ямки большого мозга, cisterna fossae lateralis cerebri, соответствует одноименной ямке и борозде.
Субарахноидальное пространство является единым для головного и спинного мозга. Образующаяся в желудочках головного мозга спинномозговая жидкость поступает в это пространство через отверстия IV желудочка, а отводится в венозную систему посредством грануляций паутинной оболочки.
Мягкая оболочка головного мозга, pia mater encephali, плотно прилегает к мозговому веществу как на свободной поверхности мозга, так и в глубине борозд. Сосудистые сплетения желудочков осуществляют секрецию спинномозговой жидкости. В толще мягкой обо- лочки расположены сети кровеносных сосудов и нервы.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
1. Назовите доли конечного мозга.
2. Какие крупные извилины расположены в лобной доле головного мозга?
3. Какие основные борозды и извилины медиальной поверхности полушария головного мозга вы знаете?
4. Назовите слои или пластинки коры конечного мозга.
5. Укажите локализацию в коре конечного мозга центральных отделов зрительного, слухового, обонятельного и вкусового анализаторов.
6. Какие структуры относят к обонятельному мозгу?
7. Назовите базальные ядра головного мозга.
8. Какие вам известны пучки длинных ассоциативных волокон?
9. Какие проводящие пути расположены в задней ножке внутренней капсулы?
10. Опишите корково-спинномозговые и корково-ядерные (пирамидные) проводящие пути.
11. Какие структуры относят к экстрапирамидной системе?
12. Какие подпаутинные цистерны вам известны?
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
ОБЩИЕ ДАННЫЕ О СТРОЕНИИ ПЕРИФЕРИЧЕСКИХ НЕРВОВ
По месту возникновения в ЦНС периферические нервы подразделяются на спинномозговые нервы, берущие начало от спинного мозга, и черепные нервы, отходящие от головного мозга.
Спинномозговые нервы, пп. spinales, образуются в результате слияния корешков спинного мозга: чувствительного заднего, radix posterior; и двигательного переднего, radix anterior. Всего насчитывается 31 пара спинномозговых нервов: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и копчиковая. Нервы сохраняют четкую сегментарность выхода и положения соответственно метамерам позвоночного ряда. Каждому сегменту спинного мозга и принадлежащим ему двум парам корешков с двумя спинномозговыми нервами - невротому - соответствуют определенный участок кости - склеротом, мышцы - миотом, кожи - дерматом. Однако к каждому метамеру (склеротому, миотому, дерматому) подходят нервные волокна не только от соответствующего ему невротома, но и от двух соседних - выше- и нижележащего.
Спинномозговые нервы являются смешанными и содержат чувствительные - афферентные, двигательные - эфферентные нервные волокна, а также волокна, относящиеся к вегетативной нервной системе. По выходе из межпозвоночного отверстия каждый спинномозговой нерв разделяется на 4 ветви: 1) переднюю, r. anterior, - к передним отделам туловища и конечностям; 2) заднюю, r. posterior, - к задним отделам туловища; 3) менингеальную, r. meningeus, возвратную - к оболочкам спинного мозга; 4) соединительные, rr. сот- municantes, - к узлам симпатического ствола (рис. 219).
Черепные нервы, пп. craniales, - берут начало в головном мозге от принадлежащих им ядер, выходят из мозга, покидают полость черепа через отверстия в его основании и разветвляются в основном на голове и шее, а блуждающий нерв - также в грудной и брюшной полостях.
Каждый нерв слагается из мякотных и безмякотных нервных волокон, состоящих из длинных отростков нервных клеток - аксонов (нейритов) и покрывающих их оболочек - нейролеммы, а для мякотных - еще и миелиновой оболочки.
Нейролемма образована вытянутыми в длину звездчатыми клетками - нейролеммоцитами. Она интимно связана с аксоном и имеет отношение к процессам обмена веществ, а также регенерации.
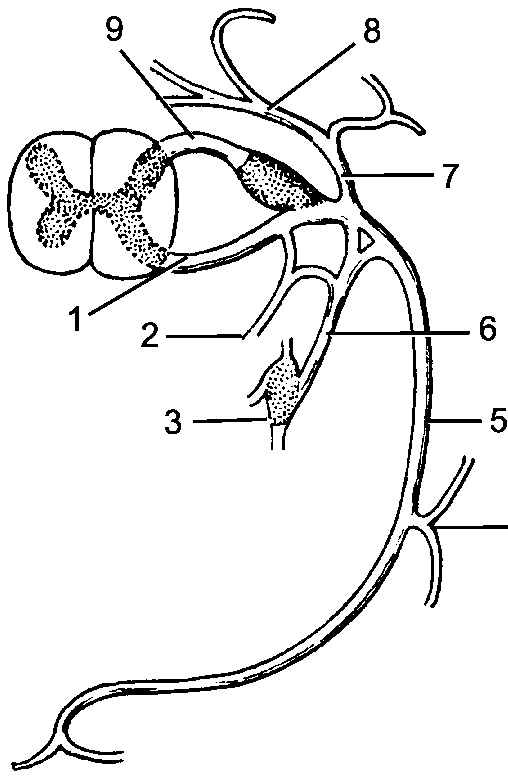
Рис. 219. Образования спинномозгового нерва и его ветвей. 1 - передний корешок; 2 - менингеальная ветвь; 3 - узел симпатического ствола; 4 - латеральная кожная ветвь; 5 - передняя ветвь; 6 - соединительная ветвь; 7 - задняя ветвь; 8 - медиальная ветвь задней ветви; 9 - задний корешок.
Миелиновая оболочка состоит из особого липоидного вещества - миелина, которое покрывает нервное волокно, но не на всем протяжении: миелин отсут- ствует при выходе аксона из нервной клетки, при переходе в нервные окончания и в местах соединения нейролеммоцитов, в так называемых перехватах волокна. В функциональном отношении мякотная оболочка может рассматри- ваться как изолятор проходящих по аксону волн возбуждения, что обеспечивает изолированность проведения биотоков.
Толщина мякотной оболочки для различных волокон неодинакова. В зависимости от ее толщины, а также диаметра самого аксона различают волокна тонкие (диаметром до 3 мкм), средние (4-10 мкм) и толстые (более 10 мкм). Скорость проведения импульсов у разных нервных волокон различна: скорость проведения по толстым волокнам 30-80 м/с, средним - 10-14 м/с, тонким - 0,7-1,3 м/с. Тонкие мякот-
ные и безмякотные волокна являются проводниками болевой чувствительности, средние - проприоцептивной и температурной чувствительности, толстые с тонкой миелиновой оболочкой - тактильной чувствительности, а волокна с толстой миелиновой оболочкой - это двигательные проводники.
Внутриствольное строение нервов. Группы нервных волокон соединяются в нерве в пучки, ограниченные пластинчатой оболочкой - периневрием. В периневральном влагалище располагаются нервные волокна и рыхлая соединительная ткань - эндоневрий, окружающий каждое нервное волокно. Пучки нервных волокон окружены рыхлой соединительной тканью - эпиневрием, которым они объединяются в нервный ствол. В эпиневрии находятся кровеносные и лимфатические сосуды и нервы, обеспечивающие иннервацию оболочек и сосудов нерва. Количество пучков в нервах различное.
Нервы по составу образующих их нервных волокон являются смешанными и содержат чувствительные, двигательные и ве-
гетативные волокна. Соотношение безмякотных и мякотных волокон в различных нервах неодинаково. В некоторых нервах: срединном, большеберцовом, кожных и сосудистых больше безмякотных волокон, в других (лучевой, глубокий малоберцовый) их сравнительно мало. Неодинаково и процентное соотношение мякотных волокон различного вида: в сосудистых и кожных нервах много тонких и средних мякотных волокон; в лучевом, глубоком малоберцовом нервах и мышечных ветвях преобладают толстые мякотные волокна; в срединном, локтевом, седалищном и большеберцовом содержится значительное количество и тонких, и толстых волокон. Поэтому различают преимущественно двигательные, чув- ствительные и смешанные нервы.
Понятие о комплексах нервов. Изучение периферической нервной системы выявило значительные различия во внешнем строении нервов. Даже одноименные нервы у разных людей могут иметь неодинаковое формирование и ветвление. Между нервами наблюдаются связи, по которым определенное количество нервных волокон переходит из одного нерва в другой.
Необходимо учитывать также, что разные нервы могут начинаться в одних и тех же сегментах спинного мозга, т.е. иметь сегментар- ную общность. При этом нервные волокна определенных сегментов могут распределяться у разных людей в начинающихся от них нервах неодинаково, т.е. входить в состав различных нервов. Один и тот же нерв у разных людей не всегда иннервирует строго определенный участок на периферии: у одного человека он может распространяться на большем, у другого - на меньшем протяжении. Со- седние нервы при этом как бы замещают друг друга.
Выделяют комплексы нервов, под которыми понимают систему нервных стволов, имеющих генетическое родство и сегментарное единство. На верхней конечности выделяют комплексы: мышечно-кожный и срединный нервы - в области плеча, срединный и локтевой - в области предплечья и кисти, лучевой и подмышечный нервы; на нижней ко- нечности: бедренный, запирательный нервы и наружный кожный нерв бедра - в передней области бедра, седалищный нерв, задний кожный нерв бедра - в задней, большеберцовый, поверхностный и глубокий малоберцовый нервы - в области голени и стопы и т.д.
Кровоснабжение нервов. Нервные стволы, как правило, проходят в составе сосудисто-нервных пучков. Основные сосудисто-нервные пучки заключены в собственные фасциальные влагалища.
Сосудисто-нервный пучок, включающий нерв, артериальные и венозные стволы и лимфатические сосуды, представляет собой ана-
томически и функционально единое целое. Между его компонентами существует взаимная связь: нервы обеспечивают иннервацию соседних артерий, вен и лимфатических сосудов, артерия снабжает перечисленные образования кровью, в вены и лимфатические сосуды происходит венозный и лимфатический отток из всего пучка в целом.
Источниками кровоснабжения нервов являются близлежащие артерии и их ветви. Артерии подходят к нервным стволам сегментарно и сразу же в эпиневрии разделяются на две ветви - восходящую и нисходящую, которые анастомозируют с аналогичными ветвями выше- и нижележащих артерий. Эти артерии образуют сеть эпиневральных артерий, которые отдают пери- и эндоневральные ветви.
Вены. Различают эндо-, пери- и эпиневральные вены. Венозные сети в нервах более обильны, чем артериальные. Эпиневральные вены вливаются в вены, расположенные рядом с нервом.
Лимфатические сосуды локализуются в эпиневрии. В периневрии между его слоями образуются лимфатические щели, сообщаю- щиеся с лимфатическими сосудами эпиневрия и эндоневральными лимфатическими щелями. По ходу нервов может распространяться инфекция.
СПИННОМОЗГОВЫЕ НЕРВЫ
РАЗВИТИЕ СПИННОМОЗГОВЫХ НЕРВОВ
Как отмечалось, в начале 1-го месяца эмбрионального развития происходит образование нервной пластинки, при замыкании которой в нервную трубку выделяются зачатки спинномозговых и симпатических узлов. В дальнейшем пути миграции нейробластов и роста их отростков образуются предпозвоночные и интрамуралъные сплетения вегетативной (автономной) нервной системы.
Различные части нервной трубки растут неравномерно, что приводит к выделению основных отделов будущего спинного мозга: боковые стенки образуют белое вещество, а вентральные и дорсальные части - передние и задние рога. Зачатки спинного мозга образованы клетками двух родов: одни - спонгиобласты - формируют нейроглию, другие - нейробласты - развиваются в нейроны (нейроциты).
На 3-4-й неделе развития отростки нейробластов нервной трубки выходят из нее и все вместе образуют метамерно расположенные
передние корешки спинного мозга. Нейробласты, лежащие в зачатках спинномозговых узлов, также отдают длинные отростки, которые формируют задние корешки. На 5-6-й неделе происходит слияние переднего и заднего корешков с образованием смешанных спинномозговых нервов и их основных ветвей: передней, задней, соединительных, менингеальной.
На 2-м месяце развития дифференцируются зачатки конечностей, в которые врастают нервные волокна из соответствующих зачаткам сегментов спинного мозга. В первой половине 2-го месяца в связи с перемещением и сдвигами метамеров, формирующих конечности, образуются нервные сплетения.
У эмбриона длиной 15-20 мм все нервные стволы конечностей и туловища соответствуют положению нервов у новорожденного. Формирование нервов туловища и нервов нижней конечности совершается подобным путем, но несколько позже (на 2 нед).
Сравнительно рано, у эмбрионов длиной 8-10 мм, наблюдается проникновение мезенхимных клеток вместе с кровеносными сосу- дами в закладки органов и конечностей. Мезенхимные клетки делятся и образуют внутриствольные оболочки нерва: эндо-, пери- и эпиневрий. Глиальные элементы зачатков (спонгиобласты) идут на построение нейролеммы - оболочек длинных отростков нервных клеток. Миелинизация нервных волокон начинается с 3-4-го месяца эмбрионального развития и заканчивается после рождения. Раньше миелинизируются черепные нервы, нервы верхней конечности, позже - нервы туловища и нижних конечностей.
ЗАДНИЕ ВЕТВИ СПИННОМОЗГОВЫХ НЕРВОВ
Задние ветви спинномозговых нервов, rami posteriores, иннервируют кожу затылка, задней поверхности шеи и спины и глубокие мышцы спины, развивающиеся из дорсальных частей миотомов.
Задние ветви шейных спинномозговых нервов имеют ряд особенностей. Задняя ветвь I спинномозгового нерва, называемая подзатылочным нервом, п. suboccipitalis, не дает кожных ветвей. Она иннервирует mm. recti capitis posteriores major et minor, semispinalis capitis, obliqui capitis superior et inferior.
Задняя ветвь II спинномозгового нерва обозначается как большой затылочный нерв, п. occipitalis major, не имеет мышечных ветвей, а иннервирует кожу затылка. Задние ветви других спинномозговых нервов отдают как кожные, так и мышечные ветви, иннервирующие кожу и мышцы задней части шеи.
Задние ветви грудных спинномозговых нервов разделяются на кожные ветви: медиальную, r. cutaneus medialis, и латеральную, r. cutaneus lateralis, которые в свою очередь отдают более мелкие ветви к собственным мышцам спины.
Задние ветви поясничных и крестцовых спинномозговых нервов также разделяются на медиальные и латеральные ветви. Латеральные ветви 3 верхних поясничных нервов иннервируют кожу верхних отделов ягодичной области и называются верхними ветвями ягодиц, пп. clunium superiores, a 3 верхних крестцовых - кожу средних отделов ягодичной области - это средние ветви ягодиц, пп. clunium medii.
ПЕРЕДНИЕ ВЕТВИ СПИННОМОЗГОВЫХ НЕРВОВ
Передние ветви спинномозговых нервов, rr. anteriores, иннервируют кожу и мышцы переднего отдела шеи и туловища, а также кожу и мышцы конечностей. Передние ветви сохраняют полную метамерию только в грудном отделе, где они называются межреберными нервами, пп. intercostales.
В области конечностей в связи с перемещением метамеров сегментность теряется и передние ветви формируют сплетения спинномозговых нервов, plexus nervorum spinalium (шейное, плечевое, поясничное, крестцовое), в которых происходит большой обмен нервными волокнами разных сегментов.
ШЕЙНОЕ СПЛЕТЕНИЕ
Шейное сплетение, plexus cervicalis, формируется передними ветвями 4 верхних шейных спинномозговых нервов (CI-CIV), имеющими между собой соединения. Сплетение залегает сбоку от поперечных отростков между позвоночными (сзади) и предпозвоночными (спереди) мышцами. Нервы выходят из-под заднего края грудино- ключично-сосцевидной мышцы, немного выше ее середины, и распространяются веерообразно вверх, вперед и вниз. От сплетения отходят следующие нервы (рис. 220):
1. Малый затылочный нерв, п. occipitalis minor (от CI-CII), распространяется вверх к сосцевидному отростку и далее в боковые отделы затылка, где иннервирует кожу указанного отдела.
2. Большой ушной нерв, п. auricularis magnus (от СIII-CIV), идет по грудино-ключично-сосцевидной мышце вверх и кпереди, к ушной раковине, иннервирует кожу ушной раковины (задняя ветвь) и кожу над околоушной слюнной железой (передняя ветвь).
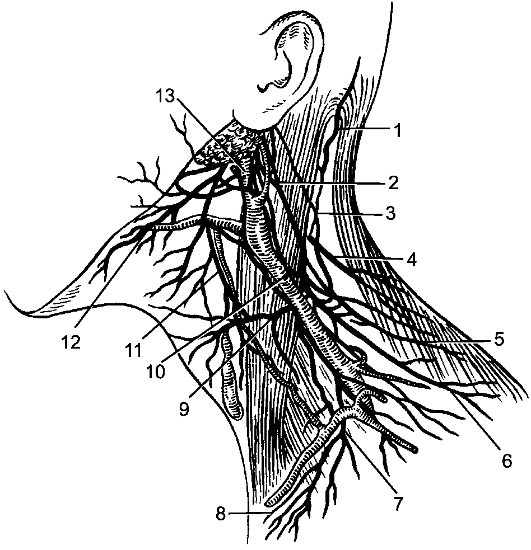
Рис. 220. Кожные ветви шейных нервов (по Л.А. Шангиной). 1 - большой затылочный нерв; 2 - большой ушной нерв; 3 - малый затылочный нерв; 4, 6 - латеральные (задние) надключичные нервы; 5 - добавочный нерв; 7 - промежуточный надключичный нерв; 8 - медиальные надключичные нервы; 9 - поперечный нерв шеи; 10, 11 - связи шейных нервов с лицевым нервом; 12 - шейная ветвь лицевого нерва; 13 - наружная яремная вена.
3. Поперечный нерв шеи, п. transversus colli (от CIII-CIV), идет кпереди и у переднего края грудино-ключично-сосцевидной мышцы разделяется на верхние и нижние ветви, иннервирующие кожу переднего отдела шеи.
4. Надключичные нервы, пп. supraclaviculares (от CIII-CIV), числом от 3 до 5 распространяются вниз веерообразно под подкожной мышцей шеи; разветвляются в коже задненижней части шеи (латеральные ветви), в области ключицы (промежуточные ветви) и верхнепередней части груди до jjj ребра (медиальные ветви).
5. Диафрагмальный нерв, п. phrenicus (от CIII-CIV и частично от CV), преимущественно двигательный нерв, идет вниз по передней лестничной мышце в грудную полость, где проходит к диафрагме впереди корня легкого между медиастинальной плеврой и перикардом. Иннервирует мышцы диафрагмы, отдает чувствительные
ветви к плевре и перикарду (rr. pericardiaci), иногда к шейно-грудному нервному сплетению. Кроме того, посылает диафрагмальнобрюшные ветви, rr. phrenicoabdominales, к диафрагмальной брюшине. Эти ветви содержат нервные узлы (ganglii phrenici) и соединяются с чревным нервным сплетением. Особенно часто такие связи име- ет правый диафрагмальный нерв, чем и объясняется френикуссимптом - иррадиация болей в область шеи при заболевании печени.
6. Нижний корешок шейной петли, radix inferior ansae cervicalis, образуется нервными волокнами от второй и третьей передних ветвей спинномозговых нервов и идет кпереди на соединение с верхним корешком, radix superior, возникающим от подъязычного нерва (XII пара черепных нервов). В результате соединения обоих корешков образуется шейная петля, ansa cervicalis, от которой отходят ветви к лопаточно-подъязычной, грудино-подъязычной, щитоподъязычным и грудино-щитовидной мышцам.
7. Мышечные ветви, rr. musculares, идут к предпозвоночным мышцам шеи, к мышце, поднимающей лопатку, а также к грудиноключично-сосцевидной и трапециевидной мышцам.
ПЛЕЧЕВОЕ ПЛЕТЕНИЕ
Плечевое сплетение, plexus brachialis, формируется передними ветвями 4 нижних шейных спинномозговых нервов и передней ветвью первого грудного спинномозгового нерва. В сплетении разли- чают 2 части: надключичную, pars supraclavicularis, и подключичную, pars infraclavicularis. Первая располагается в пределах латерального треугольника шеи, вторая - в подмышечной ямке.
Надключичная часть сплетения слагается из 3 стволов: верхнего, truncus superior, среднего, truncus medius, и нижнего, truncus inferior. Верхний и средний стволы лежат в межлестничной щели выше подключичной артерии, нижний - позади нее. От надключичной части сплетения начинаются короткие ветви (рис. 221).
Подключичная часть сплетения представлена 3 пучками: латеральным, fasciculus lateralis, задним, fasciculus posterior, и медиальным, fasciculus medialis. От подключичной части сплетения берут начало длинные нервы верхней конечности. От латерального пучка начинаются мышечно-кожный нерв и латеральный корешок срединного нерва; от медиального - медиальный корешок срединного нерва, локтевой и медиальные кожные нервы плеча и предплечья; от заднего - лучевой и подмышечный нервы.
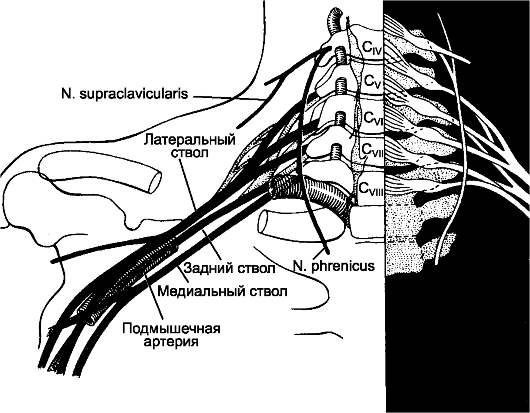
Рис. 221. Плечевое сплетение.
Короткие ветви:
1. Дорсальный нерв лопатки, п. dorsalis scapulae, иннервирует мышцу, поднимающую лопатку, большую и малую ромбовидные мышцы.
2. Длинный грудной нерв, п. thoracicus longus, иннервирует переднюю зубчатую мышцу.
3. Подключичный нерв, п. subclavius, иннервирует одноименную мышцу.
4. Надлопаточный нерв, п. suprascapularis, иннервирует надостную и подостную мышцы, капсулу плечевого сустава.
5. Подлопаточный нерв, п. subscapularis, иннервирует подлопаточную, большую круглую и широкую мышцы спины.
6. Грудные нервы - медиальный и латеральный, пп. pectorales medialis et lateralis, иннервирует большую и малую грудные мышцы.
7. Подмышечный нерв, п. axillaris, иннервирует дельтовидную мышцу, малую круглую мышцу, капсулу плечевого сустава и кожу наружной поверхности плеча.
Длинные ветви (рис. 222, 223):
1. Мышечно-кожный нерв, п. musculocutaneus, иннервирует переднюю группу мышц плеча. Продолжение этого нерва носит назва-
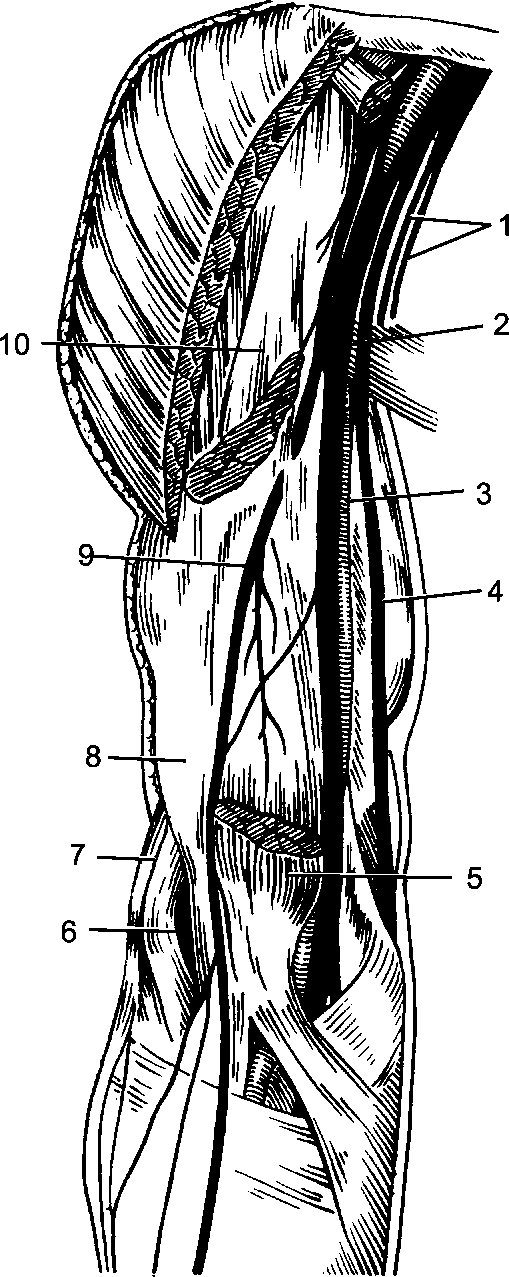
Рис. 222. Длинные ветви плечевого сплетения.
1 - медиальный кожный нерв плеча и медиальный кожный нерв предплечья; 2 - срединный нерв; 3 - плечевая артерия; 4 - локтевой нерв; 5 - двуглавая мышца плеча (дистальный конец); 6 - лучевой нерв; 7 - ветвь лучевого нерва; 8 - плечевая мышца; 9 - мышечно-кожный нерв; 10 - проксимальный конец двуглавой мышцы плеча.
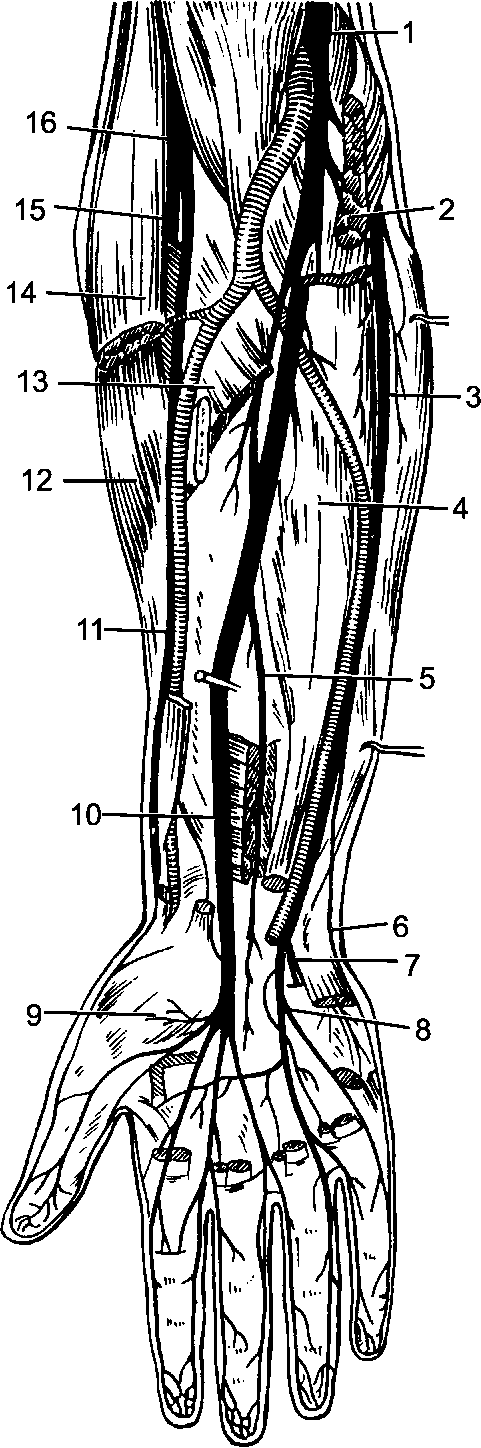
Рис. 223. Нервы предплечья и кисти. 1 - срединный нерв; 2 - круглый пронатор (отсечен); 3 - локтевой нерв; 4 - глубокий сгибатель пальцев; 5 - передний межкостный нерв; 6 - тыльная ветвь локтевого нерва; 7 - глубокая ветвь локтевого нерва; 8 - поверхностная ветвь локтевого нерва; 9 - мышцы возвышения большого пальца; 10 - квадратный пронатор; 11 - поверхностная ветвь лучевого нерва; 12 - длинный лучевой разгибатель запястья; 13 - поверхностный сгибатель пальцев (отсечен); 14 - плечелучевая мышца (отсечена); 15 - глубокая ветвь лучевого нерва; 16 - лучевой нерв.
ние латерального кожного нерва предплечья, п. cutaneus antebrachii lateralis. Он иннервирует кожу переднелатеральной поверхности предплечья.
2. Срединный нерв, п. medianus, на плече ветвей не отдает. На предплечье нерв отдает ветви к передней группе мышц, за исключением локтевого сгибателя запястья и медиальной половины глубокого сгибателя пальцев, которые иннервируются локтевым нервом. На кисти срединный нерв делится на 3 общих ладонных пальцевых нерва, пп. digitales palmares communes, которые в свою очередь делятся на собственные ладонные пальцевые нервы, пп. digi- tales palmares proprii, идущие к коже I, II, III и наружной поверхности IV пальцев.
От первого общего пальцевого нерва отходят мышечные ветви к мышцам возвышения большого пальца, за исключением мышцы, приводящей большой палец, и глубокой головки короткого сгибателя большого пальца. Ветви этого нерва идут также к I и II червеобразным мышцам.
3. Локтевой нерв, п. ulnaris, на плече ветвей не отдает. На предплечье отдает ветви к локтевому суставу, локтевому сгибателю запястья и части глубокого сгибателя пальцев. Иннервирует кожу медиальной поверхности IV и V пальцев, мышцы возвышения мизинца, межкостные мышцы, III и IV червеобразные мышцы, приводящую мышцу большого пальца и глубокую головку короткого сгибателя большого пальца.
4. Медиальный кожный нерв плеча, п. cutaneus brachii medialis, иннервирует кожу медиальной поверхности плеча.
5. Медиальный кожный нерв предплечья, п. cutaneus antebrahii medialis, иннервирует кожу медиальной поверхности предплечья.
6. Лучевой нерв, п. radialis, иннервирует задние группы мышц плеча и предплечья, кожу задней и нижнелатеральной поверхности плеча, кожу задней поверхности предплечья, кожу тыльной поверхности I, II и наружной стороны III пальцев.
МЕЖРЕБЕРНЫЕ НЕРВЫ
Межреберные нервы, nn. intercostales, являются передними ветвями 1-12-го грудных спинномозговых нервов. Они иннервируют межреберные мышцы, поперечную мышцу груди, мышцы передней брюшной стенки. Участвуют в иннервации плевры и брюшины. Кожные ветви межреберных нервов иннервируют кожу туловища
(рис. 224).
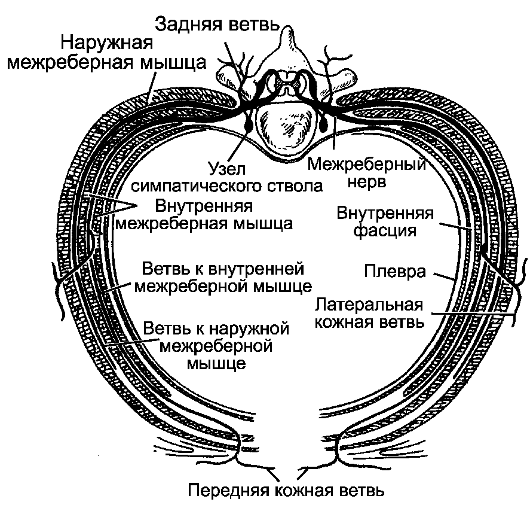
Рис. 224. Межреберные нервы.
ПОЯСНИЧНОЕ СПЛЕТЕНИЕ
Поясничное сплетение, plexus lumbalis, образовано передними ветвями 1-го, 2-го, 3-го и частично 4-го поясничных спинномозговых нервов. В образовании сплетения может принимать участие передняя ветвь 12-го грудного спинномозгового нерва (рис. 225).
Ветви поясничного сплетения:
1. Подвздошно-подчревный нерв, п. iliohypogastricus, иннервирует кожу над большим вертелом бедренной кости и в области поверхностного пахового кольца.
2. Подвздошно-паховый нерв, п. ilioinguinalis, отдает мышечные ветви к мышцам брюшной стенки, к коже мошонки или больших половых губ.
3. Бедренно-половой нерв, п. genitofemoralis, иннервирует семенной канатик, оболочки яичка и кожу бедра ниже паховой складки.
4. Латеральный кожный нерв бедра, п. cutaneus femoris lateralis, иннервирует кожу латеральной поверхности бедра.
5. Бедренный нерв, п. femoralis, отдает ветви к передней и частично медиальной группам мышц бедра. Кожные ветви иннервируют переднемедиальную поверхность кожи бедра. Одна из кожных ветвей - подкожный нерв, п. saphenus, иннервирует кожу переднемедиальной поверхности голени.
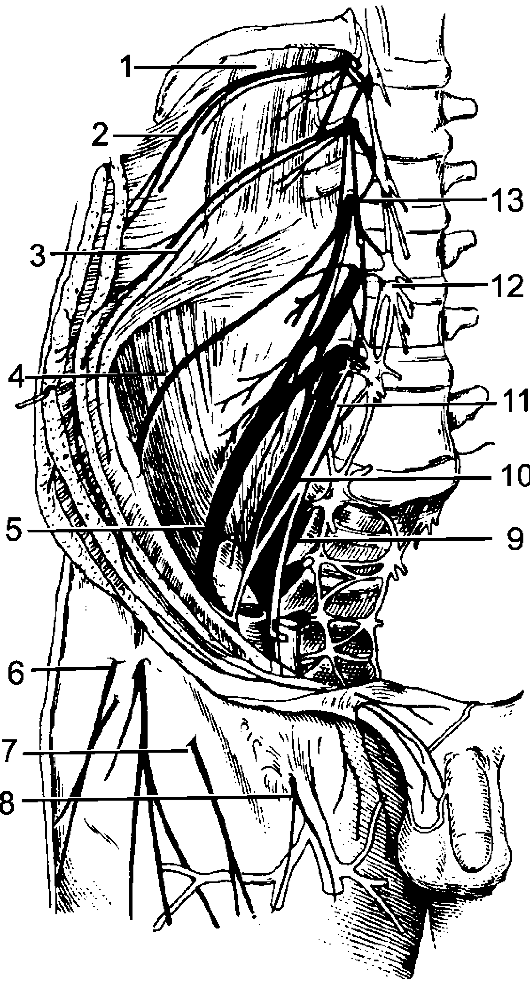
Рис. 225. Поясничное сплетение.
1 - квадратная мышца поясницы;
2 - 12-й межреберный нерв; 3 - под- вздошно-подчревный и подвздошнопаховый нервы; 4, 6 - латеральный кожный нерв бедра; 5 - бедренный нерв; 7 - передняя кожная ветвь бедренного нерва; 8 - бедренная ветвь бедренно-полового нерва; 9 - крестцовое сплетение; 10, 11 - пояснично- крестцовый ствол; 12 - поясничный узел симпатического ствола; 13 - передняя ветвь 2-го поясничного нерва.
6. Запирательный нерв, п. obturatorius, иннервирует запирательные мышцы и мышцы медиальной группы бедра, отдает ветви к тазобедренному суставу и коже медиальной поверхности бедра.
КРЕСТЦОВОЕ СПЛЕТЕНИЕ
Крестцовое сплетение, plexus sacralis, образовано передними ветвями крестцовых спинномозговых нервов. В нем различают собственно крестцовое, половое и копчиковое сплетения. От крестцового сплетения отходят короткие и длинные ветви (рис. 226).
Короткие ветви:
1. Мышечные ветви, rr. musculares, идут к грушевидным, близнецовым, внутренней запирательной и квадратной мышцам бедра.
2. Верхний ягодичный нерв, п. gluteus superior, иннервирует малую и среднюю ягодичные мышцы, а также мышцу, напрягающую широкую фасцию.
3. Нижний ягодичный нерв, п. gluteus inferior, иннервирует большую ягодичную мышцу, квадратную мышцу бедра, близнецовые мышцы и капсулу тазобедренного сустава.
Длинные ветви:
1. Задний кожный нерв бедра,
п. cutaneus femoris posterior, иннервирует кожу нижней части ягодиц, кожу промежности, задней поверхности бедра и подколенной ямки.
2. Седалищный нерв, п. ischiadicus, на бедре отдает мышечные ветви задней группе мышц бедра и ветвь к коленному суставу.
В подколенной ямке нерв делится на общий малоберцовый и большеберцовый нервы.
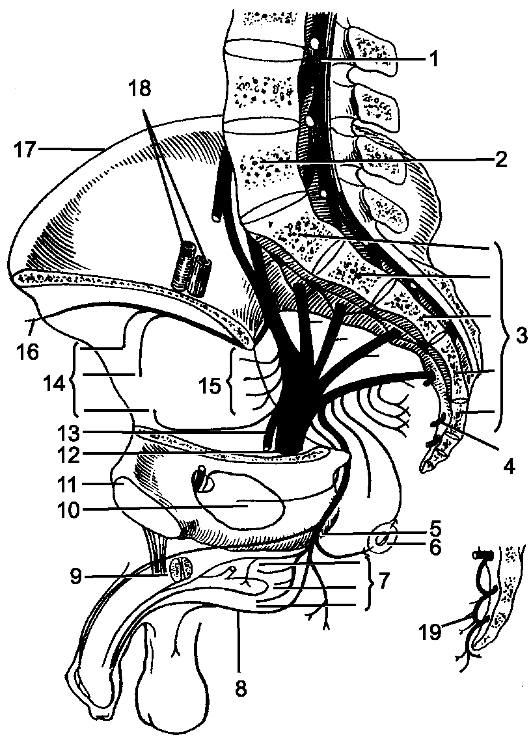
Рис. 226. Крестцовое и копчиковое сплетения.
1 - конский хвост; 2 - V поясничный позвонок; 3 - крестцовые позвонки; 4 - передняя ветвь копчикового нерва; 5 - половой нерв; 6 - прямая кишка и ее нервы; 7 - ветви нервов к мышцам промежности; 8 - ветвь к половым органам; 9 - пещеристые тела полового члена; 10 - внутренняя запирательная мышца; 11 - симфиз; 12 - седалищный нерв; 13 - задний кожный нерв бедра; 14 - ветви нервов к ягодичным мышцам; 15 - ветви нервов к мышцам задней поверхности тазобедренного сустава; 16 - ветвь к широкой фасции; 17 - подвздошный гребень; 18 - наружные подвздошные сосуды; 19 - копчиковое сплетение.
Большеберцовый нерв, п. tibialis, иннервирует заднюю группу мышц голени, капсулы коленного и голеностопного суставов, кожу заднемедиальной поверхности голени, вместе с ветвью малоберцового нерва иннервирует кожу латеральной поверхности пятки и латерального края стопы.
Конечные ветви большеберцового нерва иннервируют кожу подошвенной поверхности пальцев стопы.
Общий малоберцовый нерв, п. fibularis (peroneus) communis, ответвляется от седалищного в подколенной ямке или выше и делится на поверх-
ностный и глубокий малоберцовые нервы. От общего малоберцового нерва отходят ветви к капсуле коленного и большеберцово-малоберцового суставов и коже латеральной поверхности голени и стопы.
Поверхностный малоберцовый нерв, п. fibularis (peroneus) supeficialis, отдает ветви к латеральной группе мышц голени, к коже медиальной поверхности I пальца, латеральной поверхности II и медиальной III пальцев, к IV пальцу и медиальной поверхности V пальца.
Глубокий малоберцовый нерв, п. fibularis profundus, отдает ветви к латеральной поверхности I и медиальной II пальцев, к капсулам межпредплюсневых и предплюсне-плюсневых суставов. Мышечные ветви глубокого малоберцового нерва иннервируют переднюю группу мышц голени.
ПОЛОВОЕ СПЛЕТЕНИЕ
Половое сплетение, plexus pudendus, образуется передними ветвями третьего и четвертого крестцовых нервов и соединяется с собственно крестцовым и копчиковым сплетениями, а также с симпатическим стволом. Мышечные ветви сплетения иннервируют мышцу, поднимающую задний проход, и копчиковую мышцу.
Промежностный нерв, п. perinealis, отдает ветви к коже мошонки, а у женщин к большим половым губам, а также мышечные ветви к поверхностной поперечной мышце промежности, луковично-губчатой и седалищно-пещеристой мышцам.
Дорсальный нерв полового члена, п. dorsalis penis, проходит по спинке полового члена, отдает мышечные ветви к глубокой поперечной мышце промежности и сфинктеру мочеиспускательного канала.
КОПЧИКОВОЕ СПЛЕТЕНИЕ
Копчиковое сплетение, plexus coccygeus, образуется в результате слияния передних ветвей пятого крестцового и копчикового нервов. От сплетения отходят тонкие заднепроходно-копчиковые не- рвы и копчиковый нерв.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
1. Как образуются спинномозговые нервы?
2. Какие вы знаете задние ветви спинномозговых нервов?
3. Какие вам известны чувствительные ветви шейного сплетения?
4. Перечислите короткие ветви плечевого сплетения.
5. Что иннервирует локтевой нерв на кисти?
6. Что иннервирует бедренный нерв? От какого сплетения он отходит?
7. Какие нервы иннервируют мышцы голени по группам?
8. Какие нервы отходят от крестцового сплетения?
ЧЕРЕПНЫЕ НЕРВЫ
Cуществует 13 пар черепных нервов: 0 пара - концевой нерв, n. terminalis; I- обонятельный, п. оlfactorius; II - зрительный, п. opticus; III - глазодвигательный, п. oculomotorius; IV - блоковый, п. trochlearis; V - тройничный, п. trigeminus; VI- отводящий, п. abducens; VII - лицевой, п. facialis; Vjjj - преддверно-улитковый, п. vestibulocochlearis; IX - языкоглоточный, п. glossofaryngeus; X - блуждающий, п. vagus; XI - добавочный, п. accessorius; XII - подъязычный, п. hypoglossus.
РАЗВИТИЕ И ПРИНЦИПЫ СТРОЕНИЯ ЧЕРЕПНЫХ НЕРВОВ
Обонятельный и зрительный нервы - специфические нервы органов чувств, развиваются из переднего мозга и являются его выростами. Остальные черепные нервы дифференцировались из спинномозговых и поэтому по строению принципиально сходны с ними. Дифференцировка и превращение первичных спинномозговых нервов в черепные связаны с развитием органов чувств и жаберных дуг с принадлежащей им мускулатурой, а также с редукцией миотомов в области головы (рис. 227). Однако ни один из черепных нервов не соответствует полностью спинномозговым, так как слагается не из передних и задних корешков, а только из одного переднего или заднего. Черепные нервы jjj, jV, Vj соответствуют передним корешкам. Их ядра расположены вентрально, они иннервируют мышцы, развившиеся из 3 передних сомитов головы. Остальные передние корешки редуцируются.
Другие черепные нервы: V, Vjj, Vjjj, X, Xj и Xjj можно рассматривать как гомологи задних корешков. Эти нервы связаны с мышцами, дифференцировавшимися из мускулатуры жаберного аппарата и развившимися из боковых пластинок мезодермы. Нервы формируют две ветви - переднюю и заднюю. У высших позвоночных задняя ветвь обычно редуцирована.
Рис. 227. Черепные нервы человеческого эмбриона.
Жаберные дуги обозначены арабскими цифрами, нервы - римскими.
Некоторые черепные нервы (X, Xjj) имеют сложное происхождение, так как образуются путем слияния нескольких спинномоз- говых нервов. В связи с ассимиляцией метамеров туловища затылочной областью головы часть спинномозговых нервов перемещается краниально и заходит в область продолговатого мозга. Черепные нервы jX и Xj развились из общего источника - первичного блуждающего нерва и являются как бы его ветвями (табл. 14).
Таблица 14. Соотношение сомитов головы, жаберных дуг и черепных нервов с их корешками
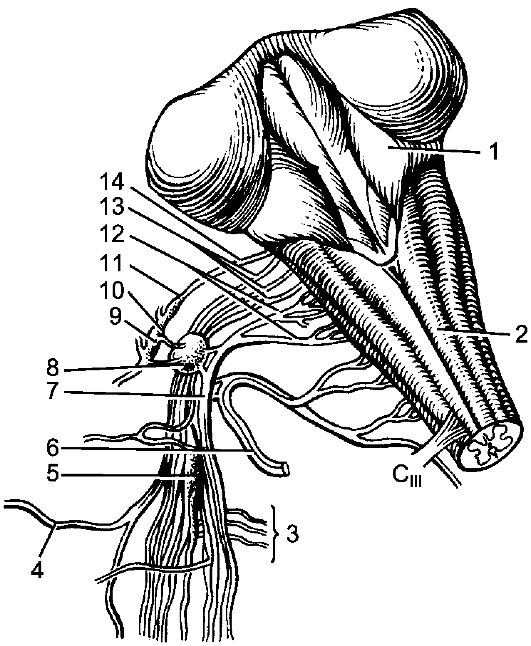
Рис. 228. Cвязи IX, Х и XI пар черепных нервов.
1 - ромбовидная ямка; 2 - спинной мозг; 3 - ветви от нижнего узла блуждающего нерва к симпатическому стволу; 4 - верхний гортанный нерв; 5 - нижний узел блуждающего нерва; 6 - наружная ветвь добавочного нерва; 7 - внутренняя ветвь добавочного нерва; 8 - верхний узел блуждающего нерва; 9 - нижний узел языкоглоточного нерва; 10 - длинная ветвь блуждающего нерва; 11 - верхний узел языкоглоточного нерва; 12 - черепные корешки добавочного нерва; 13 - блуждающий нерв; 14 - языкоглоточный нерв.
Черепные нервы по функциональной принадлежности распределяются следующим образом. К соматически-чувствительным нервам принадлежат I, II, VIII пары, к соматически-двигательным - III, IV, VI, XI, XII пары, к смешанным, содержащим соматически-двигательные и висцерально-чувствительные волокна (VII, IX, Х пары), а также висцерально-двигательные волокна, - V, VII, IX, Х пары.
C ромбовидным мозгом связаны V, VI, VII, VIII, IX, X, XI, XII пары; со средним мозгом - jjj и jV пары; с промежуточным - j и II пары черепных нервов (рис. 228).
0 ПАРА - КОНЦЕВЫЕ НЕРВЫ
Концевой нерв (0 пара), n. terminalis - это пара маленьких нервов, которые тесно прилежат к обонятельным нервам. Впервые они были
обнаружены у низших позвоночных, но их присутствие показано у плодов человека и у взрослых людей. Они содержат множество безмиелиновых волокон и связанные с ними небольшие группы биполярных и мультиполярных нервных клеток. Каждый нерв проходит вдоль меди- альной стороны обонятельного тракта, их ветви прободают решетчатую пластинку решетчатой кости и разветвляются в слизистой оболочке полости носа. Центрально нерв связан с мозгом вблизи переднего продырявленного пространства и септальной области. Его функция неизвестна, предположительно, он представляет собой головную часть симпатической нервной системы, которая распространяется на кровеносные сосуды и железы слизистой оболочки полости носа.
I ПАРА - ОБОНЯТЕЛЬНЫЕ НЕРВЫ
Обонятельный нерв, п. olfactorius, образован 15-20 обонятельными нитями, fila olfactoria, которые состоят из нервных волокон - отростков обонятельных клеток, располагающихся в слизистой оболочке верхнего отдела носовой полости. Обонятельные нити входят в полость черепа через отверстие в решетчатой пластинке и оканчиваются у обонятельных луковиц, которые продолжаются в обонятельный путь, tractus olfactorius (см. с. 650).
II ПАРА - ЗРИТЕЛЬНЫЕ НЕРВЫ
Зрительный нерв, п. opticus, состоит из нервных волокон, образуемых отростками мультиполярных нервных клеток сетчатой оболочки глазного яблока. Зрительный нерв формируется на заднем полушарии глазного яблока и проходит в глазнице к зрительному каналу, откуда выходит в полость черепа. Здесь в предперекрестной борозде оба зрительных нерва соединяются, образуя зрительный перекрест, chiasma opticum. Продолжение зрительных путей называется зрительным трактом, tractus opticus. В зрительном перекресте медиальная группа нервных волокон каждого нерва переходит в зрительный тракт противоположной стороны, а латеральная продолжается в соответствующий зрительный тракт. Зрительные тракты достигают подкорковых зрительных центров.
III ПАРА - ГЛАЗОДВИГАТЕЛЬНЫЕ НЕРВЫ
Глазодвигательный нерв, п. oculomotorius, в основном двигательный, возникает в двигательном ядре, nucleus п. oculomotorius, сред-
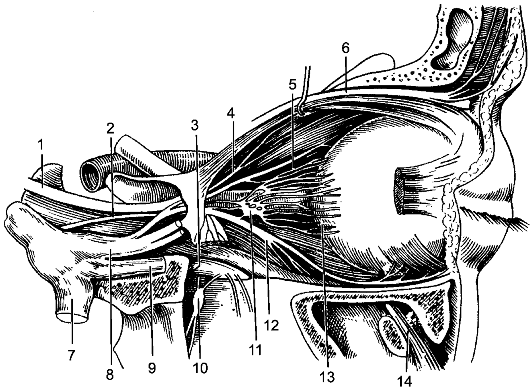
Рис. 229. Нервы глазницы; вид сбоку.
1 - глазодвигательный нерв; 2 - отводящий нерв; 3, 9 - верхнечелюстной нерв; 4 - верхняя ветвь глазодвигательного нерва; 5 - носоресничный нерв; 6 - лобный нерв; 7 - нижнечелюстной нерв; 8 - глазной нерв; 10 - крылонебный узел; 11 - ресничный узел; 12 - нижняя ветвь глазодвигательного нерва; 13 - короткие ресничные нервы; 14 - подглазничный нерв.
него мозга и парасимпатическом добавочном ядре, nucleus accessorius. Выходит на основание мозга у медиального края ножки мозга и идет вперед в верхней стенке пещеристого синуса до верхней глаз- ничной щели, через которую вступает в глазницу и разделяется на верхнюю ветвь, r. superior, - к верхней прямой мышце и мышце, поднимающей веко, и нижнюю ветвь, r. inferior, - к медиальной и нижней прямым и нижней косой мышцам. От нижней ветви отходит ветвь к ресничному узлу, являющаяся его парасимпатическим корешком (рис. 229).
IV ПАРА - БЛОКОВЫЕ НЕРВЫ
Блоковый нерв, п. trochlearis, двигательный, берет начало в двигательном ядре, nucleus n. trochlearis, расположенном в среднем мозге на уровне нижнего двухолмия. Выходит на основание мозга кнаружи от моста и продолжается вперед в наружной стенке пещеристо-
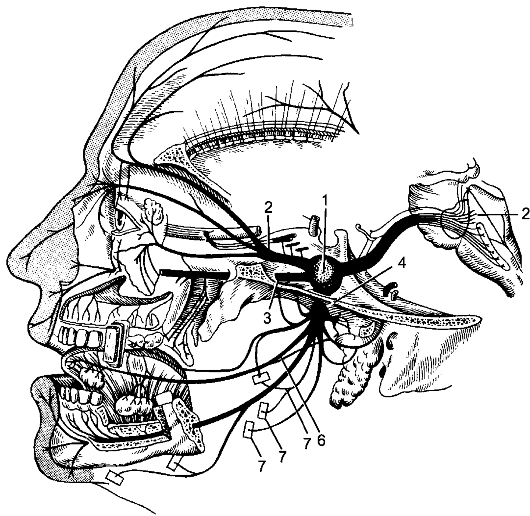
Рис. 230. Тройничный нерв.
1 - тройничный узел; 2 - глазной нерв; 3 - верхнечелюстной нерв; 4 - нижнечелюстной нерв; 5 - дно jV желудочка; 6 - язычный нерв; 7 - нервы к жевательным мышцам.
го синуса. Через верхнюю глазничную щель впадает в глазницу и разветвляется в верхней косой мышце.
V ПАРА - ТРОЙНИЧНЫЕ НЕРВЫ
Тройничный нерв, п. trigeminus, является смешанным и содержит двигательные и чувствительные нервные волокна. Иннервирует жевательные мышцы, кожу лица и переднего отдела головы, твердую оболочку головного мозга, а также слизистые оболочки носовой и ротовой полостей, зубы.
Тройничный нерв имеет сложное строение. В нем различают: 1) ядра (1 двигательное и 3 чувствительных); 2) чувствительный и двигательный корешки; 3) тройничный узел на чувствительном корешке; 4) ствол тройничного нерва; 5) 3 главные ветви тройничного нерва: глазной, верхнечелюстной и нижнечелюстной нервы (рис. 230).
Чувствительные нервные клетки, нейриты которых образуют чувствительные ветви тройничного нерва, расположены в тройнич- ном узле, ganglion trigeminale. Тройничный узел залегает на тройничном вдавлении, impressio trigeminale, передней поверхности пирамиды височной кости в тройничной полости, cavum trigeminale, образованной расщеплением твердой мозговой оболочки. Узел плоский, полулунной формы, длиной 14-29 мм и высотой 5-10 мм. У людей с брахицефалическим черепом он короткий и высокий, а у долихоцефалов - длинный и низкий.
Клетки тройничного узла являются псевдоуниполярными и отдают по одному отростку, который вблизи тела клетки разделяется на 2: центральный и периферический. Центральные нейриты формируют чувствительный корешок, radix sensoria, и через него вступают в мозговой ствол, достигая чувствительных ядер нерва: мостового ядра, nucleus pontinus n. trigemini, в мосту, спинномозгового (нижнего ядра тройничного нерва), nucleus spinalis (inferior) n. trigemini, - в нижней части мозгового моста и в продолговатом мозге, а также ядра средне- мозгового пути, nucleus mesencephalicus n. trigemini, - в среднем мозге. Периферические дендриты идут в составе перечисленных главных ветвей тройничного нерва.
Двигательные нервные волокна берут начало в двигательном ядре нерва, nucleus motorius n. trigemini, лежащем в задней части моста. Указанные волокна выходят из мозга и образуют двигательный корешок, radix motoria. Место выхода двигательного корешка из головного мозга и входа чувствительного расположено при переходе моста в среднюю мозжечковую ножку. Между чувствительными и двигательным корешками тройничного нерва нередко (в 25% случаев) есть анастомотические связи, в результате чего некоторое количество нервных волокон переходит из одного корешка в другой.
Диаметр чувствительного корешка составляет 2,0-2,8 мм, он содержит от 75 000 до 150 000 миелиновых нервных волокон диа- метром преимущественно до 5 мкм. Толщина двигательного корешка меньше - 0,8-1,4 мм. В нем проходит от 6000 до 15 000 миелиновых нервных волокон диаметром, как правило, более 5 мкм.
Чувствительный корешок с принадлежащим ему тройничным узлом и двигательный корешок в совокупности составляют ствол тройничного нерва диаметром 2,3-3,1 мм, содержащий от 80 000 до 165 000 миелиновых нервных волокон. Двигательный корешок
минует тройничный узел и входит в состав нижнечелюстного нерва.
С 3 главными ветвями тройничного нерва связаны парасимпатические нервные узлы: ресничный узел - с глазным нервом, крылонебный - с верхнечелюстным, ушной и поднижнечелюстной узлы - с нижнечелюстными нервами.
Общий план ветвления глазных ветвей тройничного нерва таков: каждый нерв (глазной, верхнечелюстной и нижнечелюстной) отдает ветвь к твердой мозговой оболочке; внутренние ветви к слизистой оболочке добавочных пазух, ротовой и носовой полостей и органам (слезная железа, глазное яблоко, слюнные железы, зубы); наружные ветви: медиальные - к коже передних областей лица и латеральные - к коже боковых областей лица.
ГЛАЗНОЙ НЕРВ
Глазной нерв, п. ophthalmicus, является первой, наиболее тонкой ветвью тройничного нерва. Функционально нерв преимущественно чувствительный. Он иннервирует кожу лба и переднего отдела височной и теменной областей, верхнего века, спинки носа, а также частично слизистую оболочку носовой полости, оболочки глазного яблока и слезную железу (рис. 231).
Нерв имеет в толщину 2-3 мм, состоит из 30-70 сравнительно мелких пучков и содержит от 20 000 до 54 000 миелиновых нервных волокон, преимущественно небольшого диаметра (до 5 мкм). По отхождении от тройничного узла нерв проходит в наружной стенке пещеристого синуса, где отдает тонкие ветви к глазодвигательному, блоковому и отводящему нервам, ветвь намета, r. tentorius, к намету мозжечка и принимает несколько ветвей от внутреннего сонного сплетения. Вблизи верхней глазничной щели глазной нерв делится на 3 ветви: слезный, лобный и носоресничный нервы (рис. 232).
1. Слезный нерв, п. lacrimalis, располагается вблизи наружной стенки глазницы, где соединяется соединительной ветвью со скуловым нервом, r. communicans cum п. zygomatico. Иннервирует слезную железу, а также кожу верхнего века и латерального угла глазной щели.
2. Лобный нерв, п. frontalis, - самая толстая ветвь глазного нерва. Проходит под верхней стенкой глазницы и делится на две ветви: надглазничный нерв, п. supraorbitalis, идущий через надглазничную вырезку к коже лба, и надблоковый нерв, п. supratrochlearis, выходя-
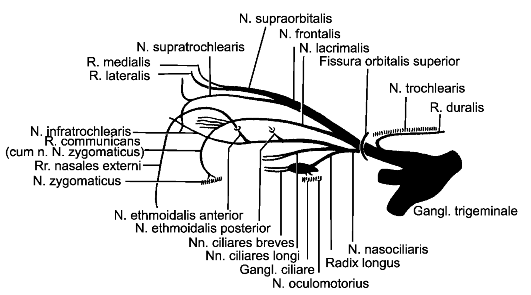
Рис. 231. Глазной нерв.
щий из глазницы у ее внутренней стенки и иннервирующий кожу верхнего века и медиального угла глаза.
3. Носоресничный нерв, п. nasociliaris, лежит в глазнице у медиальной ее стенки и под блоком верхней косой мышцы выходит из глазницы в виде конечной ветви - подблокового нерва, п. infratrochlearis, который иннервирует слезный мешок, конъюнктиву и медиальный угол глаза. На своем протяжении носоресничный нерв отдает следующие ветви: 1) длинные ресничные нервы, пп. ciliares longi, к глазному яблоку; 2) задний решетчатый нерв, п. ethmoidalis posterior, к слизистой оболочке клиновидной пазухи и задних ячеек решетчатого лабиринта; 3) передний решетчатый нерв, п. ethmoidalis anterior, к слизистой оболочке лобной пазухи и носовой полости (rr. nasales medialis et lateralis) и к коже кончика и крыла носа. Кроме того, от носоресничного нерва отходит соединительная ветвь к ресничному узлу.
Ресничный узел, ganglion ciliare (рис. 233), длиной до 2 мм лежит на латеральной поверхности зрительного нерва, приблизительно на границе между задней и средней третями длины глазницы. В ресничном узле, как и в других парасимпатических узлах тройничного нерва, находятся парасимпатические многоотростчатые (мультиполярные) нервные клетки, на которых преганглионарные волокна, образуя синапсы, переключаются на постганглионарные. Симпатические и чувствительные волокна проходят через узел транзитно.
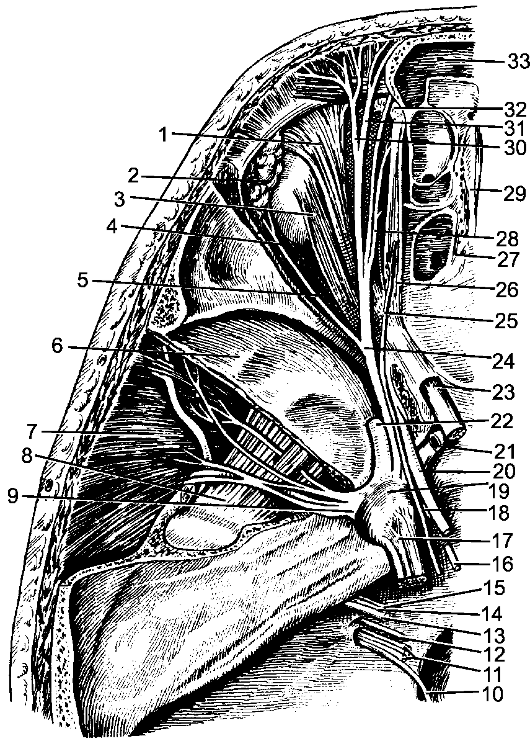
Рис. 232. Нервы глазницы; вид сверху.
1 - мышца, поднимающая верхнее веко; 2 - слезная железа; 3 - верхняя прямая мышца; 4 - слезный нерв; 5 - латеральная прямая мышца; 6 - средняя черепная ямка; 7 - височная мышца; 8 - латеральная крыловидная мышца; 9 - нижнечелюстной нерв; 10 - добавочный нерв; 11 - блуждающий нерв; 12 - языкоглоточный нерв; 13 - улитковая часть преддверно-улиткового нерва; 14 - преддверная часть преддверно-улиткового нерва; 15 - лицевой нерв; 16 - отводящий нерв; 17 - тройничный нерв; 18, 25 - блоковый нерв; 19 - тройничный нерв; 20 - глазодвигательный нерв; 21 - внутренняя сонная артерия; 22 - верхнечелюстной нерв; 23 - зрительный нерв; 24 - глазной нерв; 26 - верхняя косая мышца; 27 - решетчатая пластинка; 28 - носоресничный нерв; 29 - петушиный гребень; 30 - надглазничный нерв; 31 - лобный нерв; 32 - блок; 33 - лобная пазуха.
К узлу подходят соединительные ветви в виде его корешков: 1) чувствительный, radix nasociliaris, - из носоресничного нерва; 2) парасимпатический, radix oculomotoris, - из глазодвигательного нерва; 3) симпатический, radix sympathicus, - из сплетения, окружающего a. ophthalmica. От ресничного узла отходит от 4 до 10 коротких
Рис. 233. Ресничный узел (схема).
1 - белая соединительная ветвь; 2 - верхний шейный узел симпатического ствола; 3 - внутреннее сонное сплетение; 4 - симпатический корешок; 5 - ресничный узел; 6 - носоресничный корешок; 7 - глазодвигательный нерв; 8 - глазодвигательный корешок; 9 - короткие ресничные нервы; 10 - преганглионарные парасимпатические волокна; 11 - постганглионарные парасимпатические волокна; 12 - чувствительные волокна; 13 - постганглионарные симпатические волокна; 14 - симпатическое ядро; 15 - спинномозговой нерв; 16 - преганглионарные симпатические волокна; 17 - двигательные волокна.
ресничных нервов, пп. ciliares breves, идущих внутрь глазного яблока. Они содержат постганглионарные парасимпатические волокна, иннервирующие ресничную мышцу и сфинктер зрачка, чувствительные волокна, иннервирующие оболочки глазного яблока, а также симпатические волокна к мышце, расширяющей зрачок.
ВЕРХНЕЧЕЛЮСТНОЙ НЕРВ
Верхнечелюстной нерв, п. maxillaris, - вторая ветвь тройничного нерва, в основном чувствительный. Имеет толщину 2,5-4,5 мм и состоит из 25-70 небольших пучков, содержащих от 30 000 до 80 000 миелиновых нервных волокон, преимущественно небольшого диаметра (до 5 мкм).
Верхнечелюстной нерв иннервирует твердую оболочку головного мозга, кожу нижнего века, латерального угла глаза, передней части височной области, верхней части щеки, крыльев носа, кожу и слизистую оболочку верхней губы, слизистую оболочку задней и нижней частей носовой полости, слизистую оболочку клиновидной пазухи, неба, зубы верхней челюсти (рис. 234). По выходе из черепа через круглое отверстие нерв попадает в крыловидно-небную ямку, проходит сзади наперед и изнутри кнаружи. Длина отрезка и его положение в ямке зависят от формы черепа. При брахицефалическом черепе длина отрезка нерва в ямке составляет 15-22 мм, он расположен в ямке глубоко - до 5 см от середины скуловой дуги. Иногда нерв в крыловидно-небной ямке закрыт костным гребнем. При долихоцефалическом черепе длина рассматриваемого участка нерва 10-15 мм, а располагается он более поверхностно - до 4 см от середины скуловой дуги.
В пределах крыловидно-небной ямки верхнечелюстной нерв отдает менингеальную ветвь, r. meningeus, к твердой мозговой оболочке и разделяется на 3 ветви: 1) узловые ветви, идущие к крылонебному узлу, 2) скуловой нерв и 3) подглазничный нерв, являющийся непосредственным продолжением верхнечелюстного нерва.
1. Узловые ветви, rr. ganglionares, числом 1-7, отходят от верхнечелюстного нерва на расстоянии 1,0-2,5 мм от круглого отверстия и идут к крылонебному узлу, отдавая нервам, начинающимся от узла, чувствительные волокна. Некоторые узловые ветви минуют узел и присоединяются к его ветвям.
Крылонебный узел, ganglion pterygopalatinum, - образование парасимпатической части автономной нервной системы. Узел треугольной формы, длиной 3-5 мм, содержит мультиполярные клетки
Рис. 234. Верхнечелюстной нерв.
и имеет 3 корешка: 1) чувствительный - узловые ветви, пп. pterygopalatini; 2) парасимпатический - большой каменистый нерв, п. petrosus major (ветвь промежуточного нерва), содержит волокна к железам носовой полости, неба, слезной железе; 3) симпатический - глубокий каменистый нерв, п. petrosus profundus, отходит из внутреннего сонного сплетения, содержит постганглионарные симпатические нервные волокна из шейных узлов. От узла отходят ветви, в составе которых есть секреторные (парасимпатические и симпатические) и чувствительные волокна (рис. 235):
1) глазничные ветви, rr. orbitales, 2-3 тонких ствола, проникают через нижнюю глазничную щель и далее вместе с задним решетчатым нервом идут через мелкие отверстия клиновидно-решетчатого шва к слизистой оболочке задних ячеек решетчатого лабиринта и клиновидной пазухи;
2) задние верхние носовые ветви, rr. nasales posteriores superiores, числом 8-14 выходят из крыловидно-небной ямки через клиновидно-небное отверстие в носовую полость и делятся на две группы: латеральную и медиальную. Латеральные ветви, rr. nasales posteriores superiores laterales (6-10), идут к слизистой оболочке задних отделов верхней и средней носовых раковин и носовых ходов, задних ячеек решетчатой кости, верхней поверхности хоан и глоточного отверстия слуховой трубы. Медиальные ветви, rr. nasales posteriores superiores mediales (2-3), разветвляются в слизистой оболочке верхнего отдела
Рис. 235. Крылонебный узел (схема).
1 - верхнечелюстной нерв; 2 - преганглионарные парасимпатические волокна большого каменистого нерва; 3 - постганглионарные симпатические волокна глубокого каменистого нерва; 4 - небные нервы; 5 - постганглионарные парасимпатические волокна; 6 - скуловой нерв; 7 - соединительная ветвь скулового и слезного нервов; 8 - слезный нерв; 9 - слезная железа; 10 - нерв крыловидного канала; 11 - узловые ветви верхнечелюстного нерва; 12 - задние носовые нервы; 13 - чувствительные волокна.
перегородки носа. Одна из медиальных ветвей - носонебный нерв,
п. nasopalatinus, проходит между надкостницей и слизистой оболочкой перегородки вместе с задней артерией перегородки носа вперед, к носовому отверстию резцового канала, через который достигает слизистой оболочки передней части неба. Образует соединение с носовой ветвью верхнего альвеолярного нерва (рис. 236).
Небные нервы, пп. palatini, распространяются от узла через большой небный канал, образуя 3 группы нервов:
1) большой небный нерв, п. palatinus major, - самая толстая ветвь, выходит через большое небное отверстие на небо, где распадается на 3-4 ветви, иннервирующие большую часть слизистой оболочки неба и ее железы на участке от клыков до мягкого неба;
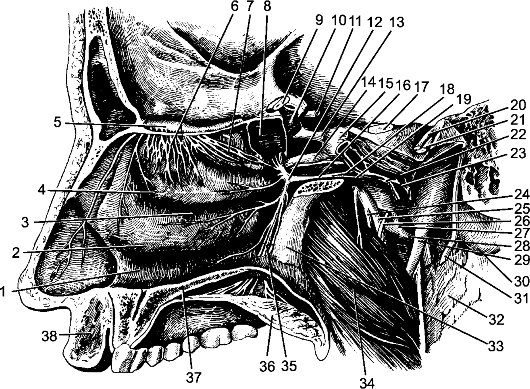
Рис. 236. Обонятельный нерв, крылонебный узел и ветви тройничного нерва. 1 - нижний носовой ход; 2, 4, 7 - нижняя, средняя и верхняя носовые раковины соответственно; 3 - средний носовой ход; 5 - обонятельная луковица; 6 - обонятельные нервы; 8 - клиновидная пазуха; 9 - зрительный нерв; 10, 23 - внутренняя сонная артерия; 11 - глазодвигательный нерв; 12 - крылонебный узел; 13 - глазной нерв; 14 - верхнечелюстной нерв; 15 - тройничный узел; 16 - нерв крыловидного канала; 17 - тройничный нерв; 18 - большой каменистый нерв; 19 - глубокий каменистый нерв; 20, 31 - лицевой нерв; 21 - преддверно-улитковый нерв; 22 - внутреннее сонное нервное сплетение; 24 - язычный нерв; 25 - ниж- ний альвеолярный нерв; 26 - барабанная струна; 27 - средняя менингеальная артерия; 28 - верхнечелюстная артерия; 29 - шиловидный отросток; 30 - сосце- видный отросток; 32 - околоушная слюнная железа; 33 - перпендикулярная пла- стинка небной кости; 34 - медиальная крыловидная мышца; 35 - небные нервы; 36 - мягкое небо; 37 - твердое небо; 38 - верхняя губа.
2) малые небные нервы, пп. palatini minores, вступают в полость рта через малые небные отверстия и ветвятся в слизистой оболочке мягкого неба и области небной миндалины;
3) нижние задние носовые ветви, rr. nasales posteriores inferiores, входят в большой небный канал, покидают его через мелкие отверстия и на уровне нижней носовой раковины вступают в носовую полость, иннервируя слизистую оболочку нижней раковины, среднего и нижнего носовых ходов и верхнечелюстной пазухи.
2. Скуловой нерв, п. zygomaticus, ответвляется от верхнечелюстного нерва в пределах крыловидно-небной ямки и проникает через нижнюю глазничную щель в глазницу, где идет по наружной стенке, выходит через скулоглазничное отверстие и делится на две ветви:
1) скулолицевая ветвь, r. zygomaticofacialis, выходит через скулолицевое отверстие на переднюю поверхность скуловой кости, в коже верхней части щеки отдает ветвь к области наружного угла глазной щели и соединительную ветвь к лицевому нерву;
2) скуловисочная ветвь, r. zygomaticotemporalis, выходит из глазницы через одноименное отверстие скуловой кости, прободает височную мышцу и ее фасцию и иннервирует кожу передней части височной и задней части лобной областей. Отдает соединительную ветвь к слезному нерву, содержащую секреторные парасимпатические волокна к слезной железе.
3. Подглазничный нерв, п. infraorbitalis, является продолжением верхнечелюстного нерва и получает свое название по отхождении от последнего указанных ранее ветвей. Подглазничный нерв покидает крыловидно-небную ямку через нижнюю глазничную щель, проходит по нижней стенке глазницы вместе с одноименными со- судами в подглазничной борозде (в 15% случаев вместо борозды имеется костный канал) и выходит через подглазничное отверстие под мышцу, поднимающую верхнюю губу, разделяясь на конечные ветви. Длина подглазничного нерва различна: у брахицефалов ствол нерва составляет 20-27 мм, а у долихоцефалов - 27-32 мм. Положение нерва в глазнице соответствует парасагиттальной плоскости, проведенной через подглазничное отверстие.
Характер отхождения ветвей может быть также различным: рассыпным, при котором от ствола отходят многочисленные тонкие нервы с большим количеством связей, или магистральным с небольшим числом крупных нервов. На своем пути подглазничный нерв отдает следующие ветви:
1) верхние альвеолярные нервы, пп. alveolares superiores, иннервируют зубы и верхнюю челюсть (рис. 237). Различают 3 группы ветвей верхних альвеолярных нервов:
а) задние верхние альвеолярные ветви, rr. alveolares superiores posteriores, ответвляются от подглазничного нерва, как правило, в крыловидно-небной ямке числом 4-8 и располагаются вместе с одноименными сосудами по поверхности бугра верхней челюсти. Часть самых задних нервов идет по наружной поверхности бугра вниз, к альвеолярному отростку, остальные входят через задние
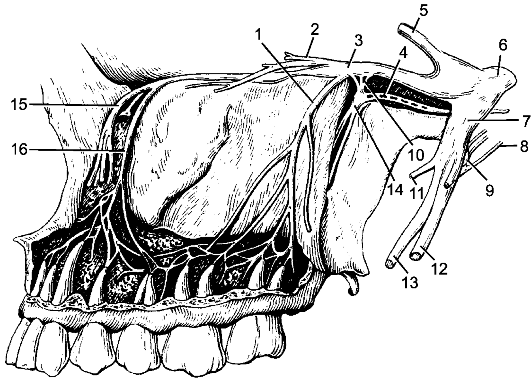
Рис. 237. Верхнечелюстной нерв.
1 - задние верхние альвеолярные ветви; 2 - скуловой нерв; 3 - верхнечелюстной нерв; 4 - нерв крыловидного канала; 5 - глазной нерв; 6 - тройничный нерв; 7 - нижнечелюстной нерв; 8 - барабанная струна; 9 - ушной узел; 10 - соединительные ветви крылонебного узла с верхнечелюстным нервом; 11 - жевательный нерв; 12 - нижний альвеолярный нерв; 13 - язычный нерв; 14 - крылонебный узел; 15 - подглазничный нерв; 16 - передние верхние альвеолярные ветви.
верхние альвеолярные отверстия в альвеолярные каналы. Ветвясь вместе с другими верхними альвеолярными ветвями, они формиру- ют нервное верхнее зубное сплетение, plexus dentalis superior, которое залегает в альвеолярном отростке верхней челюсти над верхушками корней. Сплетение достаточно густое, широкопетлистое, растянуто по всей длине альвеолярного отростка. От сплетения отходят верхние десневые ветви, rr. gingivales superiores, к периодонту и пародонту на участке верхних моляров, и верхние зубные ветви, rr. dentales superiores, к верхушкам корней больших коренных зубов, в пульпарной полости которых они разветвляются. Кроме того, задние верхние альвеолярные ветви посылают тонкие нервы к слизистой оболочке верхнечелюстной пазухи;
б) средняя верхняя альвеолярная ветвь, r. alveolaris superior medius, в виде одного или, реже, двух стволиков ответвляется от подглазничного нерва, чаще в крыловидно-небной ямке и реже в пределах глазницы, проходит в одном из альвеолярных каналов и ветвится в
костных канальцах верхней челюсти в составе верхнего зубного сплетения. Имеет соединительные ветви с задними и передними верхними альвеолярными ветвями. Иннервирует посредством верхних десневых ветвей пародонт и периодонт на участке верхних премоляров, посредством верхних зубных ветвей - верхние премоляры;
в) передние верхние альвеолярные ветви, rr. alveolares superiores anteriores, возникают от подглазничного нерва в передней части глазницы, которую покидают через альвеолярные каналы, проникая в переднюю стенку верхнечелюстной пазухи, где входят в состав верхнего зубного сплетения. Верхние десневые ветви иннервируют слизистую оболочку альвеолярного отростка и стенки альвеол на участке верхних клыков и резцов, верхние зубные ветви - верхние клыки и резцы. Передние верхние альвеолярные ветви посылают тонкую носовую ветвь к слизистой оболочке переднего отдела дна носовой полости;
2) нижние ветви век, rr. palpebrales inferiores, ответвляются от подглазничного нерва по выходе из подглазничного отверстия, проникают через мышцу, поднимающую верхнюю губу, и, разветвляясь, иннервируют кожу нижнего века;
3) наружные носовые ветви, rr. nasales externi, иннервируют кожу в области крыла носа;
4) внутренние носовые ветви, rr. nasales interni, подходят к слизистой оболочке преддверия полости носа;
5) верхние губные ветви, rr. labiales superiores, числом 3-4, идут между верхней челюстью и мышцей, поднимающей верхнюю губу, вниз; иннервируют кожу и слизистую оболочку верхней губы до угла рта.
Все перечисленные наружные ветви подглазничного нерва образуют связи с ветвями лицевого нерва.
НИЖНЕЧЕЛЮСТНОЙ НЕРВ
Нижнечелюстной нерв, п. mandibularis, - третья ветвь тройничного нерва, является смешанным нервом и формируется за счет чувствительных нервных волокон, идущих из тройничного узла, и двигательных волокон двигательного корешка. Толщина ствола нерва колеблется от 3,5 до 7,5 мм, а длина внечерепного отдела ствола 0,5-2,0 см. Нерв состоит из 30-80 пучков волокон, включающих от 50 000 до 120 000 миелиновых нервных волокон.
Нижнечелюстной нерв осуществляет чувствительную иннервацию твердой оболочки головного мозга, кожи нижней губы, подбородка, нижней части щеки, передней части ушной раковины и наружного слухового прохода, части поверхности барабанной перепонки, слизистой оболочки щеки, дна полости рта и передних двух третей языка, зубов нижней челюсти, а также двигательную иннервацию всех жевательных мышц, челюстно-подъязычной мышцы, переднего брюшка двубрюшной мышцы и мышц, напрягающих барабанную перепонку и небную занавеску.
Из полости черепа нижнечелюстной нерв выходит через овальное отверстие и попадает в подвисочную ямку, где разделяется вблизи места выхода на ряд ветвей. Ветвление нижнечелюстного нерва возможно или по рассыпному типу (чаще у долихоцефалов) - нерв распадается на множество ветвей (8-11), или по магистральному типу (чаще у брахицефалов) с ветвлением на небольшое число стволов (4-5), которые являются общими для нескольких нервов.
С ветвями нижнечелюстного нерва связаны три узла вегетативной нервной системы: ушной, ganglion oticum; поднижнечелюстной, ganglion submandibulare; подъязычный, ganglion sublinguale. От узлов идут постганглионарные парасимпатические секреторные волокна к слюнным железам.
Нижнечелюстной нерв отдает ряд ветвей (рис. 238, 239).
1. Менингеальная ветвь, r. meningeus, проходит через остистое отверстие вместе со средней менингеальной артерией в полость черепа, где разветвляется в твердой оболочке.
2. Жевательный нерв, п. massetericus, преимущественно двигательный, часто (особенно при магистральной форме ветвления нижнечелюстного нерва) имеет общее начало с другими нервами жевательных мышц. Проходит кнаружи над верхним краем латеральной крыловидной мышцы через вырезку нижней челюсти и внедряется в жевательную мышцу. Перед входом в мышцу посылает тонкую ветвь к височно-нижнечелюстному суставу, обеспечивая его чувствительную иннервацию.
3. Глубокие височные нервы, пп. temporales profundi, двигательные, проходят по наружному основанию черепа кнаружи, огибают подвисочный гребень и входят в височную мышцу с внутренней ее поверхности в переднем (п. temporalis profundus anterior) и заднем (п. temporalis profundus posterior) отделах.
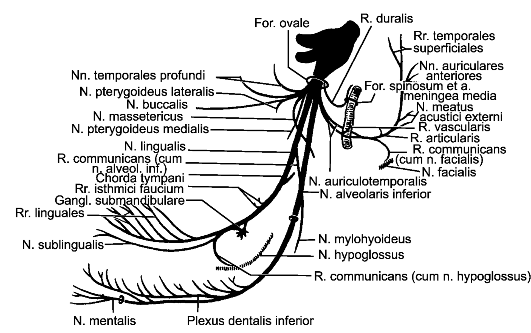
Рис. 238. Строение нижнечелюстного нерва.
3. Латеральный крыловидный нерв, п. pterygoideus lateralis, двигательный, обычно отходит общим стволом со щечным нервом, подходит к одноименной мышце, в которой разветвляется.
4. Медиальный крыловидный нерв, п. pterygoideus medialis, главным образом двигательный. Проходит через ушной узел или прилегает к его поверхности и следует вперед и вниз к внутренней поверхности одноименной мышцы, в которую проникает вблизи ее верхнего края. Кроме того, около ушного узла отдает п. tensoris tympani, n. tensoris veli palatini и соединительную ветвь к узлу.
5. Щечный нерв, п. buccalis, чувствительный, проникает между двумя головками латеральной крыловидной мышцы и идет по внутренней поверхности височной мышцы, распространяясь далее вместе со щечными сосудами по наружной поверхности щечной мышцы до угла рта. На своем пути отдает тонкие ветви, прободающие щечную мышцу и иннервирующие слизистую оболочку щеки (до десны 2-го премоляра и 1-го моляра) и ветви к коже щеки и угла рта. Образует соединительную ветвь с ветвью лицевого нерва и с ушным узлом.
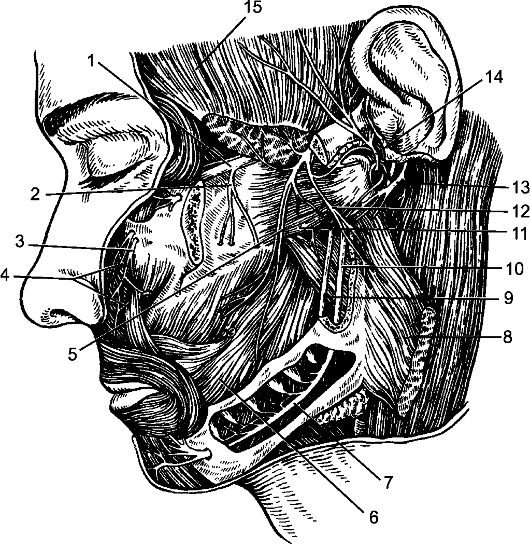
Рис. 239. Нижнечелюстной нерв.
1 - верхнечелюстной нерв; 2 - верхний альвеолярный нерв; 3, 4 - подглазничный нерв; 5 - щечный нерв; 6 - щечная мышца: 7, 10 - нижний альвеолярный нерв; 8 - жевательная мышца (отсечена и отвернута); 9 - язычный нерв; 11 - латеральная крыловидная мышца; 12 - жевательный нерв; 13 - лицевой нерв; 14 - ушновисочный нерв; 15 - височная мышца.
6. Ушно-височный нерв, п. auriculotemporalis, чувствительный, начинается от задней поверхности нижнечелюстного нерва двумя корешками, охватывающими среднюю менингеальную артерию, которые затем соединяются в общий ствол. Отдает соединительную ветвь к ушному узлу. Около шейки суставного отростка нижней челюсти ушно-височный нерв идет кверху и через околоушную слюнную железу выходит в височную область, где разветвляется на конечные ветви - поверхностные височные, r. temporales superficiales. На своем пути ушно-височный нерв отдает следующие ветви:
1) суставные, r. articulares, к височно-нижнечелюстному суставу;
2) околоушные, r. parotidei, к околоушной слюнной железе. Эти ветви содержат, кроме чувствительных, парасимпатические секреторные волокна из ушного узла;
3) нерв наружного слухового прохода, п. meatus acustici externi, к коже наружного слухового прохода и барабанной перепонки;
4) передние ушные нервы, п. auriculares anteriores, к коже переднего отдела ушной раковины и средней части височной области.
7. Язычный нерв, п. lingualis, чувствительный. Берет начало от нижнечелюстного нерва вблизи овального отверстия и располагается между крыловидными мышцами кпереди от нижнего альвеолярного нерва. У верхнего края медиальной крыловидной мышцы или несколько ниже к нерву присоединяется барабанная струна, chorda tympani, которая является продолжением промежуточного нерва. В составе барабанной струны в язычный нерв включаются секреторные волокна, следующие к поднижнечелюстному и подъязычному нервным узлам, и вкусовые волокна к сосочкам языка. Далее язычный нерв проходит между внутренней поверхностью нижней челюсти и медиальной крыловидной мышцей, над поднижнечелю- стной слюнной железой по наружной поверхности подъязычноязычной мышцы к боковой поверхности языка. Между подъязычно-язычной и подбородочно-язычной мышцами нерв рассыпается на конечные язычные ветви, rr. linguales.
По ходу нерва формируются соединительные ветви с подъязычным нервом и барабанной струной. В полости рта язычный нерв отдает следующие ветви:
1) ветви перешейка зева, rr. isthmi faucium, иннервирующие слизистую оболочку зева и заднего отдела дна полости рта;
2) подъязычный нерв, п. sublingualis, отходит от язычного нерва у заднего края подъязычного узла в виде тонкой соединительной ветви и распространяется вперед по боковой поверхности подъязычной слюнной железы. Иннервирует слизистую оболочку дна полости рта, десну и подъязычную слюнную железу;
3) язычные ветви, rr. linguales, проходят вместе с глубокими артерией и венами языка через мускулатуру языка вперед и оканчиваются в слизистой оболочке верхушки языка и его тела по пограничной линии. В составе язычных ветвей проходят вкусовые волокна к сосочкам языка, переходящие из барабанной струны.
8. Нижний альвеолярный нерв, п. alveolaris inferior, смешанный. Это наиболее крупная ветвь нижнечелюстного нерва. Ствол лежит между крыловидными мышцами позади и латеральнее язычного нерва, между нижней челюстью и клиновидно-нижнечелюстной связкой. Нерв входит вместе с одноименными сосудами в нижнечелюстной канал, где отдает множественные ветви, анастомозирующие между собой и образующие нижнее зубное сплетение, plexus dentalis inferior (в 15% случаев), или непосредственно нижние зуб-
ные и десневые ветви. Покидает канал через подбородочное отверстие, разделяясь перед выходом на подбородочный нерв и резцовую ветвь. Отдает следующие ветви:
1) челюстно-подъязычный нерв, и. mylohyoideus, возникает вблизи входа нижнего альвеолярного нерва в нижнечелюстное отверстие, располагается в одноименной борозде ветви нижней челюсти и идет к челюстноподъязычной мышце и переднему брюшку двубрюшной мышцы;
2) нижние зубные и десневые ветви, rr. dentales et gingivales inferiores, берут начало от нижнего альвеолярного нерва в нижнечелюстном канале; иннервируют десну, альвеолы альвеолярной части челюсти и зубы (премоляры и моляры);
3) подбородочный нерв, п. mentalis, является продолжением ствола нижнего альвеолярного нерва по выходе через подбородочное отверстие из канала нижней челюсти, где нерв разделяется веерообразно на 4-8 ветвей, среди которых различают: а) подбородочные, rr. mentales, к коже подбородка; б) нижние губные, rr. labiates inferiores, к коже и слизистой оболочке нижней губы.
Ушной узел, ganglion oticum, - округлое тело диаметром 3-5 мм; располагается под овальным отверстием на заднемедиальной поверхности нижнечелюстного нерва. К нему подходит малый каменистый нерв (от языкоглоточного), приносящий преганглионарные па- расимпатические волокна. От узла отходит ряд соединительных ветвей: 1) к ушно-височному нерву, в который поступают постганглионарные парасимпатические секреторные волокна, идущие затем в составе околоушных ветвей к околоушной слюнной железе; 2) к менингеальной ветви, содержащие волокна, снабжающие сосуды твердой оболочки головного мозга; 3) к барабанной струне; 4) к крылонебному и тройничному узлам (рис. 240).
Поднижнечелюстной узел, ganglion submandibulare, размером 3,0- 3,5 мм, располагается под стволом язычного нерва и связан с ним узловыми ветвями, rr. ganglionares. По этим ветвям идут к узлу и оканчиваются в нем волокна барабанной струны. Отходящие из узла постганглионарные ветви иннервируют поднижнечелюстную и подъязычную слюнные железы (см. рис. 235).
Иногда (до 30% случаев) встречается отдельный подъязычный узел, ganglion sublinguale.
VI ПАРА - ОТВОДЯЩИЕ НЕРВЫ
Отводящий нерв, n. abducens - двигательный. Ядро отводящего нерва, nucleus n. abducentis, располагается в передней части дна IV желудочка. Нерв выходит из мозга у заднего
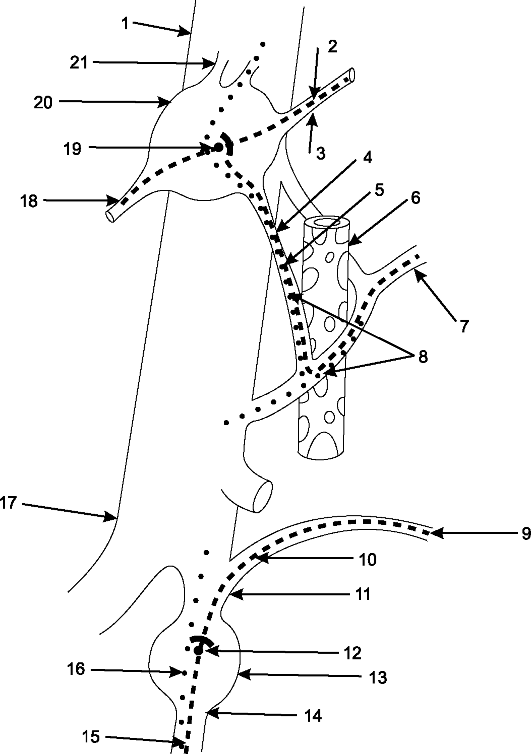
Рис. 240. Ушной и поднижнечелюстной узлы (схема).
1 - нижнечелюстной нерв; 2, 10 - преганглионарные парасимпатические волокна; 3 - малый каменистый нерв; 4 - соединительная ветвь ушного узла к ушно-височному нерву; 5, 15 - постганглионарные парасимпатические волокна; 6 - средняя артерия твердой мозговой оболочки; 7 - ушно-височный нерв; 8, 16 - чувствительные волокна; 9 - барабанная струна; 11 - узловые ветви язычного нерва; 12, 19 - переключение преганглионарных волокон на постганглионарные; 13 - поднижнечелюстной нерв; 14 - железистые ветви; 17 - язычный нерв; 18 - соединительная ветвь ушного узла к щечному нерву; 20 - ушной узел; 21 - узловые ветви нижнечелюстного нерва.
Рис. 241. Лицевой нерв (схема).
1 - дно IV желудочка; 2 - ядро лицевого нерва; 3 - шилососцевидное отверстие; 4 - задняя ушная мышца; 5 - затылочная вена; 6 - заднее брюшко двубрюшной мышцы; 7 - шилоподъязычная мышца; 8 - ветви лицевого нерва к мимической мускулатуре и платизме; 9 - мышца, опускающая угол рта; 10 - подбородочная мышца; 11 - мышца, опускающая нижнюю губу; 12 - щечная мышца; 13 - круговая мышца рта; 14 - мышца, поднимающая верхнюю губу; 15 - клыковая мышца; 16 - скуловая мышца; 17 - круговая мышца глаза; 18 - мышца, сморщивающая бровь; 19 - лобная мышца; 20 - барабанная струна; 21 - язычный нерв; 22 - крылонебный узел; 23 - тройничный узел; 24 - внутренняя сонная артерия; 25 - промежуточный нерв; 26 - лицевой нерв; 27 - преддверно-улитковый нерв.
края моста, между ним и пирамидой продолговатого мозга, и вскоре снаружи от спинки турецкого седла входит в пещеристый синус, где располагается по наружной поверхности внутренней сонной артерии. Далее проникает через верхнюю глазничную щель в глазницу и следует вперед над глазодвигательным нервом. В пещеристом синусе к нерву проходят соединительные ветви от внутреннего сонного сплетения, содержащие симпатические нервные волокна. Иннервирует наружную прямую мышцу глаза.
VII ПАРА - ЛИЦЕВЫЕ НЕРВЫ
Лицевой нерв, п. facialis, развивается в связи с образованиями второй жаберной дуги (рис. 241), поэтому он иннервирует все мимичес-
1 2 3 4 5 6 7 8
Рис. 242. Нервы каналов височной кости.
1 - стременной нерв; 2 - барабанная струна; 3 - барабанное сплетение; 4 - соединительная ветвь лицевого нерва с барабанным сплетением; 5 - узел коленца; 6 - лицевой нерв; 7 - промежуточный нерв; 8 - преддверно-улитковый нерв; 9, 19 - соединительная ветвь от узла коленца к сплетению средней менингеальной артерии; 10 - большой каменистый нерв; 11 - сонно-барабанный нерв; 12 - малый каменистый нерв; 13 - внутреннее сонное нервное сплетение; 14 - глубокий каменистый нерв; 15 - нерв крыловидного канала; 16 - крылонебные нервы; 17 - верхнечелюстной нерв; 18 - крылонебный узел; 20 - нервное сплетение вокруг средней менингеальной артерии; 21 - ушной узел; 22 - ветви ушного узла к ушно-височному нерву; 23 - соединительная ветвь между ушным узлом и барабанной струной; 24 - жевательный нерв; 25 - нижнечелюстной нерв; 26 - язычный нерв; 27 - нижний альвеолярный нерв; 28 - ушно-височный нерв; 29 - барабанный нерв; 30 - языкоглоточный нерв; 31 - верхний узел блуждающего нерва; 32 - ушная ветвь блуждающего нерва; 33 - соединительная ветвь лицевого нерва с ушной ветвью блуждающего нерва; 34 - ветви лицевого нерва к шилоподъязычной мышце; 35 - ветви лицевого нерва к заднему брюшку двубрюшной мышцы; 36 - задний ушной нерв; 37 - сосцевидный отросток.
кие мышцы. Нерв смешанный, включающий двигательные волокна из своего эфферентного мозгового ядра, а также чувствительные и вегетативные (вкусовые и секреторные) волокна, принадлежащие тесно связанному с лицевым промежуточному нерву, п. intermedius.
Двигательное ядро лицевого нерва, nucleus nervi facialis, находится на дне IV желудочка, в латеральной области ретикулярной формации. Корешок лицевого нерва выходит из мозга вместе с корешком промежуточного нерва впереди от преддверно-улиткового нерва, между задним краем моста и оливой продолговатого мозга. Далее лицевой и про- межуточный нервы входят во внутреннее слуховое отверстие и вступают в лицевой канал. В лицевом канале оба нерва образуют общий ствол, делающий два поворота соответственно изгибам канала (рис. 242).
Сначала общий ствол располагается горизонтально, направляясь над барабанной полостью кпереди и латерально. Затем соответственно изгибу лицевого канала ствол поворачивает под прямым углом назад, образуя коленце, geniculum n. facialis, и узел коленца, ganglion geniculi, принадлежащий промежуточному нерву. Пройдя над барабанной полостью, ствол делает второй поворот вниз, располагаясь позади полости среднего уха. На этом участке от общего ствола отходят ветви промежуточного нерва, лицевой нерв выходит из канала через шилососцевидное отверстие и вскоре вступает в околоушную слюнную железу. Длина ствола внечерепного отдела лицевого нерва колеблется от 0,8 до 2,3 см (чаще 1,5 см), а толщина - от 0,7 до 1,4 мм; нерв содержит 3500-9500 миелиновых нервных волокон, среди которых преобладают толстые.
В околоушной слюнной железе, на глубине 0,5-1,0 см от ее наружной поверхности, происходит разделение лицевого нерва на 2- 5 первичных ветвей, которые делятся на вторичные, образуя околоушное сплетение, plexus intraparotideus.
Различают две формы внешнего строения околоушного сплетения - сетевидную и магистральную. При сетевидной форме ствол нерва короткий (0,8-1,5 см), в толще железы разделяется на множество ветвей, имеющих между собой множественные связи, вследствие чего формируется узкопетлистое сплетение. Наблюдаются множественные связи с ветвями тройничного нерва. При магистральной (форме ствол нерва относительно длинный (1,5-2,3 см), разделяется на две ветви (верхнюю и нижнюю), которые дают по несколько вторичных ветвей; связей между вторичными ветвями мало, сплетение широкопетлистое (рис. 243).
На своем пути лицевой нерв отдает ветви при прохождении по каналу, а также по выходе из него. Внутри канала от него отходит ряд ветвей.
1. Большой каменистый нерв, п. petrosus major, берет начало вблизи узла коленца, покидает канал лицевого нерва через расщелину канала большого каменистого нерва и проходит по одноименной борозде до рваного отверстия. Проникнув через хрящ на наружное основание черепа, нерв соединяется с глубоким каменистым нервом, формируя нерв крыловидного канала, п. canalis pterygoidei, вступающий в крыловидный канал и достигающий крылонебного узла.
Нерв содержит парасимпатические волокна к крылонебному узлу, а также чувствительные волокна от клеток узла коленца. Часть чувствительных волокон в большом каменистом нерве идет из крылонебного узла в составе лицевого нерва.
Рис. 243. Различия в строении лицевого нерва.
а - сетевидное строение; б - магистральное строение.
1 - лицевой нерв; 2 - жевательная мышца.
2. Стременной нерв, п. stapedius, - тонкий ствол, ответвляется в лицевом канале у второго поворота, проникает в барабанную полость, где иннервирует стременную мышцу.
3. Барабанная струна, chorda tympani, является продолжением промежуточного нерва, отделяется от лицевого нерва в нижней части канала над шилососцевидным отверстием и входит через каналец барабанной струны в барабанную полость, где лежит под слизистой оболочкой между длинной ножкой наковальни и рукояткой молоточка. Через каменисто-барабанную щель барабанная струна выходит на наружное основание черепа и в подвисочной ямке сливается с язычным нервом.
В месте перекреста с нижним альвеолярным нервом барабанная струна дает соединительную ветвь с ушным узлом. Барабанная струна состоит из преганглионарных парасимпатических волокон к под- нижнечелюстному узлу и вкусовых чувствительных волокон к передним двум третям языка.
4. Соединительная ветвь с барабанным сплетением, r. communicans cum plexo tympanico, - тонкая ветвь; начинается от узла коленца или от большого каменистого нерва, проходит через крышу барабанной полости к барабанному сплетению.
По выходе из канала от лицевого нерва ответвляются следующие ветви.
1. Задний ушной нерв, п. auricularis posterior, отходит от лицевого нерва сразу по выходе из шилососцевидного отверстия, идет назад и вверх по передней поверхности сосцевидного отростка, разделяясь на две ветви: ушную, r. auricularis, иннервирующую заднюю ушную мышцу, и затылочную, r. occipitalis, иннервирующую затылочное брюшко надчерепной мышцы.
2. Двубрюшная ветвь, r. digastricus, возникает несколько ниже ушного нерва и, спускаясь вниз, иннервирует заднее брюшко двубрюшной мышцы и шилоподъязычную мышцу.
3. Соединительная ветвь с языкоглоточным нервом, r. communicans cum n. glossopharyngeus, ответвляется вблизи шилососцевидного отверстия и распространяется кпереди и вниз по шилоглоточной мышце, соединяясь с ветвями языкоглоточного нерва.
Ветви околоушного сплетения:
1. Височные ветви, rr. temporales, в количестве 2-4, идут вверх и разделяются на 3 группы: передние, иннервирующие верхнюю часть круговой мышцы глаза, и мышцу, сморщивающую бровь; средние, иннервирующие лобную мышцу; задние, иннервирующие рудиментарные мышцы ушной раковины.
2. Скуловые ветви, rr. zygomatici, в количестве 3-4, распространяются вперед и вверх к нижней и латеральной частям круговой мышцы глаза и скуловой мышце, которые и иннервирует.
3. Щечные ветви, rr. buccales, в количестве 3-5, идут горизонтально кпереди по наружной поверхности жевательной мышцы и снабжают ветвями мышцы в окружности носа и рта.
4. Краевая ветвь нижней челюсти, r. marginalis mandibularis, проходит по краю нижней челюсти и иннервирует мышцы, опускающие угол рта и нижнюю губу, подбородочную мышцу и мышцу смеха.
5. Шейная ветвь, r. colli, спускается на шею, соединяется с поперечным нервом шеи и иннервирует т. platysma.
Промежуточный нерв, п. intermedius, состоит из преганглионарных парасимпатических и чувствительных волокон. Чувствительные униполярные клетки расположены в узле коленца. Центральные отростки клеток восходят в составе корешка нерва и оканчиваются в ядре одиночного пути. Периферические отростки чувствительных клеток идут через барабанную струну и большой каменистый нерв к слизистой оболочке языка и мягкого неба.
Секреторные парасимпатические волокна берут начало в верхнем слюноотделительном ядре в продолговатом мозге. Корешок промежуточного нерва выходит из мозга между лицевым и преддверно-улитковым нервами, присоединяется к лицевому нерву и идет в лицевом канале. Волокна промежуточного нерва покидают ствол лицевого, переходя в барабанную струну и большой каменистый нерв, достигают поднижнечелюстного и подъязычного узлов и крылонебного узла.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
1. Какие черепные нервы по происхождению относятся к смешанным?
2. Какие черепные нервы развиваются из переднего мозга?
3. Перечислите нервы, иннервирующие мышцы глаз.
4. За счет каких ветвей осуществляется чувствительная иннервация глазных нервов? Укажите области этой иннервации.
5. Какие ветви отходят от глазного нерва?
6. Какими нервами иннервируются верхние зубы? Откуда отходят эти нервы?
7. Перечислите ветви нижнечелюстного нерва.
8. Какие нервы проходят в составе барабанной струны?
9. Какие ветви отходят от лицевого нерва внутри своего канала? Что они иннервируют?
10. Какие ветви отходят от лицевого нерва в области околоушного сплетения? Что они иннервируют?
VIII ПАРА - ПРЕДДВЕРНО-УЛИТКОВЫЕ НЕРВЫ
Преддверно-улитковый нерв, п. vestibulocochlearis, - чувствительный, состоит из двух функционально различных частей: преддверной и улитковой.
Преддверный нерв, n. vestibularis, проводит импульсы от статического аппарата преддверия и полукружных каналов лабиринта внутреннего уха. Улитковый нерв, n. cochlearis, обеспечивает передачу звуковых раздражений от спирального органа улитки. Каждая часть нерва имеет собственные чувствительные узлы, содержащие биполярные нервные клетки: преддверная часть - преддверный узел, ganglion vestibulare, расположенный на дне внутреннего слухового прохода; улитковая часть - улитковый узел [спиральный узел улит- ки], ganglion cochleare [spirale cochleae], который находится в улитке.
Преддверный узел удлиненный, в нем различают две части: верхнюю, pars superior, и нижнюю, pars inferior. Периферические отростки клеток верхней части образуют следующие нервы:
1) эллиптически-мешотчатый нерв, п. utricularis, к клеткам эллиптического мешочка преддверия улитки;
2) передний ампулярный нерв, п. ampullaris anterior, к клеткам чувствительных полосок передней перепончатой ампулы переднего полукружного канала;
3) латеральный ампулярный нерв, п. ampullaris lateralis, к латеральной перепончатой ампуле.
От нижней части преддверного узла периферические отростки клеток идут в составе сферически-мешотчатого нерва, п. saccularis, к слуховому пятну мешочка; заднего ампулярного нерва, п. ampullaris posterior, к задней перепончатой ампуле.
Центральные отростки клеток преддверного узла образуют преддверный (верхний) корешок, radix vestibularis (superior), который выходит через внутреннее слуховое отверстие позади лицевого и промежуточного нервов и вступает в мозг рядом с выходом лицевого нерва, достигая 4 преддверных ядер в мосту: медиального, лате- рального, верхнего и нижнего.
Из улиткового узла периферические отростки его биполярных нервных клеток идут к чувствительным эпителиальным клеткам спи-
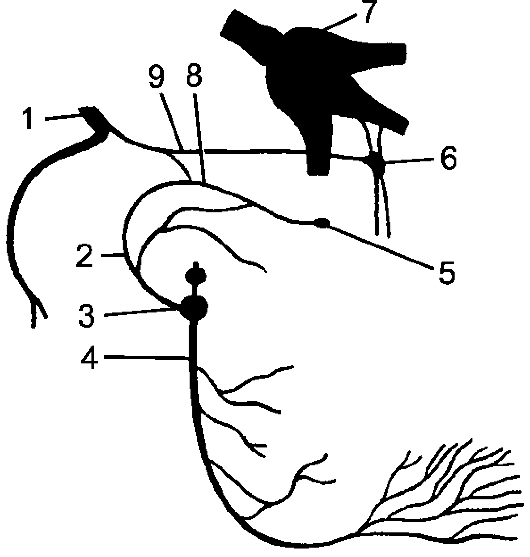
Рис. 244. Языкоглоточный нерв (схема).
1 - лицевой нерв; 2 - барабанный нерв; 3 - нижний узел языкоглоточного нерва; 4 - языкоглоточный нерв; 5 - ушной узел языкоглоточного нерва; 6 - крыло-небный узел; 7 - тройничный нерв; 8 - малый каменистый нерв; 9 - большой каменистый нерв.
рального органа улитки, образуя в совокупности улитковую часть нерва. Центральные отростки клеток улиткового узла формируют улитковый (нижний) корешок, radix cochlearis (inferior), идущий вместе с верхним корешком в мозг к дорсальному и вентральному улитковым ядрам.
IX ПАРА -
ЯЗЫКОГЛОТОЧНЫЕ НЕРВЫ
Языкоглоточный нерв, п. glossopharyngeus, - нерв третьей жаберной дуги, смешанный. Иннервирует слизистую оболочку задней трети языка, небных дужек, глотки, барабанной полости, околоушную слюнную железу и шилоглоточную мышцу (рис. 244). В составе нерва 3 вида нервных волокон: 1) чувствительные, 2) двигательные, 3) парасимпатические.
Чувствительные волокна - отростки афферентных клеток верхнего и нижнего узлов, ganglia superius et inferius. Периферические отростки следуют в составе нерва к органам, где образуют рецепторы, центральные идут в продолговатый мозг, к чувствительному ядру одиночного пути, nucleus solitarius.
Двигательные волокна начинаются от нервных клеток общего с блуждающим нервом двойного ядра, nucleus ambiguus, и проходят в составе нерва к шилоглоточной мышце.
Парасимпатические волокна берут начало в вегетативном парасимпатическом нижнем слюноотделительном ядре, nucleus salivatorius inferior, которое расположено в продолговатом мозге.
Корешок языкоглоточного нерва выходит из продолговатого мозга позади места выхода преддверно-улиткового нерва и вместе с блуждающим нервом покидает череп через яремное отверстие. В этом отверстии нерв имеет первое расширение - верхний узел, ganglion superius, а по выходе из отверстия - второе расширение - нижний узел, ganglion inferius.
Вне черепа языкоглоточный нерв лежит сначала между внутренней сонной артерией и внутренней яремной веной, а затем по-
логой дугой огибает сзади и снаружи шилоглоточную мышцу и подходит изнутри подъязычно-язычной мышцы к корню языка, разделяясь на конечные ветви.
Ветви языкоглоточного нерва:
1. Барабанный нерв, п. tympanicus, ответвляется от нижнего узла и проходит через барабанный каналец в барабанную полость, где формирует совместно с сонно-барабанными нервами барабанное сплетение, plexus tympanicus. Барабанное сплетение иннервирует слизистую оболочку барабанной полости и слуховой трубы. Барабанный нерв покидает барабанную полость через ее верхнюю стенку как малый каменистый нерв, п. petrosus minor, и идет к ушному узлу. Прегангли- онарные парасимпатические секреторные волокна, подходящие в составе малого каменистого нерва, прерываются в ушном узле, а постганглионарные секреторные волокна входят в ушно-височный нерв и достигают в его составе околоушной слюнной железы.
2. Ветвь шилоглоточной мышцы, r. musculi stylopharyngei, идет к одноименной мышце и слизистой оболочке глотки.
3. Синусная ветвь, r. sinus carotici, чувствительная, разветвляется в сонном гломусе.
4. Миндаликовые ветви, rr. tonsillares, направляются к слизистой оболочке небной миндалины и дужек.
5. Глоточные ветви, rr. pharyngei, в количестве 3-4, подходят к глотке и вместе с глоточными ветвями блуждающего нерва и симпатического ствола образуют на наружной поверхности глотки глоточное сплетение, plexus pharyngeus. От него отходят ветви к мышцам глотки и слизистой оболочке, которые в свою очередь образуют интрамуральные нервные сплетения.
6. Язычные ветви, rr. linguales, - конечные ветви языкоглоточного нерва: содержат чувствительные вкусовые волокна к слизистой оболочке задней трети языка.
X ПАРА - БЛУЖДАЮЩИЕ НЕРВЫ
Блуждающий нерв, п. vagus, смешанный, развивается из 4-5-й жаберных дуг, распространяется широко, благодаря чему и получил свое название. Иннервирует дыхательные органы, органы пищева- рительной системы (до сигмовидной ободочной кишки), щитовидную и паращитовидные железы, надпочечники, почки, участвует в иннервации сердца и сосудов.
Блуждающий нерв содержит чувствительные, двигательные и вегетативные парасимпатические и симпатические волокна, а так- же внутриствольные небольшие нервные узлы (рис. 245).
Рис. 245. Блуждающий и добавочный нервы (схема).
1 - соединительная ветвь блуждающего нерва с лицевым; 2 - языкоглоточный нерв; 3 - добавочный нерв; 4 - соединительная ветвь блуждающего нерва с подъязычным; 5 - соединительная ветвь блуждающего нерва с симпатическим стволом; 6 - язык; 7 - подъязычная кость; 8 - гортань; 9 - трахея; 10 - правый возвратный гортанный нерв;
11 - левый возвратный гортанный нерв; 12 - левый блуждающий нерв; 13 - дуга аорты; 14 - левое легкое; 15 - сердце; 16 - диафрагма; 17 - желудок; 18 - печень; 19 - правый полулунный узел чревного нервного сплетения; 20 - нервный узел на восходящей части аорты; 21 - правое легкое; 22 - пищевод; 23 - ветви правого возвратного гортанного нерва; 24 - верхний гортанный нерв; 25 - трапециевидная мышца; 26 - грудино-ключично-сосцевидная мышца; 27 - добавочный нерв; 28 - ядра блуждающего и добавочного нервов; 29 - ядро блуждающего нерва; 30 - лицевой нерв.
Чувствительные нервные волокна блуждающего нерва происходят от афферентных псевдоуниполярных нервных клеток, скопления которых формируют 2 чувствительных узла нерва: верхний, ganglion superius, расположенный в яремном отверстии, и нижний, ganglion inferius, лежащий по выходе из отверстия. Центральные отростки клеток идут в продолговатый мозг к чувствительному ядру - ядру одиночного пути, nucleus solitarius, а периферические - в составе нерва к сосудам, сердцу и внутренним органам, где заканчиваются рецепторными аппаратами.
Двигательные волокна для мышц мягкого неба, глотки и гортани берут начало от верхних клеток двигательного двойного ядра.
Парасимпатические волокна исходят из вегетативного дорсального ядра, nucleus dorsalis n. vagi, и распространяются в составе нерва к мышце сердца, мышечной ткани оболочек сосудов и внутренних органов. Импульсы, идущие по парасимпатическим волокнам, уменьшают частоту сердечных сокращений, расширяют сосуды, суживают бронхи, усиливают перистальтику трубчатых органов желудочно-кишечного тракта.
Вегетативные постганглионарные симпатические волокна поступают в блуждающий нерв по его соединительным ветвям с симпа- тическим стволом от клеток симпатических узлов и распространяются по ветвям блуждающего нерва к сердцу, сосудам и внутренним органам.
Как отмечалось, от блуждающего нерва в процессе развития отделяются языкоглоточный и добавочный нервы, поэтому блуждающий нерв сохраняет связи с этими нервами, а также с подъязычным нервом и симпатическим стволом посредством соединительных ветвей.
Блуждающий нерв выходит из продолговатого мозга позади оливы многочисленными корешками, сливающимися в общий ствол, ко- торый выходит из черепа через яремное отверстие. Далее блуждающий нерв идет книзу в составе шейного сосудисто-нервного пучка, между внутренней яремной веной и внутренней сонной артерией, а ниже уровня верхнего края щитовидного хряща - между той же веной и общей сонной артерией. Через верхнюю апертуру грудной клетки блуждающий нерв проникает между подключичными веной и артерией справа и впереди дуги аорты слева в заднее средостение. Здесь он образует впереди пищевода (левый нерв) и позади него (правый нерв) путем ветвления и связей между ветвями пищеводное нервное сплетение, plexus esophageus, которое вблизи пищеводного отверстия диафрагмы формирует 2 блуждающих ствола: передний, truncus vagalis anterior, и задний, truncus vagalis posterior, соответству-
ющие левому и правому блуждающим нервам. Оба ствола выходят из грудной полости через пищеводное отверстие, отдают ветви к желудку и заканчиваются рядом конечных ветвей в чревном сплетении. От сплетения волокна блуждающего нерва распространяются по ветвям этого сплетения. На всем протяжении блуждающего нерва от него отходят ветви (рис. 246).
Ветви головного отдела блуждающего нерва:
1. Менингеальная ветвь, r. meningeus, начинается от верхнего узла и идет через яремное отверстие, достигает твердой мозговой оболочки задней ямки черепа.
2. Ушная ветвь, r. auricularis, идет от верхнего узла по переднелатеральной поверхности луковицы яремной вены к входу в сосцевидный каналец и далее по нему к задней стенке наружного слухового прохода и части кожи ушной раковины. На своем пути образует соединительные ветви с языкоглоточным и лицевым нервами.
Ветви шейного отдела блуждающего нерва:
1. Глоточные ветви, rr. pharyngei, берут начало от нижнего узла или сразу под ним. Принимают тонкие ветви от верхнего шейного узла симпатического ствола и между наружной и внутренней сонными артериями проникают к боковой стенке глотки, на которой вместе с глоточными ветвями языкоглоточного нерва и симпати- ческого ствола образуют глоточное сплетение.
2. Верхний гортанный нерв, п. laryngeus superior, ответвляется от нижнего узла и спускается вниз и вперед по боковой стенке глотки кнутри от внутренней сонной артерии. У большого рожка подъязычной кости делится на две ветви: наружную, r. externus, и внутреннюю, r. internus. Наружная ветвь соединяется с ветвями от верхнего шейного узла симпатического ствола и идет по заднему краю щитовидного хряща к перстнещитовидной мышце и нижнему констриктору глотки, а также непостоянно отдает ветви к черпаловидным и латеральной перстнечерпаловидной мышцам. Кроме того, от нее отходят ветви к слизистой оболочке глотки и щитовидной железе. Внутренняя ветвь более толстая, чувствительная, прободает щитоподъязычную мембрану и ветвится в слизистой оболочке гортани выше голосовой щели, а также в слизистой оболочке надгор- танника и передней стенки носового отдела глотки. Образует соединительную ветвь с нижним гортанным нервом.
3. Верхние шейные сердечные ветви, rr. cardiaci cervicales superiores, - изменчивые по толщине и уровню ответвления, обычно тонкие, берут начало между верхним и возвратным гортанными нервами и идут вниз к шейно-грудному нервному сплетению.
Рис. 246. Блуждающий и языкоглоточный нервы и симпатический ствол. 1 - нижние шейные сердечные ветви блуждающего нерва; 2 - возвратный гор- танный нерв; 3 - верхние шейные сердечные ветви; 4 - глоточное сплетение; 5 - подъязычный нерв; 6 - верхний гортанный нерв; 7 - язычный нерв; 8 - глоточные ветви блуждающего нерва; 9 - языкоглоточный нерв; 10, 11 - ветви добавочного нерва; 12, 15, 17, 19 - 2-й, 3-й, 4-й и 5-й шейные спинномозговые нервы; 13 - верхний шейный узел симпатического ствола; 14, 16 - блуждающий нерв; 18 - диафрагмальный нерв; 20 - средний шейный узел симпатического ствола; 21 - плечевое сплетение; 22 - нижний шейный узел симпатического ствола; 23, 24, 26, 28 - 2-й, 3-й, 4-й и 5-й грудные узлы симпатического ствола; 25 - возвратный гортанный нерв; 27 - легочное сплетение.
4. Нижние шейные сердечные ветви, rr. cardiaci cervicales inferiores, отходят от гортанного возвратного нерва и от ствола блуждающего нерва; участвуют в формировании шейно-грудного нервного спле- тения.
Ветви грудного отдела блуждающего нерва:
1. Возвратный гортанный нерв, п. laryngus recurrens, отходит от блуждающего нерва при его входе в грудную полость. Правый возвратный гортанный нерв огибает снизу и сзади подключичную артерию, а левый - дугу аорты. Оба нерва поднимаются в борозде между пищеводом и трахеей, отдавая ветви к этим органам. Конечная ветвь - нижний гортанный нерв, п. laryngeus inferior, подходит к гортани и иннервирует все мышцы гортани, за исключением перстнещитовидной, и слизистую оболочку гортани ниже голосовых связок.
От возвратного гортанного нерва отходят ветви к трахее, пищеводу, щитовидной и паращитовидным железам.
2. Грудные сердечные ветви, rr. cardiaci thoracici, начинаются от блуждающего и левого гортанного возвратного нервов; участвуют в образовании шейно-грудного сплетения.
3. Трахейные ветви идут к грудному отделу трахеи.
4. Бронхиальные ветви направляются к бронхам.
5. Пищеводные ветви подходят к грудному отделу пищевода.
6. Перикардиальные ветви иннервируют перикард.
В пределах полостей шеи и груди ветви блуждающих, возвратных и симпатических стволов формируют шейно-грудное нервное сплетение, в составе которого выделяют органные сплетения: щитовидное, трахеальное, пищеводное, легочное, сердечное.
Ветви блуждающих стволов (брюшная часть):
1. Передние желудочные ветви начинаются от переднего ствола и образуют на передней поверхности желудка переднее желудочное сплетение.
2. Задние желудочные ветви отходят от заднего ствола и формируют заднее желудочное сплетение.
3. Чревные ветви отходят в основном от заднего ствола и принимают участие в формировании чревного сплетения.
4. Печеночные ветви входят в состав печеночного сплетения.
5. Почечные ветви формируют почечные сплетения.
XI ПАРА - ДОБАВОЧНЫЙ НЕРВ
Добавочный нерв, п. accessorius, в основном двигательный, отделившийся в процессе развития от блуждающего нерва. Начинается
двумя частями - блуждающей и спинномозговой - от соответствующих двигательных ядер в продолговатом и спинном мозге. Афферентные волокна подходят в ствол через спинномозговую часть от клеток чувствительных узлов.
Блуждающая часть выходит черепными корешками, radices craniales, из продолговатого мозга ниже выхода блуждающего нерва, спинномозговая часть формируется из спинномозговых корешков, radices spinales, выходящих из спинного мозга между задними и передними корешками. Спинномозговая часть нерва поднимается к большому затылочному отверстию, входит через него в полость черепа, где со- единяется с блуждающей частью и формирует общий ствол нерва.
В полости черепа добавочный нерв разделяется на две ветви: внутреннюю и наружную.
1. Внутренняя ветвь, r. internus, подходит к блуждающему нерву. Через эту ветвь в состав блуждающего нерва включаются двигательные нервные волокна, которые покидают его через гортанные нервы. Можно полагать, что чувствительные волокна также проходят в блуждающий и далее в гортанный нерв.
2. Наружная ветвь, r. externus, выходит из полости черепа через яремное отверстие на шею и идет сначала позади заднего брюшка двубрюшной мышцы, а далее изнутри грудино-ключично-сосцевидной мышцы. Прободая последнюю, наружная ветвь направляется вниз и заканчивается в трапециевидной мышце. Образуются соединительные связи между добавочным и шейными нервами. Иннервирует гру- дино-ключично-сосцевидную и трапециевидную мышцы.
XII ПАРА - ПОДЪЯЗЫЧНЫЙ НЕРВ
Подъязычный нерв, п. hypoglossus, преимущественно двигательный, образуется в результате слияния нескольких первичных спинномозговых сегментарных нервов, иннервирующих подъязычные мышцы. В подъязычном нерве проходят также и другие виды волокон. Чувствительные нервные волокна идут от клеток нижнего узла блуждающего нерва и, возможно, из клеток спинномозговых узлов по соединительным ветвям между подъязычным, блуждающим и шейным нервами. Симпатические волокна входят в подъязычный нерв по его соединительной ветви с верхним узлом симпатического ствола (рис. 247).
Нервные волокна, составляющие подъязычный нерв, отходят от клеток его двигательного ядра, находящегося в продолговатом мозге. Нерв выходит из него между пирамидой и оливой несколькими корешками. Сформировавшийся ствол нерва проходит через подъ-
Рис. 247. Подъязычный нерв (схема).
1 - ромбовидная ямка; 2 - канал подъязычного нерва; 3 - соединительные ветви подъязычного нерва с верхним шейным узлом симпатического ствола и нижним узлом блуждающего нерва; 4 - подъязычный нерв; 5, 6 - ветви к мышцам языка; 7 - ветви к подбородочно-подъязычной мышце; 8 - подъязычная кость; 9 - ветвь к грудино-подъязычной мышце; 10 - ветви к грудино-подъязычной мышце; 11 - ветви к грудино-щитовидной мышце; 12 - ветвь к лопаточно-подъязычной мышце; 13 - внутренняя яремная вена; 14 - нижний корешок подъязычной петли; 15 - верхний корешок подъязычной петли; 16 - внутренняя яремная вена; 17 - внутренняя сонная артерия; 18 - 1-й и 3-й шейные спинномозговые нервы; 19 - ядро подъязычного нерва.
язычный канал на шею, где располагается сначала между наружной (снаружи) и внутренней сонными артериями, а затем спускается под задним брюшком двубрюшной мышцы в виде открытой кверху дуги по боковой поверхности подъязычно-язычной мышцы, составляя верхнюю сторону треугольника Пирогова; разветвляется на конечные язычные ветви, rr. linguales, иннервирующие мышцы языка.
От середины дуги нерва вниз вдоль общей сонной артерии идет верхний корешок шейной петли, radix superior, который соединяется с ее нижним корешком, radix inferior, из шейного сплетения, в результате чего образуется шейная петля, ansa cervicalis. От шейной петли отходит несколько ветвей к мышцам шеи, расположенным ниже подъязычной кости.
Положение подъязычного нерва на шее может быть различным. У людей с длинной шеей дуга, образуемая нервом, лежит относи- тельно низко, а у людей с короткой шеей - высоко. Это важно учитывать при операциях на нерве (табл. 15).
Таблица 15. Области иннервации, волоконный состав и названия ядер черепных нервов
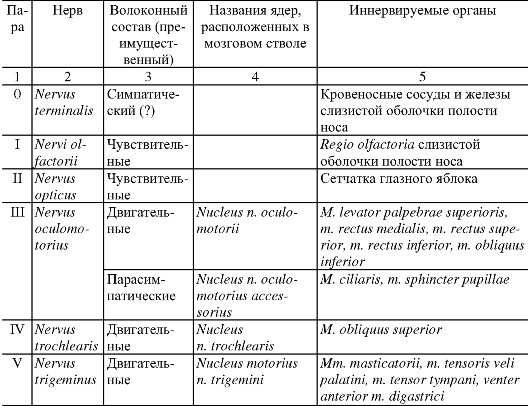
Продолжение табл. 15
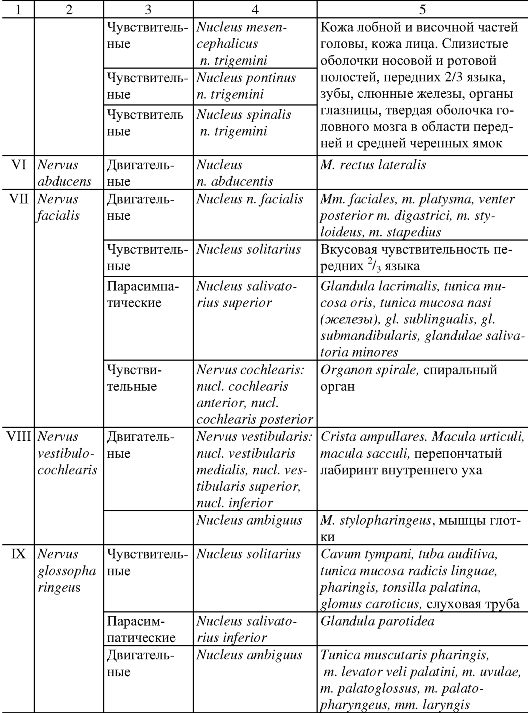
Окончание табл. 15
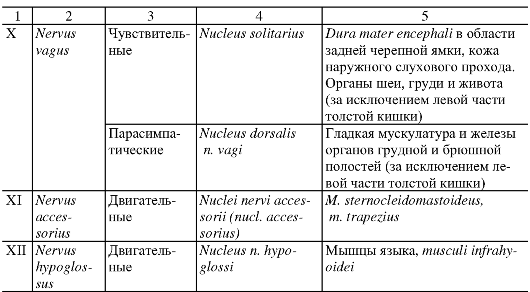
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
1. Какие нервы отходят от преддверного узла?
2. Перечислите ветви языкоглоточного нерва.
3. Какие ветви отходят от головного и шейного отделов блуждающего нерва? Что они иннервируют?
4. Перечислите ветви грудного и брюшного отдела блуждающего нерва. Что они иннервируют?
5. Что иннервируют добавочный и подъязычный нервы?
АВТОНОМНАЯ (ВЕГЕТАТИВНАЯ) НЕРВНАЯ СИСТЕМА
ОБЩИЕ ДАННЫЕ ОБ АВТОНОМНОЙ (ВЕГЕТАТИВНОЙ) НЕРВНОЙ СИСТЕМЕ
Автономная нервная система является частью нервной системы организма, которая иннервирует кровеносные и лимфатические сосуды и внутренние органы, осуществляющие так называемые растительные функции (дыхание, пищеварение, обмен веществ, выделение, размножение и др.). Кроме того, она выполняет адап- тационно-трофическую функцию - регулирует обмен веществ применительно к условиям внешней среды.
Естественно, что автономность этой нервной системы относительная и не предусматривает ее независимость от высших центров нервной системы. Термин «автономная нервная система» отражает лишь то, что она управляет непроизвольными функциями организма. Между анимальной и автономной частями нервной системы существуют тесная анатомическая и функциональная связь и взаимопроникновение. В рефлекторную дугу включаются анимальные эфферентные и автономные нейроны. В составе спинномозговых и черепных нервов проходят вегетативные нервные проводники. Деятельность обеих частей нервной системы объединяется корой полушарий большого мозга. Высшим автономным центром, осуществляющим регуляцию и интеграцию функций внутренних органов, эндокринных желез и сосудистой системы, является гипоталамус.
В автономной нервной системе выделяют центральную часть, расположенную в головном и спинном мозге, и периферическую, имеющую очень широкую область распространения, проникающую во все органы и ткани.
Анатомически вегетативная нервная система имеет следующие отличия от соматической (анимальной).
1. Территория эфферентной иннервации. Анимальная часть нервной системы иннервирует только скелетную мускулатуру (ограниченная территория иннервации), а автономная обеспечивает ее тонус (обширная территория иннервации).
2. Сохранение сегментарности. В анимальной нервной системе сохраняется отчетливая сегментарность, в автономной она полностью отсутствует.
3. Ход аксонов. В соматической нервной системе аксоны идут беспрерывно до рабочего органа, а в вегетативной эфферентный путь обязательно складывается из двух нейронов, а переключение возбуждения с проводника одного нейрона на другой происходит в вегетативных ганглиях. Различают предузловые - преганглионарные и послеузловые - постганглионарные волокна.
4. Строение нервных волокон. В соматической нервной системе волокна миелиновые, в вегетативной предузловые волокна тонкие миелиновые, послеузловые - безмиелиновые.
5. Строение рефлекторных дуг. Рефлекторная дуга в анимальный нервной системе состоит из чувствительного, вставочного и эффекторного нейронов. В автономной нервной системе рефлекторная дуга начинается в стенке внутреннего органа рецептором периферического отростка чувствительной псевдоуниполярной нервной клетки спинального ганглия. Центральный отросток клетки идет в
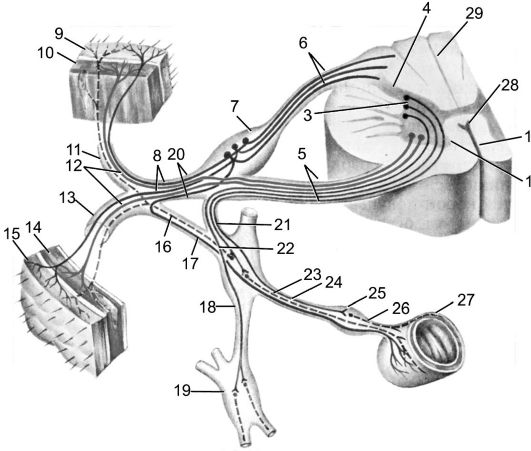
Рис. 248. Рефлекторная дуга автономной и анимальной нервных систем. 1, 28 - передняя срединная щель; 2 - передний рог; 3 - боковой рог; 4 - задний рог; 5 - брюшной корешок; 6 - спинной корешок; 7 - спинномозговой узел; 8 - афферентные волокна; 9, 15 - кожа; 10, 14 - мышца; 11 - спинная ветвь спинномозгового нерва; 12 - двигательные волокна; 13 - брюшная ветвь спинномозгового нерва; 16 - серая соединительная ветвь; 17 - висцеросенсорные волокна; 18 - межузловая ветвь; 19 - узел симпатического ствола; 20 - спинномозговой нерв; 21 - белая соединительная ветвь (преганглионарные волокна к предпозвоночным ганглиям); 22 - преганглионарное волокно к узлу симпатического ствола; 23 - постганглионарное волокно, идущее к органу; 24 - преганглионарное волокно к предпозвоночному ганглию; 25 - предпозвоночный ганглий; 26 - постганглионарное волокно, идущее от предпозвоночного узла; 27 - кишка; 29 - задняя срединная щель.
боковой рог серого вещества спинного мозга, где переключается на вставочный (ассоциативный) нейрон, длинный отросток которого в виде преганглионарного волокна подходит к эфферентным клеткам, расположенным в вегетативных ганглиях на периферии. Здесь в ганглиях происходят контакты ассоциативного и эфферентного вегетативных нейроцитов; послеузловые волокна следуют к рабочему органу. В вегетативных ганглиях образуются контакты многих нервных клеток, поэтому эти ганглии могут рассматриваться как периферические нервные центры (рис. 248, см. также цв. вкл.).
Автономная нервная система состоит из двух частей - симпатической и парасимпатической. При передаче возбуждения в нервных окончаниях этих частей автономной нервной системы образуются различные медиаторы, поэтому они выполняют разные функции. Например, при раздражении симпатической части наблюдаются повышение частоты сердечных сокращений, сужение сосудов, ослабление перистальтики кишечника, а при раздражении парасимпатической - снижение частоты сердечных сокращений, расширение сосудов, усиление перистальтики кишечника. Однако различие функций симпатической и парасимпатической частей не антагонистично, так как объекты действия импульсов с этих двух систем различные. В связи с этим можно говорить о взаимодействии двух указанных частей - о синергизме. Следует иметь в виду, что симпатическая часть автономной нервной системы имеет более широ- кую, чем парасимпатическая, территорию иннервации, так как некоторые органы (потовые и сальные железы, мышцы волосяных луковиц, скелетные мышцы) имеют только симпатическую иннервацию.
Обе части автономной нервной системы проникают друг в друга, и большинство проводников и сплетений содержит как симпатические, так и парасимпатические волокна.
В автономной нервной системе, так же как и в анимальной, различают 2 отдела: центральный, включающий ее центры, расположенные в головном и спинном мозге, и периферический, состоя- щий из ганглиев, нервов, нервных сплетений и нервных окончаний.
Периферические отделы автономной нервной системы включают: 1) ганглии; 2) внеорганные нервные сплетения; 3) внутристеночные органные (интрамуральные) сплетения; 4) нервные окончания.
1. Скопления нервных клеток (ганглии) в симпатической и парасимпатической частях автономной нервной системы расположены различно. В симпатической части они образуют по сторонам позвоночника две паравертебральные цепочки узлов, соединяющихся между собой и образующих два симпатических ствола. В парасимпатической части узлы нервных клеток связаны с III, VII, IX и Х парами черепных нервов. Кроме того, автономные узлы есть и в других участках: они входят в состав автономных нервных сплетений, а парасимпатические клетки расположены еще и в стенках органов.
2. Внеорганные автономные нервные сплетения, plexus autonomici, образуются автономными ганглиями и нервами - ветвями симпа-
тического ствола и блуждающих нервов (или тазовых внутренностных нервов), а также чувствительными волокнами анимальной нервной системы около крупных артерий; внеорганные сплетения являются смешанными и общими для обеих частей автономной нервной системы.
3. Интрамуральные нервные сплетения, plexus intramurales, залегают в различных слоях стенок органов. В полых органах, например, сплетения находятся в подсерозном, подслизистом и мышечном слоях (plexus subserosus, submucosus et myentericus). Эти сплетения образованы мелкими ганглиями из парасимпатических клеток и нервами, состоящими из симпатических, парасимпатических и чувствительных анимальных волокон, подходящих из внеорганных нервных сплетений. Интрамуральные сплетения, так же как и вне- органные, являются смешанными, общими для обеих частей вегетативной нервной системы.
4. Окончания нервных волокон автономной нервной системы представлены главным образом двигательными окончаниями - эффекторами в гладкой мышечной ткани. Кроме того, имеются и чувствительные окончания - рецепторы, образующие местные рефлекторные дуги.
РАЗВИТИЕ АВТОНОМНОЙ НЕРВНОЙ СИСТЕМЫ
Автономная нервная система, выполняющая одну из самых древних функций - адаптационно-трофическую, обеспечивает адаптацию (adapto - примеривать, прилаживать) обмена веществ организма к условиям внешней среды, поэтому и сама автономная нервная система древняя. На ранних стадиях развития жизни, например у беспозвоночных, имелась общая ганглиозно-сетевидная нервная система, выполняющая все функции нервной системы - как иннервацию мускулатуры, так и регуляцию внутренней среды организма. В ходе эволюции и усложнения строения организмов единая нервная система раз- делилась на 2 отдела - анимальный и автономный. В ходе эволюции очень быстро происходило развитие аппарата движения и органов чувств, помогающих особи лучше приспосабливаться к изменяющимся условиям среды, поэтому анимальная часть нервной системы подверглась быстрому прогрессивному развитию. Наоборот, устройство автономной нервной системы более соответствовало конструкции и функции внутренних органов. Данная часть нервной системы разви- валась медленнее. У ланцетника (бесчерепные) автономная нервная система состоит из нервных сплетений в стенке внутренних органов,
связанных с дорсальными корешками спинномозговых нервов. В составе органных нервных сплетений имеются нервные клетки, но они рассеяны диффузно и скоплений (ганглиев) не образуют. Прегангли- онарные волокна начинаются от нервных клеток мозговой трубки, лежащих по сторонам от центрального канала, и заканчиваются синапсами на нервных клетках сплетений, откуда отходят постганглионарные волокна, образующие эффекторы.
В процессе эмбриогенеза автономные нервные центры в головном и спинном мозге образуются в результате размножения и диф- ференцировки нейробластов мозговых пузырей и нервной трубки. От нейробластов, формирующих автономные центры, вырастают нейриты, которые через передние корешки следуют на периферию.
Из общего зачатка нервной системы (из продолговатого мозга и ганглиозной пластинки) по ходу задних корешков мигрируют нервные клетки (нейробласты), которые, концентрируясь по бокам от позвоночника, образуют узлы симпатического ствола (паравертебральные узлы). Нервные клетки из продолговатого мозга распространяются по ходу блуждающих нервов на периферию, где принимают участие в формировании интрамуральных нервных сплетений. Несколько позже происходит миграция клеток из паравер- тебральных узлов далее на периферию, вследствие чего образуются превертебральные ганглии в сплетениях вокруг крупных сосудов.
Из скоплений нервных клеток в области среднего и продолговатого мозга наблюдается перераспределение нервных клеток по ходу некоторых черепных нервов.
СИМПАТИЧЕСКАЯ ЧАСТЬ АВТОНОМНОЙ
НЕРВНОЙ СИСТЕМЫ
Центральный отдел симпатической части автономной нервной системы представлен симпатическими ядрами, расположенными в боковых рогах спинного мозга на уровне сегментов ThI-LIII. Нейриты клеток выходят из спинного мозга в качестве преганглионарных волокон через передние корешки, отделяются от них в виде белых соединительных ветвей, rr. communicantes albi, и идут к паравертебральным узлам симпатического ствола.
Периферический отдел симпатической части автономной нервной системы представлен двумя цепочками узлов симпатического ствола, ganglia trunci sympathici. Эти цепочки расположены по бокам позвоночника и поэтому называются паравертебральными цепочками симпатических узлов. Узлы соединяются посредством межузло-
вых ветвей, rr. interganglionares, в 2 симпатических ствола, trunci sympathici. Межузловые ветви могут соединять узлы не только одной стороны тела, но и узлы противоположных сторон (в поясничном и крестцовом отделах). От симпатических стволов отходят множественные ветви к автономным нервным сплетениям, а также серые соединительные ветви, rr. communicantes grisei, к ближайшим спинномозговым нервам; по этим ветвям к нервам подходят постганглионарные симпатические волокна к скелетным мышцам, железам и т.д. Кроме того, симпатические клетки концентрируются в узлах превертебральных автономных сплетений. Преганглионарные нервные волокна идут от ядер боковых столбов спинного мозга к нервным клеткам паравертебральных или превертебральных ганглиев и на них переключаются синаптически на постганглионарные волокна, следующие к рабочим органам. Если преганглионарные волокна переключаются в превертебральных ганглиях, то они проходят через узлы симпатического ствола, не прерываясь в них.
СИМПАТИЧЕСКИЙ СТВОЛ
Симпатический ствол, truncus sympathicus, парный, состоит из цепочки узлов, ganglia trunci sympathici, связанных межузловыми ветвями. Оба ствола лежат на боковых поверхностях позвоночника и на копчике соединяются в общий копчиковый узел.
Симпатический ствол делится на 4 отдела: шейный, грудной, поясничный и крестцовый (рис. 249).
Шейный отдел симпатического ствола залегает впереди поперечных отростков шейных позвонков на поверхности глубоких мышц шеи. В каждом шейном отделе 3 шейных узла: верхний и средний, ganglia cervicales superior et medius, и шейно-грудной (звездчатый), ganglion cervicothoracicum (stellatum). Средний шейный узел самый маленький. Звездчатый узел нередко состоит из нескольких узлов. Общее число узлов в шейном отделе может колебаться от 2 до 6. От шейных узлов отходят нервы к голове, шее и груди.
1. Серые соединительные ветви, rr. communicantes grisei, к шейному и плечевому сплетениям.
2. Внутренний сонный нерв, п. caroticus internus, отходит обычно от верхнего и среднего шейных узлов к внутренней сонной артерии и формирует вокруг нее внутреннее сонное сплетение, plexus caroticus internus, которое распространяется и на ее ветви. От сплетения ответвляется глубокий каменистый нерв, п. petrosus profundus, к крылонебному узлу.
3. Яремный нерв, п. jugularis, начинается от верхнего шейного узла, в пределах яремного отверстия разделяется на две ветви: одна
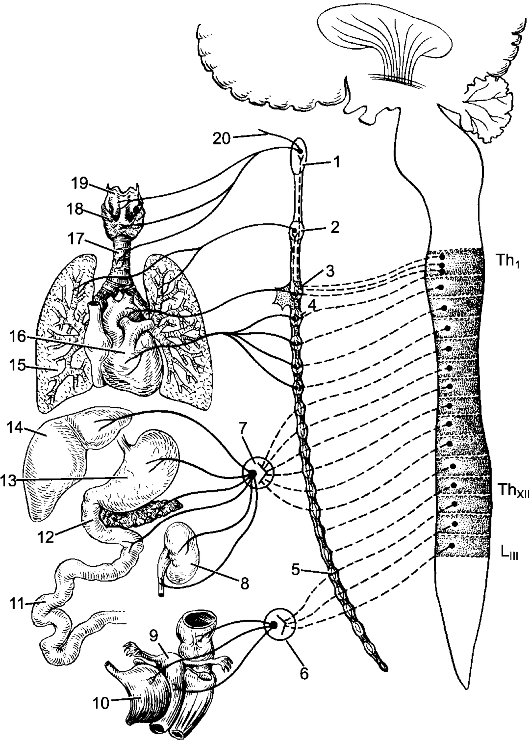
Рис. 249. Симпатический ствол (схема). Справа - спинной мозг с симпатическим центром в боковых рогах; в середине - симпатический ствол; слева - превертебральные нервные сплетения (6, 7) и иннервируемые симпатическим нервом органы. Пунктиром обозначены преганглионарные нервные волокна, сплошной линией - постганглионарные.
1 - верхний шейный узел симпатического ствола; 2 - средний шейный узел; 3 - звездчатый узел; 4 - 2-й грудной узел симпатического ствола; 5 - верхний крестцовый узел симпатического ствола; 6 - тазовое нервное сплетение; 7 - чревное нервное сплетение; 8 - почка; 9 - матка; 10 - мочевой пузырь; 11 - тощая кишка; 12 - двенадцатиперстная кишка; 13 - желудок; 14 - печень; 15 - легкое; 16 - сердце; 17 - трахея; 18 - щитовидная железа; 19 - гортань; 20 - внутренний сонный нерв.
идет к верхнему узлу блуждающего нерва, другая - к нижнему узлу языкоглоточного нерва.
4. Позвоночный нерв, п. vertebralis, отходит от шейно-грудного узла к позвоночной артерии, вокруг которой формирует позвоночное сплетение, plexus vertebralis.
5. Сердечные шейные верхние, средние и нижние нервы, пп. cardiaci cervicales superior, medius et inferior, берут начало от соответствующих шейных узлов и входят в состав шейно-грудного нервного сплетения.
6. Наружные сонные нервы, пп. carotici externi, отходят от верхнего и среднего шейных узлов к наружной сонной артерии, где участвуют в образовании наружного сонного сплетения, plexus caroticus externus, которое распространяется и на ветви артерии.
7. Гортанно-глоточные ветви, rr. laryngopharyngei, идут от верхнего шейного узла к глоточному нервному сплетению и в качестве соединительной ветви к верхнему гортанному нерву.
8. Подключичные ветви, rr. subclavii, отходят от подключичной петли, ansa subclavia, которая образована разделением межузловой ветви между средним шейным и шейно-грудными узлами.
Грудной отдел симпатического ствола включает 9-12 грудных узлов, от которых начинаются следующие ветви:
1. Белые соединительные ветви, rr. communicantes albi, соединяют грудные спинномозговые нервы с грудными узлами симпатического ствола. В составе этих ветвей и узлов идут преганглионарные волокна из ядер боковых столбов спинного мозга.
2. Серые соединительные ветви, rr. communicantes grisei, отходят от грудных узлов к грудным спинномозговым нервам.
3. Большой внутренностный нерв, п. splanchnicus major, формируется в результате слияния отдельных небольших стволов, начинающихся от 5-9-го грудных узлов симпатического ствола. Общий ствол проходит между ножками диафрагмы в брюшную полость и здесь входит в состав чревного сплетения.
4. Малый внутренностный нерв, п. splanchnicus minor, образуется, как и большой, из отдельных корешков, идущих от 10-11-го грудных узлов. Проходит вместе с большим внутренностным нервом через диафрагму и включается в состав почечного или чревного нервного сплетения.
5. Грудные сердечные нервы, пп. cardiaci thoracici, ответвляются от верхних 4-5 грудных узлов и входят в состав грудной части шейно-грудного нервного сплетения.
Поясничный отдел симпатического ствола состоит из 3-4 поясничных узлов, расположенных у медиального края большой поясничной мышцы. От узлов отходят следующие нервы.
1. Белые соединительные ветви, rr. communicantes albi, соединяющие I-II (или I-III) поясничные спинномозговые нервы с поясничными узлами.
2. Серые соединительные ветви, rr. communicantes grisei, соединяющие поясничные узлы с поясничным нервным сплетением.
3. Поясничные внутренностные нервы, пп. splanchnici lumbales, идут к чревному, почечному, надпочечниковому, аортальному, нижнему брыжеечному сплетениям.
Крестцовый отдел симпатического ствола включает в себя 3- 4 крестцовых узла, ganglia sacralia, лежащих медиальнее передних крестцовых отверстий.
Оба симпатических ствола соединяются в непарном узле, ganglion impar, на передней поверхности копчика. От узлов отходят следующие ветви.
1. Серые соединительные ветви к крестцовому нервному сплетению.
2. Крестцовые внутренностные нервы, пп. splanchnici sacrales, к тазовому нервному сплетению.
ПАРАСИМПАТИЧЕСКАЯ ЧАСТЬ АВТОНОМНОЙ НЕРВНОЙ СИСТЕМЫ
Центральный отдел парасимпатической части автономной нервной системы представлен в виде ядер в головном (мезэнцефалическое и бульбарные ядра) и спинном мозге.
Мезэнцефалическое парасимпатическое ядро - добавочное ядро глазодвигательного нерва, nucleus oculomotorius accessorius, находится на дне водопровода среднего мозга, медиальнее ядра глазодвигательного нерва. Преганглионарные парасимпатические волокна идут из этого ядра в составе глазодвигательного нерва к ресничному узлу.
В продолговатом мозге и мосту лежат следующие парасимпатические ядра:
1) верхнее слюноотделительное ядро, nucleus salivatorius superior, связанное с лицевым нервом, - в мосту;
2) нижнее слюноотделительное ядро, nucleus salivatorius inferior, связанное с языкоглоточным нервом, - в продолговатом мозге;
3) дорсальное ядро блуждающего нерва, nucleus dorsalis nervi vagi, - в продолговатом мозге.
Преганглионарные парасимпатические волокна проходят от клеток слюноотделительных ядер в составе лицевого и языкоглоточного нервов в поднижнечелюстной, подъязычный, крылонебный и ушной узлы (табл. 16).
Крестцовые парасимпатические ядра, nuclei parasympathici sacrales, лежат в боковых рогах спинного мозга на уровне II-IV крестцовых
Таблица 16. Области вегетативной иннервации, названия вегетативных ядер и узлов, волоконный состав нервов
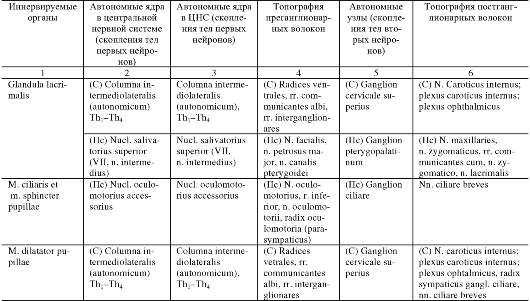
Окончание табл. 16
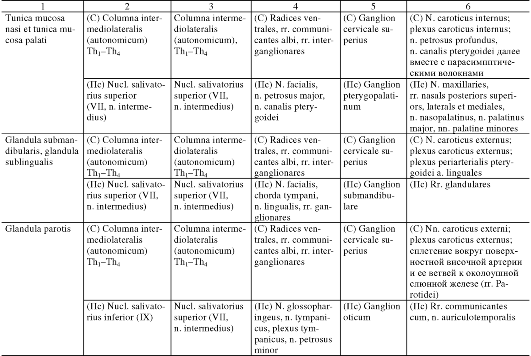
сегментов. Преганглионарные волокна выходят из мозга через передние корешки в тазовое сплетение.
Периферический отдел парасимпатической нервной системы не имеет самостоятельных образований. Парасимпатические нервные волокна от черепных ядер проходят в составе черепных нервов (III, VII, IX, X пары), а от спинномозговых промежуточных ядер - в составе тазовых внутренностных нервов, пп. splanchnici pelvini.
1. Преганглионарные нервные волокна, идущие в составе глазодвигательного нерва, следуют до ресничного узла и заканчиваются на его клетках синапсами. От узла отходят короткие ресничные нервы, пп. ciliares breves, в которых наряду с чувствительными волокнами имеются парасимпатические. Они иннервируют сфинктер зрачка и ресничную мышцу.
2. Преганглионарные волокна из клеток верхнего слюноотделительного ядра распространяются в составе промежуточного нерва, от него через большой каменистый нерв идут к крылонебному узлу, а через барабанную струну - к поднижнечелюстному и подъязычному узлам, где заканчиваются синапсами. От указанных узлов по их ветвям следуют постганглионарные волокна к рабочим органам (поднижнечелюстные и подъязычные слюнные железы, железы неба, носа и языка).
3. Преганглионарные волокна из клеток нижнего слюноотделительного ядра идут в составе языкоглоточного нерва и далее по малому каменистому нерву к ушному узлу, на клетках которого заканчиваются синапсами. Постганглионарные волокна от клеток ушного узла выходят в составе ушно-височного нерва и иннервируют околоушную железу.
4. Преганглионарные парасимпатические волокна, начинающиеся от клеток дорсального узла блуждающего нерва, проходят в составе блуждающего нерва, который является главным проводником парасимпатических волокон. Переключение на постганглионарные волокна происходит в основном в мелких ганглиях интрамуральных нервных сплетений большинства внутренних органов, поэтому постганглионарные парасимпатические волокна представляются сравнительно с преганглионарными очень короткими.
5. Преганглионарные парасимпатические волокна крестцовых парасимпатических ядер выходят из спинного мозга в составе передних корешков крестцовых спинномозговых нервов. Далее распространяются через тазовые внутренностные нервы, пп. splanchnici pelvini, в тазовое нервное сплетение. Переключение на постганглионарные парасимпатические волокна совершается или в узлах та-
зового сплетения, или в узлах интрамуральных нервных сплетений тазовых органов.
ВНЕОРГАННЫЕ И ИНТРАМУРАЛЬНЫЕ АВТОНОМНЫЕ СПЛЕТЕНИЯ
Внеорганные вегетативные нервные сплетения формируются вокруг крупных артериальных стволов шеи, груди, живота, таза из волокон обеих частей вегетативной нервной системы. Сплетения располагаются спереди от позвоночника, поэтому называются превертебральными нервными сплетениями. От внеорганных нервных сплетений нервы распространяются по сосудам, идущим в стенку органа, где входят в состав интрамуральных сплетений.
Интрамуральные нервные сплетения расположены в стенках внутренних органов. Они образуются конечными ветвями симпатической (постганглионарные волокна) и парасимпатической (преганглионарные волокна, нервные клетки и постганглионарные волокна) систем, а также чувствительными волокнами анимальной нервной системы. Указанные волокна проходят в тонких нервных стволах, которые, соединяясь, образуют в стенке органов сети. На этих стволах расположены мелкие скопления парасимпатических клеток.
ШЕЙНЫЕ И ГРУДНЫЕ АВТОНОМНЫЕ СПЛЕТЕНИЯ
Из ветвей шейных и в меньшей степени грудных узлов симпатического ствола и ветвей блуждающих нервов формируется на на- ружной поверхности органов шеи и в средостении шейногрудное нервное сплетение, plexus cervicothoracicus (рис. 250), глоточное, plexus pharyngeus, общее сонное, plexus caroticus communis, пищеводное, plexus esophageus, гортанное, plexus laryngeus, трахеальное, plexus trachealis, щитовидное, plexus thyroideus, сердечное, plexus cardiacus, легочное, plexus pulmonalis, грудное аортальное, plexus aorticus thoracicus.
БРЮШНЫЕ АВТОНОМНЫЕ СПЛЕТЕНИЯ
В полости живота главным автономным сплетением является чревное, plexus celiacus. Оно образуется вокруг чревного артериального ствола из скоплений в основном симпатических нервных клеток - чревных узлов, ganglia celiaca, и ветвей блуждающих, больших и малых внутренностных нервов, ветвей нижних грудных и верхних поясничных узлов симпатических стволов, ветви из право-
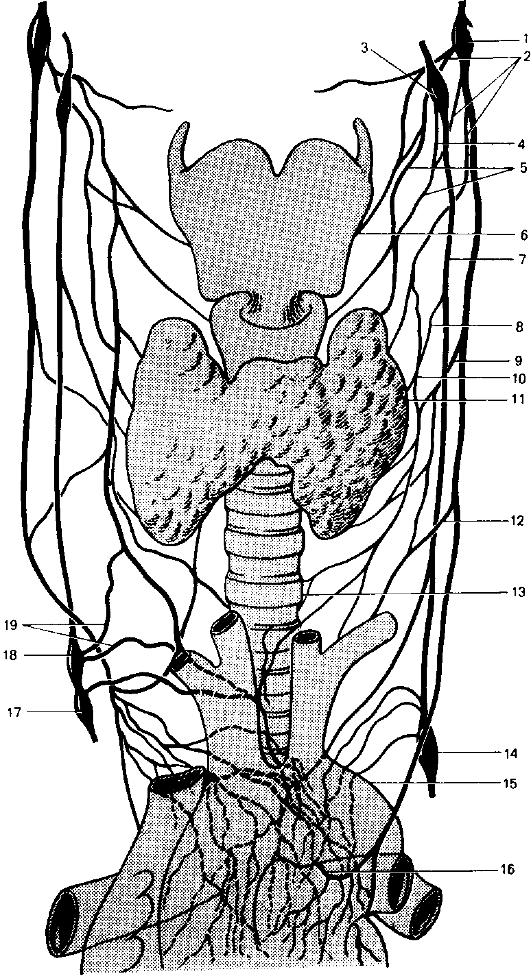
Рис. 250. Шейно-грудное нервное сплетение (схема).
1 - нижний узел блуждающего нерва; 2 - верхние шейные сердечные ветви блуждающего нерва; 3 - верхний шейный узел симпатического ствола; 4 - верхний шейный сердечный нерв; 5 - ветви к щитовидной железе; 6 - щитовидный хрящ; 7 - межузловая ветвь; 8 - средний шейный сердечный нерв; 9 - средняя шейная сердечная ветвь; 10 - смешанные ветви, образованные путем соединения верхней сердечной ветви и среднего сердечного нерва; 11 - щитовидная железа; 12 - нижняя шейная сердечная ветвь; 13 - трахея; 14 - звездчатый узел; 15 -дуга аорты; 16 - левый возвратный гортанный нерв; 17 - верхний грудной узел симпатического ствола; 18 - нижний шейный узел симпатического ствола; 19 - нижние шейные сердечные нервы.
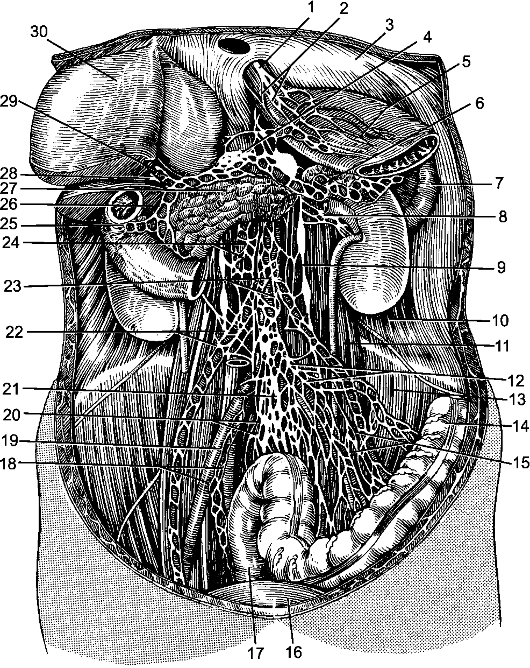
Рис. 251. Чревное сплетение и его производные.
1 - блуждающий нерв; 2 - сплетение блуждающего нерва на пищеводе; 3 - диафрагма; 4 - чревное сплетение; 5 - переднее желудочное сплетение; 6 - селезеночное сплетение; 7 - селезенка; 8 - почечное сплетение; 9 - симпатический ствол; 10, 11, 13 - мышцы задней брюшной стенки; 12 - нижнее брыжеечное сплетение; 14 - сигмовидная ободочная кишка; 15 - ветви нижнего брыжеечного сплетения; 16 - мочевой пузырь; 17 - прямая кишка; 18 - подвздошное сплетение; 19 - наружная подвздошная артерия; 20, 21 - верхнее подчревное сплетение; 22 - яичковое сплетение; 23 - межбрыжеечное сплетение; 24 - верхнее брыжеечное сплетение; 25 - двенадцатиперстная кишка; 26, 29 - панкреатическое сплетение; 27 - поджелудочная железа; 28 - печеночная артерия; 30 - печень.
го диафрагмального нерва. Число ганглиев может быть различным (два крупных ганглия или много мелких) (рис. 251).
Нервные ветви от чревного сплетения распространяются в виде периартериальных сплетений по ветвям чревного ствола и по аор-
те. Во вторичных сплетениях имеются небольшие узлы нервных клеток, в основном симпатических. Производные чревного сплетения:
1. Печеночное сплетение, plexus hepaticus, вокруг печеночной артерии.
2. Селезеночное сплетение, plexus splenicus, вокруг селезеночных сосудов.
3. Желудочные сплетения, plexus gastrici, вокруг артерий желудка.
4. Верхнее брыжеечное сплетение, plexus mesentericus superior, - сильно развитое сплетение, расположенное у верхних брыжеечных сосудов. В него входят верхние брыжеечные узлы, ganglia mesenterica superiora. Сплетение распространяется на все ветви верхней брыжеечной артерии и достигает стенки кишки.
5. Панкреатическое сплетение, plexus pancreaticus, образуется на поверхности поджелудочной железы из ветвей селезеночного и верхнего брыжеечных сплетений.
6. Почечное сплетение, plexus renalis, расположено на почечной артерии; имеет почечные узлы, ganglia renalia.
7. Надпочечниковое сплетение, plexus suprarenalis, находится на поверхности надпочечника. Является производным чревного и почечного сплетений.
8. Мочеточниковое сплетение, plexus uretericus, расположено вокруг мочеточника.
Вторым развитым автономным сплетением является межбрыжеечное сплетение, plexus intermesentericus. Оно формируется на поверхности брюшной аорты между двумя брыжеечными артериями. Корнями сплетения служат ветви чревного и почечного сплетений и поясничные внутренностные нервы. Сплетение содержит аортальные, ganglia aortica, аортально-почечные, ganglia aortorenalia, и нижние брыжеечные узлы, ganglia mesenterica inferiora, состоящие в основном из симпатических клеток.
Производные межбрыжеечного сплетения:
1. Нижнее брыжеечное сплетение, plexus mesentericus inferior, окружает одноименную артерию. Оно продолжается по ходу верхней прямокишечной артерии в одноименное сплетение - plexus rectalis superior.
2. Яичковое (яичниковое) сплетение, plexus testicularis (ovaricus), расположено по ходу соответствующих артерий. Содержит также ветви чревного сплетения.
3. Общее подвздошное сплетение, plexus iliacus communis, располагается по ходу одноименных сосудов.
4. Наружное подвздошное сплетение, plexus iliacus externus.
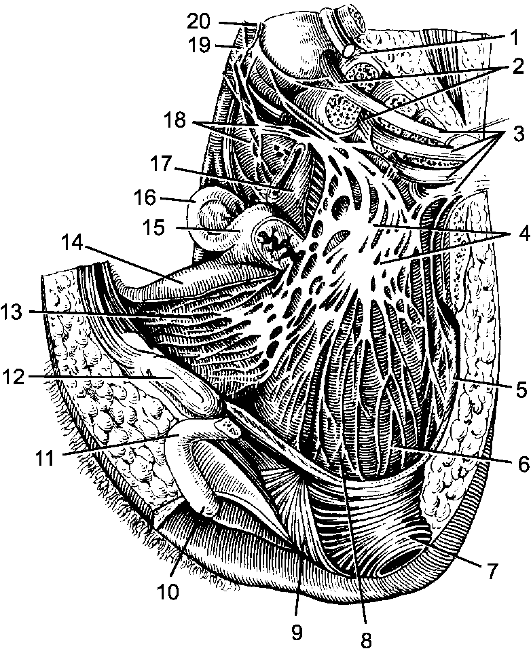
Рис. 252. Тазовое нервное сплетение.
1 - крестец; 2 - передние ветви крестцовых нервов с отходящими от них соединительными ветвями к симпатическому стволу; 3 - крестцовое сплетение; 4 - прямокишечное и маточно-влагалищное сплетения; 5, 8 - мышца, поднимающая задний проход; 6, 17 - прямая кишка; 7 - наружный сфинктер заднего прохода; 9 - глубокая поперечная мышца промежности; 10, 11 - клитор; 12 - симфиз; 13 - мочепузырное сплетение; 14 - мочевой пузырь; 15 - матка; 16 - маточная труба; 18 - верхнее прямокишечное сплетение; 19 - верхнее подчревное сплетение; 20 - аорта.
ТАЗОВЫЕ АВТОНОМНЫЕ СПЛЕТЕНИЯ
В тазу из ветвей межбрыжеечного сплетения, ветвей от крестцовых узлов симпатического ствола и тазовых внутренностных не- рвов по ходу внутренней подвздошной артерии и ее ветвей формируется сильно развитое тазовое сплетение, plexus pelvinus. В нем, кроме нервных стволов, много тазовых узлов, ganglia pelvina, содержащих симпатические и парасимпатические клетки (рис. 252).
Производные тазового сплетения:
1. Средние и нижние прямокишечные сплетения, plexus rectales medii et inferiores, распространяются по одноименным сосудам.
2. Мочепузырные сплетения, plexus vesicates, расположены на артериях мочевого пузыря.
3. Предстательное сплетение, plexus prostaticus, располагается на поверхностях предстательной железы.
4. Сплетение семявыносящего протока, plexus deferentialis.
5. Маточно-влагалищное сплетение, plexus uterovaginalis.
ИНТРАМУРАЛЬНЫЕ АВТОНОМНЫЕ СПЛЕТЕНИЯ
Подходящие вместе с артериями ветви превертебральных вегетативных сплетений, содержащие различные проводники (симпатические, парасимпатические и чувствительные), формируют в стенках органов внутристеночные, интрамуральные, сплетения. Они имеют вид сетей, образованных тонкими нервами и мелкими ганглиями, состоящими в основном из парасимпатических клеток. Сплетения в зависимости от сложности строения органа устроены различно. В паренхиматозных органах они располагаются по ходу внутриорганных сосудов в соединительной ткани между долями и дольками органа, распространяются в ткани вместе с капиллярами, а также под капсулами органов. В полых органах интрамуральные сплетения лежат между оболочками. В полых органах кишечного тракта различают сплетения: 1) подсерозное, или подфасциальное; 2) мышечно-кишечное; 3) подслизистое.
От интрамуральных сплетений отходят группы волокон и единичные волокна, образующие нервные окончания, - эффекторы и рецепторы.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ
1. Каковы отличия автономной нервной системы от анимальной?
2. Что включает в себя периферический отдел автономной нервной системы?
3. Где располагается центральный отдел симпатической нервной системы?
4. Какие ветви отходят от шейного отдела симпатического ствола? Что они иннервируют?
5. Какие ветви отходят от грудного и поясничного отделов симпатического ствола? Что они иннервируют?
6. За счет каких нервов и через какие узлы осуществляется парасимпатическая иннервация слезной железы и языка?
7. Перечислите периартериальные сплетения брюшной полости.